
Abstract
Association studies on the MTHFR polymorphisms (C677T and A1298C) in colorectal cancer (CRC) and colorectal adenoma have shown conflicting results. We performed a meta-analysis to better assess the purported associations. Overall, the 677T allele (10,131 patients and 15,362 controls) showed a small but significant protective effect against CRC compared to the 677C allele [P=0.0003, odds ratio (OR)=0.93; 95% confidence interval (CI) 0.89–0.98, P=0.22 (for heterogeneity)] for a worldwide population. Meta-analyses of other genetic contrasts suggested that the 677T allele is more likely to affect CRC in a recessive genetic model worldwide (P<0.0001, OR=0.86; 95% CI 0.76–0.96, P=0.06) and in Asians (P=0.0005, OR=0.75; 95% CI 0.64–0.88, P=0.71). Similarly, we found a significantly decreased risk of CRC for 1298C polymorphism (4,764 CRC patients and 6,592 controls) for a recessive genetic model worldwide (P=0.005, OR=0.81; 95% CI 0.70–0.94, P=0.40) and in Caucasians (P=0.04, OR=0.75 95% CI 0.57–0.99, P=0.35). No evidence of association of C677T (4,616 patients and 6,338 controls) and A1298C (1,272 patients and 1,684 controls) with colorectal adenoma were found. The evidence accumulated suggests that MTHFR may represent a low-penetrance susceptible gene for CRC, and that the two polymorphisms might protect against colorectal adenoma developing into cancer. A larger single study is required to further evaluate gene–gene and gene–environment interactions for MTHFR polymorphisms and the cancer risk in a specific population.
Similar content being viewed by others


Introduction
Colorectal cancer (CRC) is one of the most common cancers, and the development of CRC is determined by a series of risk factors, such as genetic polymorphisms, environmental factors, lifestyles, and the complex interaction between these factors (de Kok and van Maanen 2000). Colorectal adenomas are well-established precursor lesions for colorectal cancer (Fearon and Vogelstein 1990; Peipins and Sandler 1994); prevention of colorectal adenomas may decrease the occurrence of colorectal cancer.
It is widely accepted that gene methylation is implicated in carcinogenesis (Toyota et al. 1999). 5,10-Methylenetetrahydrofolate reductase (MTHFR) is involved in DNA methylation; it manipulates the levels of folate, one of the methyl group donors (Kim et al. 1996). MTHFR catalyzes the irreversible conversion of 5,10-methylenetetrahydrofolate (required for purine and thymidine synthesis) to 5-methyltetrahydrofolate, which is necessary for methionine synthesis (Ueland et al. 2001). Insufficient thymidylate results in uracil misincorporation into DNA, leading to single-strand and double-strand breaks and increasing the incidence of DNA misrepair, thus increasing the risk of genetic instability (Blount et al. 1997).
MTHFR C677T (Ala222Val) (Frosst et al. 1995) and A1298C (Glu429Ala) (van der Put et al. 1998) are two common polymorphisms. Both of these polymorphisms decrease the activity of the enzyme; this effect is more pronounced in the homozygous than the heterozygous state (Ogino and Wilson 2003). The MTHFR C677T polymorphism exerts opposite effects in colorectal carcinogenesis, depending on the balance of thymidylate synthesis and DNA methylation, which affects the folate pool (Giovannucci et al. 2003). An association between MTHFR and genetic susceptibility to CRC and colorectal adenoma has been widely documented but with inconsistent results. A single study may be underpowered to detect a possible small effect from a real association of these polymorphisms with CRC and colorectal adenoma, especially when the sample size is relatively small. Different populations and sampling methods were applied in each study, which makes it complicated to interpret the data. Thus, a rigorous quantitative synthesis method is required in order to assess the discrepancy.
Here, we performed a meta-analysis of all eligible case-control studies in order to address the association of MTHFR C677T and A1298C polymorphisms with CRC and colorectal adenoma. Our results indicated that both the 677T and 1298C alleles might have a protective effect from CRC across a worldwide population. The 677T allele may exert its effect on CRC in a recessive genetic model in Asians, while the 1298C allele may decrease CRC risk in a recessive genetic model in Caucasians. No associations of C677T and A1298C polymorphisms were found to increase the risk of colorectal adenoma, which indicated that the two polymorphisms may protect from the adenoma becoming malignant cancer. More studies are needed to further confirm the protective roles of these polymorphisms in CRC development, and their effects in different populations.
Materials and methods
Identification and eligibility of relevant studies
To identify all articles that examined the association of MTHFR C677T and A1298C polymorphisms with CRC and colorectal adenoma, we conducted a literature search of the PubMed database (from January 1991 to August 2006) using the following keywords and subject terms: “MTHFR,” “polymorphism,” and “colorectal cancer” or “colorectal adenoma.” “Colon cancer (adenoma)” and “rectal cancer (adenoma)” were used to replace “colorectal cancer (adenoma)” in further searches of related studies. References in retrieved articles were screened. Abstracts, case reports, editorials, and review articles were excluded. Studies included in the current meta-analysis had to meet all of the following criteria: (a) an unrelated case-control design was used, (b) genotype frequency was available, and (c) the genotype distribution of the control population must be in Hardy–Weinberg equilibrium (HWE).
Data extraction
Data were collected on the genotypes of C677T or A1298C based on colorectal cancer and colorectal adenoma, respectively. First author, year of publication, ethnicity of study population, and characteristics of cases and controls were described.
