
Abstract
Pediatric leukemia survival rates have improved dramatically over the past decades. However, current treatment protocols are still largely ineffective in cases of relapsed leukemia and are associated with a significant rate of chronic health conditions. Thus, there is a continued need for new therapeutic options. Here, we show that mer receptor tyrosine kinase (MerTK) was abnormally expressed in approximately one half of pediatric T-cell leukemia patient samples and T-cell acute lymphoblastic leukemia (T-ALL) cell lines. Stimulation of MerTK by the ligand Gas6 led to activation of the prosurvival proteins Erk 1/2 and Stat5, and MerTK-dependent activation of the STAT pathway in leukemia represents a novel finding. Furthermore, inhibition of MerTK expression increased the sensitivity of T-ALL cells to treatment with chemotherapeutic agents and decreased the oncogenic potential of the Jurkat T-ALL cell line in a methylcellulose colony-forming assay. Lastly, inhibition of MerTK expression significantly increased median survival in a xenograft mouse model of leukemia (30.5 days vs 60 days, P<0.0001). These results suggest that inhibition of MerTK is a promising therapeutic strategy for the treatment of leukemia and may allow for dose reduction of currently used chemotherapeutics resulting in decreased rates of therapy-associated toxicities.
Similar content being viewed by others
Introduction
Cancer is the leading cause of disease-related deaths in children, and leukemia is the most common form of childhood cancer.1, 2 Leukemia survival rates have improved significantly in the last three decades as treatment protocols have been optimized. However, this success is associated with a two to fourfold increased rate of adverse late effects of therapy.3, 4 Furthermore, children with T-cell acute lymphoblastic leukemia (T-ALL) who experience a bone marrow relapse have a particularly poor prognosis,5, 6 and currently the best therapeutic option is hematopoietic stem cell transplantation. Unfortunately, induction of complete remission, a necessary first step for transplantation, is extremely difficult in relapsed T-ALL.7 Consequently, there continues to be a need for new therapeutic strategies, such as molecularly targeted therapies, that can interact synergistically with current treatment protocols and result in improved efficacy and reduced side effects.
Mer receptor tyrosine kinase (MerTK), a member of the TAM (Tyro3/Axl/Mer) family of receptor tyrosine kinases, was initially cloned from a B-lymphoblastoid complementary DNA library.8 In normal hematopoiesis, MerTK is important in macrophage and dendritic cell inflammatory response and MerTK promotes phagocytosis of apoptotic cells. Other physiological roles for MerTK in hematopoietic cells, include natural killer-cell differentiation9 and platelet aggregation.10 MerTK is activated by the ligands Protein S and Gas6, two structurally similar vitamin K-dependent proteins.11, 12 Both ligands are produced by a variety of tissues and are present in serum.13, 14 Interestingly, overexpression of both Gas6 and Protein S is correlated with poor prognosis in a variety of cancers.15 More recently, Tubby, tubby-like protein 1 and galectin-3 have been described as novel Mer ligands important in phagocytosis.16, 17 Although galectin-3 has been shown to be important in tumorigenesis,18 it is unclear whether Tubby or tubby-like protein 1 have a role in cancer.
The oncogenic potential of MerTK is supported by various lines of evidence. MerTK activation results in upregulation of the MAPK and PI3K/Akt antiapoptotic signaling pathways, thereby promoting survival of tumor cells. Expression of activated MerTK is transforming in Ba/F3 pro-B lymphocytes and NIH 3T3 fibroblasts;19, 20 MerTK overexpression has been reported in a variety of human cancers, including B- and T-ALL21, 22 and ectopic expression of MerTK in lymphocytes in a transgenic mouse model promotes the development of leukemia/lymphoma.23 Additionally, our lab and others have shown that MerTK inhibition leads to increased sensitivity of glioblastoma24 and non small-cell lung cancer 25 cell lines to treatment with chemotherapeutic agents. Taken together, these data indicate important roles for MerTK in leukemogenesis and chemoresistance.
Here, we extend our previous work on MerTK expression in T-ALL.21 We show increased expression of MerTK in a prospective analysis of pediatric T-cell ALL at the time of diagnosis. Furthermore, we show that MerTK stimulation leads to activation of the MAPK pathway and the signal transducers and activators of transcription (STAT) proteins, two prosurvival signaling pathways. Lastly, we demonstrate that MerTK inhibition sensitizes leukemia cells to treatment with chemotherapeutic drugs, and reduces their leukemogenic potential in a xenograft mouse model of leukemia. These data support the development of MerTK-targeted therapies for the treatment of pediatric T-ALL.
Materials and methods
Patient samples and cell culture
Diagnostic bone marrow or peripheral blood samples were obtained at the Children’s Hospital Colorado (Aurora, CO, USA) according to an institutionally approved COMIRB protocol. The CEM, HSB-2, Jurkat, Loucy, Molt3 and Molt4 cell lines were obtained from the American Type Culture Collection (Manassas, VA, USA). The ALL-SIL, DND41, HPB-ALL, SupT11 and TALL-1 cell lines were obtained from the German Collection of Microorganisms and Cell Cultures (Braunschweig, Lower Saxony, Germany). All cell lines were maintained in RPMI medium (Hyclone Laboratories, Logan, UT, USA) supplemented with 10% heat inactivated fetal bovine serum (Atlanta Biologicals, Lawrenceville, GA, USA) and penicillin (100 Units/ml)/streptomycin (100 μg/ml) (Hyclone Laboratories, Logan, UT, USA). The identity of all cell lines was confirmed by genomic fingerprinting via short tandem repeat microsatellite loci analysis.
