
Abstract
Cancer initiation and progression are defined by the behavior of cancer cells per se and the development of tumor tissues, both of which are modulated by crosstalk between cancer cells and the surrounding microenvironment. Advances in cancer research have highlighted the significance of constant evolution of the tumor microenvironment, leading to tumor formation, metastasis and refractoriness to therapy. MicroRNAs (miRNAs) are small non-coding RNAs that function as major players of posttranscriptional gene regulation in diverse biological processes. They function as both tumor suppressors and promoters in many aspects of the autonomous behavior of cancer cells. Theoretically, dysfunction in the gene regulatory networks of cancer cells is one of the major driving forces for alterations of ostensibly normal surrounding cells. In this context, the core targets of miRNAs, termed miRNA regulons, are currently being expanded to include various modulators of the tumor microenvironment. Recent advances have highlighted two important roles played by miRNAs in the evolution of tumor microenvironments: miRNAs in tumor cells transform the microenvironment via non-cell-autonomous mechanisms, and miRNAs in neighboring cells stabilize cancer hallmark traits. These observations epitomize the distal and proximal functions of miRNAs in tumor microenvironments, respectively. Such regulation by miRNAs affects tumor angiogenesis, immune invasion and tumor–stromal interactions. This review summarizes recent findings on the mechanisms of miRNA-mediated regulation of tumor microenvironments, with a perspective on the design of therapeutic interventions.
Similar content being viewed by others


Introduction
Recent advances in cancer therapeutics have yielded several new-generation therapeutic modalities, and cancer prognoses have improved significantly. However, further advances are required before major improvements are achievable, especially in patients with advanced and/or intractable cancers, which are refractory to conventional therapeutic options. Although cancer therapies ideally seek eradication of all cancer cells in tumor tissues, recent work has emphasized that targeting of cancer cells is not equivalent to targeting of tumor tissues. The work suggests that cancer initiation and progression are defined by not only the behavior of cancer cells per se but also the development of tumor tissues, controlled in turn by crosstalk between cancer cells and the surrounding microenvironment.1, 2, 3 Recent cancer research has emphasized the significance of constant evolution of the tumor microenvironment, facilitating tumor formation, metastasis and development of refractoriness to cancer therapy.3
Tumor microenvironments are highly heterogeneous, containing various cell types, including fibroblasts, endothelial cells, pericytes, immune cells and local and bone marrow-derived stromal stem and progenitor cells, and a surrounding extracellular matrix (ECM) (Figure 1).1, 2, 3 These cells originate from normal cells but become altered during tumor development. For example, many types of solid tumor are accompanied by variable extents of stromal cell infiltration and ECM deposition, termed desmoplasia. Such cancer tissue remodeling allows tumor cells to grow and disseminate and contributes to an increase in interstitial fluid pressure, which can impede delivery of cancer drugs.2, 3, 4 Cancer-associated fibroblasts (CAFs) make versatile contribution to these responses.2,3

Components of tumor microenvironment. Tumor microenvironment is very heterogeneous and comprised of various cell types, such as CAFs, endothelial cells, pericytes, immune cells, including various types of lymphocytes, Treg, TAMs and MDSCs, and local and bone marrow-derived stromal stem and progenitor cells, and surrounding ECM.
MicroRNAs (miRNAs) are small non-coding RNAs that function as major players of posttranscriptional gene regulation within diverse cell types.5 Reflecting the importance in cell biology, they also have critical roles in cancer biology. In addition to various protein-coding oncogenes and tumor-suppressor genes, previous studies have demonstrated multifaceted roles of miRNAs in cancer cell behaviors, which constitute major aspects of conventional hallmarks of cancer, including sustained proliferation, resistance to cell death, evasion of growth suppressors, establishment of replicative immortality and acquisition of invasive phenotypes.1,6 For example, miR-34a is associated with growth control, miR-21 with cell proliferation and the miR-200 family with epithelial–mesenchymal transition (EMT).7
In addition, recent work has shown that miRNAs of cancer cells modulate the microenvironment via non-cell-autonomous mechanisms, and alterations in the miRNA profiles of neighboring cells that lack genetic abnormalities favor the acquisition of cancer hallmark traits, thereby unexpectedly expanding the roles of miRNAs in tumor microenvironments.8, 9, 10, 11, 12 Such actions include regulation of tumor angiogenesis, tumor immune invasion and tumor–stromal interactions. The present review focuses on the relationship between miRNA alterations in cancer cells and tumor microenvironments, and the mechanisms of miRNA-mediated regulation of tumor microenvironments, with the perspective of possible therapeutic interventions.
Dysfunction of miRNAS in cancer
In mammalian cells, the primary transcripts of miRNAs (primary miRNAs; pri-miRNAs) are transcribed by RNA polymerase II or III from miRNA-coding sequences, residing principally in intergenic regions or within introns of genes. Most pri-miRNAs are processed into short hairpin RNAs (precursor miRNAs; pre-miRNAs) by the Drosha/DGCR8 complex with RNase III activity in the nucleus. Pre-miRNAs are transported to the cytoplasm and further cleaved by a second RNase III Dicer to yield 21–25-nucleotide-long miRNA duplexes. Single RNA strands derived from such duplexes are incorporated as mature miRNAs within Argonaute proteins (Ago1-4) to form RNA-induced silencing complexes. In general, RNA-induced silencing complexes suppress the expression of various target genes through sequence complementarity between miRNAs and target mRNAs.5,13
Various mechanisms contribute to aberrant miRNA expression in cancer.13,14 As with abnormalities in oncogenes and tumor-suppressor genes, alterations in miRNAs can be explained in part by several mechanisms, including chromosomal deletion, amplification, mutation, epigenetic silencing and transcriptional dysregulation of pri-miRNA transcripts. On the other hand, concerning the characteristic process of miRNA biogenesis, impairment of miRNA processing might explain the downregulation of both broad groups of miRNAs and certain specific miRNAs.13,15 In addition, various miRNA-processing regulators, such as LIN28, KSRP and p53, have been identified to date at each step of miRNA biogenesis, suggesting multidimensional regulation of miRNA biogenesis.13,16, 17, 18 Furthermore, a recent report showed that Hippo signaling pathway regulated pri-miRNA processing in a cell-density-dependent manner, linking cell–cell contact to miRNA processing.19
Non-cell-autonomous function of cancer-related miRNAS
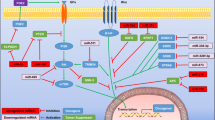
Most, if not all, of cancers are defined by the evolution of genomic abnormalities. Thus, in theory, alterations of ostensibly normal cells in tumor microenvironments will be largely attributable to dysfunction of the gene regulatory network of cancer cells per se. Recent advances have shown that miRNA dysfunction in tumor cells modulates the tumor microenvironment via non-cell-autonomous mechanisms (Figure 2), thus supporting this concept.10 In this context, the core targets of miRNAs, termed miRNA regulons, are currently being expanded to include various modulators of tumor microenvironments.8, 9, 10, 11, 12 This type of regulation has been also described in the context of protein-coding oncogenes and tumor-suppressor genes. For example, it has been shown that KRAS, mutation in which is an early event in the development of pancreatic ductal adenocarcinoma, enhances granulocyte macrophage colony-stimulating factor (GM-CSF) production from pancreatic ductal epithelial cells and thus increases recruitment of immunosuppressive Gr1+CD11b+ myeloid cells to suppress anti-tumor immunity.20,21 In this section, we focus on such distal impacts of cancer-associated miRNAs on three major components of the tumor milleu; these are the vasculature, ECM and immune cells.