Statistical analysis
The meta-analysis examined the overall association of the C677T allele T with the risk of CRC and colonrectal adenoma compared to that for allele C; homozygote TT was contrasted with CC, recessive (TT vs. CT + CC) and dominant (TT + CT vs. CC) models for allele T. The same contrasts were performed for allele C of the A1298C polymorphism.
Odds ratios (OR) corresponding to a 95% confidence interval (CI) were applied to assess the strength of association of C677T or A1298C with CRC and colorectal adenoma since case-control studies were used, and OR was calculated according to the method of Woolf (1955). A chi-square-based Q statistic test was performed to assess the between-study heterogeneity (Lau et al. 1997). Heterogeneity was considered significant for P<0.10 because of the low power of the statistic. A fixed-effect model using the Mantel–Haenszel method and a random-effects model using the DerSimonian and Laird method were used to pool the results (Petitti 1994). In the absence of between-study heterogeneity, the two methods provide similar results. Random effects are more appropriate when heterogeneity is present. The significance of the pooled OR was determined by the Z test, a P value of <0.05 was considered significant. For each genetic contrast, subgroup analysis according to ethnicity was only considered for Asian and Caucasian populations, in order to estimate ethnic-specific OR. The A1298C comparisons for colorectal adenoma were not stratified for subgroup analysis because of the limited studies included.
Publication bias was investigated by funnel plot, in which the standard error in log(OR) in each study was plotted against OR. An asymmetric plot suggested possible publication bias. Funnel plot asymmetry was assessed via Egger’s linear regression test, a linear regression approach to measuring funnel plot asymmetry on the natural logarithm scale of the OR (Egger et al. 1997). The significance of the intercept was determined by the t-test as suggested by Egger, and a P value of <0.05 was considered significant.
Hardy–Weinberg equilibrium was tested for by the chi-square test for goodness of fit using a web-based program (http://ihg.gsf.de/cgi-bin/hw/hwa1.pl). Analyses were performed using the software Stata version 7, ReviewManage 4.2. All P values were two-sided.
Results
Selection of studies
Twenty-eight articles were retrieved based on the search criteria for colorectal cancer, and 20 met our inclusion criteria. For various reasons, eight articles were not included. Genotype distributions in the control populations in five studies (Le Marchand et al. 2005; Jiang et al. 2005; Ma et al. 1997; Chen et al. 2002; Miao et al. 2005) significantly deviated from HWE. No detailed genotyping information was available from Marugame et al. (2003) and Kawakami et al. (2003). One article (Slattery et al. 1997) was replaced with its updated study (Slattery et al. 1999). All 20 eligible articles described C677T (Chen et al. 1996; Park et al. 1999; Slattery et al. 1999; Ryan et al. 2001; Keku et al. 2002; Le Marchand et al. 2002; Sachse et al. 2002; Shannon et al. 2002; Heijmans et al. 2003; Plaschke et al. 2003; Pufulete et al. 2003; Toffoli et al. 2003; Curtin et al. 2004; Jiang et al. 2004; Kim et al. 2004; Ulvik et al. 2004; Yin et al. 2004; Matsuo et al. 2005; Otani et al. 2005; Wang et al. 2006), and 11 of them also provide data on A1298C (Table 1). Among the 20 eligible articles included, 70% (14/20) stated that the age and sex status were matched between the case and control populations. PCR-RFLP was used to validate genotype in all but two articles. Ulvik et al. (2004) used real-time PCR and Otani et al. (2005) used Taqman SNP genotyping assay. All of the articles used blood samples for genotyping except Shannon et al. (2002), which used frozen tissue samples.
Thirteen articles were retrieved for colorectal adenoma. Crabtree et al. (2004) only examined the MTHFR polymorphism with respect to the severity of the adenoma. Ulvik et al. (2001) contained no detailed information on genotyping. These two studies were discarded. Among 11 eligible articles (Chen et al. 1998; Ulrich et al. 1999; Levine et al. 2000; Marugame et al. 2000; Giovannucci et al. 2003; Pufulete et al. 2003; Boyapati et al. 2004; Goode et al. 2004; van den Donk et al. 2005; Hirose et al. 2005; Mitrou et al. 2006), 27% (3/11) stated that the age and sex status were matched between the case and control populations. PCR-RFLP was used to validate the genotype in all of the studies. All articles included in the meta-analysis provided information on C677T, and three of them also genotyped A1298C (Table 1).
Keku et al. (2002) provided data on two ethnicities (African–Americans and Caucasians), while Le Marchand et al. (2002) reported on three separate populations: Japanese, Caucasians, Hawaiians. Thus, each subpopulation in these two articles was treated as a separate study in the meta-analysis. Studies that provide genotyping data for mixed populations are indicated as having “unknown” ethnicity in Table 1.
Summary statistics
A total of 10,131 CRC patients and 15,362 controls (C677T), 4,764 CRC patients and 6,592 controls (A1298C), 4,616 adenoma patients and 6,338 controls (C677T), and 1,272 adenoma patients and 1,684 controls (A1298C) were investigated. Electronic Supplementary Material Table 1 shows the genotype and allele frequencies of both CRC and colorectal adenoma patients as well as the controls in the eligible studies. The allele frequencies were calculated for controls from the corresponding genotype distributions. The variant allele frequencies of 677T and 1298C in the individual studies are also shown in Table 1.