Flow cytometric analysis of cell surface proteins
Flow cytometric analysis of cell lines was performed as previously described.26 For flow cytometric analysis of T-ALL patient samples, cells were stained with mouse anti-human Mer 590,27 biotinylated anti-human Axl (R&D Systems, Minneapolis, MN, USA), ECD-linked anti-human CD45 (Beckman Coulter, Brea, CA, USA) and PC7-linked anti-human CD7 (Beckman Coulter). PE-linked donkey anti-mouse (Jackson ImmunoResearch Laboratories, West Grove, PA, USA) and FITC-linked Streptavidin (BD Biosciences, San Jose, CA, USA) were used to detect anti-hMer and anti-hAxl antibodies, respectively. Stained cells were washed and resuspended in phosphate-buffered saline and analyzed using a FC 500 flow cytometer (Beckman Coulter) and CXP data analysis software (R&D Systems). Mer and Axl expression was determined on live, CD45/CD7 double-positive cells, with ⩾20% expression considered positive.
Western blot analysis
Whole-cell lysates were prepared in lysis buffer (50 mM HEPES pH 7.5, 150 mM NaCl, 10 mM EDTA, 10% glycerol and 1% Triton X-100) supplemented with phosphatase inhibitors (0.1 mM Sodium pervanadate and 0.1 mM Na2MoO4) and protease inhibitors (Complete mini, Roche Molecular Biochemicals, Indianapolis, IN, USA). Sodium pervanadate was prepared per the Alliance for Cellular Signaling (http://www.signaling-gateway.org/) protocol. Samples were analyzed by SDS−polyacrylamide gel electrophoresis and immunoblotting with the following primary antibodies: anti-Actin (sc-1616, Santa Cruz Biotechnology, Santa Cruz, CA, USA); anti-phospho-Mer (pMer Y749/Y753/Y754, PhosphoSolutions, Aurora, CO, USA); anti-hMer (no. 1633-1, Epitomics, Burlingame, CA, USA); anti-phospho-p44/42 MAPK (Thr202/Tyr204, no. 9106), anti-p44/42 MAPK (no. 9102), anti-PARP (no. 9542), anti-Caspase 3 (no. 9662), anti-phospho-Stat5 (Y694, no. 9359), anti-Stat5 (no. 9358) (Cell Signaling Technology, Danvers, MA, USA). Primary antibodies were labeled with horseradish peroxidase-conjugated secondary antibodies (donkey-anti-goat, sc-2020, Santa Cruz Biotechnology; goat-anti-mouse, 170–6516, Bio-Rad Laboratories, Hercules, CA, USA; or donkey-anti-rabbit, 711-035-152, Jackson ImmunoResearch Laboratories, West Grove, PA, USA) and proteins were visualized by enhanced chemiluminescence (Perkin-Elmer, Waltham, MA, USA).
Human phospho-kinase array analysis
Jurkat cells were serum starved for 3 h, and then separate aliquots were treated with vehicle control or 200 nM rhGas6 (R&D Systems) for ten minutes. Cell lysate isolation and the phospho-array screen were conducted according to the Human Phospho-Kinase Array Kit (R&D Systems) protocol.
Lentiviral small hairpin RNA (shRNA) vectors, Mer expression vector, luciferase vector and target cell transduction
Lentiviral shRNA vectors shMer1 (targeting Mer) and shControl1 (targeting GFP) have been previously described.26 The lentiviral shRNA vector shControl2 (SHC002, Sigma, St. Louis, MO, USA) expresses an shRNA construct that does not target any human gene. shRNA expressing Jurkat cell lines have been previously described.26 shRNA expressing HSB2 cell lines were derived in the same manner as with the Jurkat cell lines. The identity of all shRNA expressing cell lines was confirmed by genomic fingerprinting. The construction of the Mer expression vector and generation of the stable Jurkat Mer add-back cell lines has been previously described.26 Details of luciferase tagging of Jurkat parental and shRNA derivative cell lines will be provided elsewhere.
Cellular metabolism assay
Initial experiments were performed to determine the maximum number of cells that could be plated while still remaining within the linear range of the colorimetric assay. Jurkat and HSB2 (both at 5 × 105 cells/ml) were plated in 96-well plates and cultured for 4 h or overnight, respectively. Chemotherapeutic agents or vehicle only were added at the appropriate concentrations and the cells were cultured for an additional 48 h. MTT (3-(4,5-Dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide, Sigma) reagent was added to the cells to a final concentration of 0.65 mg/ml for the final 3 h of culture. Solubilization solution (2X, 10% SDS in 0.01M HCl) was added overnight and optical density was determined at 562 nm with a reference wavelength of 650 nm. Relative cell numbers were calculated by subtraction of background absorbance and normalization to untreated controls. IC50 values were determined by nonlinear regression of the MTT assay data using Prism (Version 5.0, GraphPad Software, LaJolla, CA, USA).