Non-cell-autonomous roles of cancer cell miRNAs in regulation of tumor microenvironment. Alteration of miR-9 (a), miR-126 (b), miR-135b (c), miR-29b (d), miR-30b/d (e), miR-34a (f) and miR-199a-5p/3p and miR-1908 (g) in cancer cells elicit distal impacts on tumor microenvironment to promote tumor progression through various non-cell-autonomous mechanisms. Oncogenic miRNAs and tumor-suppressive miRNAs are represented by black and red, respectively.
Distal regulation of tumor angiogenesis by miR-9 and miR-126
Epithelial–mesenchymal transition (EMT) is an important step in cancer metastasis. Among a set of EMT-regulatory miRNAs, including miR-200, miR-103/107, miR-205, let-7 and miR-9,22 miR-9 has been shown to simultaneously modulate tumor angiogenesis through regulation of vascular endothelial growth factor (VEGF)-A (Figure 2a).23 In breast cancer cells, miR-9, induced by MYC and MYCN, targeted E-cadherin and increased cell motility and invasiveness. E-cadherin downregulation by miR-9 activated β-catenin signaling, in turn upregulating VEGF-A expression and increasing tumor angiogenesis.23 This report raised the possibility that miRNA targets diversify onto cell-autonomous regulators and non-cell-autonomous regulators in tumor progression.
This possibility was further solidified by studies on miR-126, the expression of which is frequently suppressed in various types of human cancer, including breast, gastric and colon cancers.24, 25, 26 Tavazoie et al.24 showed that miR-126 non-cell-autonomously restricted metastasis via regulation of multiple targets involved in endothelial recruitment (Figure 2b).27 They showed that endogenous miR-126 in breast cancer cells contributed to suppression of metastatic colonization and that the metastatic nodules developing upon miR-126 silencing displayed a denser vasculature. Mechanistically, miR-126 downregulates secretion of a soluble form of c-Mer tyrosine kinase receptor (MERTK), and insulin-like growth factor binding protein 2 (IGFBP2), from metastatic cells, by suppressing MERTK and inhibiting IGFBP2 and a regulatory gene thereof, PITPNC1 (encoding phosphatidylinositol transfer protein, cytoplasmic 1), respectively. Subsequently, downregulation of endogenous miR-126 promotes endothelial recruitment by increasing IGFBP2/IGF1/IGF1R signaling and attenuating GAS6/MERTK signaling, regulated by competition between the MERTK ligand GAS6 and MERTK, in endothelial cells. They further showed that overexpression of eight target genes of miR-126, including MERTK, IGFBP2 and PITPNC1, in primary breast cancer, was associated with shorter metastasis-free survival in multiple cohorts.27 This set of eight genes constitutes the core target genes of miR-126, namely the miR-126 regulon, which links the anti-metastatic activity of miR-126 to cancer–endothelial interactions.
In addition, Wang and colleagues reported a unique function of miR-126 in breast cancer metastasis.28 Although the precise mechanism(s) determining asymmetric miRNA biogenesis from a single miRNA precursor have not yet been defined, the miR-126 precursor produces comparable amounts of 5p and 3p miRNA species. They showed that the two mature miRNAs derived from pre-miR-126, miR-126 and miR-126*, cooperatively targeted expression of stromal cell-derived factor-1 alpha (SDF-1α)/ chemokine (C-X-C motif) ligand 12 (CXCL12), and subsequently chemokine (C-C motif) ligand 2 (CCL2), in cancer cells. Through these inhibitory effects, they suppressed recruitment of mesenchymal stem cells and inflammatory monocytes into tumor stroma, ultimately suppressing lung metastasis by breast cancer cells. In their report, miR-126/-126* appeared to exert metastasis-suppressive activities predominantly in primary tumor site, rather than metastatic nodules.28
Regulation of tumor cell immunophenotype by miR-135b
Although the roles of miRNAs in tumor microenvironments have emerged in recent reports, especially about solid tumors,8, 9, 10, 11 our group discovered a unique environmental involvement of miRNAs in hematological malignancies, where miRNA alteration accounts for regulation of immunophenotype of cancer cells per se and modulation of tumor microenvironments (Figure 2c).29 We showed that miR-135b, overexpressed in various types of cancer, including colon and lung cancers,30,31 was highly expressed in nucleophosmin-anaplastic lymphoma kinase (NPM-ALK)-positive anaplastic large cell lymphomas (ALCLs) and mediated downstream signaling of the NPM-ALK-STAT3 (signal transducer and activator of transcription factor 3) pathway.29 In this tumor type, NPM-ALK oncogene strongly promotes the expression of LEMD1, a host gene of miR-135b, and that of miR-135b, through activation of STAT3. FOXO1 was identified as a target of miR-135b, suggesting contribution to the oncogenic activities of NPM-ALK.
In parallel, we demonstrated immunomodulatory properties of miR-135b. Interestingly, miR-135b conferred interleukin (IL)-17-producing immunophenotype, which has recently been demonstrated by genome-wide expression profiling of various peripheral T-cell lymphomas, on ALCL cells.29 This skewing of ALCL immunophenotype, overlapping with that of T-helper (Th) 17 cells, was associated with targeting of Th2 master regulators STAT6 and GATA3 by miR-135b, indicating that miR-135b contributes to generation of an IL-17-producing immunophenotype by perturbing mutually antagonistic differentiation programs active during normal lymphocyte differentiation.32,33 miR-135b suppression inhibited expression levels of IL-17A, IL-17F, IκBζ, IL-6 and IL-8 in ALCL cells. miR-135b silencing also attenuated the expression of granzyme B and perforin 1, cytotoxic molecules highly expressed in ALCLs, implying that miR-135b exerts a broad range of effects on the ALCL immunophenotype. In line with the inflammatory role of IL-17, miR-135b blockade attenuated the paracrine inflammatory response in co-cultures of ALCL cells and fibroblasts and reduced tumor angiogenesis and in vivo growth. Although the mechanisms underlying lymphoma immunophenotypes related to the corresponding normal lymphocytes are largely unclear, the study illuminated the unique contribution made by an oncogenic kinase-linked miRNA to the modulation of tumor immune phenotype.