The C677 allele was a little less common among controls of Asian descent (62.4%, 95% CI 54.0–70.8) than in controls of European descent (67.8%, 95% CI 65.1–70.5). The A1298 allele was slightly more common in controls of Asian descent (77.7%, 95% CI 68.0–87.3) than in controls of European descent (71.0%, 95% CI 65.8–76.2). Overall, the prevalence of 677TT homozygosity was 15.6% and 16.8% in control subjects of Asian and European descent, respectively. The prevalence of 1298CC homozygosity was 5.3 and 9.0% in control subjects of Asian and European descent, respectively.
Genetic contrasts
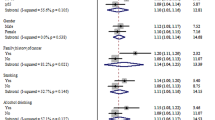
No significant heterogeneity existed between the 23 studies upon comparing the C677T C to the T allele in CRC (P=0.22). First, a fixed-effect model was used to pool the results (Fig. 1A). There was evidence that the T allele resulted in decreased susceptibility to CRC in a worldwide population. The overall OR was 0.93, 95% CI (0.89–0.98) by fixed effects (P=0.004). No significant between-study heterogeneity was found within each subgroup. The decreased effect of the T allele on the susceptibility was seen in both subgroups, with eight comparisons in populations of Asian descent (P=0.001, OR=0.87, 95% CI 0.81–0.95) and nine comparisons in those of Caucasian descent (P=0.03, OR=0.93, 95% CI 0.88–0.99). Meta-analyses for other genetic contrasts further suggested that the 677T allele was more likely to affect the CRC risk in a recessive genetic model worldwide (P=0.009, OR=0.86, 95% CI 0.76–0.96, random effects model), with between-study heterogeneity present (P=0.06). This result implies that a dose effect may exist for this polymorphism. Interestingly, this effect is only significant in Asians (P=0.0005, OR=0.75, 95% CI 0.64–0.88, P=0.71 for heterogeneity), not in Caucasians (P=0.16, OR=1.02, 95% CI 0.79–1.32, P=0.006 for heterogeneity). The TT versus CC contrast also showed that the association between C677T polymorphism and CRC risk was maintained in Asians but not Caucasians (Table 2).

Overall meta-analysis for C677T polymorphism (T vs. C allele) in CRC and colorectal adenoma. Point estimates of the OR for each study and the accompanying 95% CI values obtained with a fixed effects model are shown. A Analysis of the comparison with CRC, and B analysis of the comparison with colorectal adenoma. n indicates the total number of T or C alleles, N indicates the total number of T plus C alleles
Eleven studies investigated the association between C677T polymorphism and colorectal adenoma. No heterogeneity was found among the studies (P=0.87) when comparing the C to the T allele. No association was found with colorectal adenoma when a fixed model was used to perform a worldwide allele comparison (Fig. 1B, P=0.52, OR=0.98, 95% CI 0.93–1.04), as well as comparisons for Asian (P=0.78, OR=0.98, 95% CI 0.85–1.13) and Caucasian (P=0.35, OR=0.96, 95% CI 0.88–1.05) populations. No evidence of association between C677T and colorectal adenoma was discerned upon comparing with other genetic models.
There was heterogeneity among the 14 studies comparing the C versus the A allele (P=0.09) for A1298C polymorphism. Thus, the random effects model is more appropriate than the fixed model. The C allele tends to decrease the risk of CRC, but no significant association of the C allele with CRC susceptibility was found in a worldwide population (Fig. 2A, P=0.08, OR=0.93, 95% CI 0.85–1.01) and in an Asian population (P=0.2, OR=0.89, 95% CI 0.74–1.06, P=0.03 for heterogeneity). No heterogeneity (P=0.22) existed among the studies of Caucasian populations, and the fixed model did not reveal an association between 1298C and CRC risk (P=0.08, OR=0.90, 95% CI 0.79–1.01). Evidence for an association with CRC was discerned upon comparing the 1298CC genotype with the 1298AA genotype in a worldwide population (P=0.005, OR=0.80, 95% CI 0.69–0.94, P=0.18 for heterogeneity) and a Caucasian subgroup (P=0.04, OR=0.74, 95% CI 0.55–0.98, P=0.23 for heterogeneity). No association with 1298CC that decreases CRC risk in Asians was found using a random-effects model (P=0.35, OR=0.78, 95% CI 0.46–1.33, P=0.05 for heterogeneity) existed in the homozygote contrast. Importantly, the 1298C allele also tends to decrease CRC risk in a recessive genetic model worldwide (P=0.005, OR=0.81, 95% CI 0.70–0.94, P=0.40 for heterogeneity). This effect is only significant in the Caucasian subgroup (P=0.04, OR=0.75, 95% CI 0.57–0.99, P=0.35 for heterogeneity), not in Asians (P=0.17, OR=0.81, 95% CI 0.60–1.09, P=0.11 for heterogeneity) using a fixed effects model.

Overall meta-analysis for A1298C polymorphism (C vs. A allele) in CRC and colorectal adenoma. Point estimates of the OR values for the studies are shown, as well as the accompanying 95% CI values obtained using a random effects model in (A; analysis of the comparison with CRC) and using a fixed effects model in (B; analysis of the comparison with colorectal adenoma). n indicates the total number of C or A alleles, N indicates the total number of C plus A alleles
Only three studies reported A1298C polymorphism in colorectal adenoma, one for an unknown population and the other two for populations of Caucasian descent. Ethnic subgroup analysis was not performed due to limited studies. No association was found between A1298C and colorectal adenoma in a worldwide population when comparing the A and C alleles (Fig. 2B, P=0.15, OR=1.09, 95% CI 0.97–1.22, P=0.24 for heterogeneity), as well as for other genetic contrasts (Table 2).