Flow cytometric analysis of apoptosis
Jurkat or HSB2 parental and shRNA derivative cells were plated at 5 × 105 cells/ml and cultured for 48 h in the presence of the indicated agents or vehicle control. Cells were collected by centrifugation (240 × g for 5 min), resuspended in phosphate-buffered saline containing 1 μM YO-PRO-1 (Invitrogen, Grand Island, NY, USA) and 1.5 μM propidium iodide (PI) (Invitrogen), and incubated on ice for 20–30 min. Fluorescence was detected and analyzed using a FC 500 flow cytometer and CXP data analysis software (Beckman Coulter).
Methylcellulose assay
Five hundred Jurkat parental and shRNA derivative cells were plated in methylcellulose-based medium (Cat no. 1101, Reachbio, Seattle, WA, USA) according to manufacturer’s protocol. Cells were grown for 8 days, then stained with MTT (0.5 mg/ml) overnight and visualized with a colony counter (GelCount, Oxford Optronix, Oxford, UK). Colonies greater than 50 μm in diameter were counted.
Xenograft model of leukemia and in vivo luciferase imaging
NOD scid gamma mice (Stock no. 5557, The Jackson Laboratory, Bar Harbor, ME, USA) were irradiated with 200 rads and injected intravenously with Jurkat or shRNA expressing cell lines (5 × 106 cells). Animals were monitored daily and sacrificed upon signs of leukemia onset (weight loss, decreased activity and/or hind limb paralysis). Whole blood, bone marrow and spleen were harvested at the time of sacrifice and analyzed for human cell-surface proteins CD2 and CD45 by flow cytometry. Mer surface expression was confirmed by flow cytometry and western blot. Control animals injected with phosphate-buffered saline did not develop leukemia. For luciferase imaging of leukemia progression, monoclonal Jurkat and shRNA-derivative cell lines expressing similar levels of luciferase were generated using a lentiviral system which will be described elsewhere. Mice inoculated with luciferase-expressing cell lines were injected with K+Salt D-Luciferin (Caliper Life Sciences, Hopkinton, MA, USA) and imaged with a Xenogen IVIS200 (Caliper Life Sciences, Hopkinton, MA, USA) imager. All experiments involving animals followed the regulatory standards approved by the University of Colorado Institutional Animal Care and Use Committee.
Statistical analysis
For determination of IC50 values, experiments were repeated 4–12 times. When the variability within a cell line was approximately the same among all of the cell lines being compared, repeated analysis of variance was used to estimate and compare the mean IC50 values. Otherwise, the difference in IC50 between cell lines was calculated and a t-test was used to determine if the mean difference was significantly different from 0. SAS 9.2 software (SAS Institute Inc., Cary, NC, USA. SAS/STAT 9.2) was used for the analyses. All other statistical analyses was carried out using Prism software (Version 5.0, GraphPad Software, LaJolla, CA, USA) Results were considered significant at P <0.05.
Results
Mer receptor tyrosine kinase is expressed in pediatric T-ALL
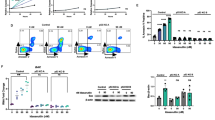
We have previously shown that MerTK is ectopically expressed in pediatric T-ALL patient samples using a retrospective analysis of banked T-cell leukemia patient samples (21). In this report, we expand on that observation by analyzing MerTK expression in a panel of eleven T-ALL cell lines. Western blot analysis of protein extract from each of the cell lines showed that 6 out of the 11 lines (54.5%) expressed MerTK protein at varying levels, with DND41 expressing the least and Loucy expressing the most (Figure 1a). The results of the western blot analysis and the relative levels of MerTK protein expression were also confirmed by flow cytometric analysis of surface protein expression (data not shown). The observed variability in MerTK expression levels is consistent with reports in other tumor types, including gliobastoma and non small-cell lung cancer, where negative as well as low and high MerTK expressing cell lines have been identified.23, 25

MerTK expression in T-ALL cell lines and diagnostic patient samples. (a) MerTK expression in a panel of T-ALL cell lines was detected by western blot analysis. (b) MerTK expression in diagnostic pediatric T-ALL patient samples was detected by flow cytometry analysis. Expression was determined after gating on live CD45 and CD7 positive lymphoblasts.
MerTK expression was also analyzed prospectively in a set of diagnostic samples obtained at the time of patient admission (see Table 1). Upon determination of the T-ALL diagnosis, CD45/CD7 double positive cells were tested for MerTK and Axl expression using flow cytometry (Figure 1b). As shown in Table 1, 5 out of 12 patient samples (41.7%) analyzed in the Children’s Hospital Colorado clinical pathology laboratory were classified as positive for MerTK expression, based on the established clinical criteria that at least 20% of cells must express a specific marker to be considered positive. As shown in Figure 1b, in MerTK positive samples the fluorescence intensity of the entire population shifted along the MerTK expression axis, suggesting that MerTK expression was increased in all cells in patient samples and was not solely due to the presence of a MerTK-expressing subclone. These prospective findings in patient samples, as well as the data for cell lines, are consistent with the findings previously reported by our lab, which showed MerTK mRNA expression in 19/34 (55.8%) banked patient samples and MerTK protein expression in 8/16 (50%) banked leukemia patient samples.21
Multiple signaling pathways are activated upon Gas6 stimulation of Mer receptor tyrosine kinase
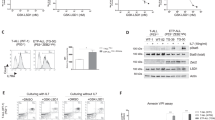
Previous reports have shown that a variety of signaling pathways are activated downstream of Gas6/MerTK signaling (reviewed in28, 29). We first analyzed the status of MerTK activation in the Jurkat and HSB2 cell lines and the ability of the receptor to respond to Gas6 stimulation (Figure 2a). MerTK was phosphorylated in both the Jurkat and HSB2 cell lines in response to Gas6 treatment. Additionally, in the Jurkat cell line there was low but detectable constitutive activation of MerTK, which was not present in the HSB2 cell line. To identify the pathways that might specifically be involved in the role of MerTK in T-ALL we used a human phospho-kinase array to analyze extracts from Jurkat cells treated with Gas6 or vehicle only (Figure 2b). As expected, we detected an increase in phosphorylated ERK and AKT in the Gas6 treated sample indicating the activation of the MAPK and PI3K/AKT pathways. Interestingly, an increase in pSTAT5 and pSTAT6 was also noted (Figure 2b), suggesting the activation of the Stat pathway in a MerTK dependent manner.