Distal impacts of miR-29 on angiogenesis and ECM remodeling
An association between miRNAs and GATA3 transcriptional factor has also been described in the setting of breast cancer development.34 In the mammary gland, GATA3 is required for luminal epithelial cell differentiation and maintenance, and its expression progressively decreases during progression of luminal breast cancer in association with a worse prognosis. A recent report demonstrated that GATA3 promoted differentiation of breast cancer cells, inhibited metastasis and modified the tumor microenvironment, through induction of miR-29b (Figure 2d).34 Thus the GATA3–miR29b axis functions as a tumor-suppressive arm, and loss of miR-29b in cells expressing GATA3 promotes a mesenchymal phenotype and metastasis. As mechanistic insights, various targets of miR-29b, including VEGF-A, ANGPTL4, platelet-derived growth factor, LOX and matrix metalloproteinase 9 (MMP9), which are involved in angiogenesis, collagen remodeling and proteolysis to promote metastasis, were identified. miR-29b introduction in a murine orthotopic breast cancer model reduced blood vessel development and the extent of fibrillar collagen synthesis without a concomitant effect on primary tumor size and reduced the incidence and size of metastasis. This metastasis inhibitory effect of miR-29b was mitigated by re-introduction of the miR-29b regulons VEGF-A, ANGPTL4, LOX and MMP9, strengthening the importance of microenvironmental target regulation by miR-29b.34 This study revealed a pleiotropic role for miR-29b in the modulation of the tumor microenvironment; both angiogenesis and ECM remodeling were affected. As miR-29b was also suppressed in NPM-ALK-positive ALCLs, in which GATA3 is downregulated,35 a GATA3–miR-29b axis may be operational in a range of cancer types. Additionally, in nasopharyngeal carcinomas, miR-29c downregulation has been reported to induce the expression of ECM proteins, including COL1A2, COL3A1, COL4A1 and laminin γ1, as well.36
Distal modulation of tumor–immune crosstalk by miRNAs
An important aspect of the evolution of tumor microenvironments is evasion of anti-tumor immune responses.3 Recent work has suggested that such immune escape of tumors can be broadly classified into two categories, depending on the characteristics of the tumor microenvironment.37 One major type exhibits a T-cell-inflamed phenotype with infiltration of T cells and a broad chemokine profile. Such tumors appear to endure immune attack predominantly by engaging inhibitory effectors of the immune system, such as PD-L1, IDO and regulatory T (Treg) cells. The other major type lacks this T-cell-inflamed phenotype and appears to prevent immunological attack through immune system ignorance and exclusion. In tumors in which the latter mechanism is operative, dense stroma and accumulation of immunosuppressive myeloid or macrophage populations might be observed, instead of T-cell infiltration. miRNA dysregulation in cancer cells appears to contribute to both types of immune escape. Regulation of SDF-1α by miR-126/126* and modulation of tumor stroma reactions by miR-29b may be associated with the latter category of escape.28,34
Recent analyses have shed light on the contributions made by miRNAs to activation of the inhibitory arm of the immune system in the former category of escape. In human melanoma, high expression levels of miR-30b and miR-30d were associated with frequent metastasis, early recurrence and lower overall survival (Figure 2e).38 Functional analyses revealed that miR-30b/-30d directly targeted the GalNAc transferase GALNT7 and subsequently promoted the secretion of the immunosuppressive cytokine IL-10.38 This reduced immune cell activation and enhanced recruitment of Treg cells, promoting metastasis.
In addition, a study in hepatocellular carcinoma also revealed enhancement of Treg cell function by a miRNA-mediated non-cell-autonomous mechanism (Figure 2f).39 In hepatocellular carcinoma, portal vein tumor thrombus (PVTT) is associated with a poor prognosis. Persistent presence of heptitis B virus (HBV) in liver tissue enhances transforming growth factor (TGF)-β activity, and in turn, TGF-β suppresses miR-34a.39 Although miR-34a is well known to be a transcriptional target of p53, and a representative tumor-suppressive miRNA,15 miR-34a had no significant effect on cell proliferation but suppressed the production of CCL22 important for Treg cell recruitment. An inverse correlation between the expression level of miR-34a and CCL22 or FoxP3 was observed in HBV+ primary tumor and PVTT samples. Restoration of miR-34a inhibited the growth of murine liver tumor cells, infiltration of Treg cells and metastasis, in immune-competent mice.39 Taken together, the data showed active participation of miRNAs in immune escape and also revealed one of the mechanisms of immune suppression by TGF-β, which is abundantly expressed in tumor stroma and has a versatile role in tumor development.40
Furthermore, other reports have also revealed the involvement of multiple miRNAs in immune responses. In glioma, miR-124 downregulation is associated with immunosuppressive activities of glioma stem cells and suppression of the effector response of T cells.41 miR-17-5p and miR-20 in miR-17-92 polycistronic cluster were shown to suppress cell migration and invasion by altering the secretion levels of IL-8, CK8 and CXCL1 from breast cancer cells.42 In head and neck cancer, the tumor-suppressive miR-145 has been reported to target SOX9 and ADAM17 and subsequently suppress IL-6 production.43
Convergent modulation of environmental regulators by multiple miRNAs
Independent miRNAs potentially target a range of genes exerting various functions, including both tumor suppressors and promoters. The sets of miRNA regulons mentioned above serve as examples that the targets divergently biased for tumor promotion or suppression. On the other hand, another scenario has also been described: one set of miRNAs convergently target metastasis-associated genes.44 In melanoma, a set of pro-metastatic miRNAs, including miR-199a-5p, miR-199a-3p and miR-1908, has been shown to combinatorially target apolipoprotein E (ApoE) (Figure 2g).44 Secretion of ApoE from cancer cells suppresses cancer cell invasion and endothelial cell recruitment, thereby inhibiting metastasis and angiogenesis. Regulation of SDF-1α by miR-126/126* is also categorized as such a type of regulation.28 When independent miRNAs lack the power to drive significant biological effects, a combination of alterations in multiple miRNAs may modulate the threshold to achieve metastasis.