Sensitivity analysis
Sensitivity analysis was performed by sequential omission of individual studies from various contrasts performed on a worldwide population and on Caucasian or Asian subgroups. The pooled ORs (including 95% CI) were consistently below 1.0 in all of the comparisons for the three groups (worldwide population, Caucasian, Asian), with either fixed or random effect models applied, except for the dominant genetic model of C677T. The results did not alter the protecting role exhibited by the 677T allele against the CRC risk when using a recessive genetic model, indicating that the significance of the pooled ORs was not excessively influenced by any single study (data not shown). When individual studies were sequentially omitted for A1298C under homozygote and recessive contrasts in the worldwide population and for Caucasians, the pooled ORs were consistently below 1.0, suggesting that the protecting role exhibited by 1298C against the CRC risk when using a recessive genetic model is not affected by a single study. The associations of the C677T and A1298C polymorphisms with colorectal adenoma did not change during the sensitivity analysis.
Publication bias
A funnel plot for the comparison of the 677C allele with the 677T allele in the OR analysis and Egger’s test provided no evidence for funnel plot symmetry for both colorectal adenoma (t=0.42, P=0.69) and CRC (t=1.50, P=0.15). Similarly, no publication bias was detected for the A versus C allele contrast of A1298C polymorphism for both colorectal adenoma (t=−0.36, P=0.72) and CRC (t=0.87, P=0.54).
Discussion
This meta-analysis examined the association of MTHFR C677T and A1298C polymorphisms with susceptibility to CRC and colorectal adenoma. Little et al. (2003) found a protective effect of MTHFR C677T against CRC risk using a meta-analysis on 14 studies in 2003, including Ma et al. (1997), which deviated from HWE. A protective effect of MTHFR A1298C against colorectal cancer risk was also found using six studies. No association of MTHFR C677T with colorectal adenoma was found in their meta-analysis using six studies. Our meta-analysis results are consistent with those of Little et al. (2003), which were obtained by recruiting more up-to-date studies. We also investigated the association of MTHFR A1298C with colorectal adenoma risk. Importantly, we analyzed the effect of C677T and A1298C on CRC and colorectal adenoma risk by stratifying the ethnic populations studied.
A significant association between the MTHFR C677T polymorphism and CRC in a worldwide population for the overall effect of allele T versus C was found. Subgroup analysis suggested 677C polymorphism was a protecting factor against susceptibility to CRC for Asians when either homozygous (TT vs. CC) or recessive genetic model contrasts were performed. Significant associations were found between MTHFR A1298C polymorphism and CRC for the homozygote contrast and recessive genetic models. However, the Asian population does not show an association in the homozygote and recessive genetic analysis, in contrast to Caucasians, which retain the protecting effect of the 1298C against CRC risk. Thus, both the 677T allele and the 1298C allele had a decreased effect on CRC risk under the recessive genetic model, but the effect may be more pronounced in Asians and Caucasians, respectively. We investigated five studies which provided case and control numbers of the nine combined genotypes of C677T and A1298C (Curtin et al. 2004; Jiang et al. 2004; Keku et al. 2002; Plaschke et al. 2003; Yin et al. 2004). The probabilities of 677TC + 1298CC, 677TT + 1298AC, and 677TT + 1298CC genotypes were very rare from 3,243 cases and 4,198 controls of those studies. Ogino and Wilson (2003) calculated the haplotype frequencies of the C677T and A1298C polymorphisms in pooled general populations derived from data published in 16 articles. They found that most 677T and 1298C alleles were associated with the 1298A and 677C alleles, respectively. There may be an increased frequency of the very rare cis 677T/1298C haplotype in some parts of the UK and Canada, possibly due to a founder effect. Chen et al. (2002) reported the linkage disequilibrium between C677T and A1298C polymorphisms in MTHFR. Therefore, the effects on the CRC risk are independent for the two polymorphisms; both the 677T and 1298C alleles may be causal variants.
The 677T allele and the 1298C allele may decrease the risk of CRC, but they show no association with the risk of colorectal adenoma in a worldwide population and in the subgroups in our meta-analysis. The results suggest that the two polymorphisms may have a protecting effect, preventing colorectal adenoma from transforming into malignant CRC, since colorectal adenomas are precursor lesions of colorectal cancer (Fearon and Vogelstein 1990; Peipins and Sandler 1994). It is possible that MTHFR polymorphisms could modify the association between diet and colorectal adenomas. However, various results have been reported and no conclusions are available as yet. The MTHFR TT genotype in combination with a low folate status may be a risk factor for colorectal adenomas (Ulrich et al. 1999; Levine et al. 2000), or there may be no interaction (van den Donk et al. 2005).
Reasons for the conflicting results where MTHFR C677T and A1298C play different roles in different studies may be genetic heterogeneity in different populations and clinical heterogeneity in different studies. Potentially, differences in patient populations (e.g., in terms of age and years from onset, gender difference, and lifestyle) might cause different results. Shannon et al. (2002) showed that the MTHFR 677TT genotype conferred an increased risk of colorectal cancer on subjects older but not younger than 70 years. The risk effect may be dependent on gene methylation, which is affected by folate intake and its interaction with other risk factors. Chen et al. (1996) suggested a possible interaction between the MTHFR C677T polymorphism and folate status in CRC. TT genotype was protective in folate-replete subjects, whereas the combination of TT and low folate status conferred no protection, and even increased risk. Otani et al. (2005) reported no association between MTHFR C677T and A1298C–nutrient interaction in CRC, including vitamin B6, vitamin B12 and folate. Moreover, the association of alcohol consumption with colorectal cancer risk has been related to its anti-folate effects and subsequent effects on DNA methylation (Freudenheim et al. 1991; Giovannucci et al. 1993). Thus, diet, particularly low folic acid intake, can modify the effects of the MTHFR polymorphisms. However, no definite conclusion about the relation between folate intake and CRC risk has been possible so far. More careful stratification analysis that takes into account the clinical character and diet, smoking status and alcohol consumption are needed.