MAP Kinase and Stat pathway activation in response to MerTK Gas6 stimulation. (a) Analysis of phosphorylation status of MerTK in Jurkat and HSB2 cell lines during log growth (lane 1), or incubated in serum-free media for 3 h and then treated with vehicle control (lane 2) or Gas6 (200 nM) (lane 3). (b) Human phospho-kinase array analysis of the Jurkat cell line treated with Ga6 (200 nM) or vehicle control. (c and d) Western blot analysis of ERK and Stat5 activation in response to Gas6 (200 nM) treatment in (c) Jurkat and (d) HSB2 parental and shRNA derivative cell lines.
The results from the phospho-array screen were verified by western blot analysis in the T-ALL cell lines Jurkat and HSB2. Additionally, to confirm the requirement of MerTK in the Gas6 induced signaling, we derived cell lines where MerTK expression is inhibited by an shRNA construct (shMer1) targeting the 3′ UTR of the MerTK transcript. As shown in Figures 2c and d, MerTK protein expression was essentially ablated in two independent clones of Jurkat and HSB2 expressing shRNA directed against MerTK (shMer1A and shMer1B), while the cell lines expressing non-silencing control shRNA constructs (Jurkat shCntrl1 and HSB2 shCntrl2) showed no change in the level of MerTK protein.
Treatment of Jurkat and HSB2 parental and shControl cell lines with Gas6 led to an increase in the phosphorylation of Erk 1/2 (Thr202/Tyr204), whereas in the MerTK knockdown cell lines the activation of Erk was attenuated (Figure 2c and d), indicating that MerTK is required for Erk signaling in these cell lines. Similarly, when we analyzed the phosphorylation status of Stat5 in the Jurkat cell lines, both the Jurkat parental and shCntrl1 cell lines showed an increase in phosphorylated Stat5 (Tyr694) (pStat5), and this signal was absent in the Jurkat MerTK knockdown cell lines (Figure 2c).
In contrast to the Jurkat cell lines, the HSB2 parental and shCntrl2 cell lines showed a significant basal level of pStat5 (Figure 2d), which was largely unresponsive to treatment with Gas6. Interestingly, in the HSB2 MerTK knockdown cell lines, basal levels of pStat5 were reduced compared with the controls. However, when the total levels of Stat5 protein were analyzed, the knockdown cell lines showed a lower level of total Stat5 than in the HSB2 parental or shCntrl2 cell lines. These findings suggest that MerTK may have a role in the regulation of Stat5 activation and expression.
Inhibition of MerTK increased the chemosensitivity of T-ALL cell lines
As MerTK stimulation leads to the activation of Erk 1/2 (Figure 2) and other prosurvival pathways,19, 30 we decided to determine the effect of MerTK inhibition on the response of the Jurkat and HSB2 cell lines to treatment with cytotoxic agents that are currently used clinically.31 Using the MTT assay, the IC50 of the HSB2 parental and shRNA derivative cell lines in response to treatment with Cytarabine (Ara-C), Etoposide or 6-mercaptopurine (6-MP) were determined; similarly, the response of the Jurkat cell lines to treatment with Methotrexate was also analyzed.
As can be seen in Figures 3a, b and c), inhibition of MerTK increased the sensitivity of HSB2 to Ara-C, Etoposide and 6-MP. Comparison of the IC50 values (Table 2), generated by nonlinear regression of the MTT assay data, showed that the HSB2 shMer knockdown cell lines were significantly more sensitive to Etoposide (IC50=20.5 and 25.8 nM) compared with HSB2 shCntrl2 cells (IC50=44.1 nM; P=0.014 and 0.047 vs HSB2 shMer1A and shMer1B, respectively). In response to 6-MP, there was a statistically significant ∼1.8-fold increase in the sensitivity of the HSB2 shMer cells (IC50=2.78 and 2.63 μM) in comparison with the HSB2 shCntrl2 cells (4.79 μM; P=0.019 and 0.033 vs HSB2 shMer1A and shMer1B, respectively). In response to treatment with Ara-C, there was a ∼fourfold decrease in the IC50 of the HSB2 shMer cells (7.5 and 9.8 nM), relative to shCntrl2 cells (39.3 nM); however, this decrease was not statistically significant (P=0.125 and 0.162 vs HSB2 shMer1A and shMer1B, respectively). We saw a similar result when the Jurkat cell lines were tested in response to treatment with Methotrexate (Figure 3d), with both Jurkat shMer knockdown lines (IC50=44.8 and 43 nM) significantly more sensitive than the Jurkat shCntrl cell line (IC50=64.3 nM; P=0.005 and 0.033 vs Jurkat shMer1A and shMer1B, respectively) (Table 2).