Proximal roles of miRNAS in tumor stromal cells
Various cell types, including fibroblasts, endothelial cells, pericytes and immune inflammatory cells, contribute to the formation of tumor microenvironments favorable for cancer growth. Of these, CAFs, in contrast to normal fibroblasts, enhance ECM production and secrete cancer-activating cytokines and chemokines, significantly promoting tumorigenesis.3,45, 46, 47, 48 It remains largely unclear how CAFs arise from normal fibroblasts. In certain cancer types, CAFs are thought to be generated from normal fibroblasts by tumor-derived paracrine signals.45,46 Nonetheless, it has also been known that the tumor-supporting capacity of CAFs is sustainable over multiple passages in the absence of cancer cells.45 Although early studies suggested the presence of genetic abnormalities in tumor-suppressors p53 and phophatase and tensin homolog (PTEN) in CAFs,49 detailed genetic analyses have indicated that genetic alterations are in fact rare,50,51 suggesting that other epigenetic mechanisms stabilize the CAF phenotype. In this context, recent studies have revealed that the miRNA profiles of normal fibroblasts and CAFs differ and that several features of the CAF phenotype are attributable to miRNA dysregulation.9 In addition, miRNA function has been linked to variations in other tumor-associated cell types, including osteoclasts, myeloid-derived suppressor cells (MDSCs), and tumor-associated macrophages (TAMs). We next focus on the dysregulation of miRNAs in stromal cells and the proximal effects of such changes in tumor microenvironments (Figure 3).

Proximal functions of miRNAs in tumor stromal cells. Specific changes of multiple miRNAs in CAFs (a), osteoclasts (b) and MDSCs (c) induce phenotypic changes of these cell types, leading to tumor progression. Oncogenic miRNAs and tumor-suppressive miRNAs are represented by black and red, respectively.
Reprogramming of CAFs by miR-31, miR-214, and miR-155
Mitra et al.52 recently conducted miRNA expression profiling of primary CAFs vs adjacent normal fibroblasts in ovarian cancer patients, and induced CAFs (iCAFs), generated from normal fibroblasts upon co-culture with tumor cells, vs normal fibroblasts (Figure 3a). This study showed downregulation of miR-214 and miR-31 and upregulation of miR-155 in CAFs. Interestingly, introduction of miR-214 and miR-31 and silencing of miR-155 converted the CAF phenotype to normality, and vice versa, suggesting a direct involvement of miRNAs in reversible conversion of normal fibroblasts into CAFs. Patient CAFs, iCAFs and miRNA-reprogrammed CAFs (miR-CAFs) all exhibited high expression levels of several pro-tumorigenic chemokines, including CCL5, CCL20 and CXCL8/IL-8. CCL5 was shown to be a direct target of miR-214. Moreover, patient CAFs and miR-CAFs enhanced the growth of co-injected ovarian cancer cells and increased invasion by co-cultured cancer cells; these effects were abrogated upon neutralization of CCL5, indicating that CCL5 is a key tumor modulator in miR-CAFs.52 The results indicated that ovarian cancer cells reprogram fibroblasts to CAFs via the action of miRNAs. In addition, downregulation of miR-31 has been reported in CAFs isolated from endometrial cancers, and miR-31 downregulation upregulated the target SATB2 (special AT-rich sequence-binding protein 2), enhancing tumor cell migration.53 Downregulation of miR-148a in CAFs of endometrial cancer has also been reported.54
Proximal roles of PTEN-miR-320 axis in CAF
CAFs frequently show characteristic upregulation of CAF-related genes, such as α-smooth muscle actin, fibroblast-specific protein, platelet-derived growth factor-B and fibroblast activation protein.47,55 In addition, alterations in the expression levels of several other markers, including periostin, p53, PTEN and podoplanin, have been reported to correlate with the prognostic outcomes of solid tumors.55, 56, 57 Accordingly, abrogation of PTEN in stromal cells has been shown to promote the development of epithelial mammary tumors.57 A recent study found that a specific miRNA was responsible for the tumor-suppressive activity of PTEN (Figure 3a).58 Of several miRNAs exhibiting distinct expression patterns in PTEN-depleted fibroblasts, miR-320 was identified as a tumor suppressor acting downstream of PTEN. miR-320 targets ETS2 (v-ets erythroblastosis virus E26 oncogene homolog 2), MMP9 and Emilin2, and downregulation thereof induces a tumor-specific secretome, composed of MMP9, MMP2, BMP1 and LOXL2, leading to enhanced angiogenesis and tumor cell invasion. Expression of the miR-320-Ets2-related secretory regulons separated human normal breast stroma from tumor stroma and correlated with the outcomes of breast cancer patients.58
Co-downregulation of miR-15a and miR-16-1 in tumor cells and CAFs
Another report demonstrated that certain miRNAs exhibit a pan-tumor-suppressive function in both cancer cells and CAFs.59 miR-15a and miR-16-1 are well-characterized tumor-suppressive miRNAs encoded in the chromosomal region 13q14 and are frequently lost in chronic lymphocytic leukemia and prostate cancer.60,61 miR-15a and miR-16-1 function as tumor suppressors in prostate cancer by targeting BCL2, cyclin D1 and Wnt3a.61 Musumeci et al.59 reported that miR-15 and miR-16 were also downregulated in the CAFs of most of prostate tumor patients (Figure 3a). Restoration of miR-15a and miR-16-1 suppressed proliferation and migration of CAFs as similarly observed in cancer cells. Furthermore, miR-15a and miR-16 suppressed two novel targets, fibroblast growth factor 2 (FGF2) and its receptor FGFR1, which enhance the proliferation and migration of both stromal and cancer cells. Reconstitution of miR-15 and miR-16 suppressed the tumor-promoting activity of stromal cells in vivo.59 These findings suggest that some miRNAs can function as pan-tumor suppressors in cancer cells and tumor microenvironments, although the mechanisms of deregulation may be distinct between cancer cells and microenvironments. From the standpoint of therapeutic intervention, simple restoration of this type of miRNA might aid co-targeting cancer cells and tumor microenvironment.
miRNA-mediated osteoclast differentiation in bone metastasis
Osteolytic bone metastasis is frequently observed in many cancer types, including breast and lung cancers, and is associated with aberrant osteoclast activation.48,62,63 In a recent study, treatment of preosteoclast cells with conditioned media from highly bone metastatic breast and bladder cancer cell lines was shown to induce characteristic changes in miRNA expression, similar to those noted upon stimulation with RANKL (receptor activator of nuclear factor κB ligand), a major stimulator of osteoclast differentiation (Figure 3b).64 Bone-metastatic cancer cells triggered such miRNA expression changes partly through soluble intercellular adhesion molecule 1 (sICAM1)-mediated NF-κB activation. Overexpression of multiple miRNAs that were downregulated during osteoclastogenesis suppressed various osteoclast genes, including Calcr (encoding the calcitonin receptor) and TRAF6, and inhibited osteoclast differentiation. Intravenous delivery of these miRNAs such as miR-141 and miR-219 reduced osteolytic bone metastasis. Interestingly, the serum levels of two miRNAs miR-16 and miR-378, which increased during osteoclastogenesis, were associated with bone metastasis, suggesting that the miRNAs might serve as useful biomarkers.64 These findings emphasize the key roles played by miRNAs in aberrant osteoclastogenesis during bone metastasis.