The 677T and 1298C SNPs in the MTHFR gene both decrease the activity of the enzyme, leading to hyperhomocysteinemia, particularly in folate-deficient states (Frosst et al. 1995; van der Put et al. 1998). Elevated plasma homocysteine concentration had been suggested to be a risk factor for schizophrenia (Muntjewerff et al. 2006), coronary heart disease (Klerk et al. 2002), and stroke (Casas et al. 2005), particularly in low folate status. 677TT homozygotes show significantly greater mean homocysteine and risks of those three diseases than people who are 677CC homozygotes. However, Lewis et al. (2005) found no strong association of the MTHFR 677T polymorphism with coronary heart disease in Europe, North America, or Australia using a meta-analysis, which might due to the higher folate intake in North America and Europe. Here, folate provides one-carbon groups for the methylation of homocysteine to form methionine, which then decreases the concentration of homocysteine (Hankey and Eikelboom 2005). Zintzaras recently used meta-analysis to examine the association of MTHFR C677T and A1298C polymorphisms with breast cancer and gastric cancer, respectively (Zintzaras 2006a, 2006b). No association was found with breast cancer, but an increased risk from MTHFR 677T and 1298C was indicated for gastric cancer, in contrast to our meta-analysis results which indicate that MTHFR 677T and 1298C might be protective against CRC. Decreased levels of methyltetrahydrofolate may adversely affect the methylation of oncogenes and tumor suppressor genes, contributing to carcinogenesis. Also, depletion of methylenetetrahydrofolate interferes with thymidylate biosynthesis, which may lead to an accumulation of deoxyuridylate DNA, and subsequent removal of this abnormal base may destroy the integrity of DNA (Houlston and Tomlinson 2001). Thus, the balance between DNA synthesis and DNA methylation, which is determined by the MTHFR polymorphisms, may influence the cancer risk, which is also affected by folate intake.
Only two of the studies involved in our meta-analysis investigated gene–gene interactions. Ulvik et al. (2004) found no association of MTHFR C677T and MTR (methionine synthase) A2756G with CRC risk, and Hirose et al. (2005) revealed a decreased risk of CRC from ALDH2*2 and MTHFR 677T. More research on gene–gene interactions such as those involving MTR, thymidylate synthase, and ALDH2 will provide a more comprehensive insight into the associations studied here.
In conclusion, our meta-analysis (along with other studies) suggests that the MTHFR 677T and 1298C alleles have no association with colorectal adenoma risk, and they may both provide protecting effects against CRC risk in a recessive genetic model; however, the protecting effect of 677T was only significant in Asians, while 1298C was only significant in Caucasians. Whether other MTHFR polymorphisms provide protective effects against CRC risk in different populations in a recessive genetic model requires further investigation. More studies or large case-control studies should be performed to clarify the possible roles of C677T and A1298C in CRC.
Reference
Blount BC, Mack MM, Wehr CM, MacGregor JT, Hiatt RA, Wang G, Wickramasinghe SN, Everson RB, Ames BN (1997) Folate deficiency causes uracil misincorporation into human DNA and chromosome breakage: implications for cancer and neuronal damage. Proc Natl Acad Sci USA 94:3290–3295
Boyapati SM, Bostick RM, McGlynn KA, Fina MF, Roufail WM, Geisinger KR, Hebert JR, Coker A, Wargovich M (2004) Folate intake, MTHFR C677T polymorphism, alcohol consumption, and risk for sporadic colorectal adenoma (United States). Cancer Causes Control 15:493–501
Casas JP, Bautista LE, Smeeth L, Sharma P, Hingorani AD (2005) Homocysteine and stroke: evidence on a causal link from mendelian randomisation. Lancet 365:224–232
Chen J, Giovannucci E, Hankinson SE, Ma J, Willett WC, Spiegelman D, Kelsey KT, Hunter DJ (1998) A prospective study of methylenetetrahydrofolate reductase and methionine synthase gene polymorphisms, and risk of colorectal adenoma. Carcinogenesis 19: 2129–2132
Chen J, Giovannucci E, Kelsey K, Rimm EB, Stampfer MJ, Colditz GA, Spiegelman D, Willett WC, Hunter DJ (1996) A methylenetetrahydrofolate reductase polymorphism and the risk of colorectal cancer. Cancer Res 56:4862–4864
Chen J, Ma J, Stampfer MJ, Palomeque C, Selhub J, Hunter DJ (2002) Linkage disequilibrium between the 677C > T and 1298A > C polymorphisms in human methylenetetrahydrofolate reductase gene and their contributions to risk of colorectal cancer. Pharmacogenetics 12:339–342