Inhibition of MerTK leads to increased sensitivity of HSB2 and Jurkat cell lines to chemotherapeutic agents. MTT analysis of HSB2 and Jurkat parental and shRNA derivative cell lines in response to increased concentrations of chemotherapeutic agents. (a, b, c) Analysis of HSB2 treated with Ara-C, Etoposide or 6-MP, respectively. (d) Analysis of Jurkat treated with Methotrexate. Error bars represent standard error of the mean derived from at least four independent experiments.
The observed changes in chemosensitivity of the shMer knockdown cell lines in the MTT analysis can be attributed to either a decrease in proliferation or an increase in cell death. To determine whether decreased Mer expression enhanced cell death in response to treatment with chemotherapeutic agents, the levels of apoptosis were analyzed with a fluorescence-based assay, using YOPRO-1 and PI to stain apoptotic and dead cells, respectively. As shown in (Figure 4a), both HSB2 shMer knockdown cell lines had a statistically significant increase in the percentage of dead cells in response to treatment with 20 nM Ara-C (70.8 and 44%) or 6-MP (52–68.5 and 32.9–54.6%), compared with the HSB2 shCntrl2 cell line (21% for 20 nM Ara-C and 16.5–35.9% for 6-MP, P<0.001). Similarly, when Jurkat cells were treated with 60 nM Methotrexate (Figure 4b), there was a significantly increased level of apoptotic cells in the shMer knockdown cell lines (31.9 and 30.7%) compared with the Jurkat shCntrl1 cell line (13%; P<0.01). Analysis of the results from the YOPRO/PI assay using the Bliss independence model demonstrated an additive or synergistic increase in sensitivity to chemotherapy treatment in both the HSB2 and Jurkat cell lines depending on the concentrations of chemotherapy used (Supplementary Table 1).

Inhibiton of MerTK leads to increased apoptosis in HSB2 and Jurkat cell lines in response to treatment with chemotherapeutic agents. (a and b) Flow cytometry analysis of apoptotic and dead cells stained with YOPRO and PI. Error bars represent s.e. of the mean of at least three independent experiments. Statistical comparison vs shControl by two-way repeated measures analysis of variance followed by Bonferroni posttests(**P<0.01, ***P<0.001) (c and d) western blot analysis of Caspase 3 activation and PARP cleavage in (c) HSB2 and (d) Jurkat parental and shRNA derivative cell lines in response to Ara-C or Methotrexate (MTX) treatment, respectively.
To confirm the results from the YOPRO/PI analysis on a molecular level, the status of the apoptosis signaling effectors Caspase 3 and PARP were analyzed. As shown in Figure 4c, when HSB2 shMer cells were treated with Ara-C, there was an increase in cleaved Caspase 3 and cleaved PARP compared with the parental and shCntrl2 cell lines, consistent with the increased apoptosis observed in the YOPRO/PI assay (Figure 4a). Similar results were observed in the Jurkat cell lines (Figure 4d), with both the Jurkat shMer cell lines showing increased activation of Caspase 3 and PARP cleavage when treated with Methotrexate relative to the Jurkat parental and shCntrl1 cell lines.
Inhibition of MerTK decreases the oncogenic potential of the Jurkat cell line
To determine whether MerTK inhibition resulted in any changes in the clonogenic potential of T-ALL, the colony forming potential of the Jurkat parental and shRNA derived cell lines in methylcellulose media was analyzed. As shown in Figure 5a there was a significant decrease in the ability of the shMer knockdown cell lines to form colonies (75 and 67colonies) relative to the shCntrl1 cell line (185 colonies, P<0.01), indicating that MerTK activity contributes to the clonogenic potential of the Jurkat cell line. The colony formation assay was not carried out with the HSB2 cell line, as the cells did not form distinct colonies in this assay.

Inhibition of MerTK impairs the clonogenic potential of the Jurkat cell line. (a) Jurkat parental and shRNA derivative cells were plated in methylcellulose media, grown for 8 days, and colonies were counted. (b) Analysis of methylcellulose colony formation in Jurkat parental and shRNA derivative cells transduced with an empty (pLNCX) or a MerTK expression vector. Error bars represent the s.e. of the mean of at least three independent experiments. Statistical comparison vs shCntrl1 by one-way repeated measures analysis of variance followed by Bonferroni posttests. The differences in colony number between the Jurkat parental, shCntrl1 and shMer1A+MerWT lines were not statistically significant.
In order to control for the possibility of off-target effects of the shRNA used for Mer knockdown, we generated a MerTK add-back cell line derived from the Jurkat shMer1A cell line. As is shown in Figure 5b, reintroduction of MerTK expression in the knockdown cell line restores the clonogenic potential of the cells to the same level as the Jurkat parental and shCntrl1 cell lines with the empty vector only. Additionally, the increased chemosensitivity in the knockdown cell lines is reversed by exogenous MerTK expression (Supplementary Figure 1), suggesting that the phenotypes observed as a result of shRNA-mediated knockdown are not due to off-target effects and are specific to MerTK inhibition.