Proximal functions of miRNAs in MDSCs
In tumor-infiltrating immune cells, MDSCs have key roles in immune suppression, in turn allowing tumor progression.48,62,65, 66, 67, 68 MDSCs are myeloid-related cells characterized by the expression of Gr-1 and CD11b markers and the ability to suppress T lymphocyte activation. MDSCs are thought to arise upon deviation of immature myeloid progenitor cells continually generated in bone marrow from the normal differentiation programs toward macrophages, dendritic cells and granulocytes.67,68 MDSC numbers increase in the bone marrow and blood of tumor-bearing mice and cancer patients and accumulate in tumor microenvironments.62,65 MDSCs are heterogenous in nature and are mainly divided into Ly6Glow/Ly6Chigh monocytic MDSCs and Ly6Ghigh/Ly6Clow granulocytic or polymorphonuclear MDSCs.67,69,70 These cells suppress the anti-tumor functions of CD4+ and CD8+ T cells via production of arginase, nitric oxide (NO) and reactive oxygen species (ROS), expand the pool of Treg cells and inhibit the activities of natural killer cells.68 In addition, MDSCs promote tumor progression by enhancing MMP9 production in an immune-independent manner.
Liu et al.71 recently performed miRNA profiling of Gr1+CD11b+MDSCs in mice bearing syngeneic 4T1 mammary tumors and identified miR-494 as the most prominently upregulated miRNA in tumor-expanded MDSCs (Figure 3c). Induction of miR-494 was observed in both monocytic and granulocytic MDSCs and was further confirmed in five other tumor models, including lung cancer, melanoma, lymphoma and colon cancer. Among tumor-derived factors, including IL-6, GM-CSF, and TGF-β1, TGF-β1 was responsible for upregulation of miR-494 in MDSCs. In turn, miR-494 inhibited PTEN, increased Akt activity and supported MDSC survival. Moreover, miR-494 enhanced expression of arginase, required for immunosuppression, and that of multiple MMPs, such as MMP2, MMP13 and MMP14. Importantly, silencing of miR-494 reversed the immune-suppressive capacity of MDSCs and inhibited tumor growth and metastasis, suggesting that miR-494 played a key role in expansion and maintenance of tumor-associated MDSCs. In addition, Li et al.72 reported that miR-155 and miR-21 were highly induced during induction of MDSC by GM-CSF and IL-6 (Figure 3c). High-level expression of miR-155 and miR-21 was confirmed in tumor-bearing mice and was shown to promote the production of both monocytic and granulocytic MDSCs. TGF-β also promoted their expression to expand MDSCs. Mechanistically, miR-155 and miR-21 target SHIP-1 and PTEN, respectively, and finally activate the STAT3 signaling pathway crucial for MDSC function.72 It should be further investigated whether these findings are applicable to human MDSCs.
In MDSCs, STAT1 and STAT3 transcription factors are important for the production of NO and ROS, respectively, and several effects of MDSCs are mediated by the Janus-activated kinase 2–STAT3 axis, which is stimulated by tumor-derived factors.68,69,73 Two miRNAs, miR-17-5p and miR-20a, were previously shown to suppress STAT3 in MDSCs.74 These miRNAs decreased under coexistence with tumors. Both miR-17-5p and miR-20a reduced production of ROS and H2O2 and alleviated the suppressive function of granulocytic MDSCs on antigen-specific CD4+ and CD8+ T cells.74 These findings thus suggest that miR-17-5p and miR-20a may be additional druggable targets to modulate MDSC function.
Autoregulation of TAMs by miR-511-3p
TAMs are another important type of bone marrow-derived cells in tumor microenvironments.48,62,68 Although heterogeneous in nature, TAMs are thought to share characteristics with alternatively activated M2 macrophages favorably skewed by IL-4, IL-10, IL-13, glucocorticoids and TGF-β. In contrast to classically activated M1 macrophages, which support tumor rejection, TAMs are proposed to produce a range of tissue-remodeling and immunomodulatory factors, including VEGF-A, FGF2, CXCL8, hepatocyte growth factor and MMPs, and to enhance tumor invasion, ECM synthesis, angiogenesis and immune suppression.62,68,75 Although the expression patterns and roles of miRNAs in TAMs remain largely unknown, a recent report showed that miR-511-3p exerted regulatory functions in TAMs.76 In mice, TAMs are characterized by high-level expression of the mannose receptor (MRC1/CD206). miR-511-3p, encoded within the intron of MRC1, is abundantly expressed in MRC1+ alternatively activated macrophages and TAMs. Overexpression of miR-511-3p in TAMs suppressed pro-tumoral genes and inhibited tumor growth in vivo, suggesting that miR-511-3p rather limits the pro-tumoral function of TAMs and forms a negative feedback loop together with MRC1.76 Modulation of miR-511-3p may be beneficial to switch TAMs from pro- to anti-tumoral phenotype. In addition, miR-155 has been reported to modulate TAM phenotypes.77
Reciprocal modulation of miRNA networks in tumor microenvironments: roles of extracellular vesicles
As described above, alteration of miRNA expression patterns in cancer and stromal cells can exert distal and proximal effects on the tumor stroma, respectively. In addition, tumor stromal cells appear to modulate the miRNAs of cancer cells conversely (Figure 4a). In ovarian cancer, MDSCs enhance the stem-cell-like properties of cancer cells via induction of miR-101 in co-cultured cancer cells.78 Other studies have found that TAMs increase the expression levels of CD44 and Bmi1 by suppressing miR-328 and miR-30e*, respectively, in gastrointestinal cancer.79,80 Collagen remodeling has also been reported to be associated with suppression of the tumor-suppressive let-7 miRNA in pancreatic ductal adenocarcinomas.81 These results collectively suggest reciprocal modulation of the miRNA networks of cancer cells and tumor stroma.

Reciprocal modulation of miRNA networks in tumor microenvironment. (a) Stromal cells modulate miRNAs in tumor cells. (b) Cancer cells directly transferred miRNAs to stromal cells, and vice versa.
Moreover, recent work has highlighted more direct roles played by miRNAs in cellular communication. Currently, miRNAs are shown to be present in various forms in extracellular fluids.82, 83, 84, 85, 86 These secretory miRNAs are contained in exosomes, microvesicles, apoptotic bodies and high-density lipoproteins or bound to RNA-binding proteins.82, 83, 84, 85, 86 Vesicles released from cells, including exosomes, microvesicles and apoptotic bodies, vary from 30 nm to 5 μm in diameter and are collectively called extracellular vesicles.87,88 In particular, exosomes, smallest membrane vesicles (30–100 nm in size), have been intensely studied as direct conveyors of miRNAs mediating cellular communication.89 Although a relative extent of effects of secretory miRNAs on intracellular miRNA networks remains unclear, recent studies showed that such extracellular miRNAs, especially those embedded in extracellular vesicles, were transferred to recipient cells and mediated gene regulation.89,90 In addition, miRNAs in extracellular fluids hold a promising route to develop biomarkers.