Crabtree MD, Fletcher C, Churchman M, Hodgson SV, Neale K, Phillips RK, Tomlinson IP (2004) Analysis of candidate modifier loci for the severity of colonic familial adenomatous polyposis, with evidence for the importance of the N-acetyl transferases. Gut 53:271–276
Curtin K, Bigler J, Slattery ML, Caan B, Potter JD, Ulrich CM (2004) MTHFR C677T and A1298C polymorphisms:diet, estrogen, and risk of colon cancer. Cancer Epidemiol Biomarkers Prev 13:285–292
de Kok TM, van Maanen JM (2000) Evaluation of fecal mutagenicity and colorectal cancer risk. Mutat Res 463:53–101
Egger M, Davey Smith G, Schneider M, Minder C (1997) Bias in meta-analysis detected by a simple, graphical test. BMJ 315:629–634
Fearon ER, Vogelstein B (1990) A genetic model for colorectal tumorigenesis. Cell 61:759–767
Freudenheim JL, Graham S, Marshall JR, Haughey BP, Cholewinski S, Wilkinson G (1991) Folate intake and carcinogenesis of the colon and rectum. Int J Epidemiol 20:368–374
Frosst P, Blom HJ, Milos R, Goyette P, Sheppard CA, Matthews RG, Boers GJ, den Heijer M, Kluijtmans LA, van den Heuvel LP et al. (1995) A candidate genetic risk factor for vascular disease: a common mutation in methylenetetrahydrofolate reductase. Nat Genet 10:111–113
Giovannucci E, Chen J, Smith-Warner SA, Rimm EB, Fuchs CS, Palomeque C, Willett WC, Hunter DJ (2003) Methylenetetrahydrofolate reductase, alcohol dehydrogenase, diet, and risk of colorectal adenomas. Cancer Epidemiol Biomarkers Prev 12:970–979
Giovannucci E, Stampfer MJ, Colditz GA, Rimm EB, Trichopoulos D, Rosner BA, Speizer FE, Willett WC (1993) Folate, methionine, and alcohol intake and risk of colorectal adenoma. J Natl Cancer Inst 85:875–884
Goode EL, Potter JD, Bigler J, Ulrich CM (2004) Methionine synthase D919G polymorphism, folate metabolism, and colorectal adenoma risk. Cancer Epidemiol Biomarkers Prev 13:157–162
Hankey GJ, Eikelboom JW (2005) Homocysteine and stroke. Lancet 365:194–196
Heijmans BT, Boer JM, Suchiman HE, Cornelisse CJ, Westendorp RG, Kromhout D, Feskens EJ, Slagboom PE (2003) A common variant of the methylenetetrahydrofolate reductase gene (1p36) is associated with an increased risk of cancer. Cancer Res 63:1249–1253
Hirose M, Kono S, Tabata S, Ogawa S, Yamaguchi K, Mineshita M, Hagiwara T, Yin G, Lee KY, Tsuji A, Ikeda N (2005) Genetic polymorphisms of methylenetetrahydrofolate reductase and aldehyde dehydrogenase 2, alcohol use and risk of colorectal adenomas: Self-Defense Forces Health Study. Cancer Sci 96:513–518
Houlston RS, Tomlinson IP (2001) Polymorphisms and colorectal tumor risk. Gastroenterology 121:282–301
Jiang Q, Chen K, Ma X, Li Q, Yu W, Shu G, Yao K (2005) Diets, polymorphisms of methylenetetrahydrofolate reductase, and the susceptibility of colon cancer and rectal cancer. Cancer Detect Prev 29:146–154
Jiang QT, Chen K, Ma XY, Miao XP, Yao KY, Yu WP, Li LY, Zhu YM, Zhou HG (2004) A case-control study on the polymorphisms of methylenetetrahydrofolate reductases, drinking interaction and susceptibility in colorectal cancer. Zhonghua Liu Xing Bing Xue Za Zhi 25:612–616
Kawakami K, Ruszkiewicz A, Bennett G, Moore J, Watanabe G, Iacopetta B (2003) The folate pool in colorectal cancers is associated with DNA hypermethylation and with a polymorphism in methylenetetrahydrofolate reductase. Clin Cancer Res 9:5860–5865
Keku T, Millikan R, Worley K, Winkel S, Eaton A, Biscocho L, Martin C, Sandler R (2002) 5,10-Methylenetetrahydrofolate reductase codon 677 and 1298 polymorphisms and colon cancer in African Americans and whites. Cancer Epidemiol Biomarkers Prev 11:1611–1621
Kim DH, Ahn YO, Lee BH, Tsuji E, Kiyohara C, Kono S (2004) Methylenetetrahydrofolate reductase polymorphism, alcohol intake, and risks of colon and rectal cancers in Korea. Cancer Lett 216:199–205
Kim YI, Pogribny IP, Salomon RN, Choi SW, Smith DE, James SJ, Mason JB (1996) Exon-specific DNA hypomethylation of the p53 gene of rat colon induced by dimethylhydrazine. Modulation by dietary folate. Am J Pathol 149:1129–1137
Klerk M, Verhoef P, Clarke R, Blom HJ, Kok FJ, Schouten EG (2002) MTHFR 677C → T polymorphism and risk of coronary heart disease: a meta-analysis. JAMA 288:2023–2031
Lau J, Ioannidis JP, Schmid CH (1997) Quantitative synthesis in systematic reviews. Ann Intern Med 127:820–826
Le Marchand L, Donlon T, Hankin JH, Kolonel LN, Wilkens LR, Seifried A (2002) B-vitamin intake, metabolic genes, and colorectal cancer risk (United States). Cancer Causes Control 13:239–248