To further investigate the effect of MerTK on the oncogenic potential in T-ALL, a mouse xenograft model of leukemia was developed by transplating Jurkat parental and shRNA-derived cell lines into NOD scid gamma mice. A significant survival advantage was observed when mice were injected with Jurkat shMer cell lines, compared with those injected with Jurkat shCntrl1 (P<0.0001) (Figure 6a). Kaplan−Meier survival curves were used to determine a media survival of 60 and 59 days for mice injected Jurkat shMer1A and shMer1B, respectively, compared with a median survival of 30.5 days for mice injected with the Jurkat shCntrl1 cell line. Interestingly, a slight difference in survival was also noted between mice injected with the control cell line compared with those injected with the Jurkat parental cell line (27 vs 30.5 days).

Inhibition of MerTK significantly improves survival and delays disease progression in a Jurkat induced xenograft model of human leukemia. (a) Kaplan−Meier survival curves derived from NOD scid gamma mice mice injected with Jurkat parental or shRNA derivative cell lines. (b) Luciferase imaging of leukemia progression in NOD scid gamma mice mice injected with luciferase-tagged Jurkat and shRNA derivative cell lines and luciferin.
In order to visualize the extent of the changes in leukemia onset and progression in a more dynamic manner, mice were transplanted with monoclonal Jurkat cell lines (parental, shControl and shMer) expressing luciferase and luciferase activity was visualized over time as leukemia developed. Consistent with the data from the survival curve, mice injected with the parental cell line showed a slightly greater leukemia burden earlier on relative to the shCntrl1-injected mice (Figure 6b). Furthermore, we are able to visually observe the significant difference in survival induced by MerTK knockdown, as the mice that were injected with the Jurkat shMer1A/Luc cell line had dramatically less leukemia burden at days 17 and 24 postinjection relative to either the parental or shCntrl1 cell lines.
Discussion
In this paper, we have extended our previous findings demonstrating ectopic expression of MerTK in pediatric T-cell ALL.21 We detected MerTK expression in 5 out of 12 (41.7%) pediatric patient samples at the time of diagnosis, and in 6 out of 11 (54.5%) T-ALL cell lines analyzed. These findings are consistent with the data previously reported (MerTK expression detected in 50–55.8% of banked pediatric T-ALL patient samples),21 and also demonstrate the relative ease with which MerTK expression analysis can be incorporated into current flow cytometry diagnostic protocols.
To better understand the mechanism by which MerTK activation provides an advantage in T-cell leukemia cells, we screened a human phospho-kinase array for changes in phosphorylation status of proteins in response to Gas6-mediated activation of MerTK. Of the 38 different phospho-proteins present on the array, six proteins exhibited a change in phosphorylation status, including the ERK1/2 and AKT kinases and the STAT5 protein. Activation of the STAT pathway has been observed in AML and ALL, and is known to occur through a variety of mechanisms, including expression of Jak kinase fusion proteins (for example, TEL-JAK2).32 Although there are a few reports of STAT activation downstream of the TAM (Tyro3/Axl/Mer) receptors,33 MerTK-dependent STAT phosphorylation has only been observed in COS cells overexpressing a constitutively active chimeric MerTK receptor.34 In this report, we show that Stat5 is phosphorylated in response to Gas6 treatment and that MerTK inhibition reduces this response. Our data are the first to demonstrate a ligand-dependent role for endogenous MerTK protein in STAT signaling. In addition, STAT signaling as a consequence of MerTK activation has not been previously reported in leukemia. The mechanism of MerTK-dependent Stat5 phosphorylation will require further study, as we were not able to detect changes in Jak kinase activation (data not shown). It is possible that Stat5 phosphorylation could be mediated by another kinase, such as Src or MAP kinases,35 which have also been shown to be activated in response to MerTK stimulation.
Both the ERK1/2 and AKT kinases are well known modulators of antiapoptotic signals in cancer cells and have been shown to be downstream of MerTK stimulation. Both pathways can be activated by a variety of inputs and mechanisms. For example, in Jurkat cells AKT is constitutively active due to a mutation in PTEN, a phosphatase that regulates AKT activity.36 Thus, AKT activity in Jurkat cells was not significantly affected by MerTK stimulation or inhibition (data not shown). ERK1/2 activation, however, was responsive to Gas6 treatment in the Jurkat and HSB2 cells, and was blunted in the shRNA derived Mer knockdown cell lines.
The results of our signaling analysis suggests that targeted inhibition of MerTK would reduce prosurvival signaling in cells and result in a more effective outcome in response to cytotoxic treatment. Accordingly, both Jurkat and HSB2 MerTK knockdown cells were more sensitive to treatment with chemotherapeutic agents currently used for the treatment of leukemia.31 The increased sensitivity was mediated by a higher rate of apoptosis, which was detected by increased levels of caspase 3 and PARP cleavage. Interestingly, MerTK inhibition also reduced the clonogenic potential of the Jurkat cell line, both in an in vitro assay and in a mouse model of leukemia. Importantly, we were able to rescue the increased chemosensitivity and reduced clonogenic potential in the MerTK knockdown cell lines by exogenous expression of MerTK, indicating that the results presented here are unlikely to be due to off-target effects of the shRNA construct.
In conclusion, in this report we have confirmed that MerTK is ectopically expressed in ∼50% of patients with T-cell ALL, and we demonstrated that MerTK expression analysis can be incorporated into current flow cytometry diagnostic protocols. We have shown that MerTK activation leads to upregulation of prosurvival signaling pathways in T-cell lines, and identified novel downstream signaling via Stat5. Furthermore, we have shown that inhibition of MerTK increases the sensitivity of leukemia cells to treatment with cytotoxic agents, decreases their colony-forming potential, and MerTK inhibition also decreased the leukemogenic potential of T-cell lines in a mouse model. We believe these results offer compelling evidence that MerTK is a viable target for the development of targeted inhibitors that can be used in combination with cytotoxic therapies for T-cell leukemia, and that these inhibitors will increase the benefit of current treatment protocols while allowing for dose reduction and a decrease in the severity of the side effects that are now observed.