In tumor microenvironments, extracellular miRNAs have been suggested to influence tumor progression via bidirectional tumor-to-stromal and stromal-to-tumor communication (Figure 4b). Kosaka et al.91 showed that neutral sphyngomyelinase 2 (nSMase2), which regulates ceramide biogenesis, is involved in exosomal miRNA secretion. The nSMase2 enzyme level was enhanced in cancer cells, and nSMase2 promoted tumor angiogenesis and metastasis in a breast cancer model.89,91 Endothelial activation required nSMase2 within tumor cells and was enhanced by exosomes derived from metastatic cancer cells. Furthermore, they showed that miR-210 secreted from cancer cells served as an angiogenic miRNA in recipient endothelial cells (Figure 4b).91 Although the evidence is scanty, the possibility of intercellular communication in the opposite direction has also been proposed.92 miR-223 was reported to transit from IL-4-activated macrophages to breast cancer cells, via exosomes, and to function in co-cultured cancer cells. In contrast, miR-223 was reported to be transferred from macrophages to hepatocellular carcinoma cells, depending on cellular contact and gap junctions, but not exosomes,93 suggesting that the transfer modes of extracellular miRNAs may be complex. In addition, miR-1, which is downregulated in human glioblasoma multiforme, was recently shown to mediate extracellular vesicle function.94 Overexpression of miR-1 in glioblastoma cells attenuated in vivo tumor growth, angiogenesis and invasiveness. Such effects were associated with extracellular vesicle transport of miR-1. miR-1 was shown to suppress Annexin A2, one of the most abundant proteins in glioblastoma-derived extracellular vesicles, and other proteins and was proposed to modulate the functions of extracellular vesicles per se.94 Further investigations will yield valuable insights into the roles played by extracellular miRNAs in tumor microenvironments.
Environmental factors and bias influencing miRNA expression in tumor microenvironments
Numerous reports have described alterations in the miRNA profiles of many types of cancer. Such alterations in human cancers are induced by dysregulation of specific signaling pathways in independent cancer types but may also be consequences of more general events, such as hypoxia and acidity, arising in tumor microenvironments.95 Multiple miRNAs, including miR-210, miR-155, miR-372/373 and miR-200b, have been reported to exhibit dynamic changes in the expression levels in response to hypoxia with variable results in literature.12,96, 97, 98, 99, 100 Among them, miR-210 has been suggested to be a key regulator of hypoxia in human cancers.97,101,102 miR-210 is consistently induced at the transcriptional level by hypoxia-inducible factors and, in turn, regulates multiple normoxic genes and tumor-associated pathways. Although miR-210 upregulation is frequently observed in various types of cancer, reports on the function of miR-210 in tumorigenesis vary in terms of the conclusions reached.102,103 miR-210 has also been reported to target ephrin A3 (EFNA3) and to promote angiogenesis, suggesting that miR-210 has various roles in hypoxic tumor and stromal cells. In addition, a recent report found that hypoxia influenced the extent of crosstalk between miRNAs and RNA-binding proteins.104 In monocytic cells, endogenously expressed miR-297 and miR-299 suppressed VEGF-A expression in normoxia by binding to CA-rich element in the VEGF-A 3′-untranslated region. Hypoxia induces cytoplasmic translocation of nuclear ribonucleoprotein L, which also binds the CA-rich element, and attenuates VEGF-A suppression by compromising miRNA functions.104 Thus the tumor microenvironment may modulate not only miRNA expression but also miRNA function, by regulating the actions of miRNA modulators.
On the other hand, alterations in the miRNA profiles of whole tumor tissues should be carefully interpreted, because the consequences of changes in cancer and stromal cells will be influenced by the stromal contents. This is because specific miRNA alterations can be induced in both cancer and stromal cells, as described above. Although cell-type-specific miRNA changes within tumor tissues remain poorly understood, some studies have indeed shown confinement of altered miRNA expression to distinct cellular compartments of various solid tumors. Sempere et al.105,106,107 reported that alterations in miRNA expression patterns were confined to a specific epithelial subpopulation of breast cancer and that such changes depended on cell types and tumor types, as revealed by combining data from in situ hybridization and immunohistochemical assays. They reported that high-level expression of miR-21, frequently observed in various hematological and solid tumors, was observed preferentially in cancer cells of lung, prostate and pancreatic cancers but in the CAFs of breast and colorectal cancers.106 In contrast, expression of miR-155, often upregulated in various cancers, was predominantly restricted to immune-related cells.106 In addition, predominant overexpression of miR-21 in CAFs was reported in esophageal squamous cell carcinoma.108 Thus, global profiling results performed on entire tumor digests should be interpreted carefully together with further innovation to understand context-dependent roles of miRNA in tumor microenvironments. Although multiple genes can be potentially targeted by each miRNA, the roles played by miRNAs in the pathogenesis have been suggested to be highly context dependent.109 Several recent integration frameworks, including GenMiR++, FAME (functional assignment of miRNAs via enrichment), CoSMic (context-specific miRNA analysis) and GFA (GSEA-FAME analysis), which integrate miRNA target-prediction information and biological expression data, may advance our understanding of the context-dependent roles of miRNAs.35,110, 111, 112, 113, 114
Targeting of tumor microenvironments by miRNAS
Although miRNA research on tumor microenvironments has extended the list of miRNAs that may serve as therapeutic targets, several miRNAs have been identified as direct modulators of these microenvironments besides mediators of crosstalk between cancer cells and microenvironments. miR-125b was upregulated upon stimulation with VEGF-A and ischemia and targeted VE-cadherin expression in endothelial cells.115 Systemic administration of miR-125b induced formation of non-functional blood vessels and inhibited in vivo tumor growth, suggestive of therapeutic potential. On the other hand, miR-155 has been shown to be an immunostimulatory miRNA. Selective loading of miR-155 into tumor-infiltrating dendritic cells using polyethylenimine-based nanocomplexes converted the phenotype from immunosuppressive to immunostimulatory and triggered anti-tumor immunity in ovarian cancer models.116 In addition, systemic injection of immunostimulatory miR-124, which is absent in a range of gliomas, and adoptive transfers of miR-124-introduced T cells were shown to exhibit anti-tumor activities in a T-cell-dependent manner.41 These findings indicate that components of tumor microenvironments can be directly targeted by miRNA-mediated cancer therapeutics.