Le Marchand L, Wilkens LR, Kolonel LN, Henderson BE (2005) The MTHFR C677T polymorphism and colorectal cancer: the multiethnic cohort study. Cancer Epidemiol Biomarkers Prev 14:1198–1203
Levine AJ, Siegmund KD, Ervin CM, Diep A, Lee ER, Frankl HD, Haile RW (2000) The methylenetetrahydrofolate reductase 677C → T polymorphism and distal colorectal adenoma risk. Cancer Epidemiol Biomarkers Prev 9:657–663
Lewis SJ, Ebrahim S, Davey Smith G (2005) Meta-analysis of MTHFR 677C → T polymorphism and coronary heart disease: does totality of evidence support causal role for homocysteine and preventive potential of folate? BMJ 331:1053–1058
Little J, Sharp L, Duthie S, Narayanan S (2003) Colon cancer and genetic variation in folate metabolism: the clinical bottom line. J Nutr 133:3758S–3766S
Ma J, Stampfer MJ, Giovannucci E, Artigas C, Hunter DJ, Fuchs C, Willett WC, Selhub J, Hennekens CH, Rozen R (1997) Methylenetetrahydrofolate reductase polymorphism, dietary interactions, and risk of colorectal cancer. Cancer Res 57:1098–1102
Marugame T, Tsuji E, Inoue H, Shinomiya S, Kiyohara C, Onuma K, Hamada H, Koga H, Handa K, Hayabuchi H, Kono S (2000) Methylenetetrahydrofolate reductase polymorphism and risk of colorectal adenomas. Cancer Lett 151:181–186
Marugame T, Tsuji E, Kiyohara C, Eguchi H, Oda T, Shinchi K, Kono S (2003) Relation of plasma folate and methylenetetrahydrofolate reductase C677T polymorphism to colorectal adenomas. Int J Epidemiol 32:64–66
Matsuo K, Ito H, Wakai K, Hirose K, Saito T, Suzuki T, Kato T, Hirai T, Kanemitsu Y, Hamajima H, Tajima K (2005) One-carbon metabolism related gene polymorphisms interact with alcohol drinking to influence the risk of colorectal cancer in Japan. Carcinogenesis 26:2164–2171
Miao XP, Yang S, Tan W, Zhang XM, Ye YJ, Lin DX, Wang S (2005) Association between genetic variations in methylenetetrahydrofolate reductase and risk of colorectal cancer in a Chinese population. Zhonghua Yu Fang Yi Xue Za Zhi 39:409–411
Mitrou PN, Watson MA, Loktionov AS, Cardwell C, Gunter MJ, Atkin WS, Macklin CP, Cecil T, Bishop TD, Primrose J, Bingham SA (2006) MTHFR (C677T and A1298C) polymorphisms and risk of sporadic distal colorectal adenoma in the UK Flexible Sigmoidoscopy Screening Trial (United Kingdom). Cancer Causes Control 17:793–801
Muntjewerff JW, Kahn RS, Blom HJ, den Heijer M (2006) Homocysteine, methylenetetrahydrofolate reductase and risk of schizophrenia: a meta-analysis. Mol Psychiat 11:143–149
Ogino S, Wilson RB (2003) Genotype and haplotype distributions of MTHFR677C > T and 1298A > C single nucleotide polymorphisms: a meta-analysis. J Hum Genet 48:1–7
Otani T, Iwasaki M, Hanaoka T, Kobayashi M, Ishihara J, Natsukawa S, Shaura K, Koizumi Y, Kasuga Y, Yoshimura K, Yoshida T, Tsugane S (2005) Folate, vitamin B6, vitamin B12, and vitamin B2 intake, genetic polymorphisms of related enzymes, and risk of colorectal cancer in a hospital-based case-control study in Japan. Nutr Cancer 53:42–50
Park KS, Mok JW, Kim JC (1999) The 677C > T mutation in 5,10-methylenetetrahydrofolate reductase and colorectal cancer risk. Genet Test 3:233–236
Peipins LA, Sandler RS (1994) Epidemiology of colorectal adenomas. Epidemiol Rev 16:273–297
Petitti D (1994) Meta-analysis, decision analysis, and cost-effectiveness analysis. Oxford University Press, New York
Plaschke J, Schwanebeck U, Pistorius S, Saeger HD, Schackert HK (2003) Methylenetetrahydrofolate reductase polymorphisms and risk of sporadic and hereditary colorectal cancer with or without microsatellite instability. Cancer Lett 191:179–185
Pufulete M, Al-Ghnaniem R, Leather AJ, Appleby P, Gout S, Terry C, Emery PW, Sanders TA (2003) Folate status, genomic DNA hypomethylation, and risk of colorectal adenoma and cancer: a case control study. Gastroenterology 124:1240–1248
Ryan BM, Molloy AM, McManus R, Arfin Q, Kelleher D, Scott JM, Weir DG (2001) The methylenetetrahydrofolate reductase (MTHFR) gene in colorectal cancer: role in tumor development and significance of allelic loss in tumor progression. Int J Gastrointest Cancer 30:105–111
Sachse C, Smith G, Wilkie MJ, Barrett JH, Waxman R, Sullivan F, Forman D, Bishop DT, Wolf CR (2002) A pharmacogenetic study to investigate the role of dietary carcinogens in the etiology of colorectal cancer. Carcinogenesis 23:1839–1849
Shannon B, Gnanasampanthan S, Beilby J, Iacopetta B (2002) A polymorphism in the methylenetetrahydrofolate reductase gene predisposes to colorectal cancers with microsatellite instability. Gut 50:520–524