References
Jemal A, Siegel R, Ward E, Hao Y, Xu J, Thun MJ . Cancer statistics, 2009. CA Cancer J Clin 2009; 59: 225–249.
Kaatsch P . Epidemiology of childhood cancer. Cancer Treat Rev 2010; 36: 277–285.
Landier W, Bhatia S . Cancer survivorship: a pediatric perspective. Oncologist 2008; 13: 1181–1192.
Oeffinger KC, Mertens AC, Sklar CA, Kawashima T, Hudson MM, Meadows AT et al. Chronic health conditions in adult survivors of childhood cancer. N Engl J Med 2006; 355: 1572–1582.
Goldberg JM, Silverman LB, Levy DE, Dalton VK, Gelber RD, Lehmann L et al. Childhood T-cell acute lymphoblastic leukemia: the Dana-Farber Cancer Institute acute lymphoblastic leukemia consortium experience. J Clin Oncol 2003; 21: 3613–3622.
Henze G, Fengler R, Hartmann R, Kornhuber B, Janka-Schaub G, Niethammer D et al. Six-year experience with a comprehensive approach to the treatment of recurrent childhood acute lymphoblastic leukemia (ALL-REZ BFM 85). A relapse study of the BFM group. Blood 1991; 78: 1166–1172.
Raetz EA, Borowitz MJ, Devidas M, Hunger SP, Winick N, Camitta B et al. Outcomes of Children with First Marrow Relapse: Results from Children’s Oncology Group (COG) Study AALL01P2. Blood (ASH Annual Meeting Abstracts) 2006; 108: 1871.
Graham DK, Dawson TL, Mullaney DL, Snodgrass HR, Earp HS . Cloning and mRNA expression analysis of a novel human protooncogene, c-mer. Cell Growth Differ 1994; 5: 647–657.
Lemke G, Rothlin CV . Immunobiology of the TAM receptors. Nat Rev Immunol 2008; 8: 327–336.
Sather S, Kenyon KD, Lefkowitz JB, Liang X, Varnum BC, Henson PM et al. A soluble form of the Mer receptor tyrosine kinase inhibits macrophage clearance of apoptotic cells and platelet aggregation. Blood 2007; 109: 1026–1033.
Nagata K, Ohashi K, Nakano T, Arita H, Zong C, Hanafusa H et al. Identification of the product of growth arrest-specific gene 6 as a common ligand for Axl, Sky, and Mer receptor tyrosine kinases. J Biol Chem 1996; 271: 30022–30027.
Stitt TN, Conn G, Gore M, Lai C, Bruno J, Radziejewski C et al. The anticoagulation factor protein S and its relative, Gas6, are ligands for the Tyro 3/Axl family of receptor tyrosine kinases. Cell 1995; 80: 661–670.
Balogh I, Hafizi S, Stenhoff J, Hansson K, Dahlback B . Analysis of Gas6 in human platelets and plasma. Arterioscler Thromb Vasc Biol 2005; 25: 1280–1286.
Manfioletti G, Brancolini C, Avanzi G, Schneider C . The protein encoded by a growth arrest-specific gene (gas6) is a new member of the vitamin K-dependent proteins related to protein S, a negative coregulator in the blood coagulation cascade. Mol Cell Biol 1993; 13: 4976–4985.
Linger RM, Keating AK, Earp HS, Graham DK . Taking aim at Mer and Axl receptor tyrosine kinases as novel therapeutic targets in solid tumors. Expert Opin Ther Targets 2010; 14: 1073–1090.
Caberoy NB, Alvarado G, Bigcas JL, Li W . Galectin-3 is a new MerTK-specific eat-me signal. J Cell Physiol 2012; 227: 401–407.
Caberoy NB, Zhou Y, Li W . Tubby and tubby-like protein 1 are new MerTK ligands for phagocytosis. EMBO J 2010; 29: 3898–3910.
Newlaczyl AU, Yu LG . Galectin-3—a jack-of-all-trades in cancer. Cancer Lett 2011; 313: 123–128.
Georgescu MM, Kirsch KH, Shishido T, Zong C, Hanafusa H . Biological effects of c-Mer receptor tyrosine kinase in hematopoietic cells depend on the Grb2 binding site in the receptor and activation of NF-kappaB. Mol Cell Biol 1999; 19: 1171–1181.
Ling L, Kung HJ . Mitogenic signals and transforming potential of Nyk, a newly identified neural cell adhesion molecule-related receptor tyrosine kinase. Mol Cell Biol 1995; 15: 6582–6592.
Graham DK, Salzberg DB, Kurtzberg J, Sather S, Matsushima GK, Keating AK et al. Ectopic expression of the proto-oncogene Mer in pediatric T-cell acute lymphoblastic leukemia. Clin Cancer Res 2006; 12: 2662–2669.
Yeoh EJ, Ross ME, Shurtleff SA, Williams WK, Patel D, Mahfouz R et al. Classification, subtype discovery, and prediction of outcome in pediatric acute lymphoblastic leukemia by gene expression profiling. Cancer Cell 2002; 1: 133–143.