Conclusion
In this review, we have summarized recent advances in the roles played by miRNAs in tumor microenvironments. These advances have unveiled both the indirect and distal impacts of miRNAs on the tumor stroma via non-cell-autonomous regulation of cancer cell function and the direct and proximal roles of miRNAs in tumor stromal cells.
In the former part (Figure 2), oncogenic miR-9 and tumor-suppressive miR-126 regulate angiogenesis via their regulons (miR-9—E-cadherin; miR-126—MERTK, IGFBP2 and PITPNC1), and tumor-suppressive miR-29b and the regulons (VEGF-A, ANGPTL4, LOX and MMP9) modulate ECM remodeling and angiogenesis in breast cancer, finally leading to global regulation of tumor metastasis. In NPM-ALK-positive ALCL, oncogenic miR-135b empowers lymphoma cells to stimulate paracrine inflammatory reactions by controlling STAT6 and GATA3. In melanoma, oncogenic miR-30b/d and a tumor-suppressive miRNA group (miR-199a-5p/3p and miR-1908) control immunosuppression and angiogenesis through regulation of GALNT7 and ApoE, respectively. In addition, tumor-suppressive miR-34a links TGF-β signaling to CCL22-mediated immunosuppression in hepatocellular carcinoma. These non-cell-autonomous effects embrace many aspects of the tumor microenvironment, including tumor angiogenesis, tumor immune invasion and tumor–stromal interactions. The latter part (Figure 3) highlighted that tumor progression is supported by alterations of multiple miRNAs in CAFs (miR-214, -31, -155, -320, -15a and -16-1) in various cancer types, including ovarian and prostate cancers, and that miRNAs also contribute to aberrant functions of other cell types, including osteoclasts, MDSCs and TAMs. These findings collectively revealed that miRNAs have widespread impacts on environmental changes in tumor, which are initiated by dysfunction in gene regulatory networks. Furthermore, the studies reviewed demonstrated that these miRNAs and/or regulons in both cancer and stromal cells can be not only clinical biomarkers but also potential therapeutic targets to target tumor microenvionments.
Future studies will shed light on how such alterations are spatiotemporally induced and how miRNA abnormalities affect therapeutic responses and resistance. In addition, recent studies have highlighted miRNA-mediated reciprocal interactions between cancer and stomal cells and the importance of extracellular miRNAs in such cellular communication (Figure 4). Further investigations will bring deep insight for distinct involvement of various forms of miRNAs in extracellular fluids. This will enhance our understanding of the in vivo kinetics of miRNA-like molecules and allow development of therapeutic approaches using miRNA-like agents. In summary, targeting of miRNA-mediated gene networks in various tumor compartments may yield novel solutions to treat cancer tissues and achieve better therapeutic responses.
References
Hanahan D, Weinberg RA . The hallmarks of cancer. Cell 2000; 100: 57–70.
Hanahan D, Weinberg RA . Hallmarks of cancer: the next generation. Cell 2011; 144: 646–674.
Junttila MR, de Sauvage FJ . Influence of tumour micro-environment heterogeneity on therapeutic response. Nature 2013; 501: 346–354.
Heldin CH, Rubin K, Pietras K, Ostman A . High interstitial fluid pressure—an obstacle in cancer therapy. Nat Rev Cancer 2004; 4: 806–813.
He L, Hannon GJ . MicroRNAs: small RNAs with a big role in gene regulation. Nat Rev Genet 2004; 5: 522–531.
Esquela-Kerscher A, Slack FJ . Oncomirs—microRNAs with a role in cancer. Nat Rev Cancer 2006; 6: 259–269.
Iorio MV, Croce CM . MicroRNA dysregulation in cancer: diagnostics, monitoring and therapeutics. A comprehensive review. EMBO Mol Med 2012; 4: 143–159.
Nicoloso MS, Spizzo R, Shimizu M, Rossi S, Calin GA . MicroRNAs—the micro steering wheel of tumour metastases. Nat Rev Cancer 2009; 9: 293–302.
Li X, Wu Z, Fu X, Han W . A microRNA component of the neoplastic microenvironment: microregulators with far-reaching impact. Biomed Res Int 2013; 2013: 762183.
Pencheva N, Tavazoie SF . Control of metastatic progression by microRNA regulatory networks. Nat Cell Biol 2013; 15: 546–554.
Zhang Y, Yang P, Wang XF . Microenvironmental regulation of cancer metastasis by miRNAs. Trends Cell Biol 2014; 24: 153–160.
Wentz-Hunter KK, Potashkin JA . The role of miRNAs as key regulators in the neoplastic microenvironment. Mol Biol Int 2011; 2011: 839872.
Suzuki HI, Miyazono K . Emerging complexity of microRNA generation cascades. J Biochem 2011; 149: 15–25.
Croce CM . Causes and consequences of microRNA dysregulation in cancer. Nat Rev Genet 2009; 10: 704–714.
Suzuki HI, Miyazono K . Dynamics of microRNA biogenesis: crosstalk between p53 network and microRNA processing pathway. J Mol Med (Berl) 2010; 88: 1085–1094.
Suzuki HI, Yamagata K, Sugimoto K, Iwamoto T, Kato S, Miyazono K . Modulation of microRNA processing by p53. Nature 2009; 460: 529–533.
Suzuki HI, Arase M, Matsuyama H, Choi YL, Ueno T, Mano H et al. MCPIP1 ribonuclease antagonizes dicer and terminates microRNA biogenesis through precursor microRNA degradation. Mol Cell 2011; 44: 424–436.
Suzuki HI, Miyazono K . Control of microRNA maturation by p53 tumor suppressor and MCPIP1 ribonuclease. Enzymes 2012; 32: 163–183.
Mori M, Triboulet R, Mohseni M, Schlegelmilch K, Shrestha K, Camargo FD et al. Hippo signaling regulates microprocessor and links cell-density-dependent miRNA biogenesis to cancer. Cell 2014; 156: 893–906.
Pylayeva-Gupta Y, Lee KE, Hajdu CH, Miller G, Bar-Sagi D . Oncogenic Kras-induced GM-CSF production promotes the development of pancreatic neoplasia. Cancer Cell 2012; 21: 836–847.
Bayne LJ, Beatty GL, Jhala N, Clark CE, Rhim AD, Stanger BZ et al. Tumor-derived granulocyte-macrophage colony-stimulating factor regulates myeloid inflammation and T cell immunity in pancreatic cancer. Cancer Cell 2012; 21: 822–835.
Ceppi P, Peter ME . MicroRNAs regulate both epithelial-to-mesenchymal transition and cancer stem cells. Oncogene 2014; 33: 269–278.