Slattery ML, Potter J, Caan B, Edwards S, Coates A, Ma KN, Berry TD (1997) Energy balance and colon cancer—beyond physical activity. Cancer Res 57:75–80
Slattery ML, Potter JD, Samowitz W, Schaffer D, Leppert M (1999) Methylenetetrahydrofolate reductase, diet, and risk of colon cancer. Cancer Epidemiol Biomarkers Prev 8:513–518
Toffoli G, Gafa R, Russo A, Lanza G, Dolcetti R, Sartor F, Libra M, Viel A, Boiocchi M (2003) Methylenetetrahydrofolate reductase 677 C → T polymorphism and risk of proximal colon cancer in north Italy. Clin Cancer Res 9:743–748
Toyota M, Ahuja N, Ohe-Toyota M, Herman JG, Baylin SB, Issa JP (1999) CpG island methylator phenotype in colorectal cancer. Proc Natl Acad Sci USA 96:8681–8686
Ueland PM, Hustad S, Schneede J, Refsum H, Vollset SE (2001) Biological and clinical implications of the MTHFR C677T polymorphism. Trends Pharmacol Sci 22:195–201
Ulrich CM, Kampman E, Bigler J, Schwartz SM, Chen C, Bostick R, Fosdick L, Beresford SA, Yasui Y, Potter JD (1999) Colorectal adenomas and the C677T MTHFR polymorphism: evidence for gene-environment interaction? Cancer Epidemiol Biomarkers Prev 8:659–668
Ulvik A, Evensen ET, Lien EA, Hoff G, Vollset SE, Majak BM, Ueland PM (2001) Smoking, folate and methylenetetrahydrofolate reductase status as interactive determinants of adenomatous and hyperplastic polyps of colorectum. Am J Med Genet 101:246–254
Ulvik A, Vollset SE, Hansen S, Gislefoss R, Jellum E, Ueland PM (2004) Colorectal cancer and the methylenetetrahydrofolate reductase 677C → T and methionine synthase 2756A → G polymorphisms: a study of 2,168 case-control pairs from the JANUS cohort. Cancer Epidemiol Biomarkers Prev 13:2175–2180
van den Donk M, Buijsse B, van den Berg SW, Ocke MC, Harryvan JL, Nagengast FM, Kok FJ, Kampman E (2005) Dietary intake of folate and riboflavin, MTHFR C677T genotype, and colorectal adenoma risk: a Dutch case-control study. Cancer Epidemiol Biomarkers Prev 14:1562–1566
van der Put NM, Gabreels F, Stevens EM, Smeitink JA, Trijbels FJ, Eskes TK, van den Heuvel LP, Blom HJ (1998) A second common mutation in the methylenetetrahydrofolate reductase gene: an additional risk factor for neural-tube defects? Am J Hum Genet 62:1044–1051
Wang J, Gajalakshmi V, Jiang J, Kuriki K, Suzuki S, Nagaya T, Nakamura S, Akasaka S, Ishikawa H, Tokudome S (2006) Associations between 5,10-methylenetetrahydrofolate reductase codon 677 and 1298 genetic polymorphisms and environmental factors with reference to susceptibility to colorectal cancer: a case-control study in an Indian population. Int J Cancer 118:991–997
Woolf B (1955) On estimating the relation between blood group and disease. Ann Hum Genet 19:251–253
Yin G, Kono S, Toyomura K, Hagiwara T, Nagano J, Mizoue T, Mibu R, Tanaka M, Kakeji Y, Maehara Y, Okamura T, Ikejiri K, Futami K, Yasunami Y, Maekawa T, Takenaka K, Ichimiya H, Imaizumi N (2004) Methylenetetrahydrofolate reductase C677T and A1298C polymorphisms and colorectal cancer: the Fukuoka Colorectal Cancer Study. Cancer Sci 95:908–913
Zintzaras E (2006a) Association of methylenetetrahydrofolate reductase (MTHFR) polymorphisms with genetic susceptibility to gastric cancer: a meta-analysis. J Hum Genet 51:618–624
Zintzaras E (2006b) Methylenetetrahydrofolate reductase gene and susceptibility to breast cancer:a meta-analysis. Clin Genet 69:327–336
Acknowledgments
This research is supported by grant from the National Natural Science Foundation of China (30371422).
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Rights and permissions
About this article
Cite this article
Huang, Y., Han, S., Li, Y. et al. Different roles of MTHFR C677T and A1298C polymorphisms in colorectal adenoma and colorectal cancer: a meta-analysis. J Hum Genet 52, 73–85 (2007). https://doi.org/10.1007/s10038-006-0082-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10038-006-0082-5
Keywords
This article is cited by
-
MTHFR C677T and A1298C polymorphism’s effect on risk of colorectal cancer in Lynch syndrome
Scientific Reports (2023)
-
Genetic impact of methylenetetrahydrofolate reductase (MTHFR) polymorphism on the susceptibility to colorectal polyps: a meta-analysis
BMC Medical Genetics (2019)
-
Alcohol consumption, genetic variants in the alcohol- and folate metabolic pathways and colorectal cancer risk: the JPHC Study
Scientific Reports (2016)
-
Puzzling role of genetic risk factors in human longevity: “risk alleles” as pro-longevity variants
Biogerontology (2016)
-
Association between MTHFR polymorphisms and overall survival of colorectal cancer patients in Northeast China
Medical Oncology (2013)