Keating AK, Salzberg DB, Sather S, Liang X, Nickoloff S, Anwar A et al. Lymphoblastic leukemia/lymphoma in mice overexpressing the Mer (MerTK) receptor tyrosine kinase. Oncogene 2006; 25: 6092–6100.
Keating AK, Kim GK, Jones AE, Donson AM, Ware K, Mulcahy JM et al. Inhibition of Mer and Axl receptor tyrosine kinases in astrocytoma cells leads to increased apoptosis and improved chemosensitivity. Mol Cancer Ther 2010; 9: 1298–1307.
Linger RM, Cohen RA, Cummings CT, Sather S, Migdall-Wilson J, Middleton DH et al. Mer or Axl receptor tyrosine kinase inhibition promotes apoptosis, blocks growth and enhances chemosensitivity of human non-small cell lung cancer. Oncogene 2012, (e-pub ahead of print 13 August 2012; doi:10.1038/onc.2012.355).
Migdall-Wilson J, Bates C, Schlegel J, Brandao L, Linger RM, DeRyckere D et al. Prolonged exposure to a Mer ligand in leukemia: Gas6 favors expression of a partial Mer glycoform and reveals a novel role for Mer in the nucleus. PLoS One 2012; 7: e31635.
Rogers AE, Le JP, Sather S, Pernu BM, Graham DK, Pierce AM et al. Mer receptor tyrosine kinase inhibition impedes glioblastoma multiforme migration and alters cellular morphology. Oncogene 2012; 31: 4171–4181.
Brandao L, Migdall-Wilson J, Eisenman K, Graham DK . TAM receptors in leukemia: expression, signaling, and therapeutic implications. Crit Rev Oncog 2011; 16: 47–63.
Linger RM, Keating AK, Earp HS, Graham DK . TAM receptor tyrosine kinases: biologic functions, signaling, and potential therapeutic targeting in human cancer. Adv Cancer Res 2008; 100: 35–83.
Demarchi F, Verardo R, Varnum B, Brancolini C, Schneider C . Gas6 anti-apoptotic signaling requires NF-kappa B activation. J Biol Chem 2001; 276: 31738–31744.
Estlin EJ . Continuing therapy for childhood acute lymphoblastic leukaemia: clinical and cellular pharmacology of methotrexate, 6-mercaptopurine and 6-thioguanine. Cancer Treat Rev 2001; 27: 351–363.
Lacronique V, Boureux A, Valle VD, Poirel H, Quang CT, Mauchauffe M et al. A TEL-JAK2 fusion protein with constitutive kinase activity in human leukemia. Science 1997; 278: 1309–1312.
Rothlin CV, Ghosh S, Zuniga EI, Oldstone MB, Lemke G . TAM receptors are pleiotropic inhibitors of the innate immune response. Cell 2007; 131: 1124–1136.
Zong C, Yan R, August A, Darnell JE, Hanafusa H . Unique signal transduction of Eyk: constitutive stimulation of the JAK-STAT pathway by an oncogenic receptor-type tyrosine kinase. EMBO J 1996; 15: 4515–4525.
Rawlings JS, Rosler KM, Harrison DA . The JAK/STAT signaling pathway. J Cell Sci 2004; 117 (Pt 8): 1281–1283.
Shan X, Czar MJ, Bunnell SC, Liu P, Liu Y, Schwartzberg PL et al. Deficiency of PTEN in Jurkat T cells causes constitutive localization of Itk to the plasma membrane and hyperresponsiveness to CD3 stimulation. Mol Cell Biol 2000; 20: 6945–6957.
Acknowledgements
We would like to thank the Core Facilities at the University of Colorado Cancer Center (flow cytometry) and Barbara Davis Center (DNA sequencing) for expert technical assistance. Funding: This article was supported in part by grants from the NIH (RO1CA137078, D.K.G.) and the National Heart, Lung and Blood Institute (F32HL096416, L.N.B.). Supported in part by NIH/NCRR Colorado CTSI Grant Number UL1 RR025780. Contents are the authors’ sole responsibility and do not necessarily represent official NIH views. Douglas K. Graham is the Damon Runyon Novartis Clinical Investigator, supported in part by the Damon Runyon Cancer Research Foundation (CI-39-07, D.K.G.).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies this paper on Blood Cancer Journal website
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article
Cite this article
Brandao, L., Winges, A., Christoph, S. et al. Inhibition of MerTK increases chemosensitivity and decreases oncogenic potential in T-cell acute lymphoblastic leukemia. Blood Cancer Journal 3, e101 (2013). https://doi.org/10.1038/bcj.2012.46
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/bcj.2012.46
Keywords
This article is cited by
-
TAM family kinases as therapeutic targets at the interface of cancer and immunity
Nature Reviews Clinical Oncology (2023)
-
Phosphatidylserine receptor-targeting therapies for the treatment of cancer
Archives of Pharmacal Research (2019)
-
Targeting Gas6/TAM in cancer cells and tumor microenvironment
Molecular Cancer (2018)
-
The proto-oncogene Mer tyrosine kinase is a novel therapeutic target in mantle cell lymphoma
Journal of Hematology & Oncology (2018)
-
Molecular insights of Gas6/TAM in cancer development and therapy
Cell Death & Disease (2017)

{kind=link}