Ma L, Young J, Prabhala H, Pan E, Mestdagh P, Muth D et al. miR-9, a MYC/MYCN-activated microRNA, regulates E-cadherin and cancer metastasis. Nat Cell Biol 2010; 12: 247–256.
Tavazoie SF, Alarcon C, Oskarsson T, Padua D, Wang Q, Bos PD et al. Endogenous human microRNAs that suppress breast cancer metastasis. Nature 2008; 451: 147–152.
Feng R, Chen X, Yu Y, Su L, Yu B, Li J et al. miR-126 functions as a tumour suppressor in human gastric cancer. Cancer Lett 2010; 298: 50–63.
Guo C, Sah JF, Beard L, Willson JK, Markowitz SD, Guda K . The noncoding RNA, miR-126, suppresses the growth of neoplastic cells by targeting phosphatidylinositol 3-kinase signaling and is frequently lost in colon cancers. Genes Chromosomes Cancer 2008; 47: 939–946.
Png KJ, Halberg N, Yoshida M, Tavazoie SF . A microRNA regulon that mediates endothelial recruitment and metastasis by cancer cells. Nature 2012; 481: 190–194.
Zhang Y, Yang P, Sun T, Li D, Xu X, Rui Y et al. miR-126 and miR-126* repress recruitment of mesenchymal stem cells and inflammatory monocytes to inhibit breast cancer metastasis. Nat Cell Biol 2013; 15: 284–294.
Matsuyama H, Suzuki HI, Nishimori H, Noguchi M, Yao T, Komatsu N et al. miR-135b mediates NPM-ALK-driven oncogenicity and renders IL-17-producing immunophenotype to anaplastic large cell lymphoma. Blood 2011; 118: 6881–6892.
Sarver AL, French AJ, Borralho PM, Thayanithy V, Oberg AL, Silverstein KA et al. Human colon cancer profiles show differential microRNA expression depending on mismatch repair status and are characteristic of undifferentiated proliferative states. BMC Cancer 2009; 9: 401.
Lin CW, Chang YL, Chang YC, Lin JC, Chen CC, Pan SH et al. MicroRNA-135b promotes lung cancer metastasis by regulating multiple targets in the Hippo pathway and LZTS1. Nat Commun 2013; 4: 1877.
Harrington LE, Hatton RD, Mangan PR, Turner H, Murphy TL, Murphy KM et al. Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nat Immunol 2005; 6: 1123–1132.
van Hamburg JP, Mus AM, de Bruijn MJ, de Vogel L, Boon L, Cornelissen F et al. GATA-3 protects against severe joint inflammation and bone erosion and reduces differentiation of Th17 cells during experimental arthritis. Arthritis Rheum 2009; 60: 750–759.
Chou J, Lin JH, Brenot A, Kim JW, Provot S, Werb Z . GATA3 suppresses metastasis and modulates the tumour microenvironment by regulating microRNA-29b expression. Nat Cell Biol 2013; 15: 201–213.
Suzuki HI, Matsuyama H, Noguchi M, Yao T, Komatsu N, Mano H et al. Computational dissection of distinct microRNA activity signatures associated with peripheral T cell lymphoma subtypes. Leukemia 2013; 27: 2107–2111.
Sengupta S, den Boon JA, Chen IH, Newton MA, Stanhope SA, Cheng YJ et al. MicroRNA 29c is down-regulated in nasopharyngeal carcinomas, up-regulating mRNAs encoding extracellular matrix proteins. Proc Natl Acad Sci USA 2008; 105: 5874–5878.
Gajewski TF, Schreiber H, Fu YX . Innate and adaptive immune cells in the tumor microenvironment. Nat Immunol 2013; 14: 1014–1022.
Gaziel-Sovran A, Segura MF, Di Micco R, Collins MK, Hanniford D, Vega-Saenz de Miera E et al. miR-30b/30d regulation of GalNAc transferases enhances invasion and immunosuppression during metastasis. Cancer Cell 2011; 20: 104–118.
Yang P, Li QJ, Feng Y, Zhang Y, Markowitz GJ, Ning S et al. TGF-beta-miR-34a-CCL22 signaling-induced Treg cell recruitment promotes venous metastases of HBV-positive hepatocellular carcinoma. Cancer Cell 2012; 22: 291–303.
Bierie B, Moses HL . Tumour microenvironment: TGFbeta: the molecular Jekyll and Hyde of cancer. Nat Rev Cancer 2006; 6: 506–520.
Wei J, Wang F, Kong LY, Xu S, Doucette T, Ferguson SD et al. miR-124 inhibits STAT3 signaling to enhance T cell-mediated immune clearance of glioma. Cancer Res 2013; 73: 3913–3926.
Yu Z, Willmarth NE, Zhou J, Katiyar S, Wang M, Liu Y et al. microRNA 17/20 inhibits cellular invasion and tumor metastasis in breast cancer by heterotypic signaling. Proc Natl Acad Sci USA 2010; 107: 8231–8236.
Yu CC, Tsai LL, Wang ML, Yu CH, Lo WL, Chang YC et al. miR145 targets the SOX9/ADAM17 axis to inhibit tumor-initiating cells and IL-6-mediated paracrine effects in head and neck cancer. Cancer Res 2013; 73: 3425–3440.
Pencheva N, Tran H, Buss C, Huh D, Drobnjak M, Busam K et al. Convergent multi-miRNA targeting of ApoE drives LRP1/LRP8-dependent melanoma metastasis and angiogenesis. Cell 2012; 151: 1068–1082.
Orimo A, Gupta PB, Sgroi DC, Arenzana-Seisdedos F, Delaunay T, Naeem R et al. Stromal fibroblasts present in invasive human breast carcinomas promote tumor growth and angiogenesis through elevated SDF-1/CXCL12 secretion. Cell 2005; 121: 335–348.
Erez N, Truitt M, Olson P, Arron ST, Hanahan D . Cancer-associated fibroblasts are activated in incipient neoplasia to orchestrate tumor-promoting inflammation in an NF-kappaB-dependent manner. Cancer Cell 2010; 17: 135–147.
Kalluri R, Zeisberg M . Fibroblasts in cancer. Nat Rev Cancer 2006; 6: 392–401.
Quail DF, Joyce JA . Microenvironmental regulation of tumor progression and metastasis. Nat Med 2013; 19: 1423–1437.
Kurose K, Gilley K, Matsumoto S, Watson PH, Zhou XP, Eng C . Frequent somatic mutations in PTEN and TP53 are mutually exclusive in the stroma of breast carcinomas. Nat Genet 2002; 32: 355–357.
Allinen M, Beroukhim R, Cai L, Brennan C, Lahti-Domenici J, Huang H et al. Molecular characterization of the tumor microenvironment in breast cancer. Cancer Cell 2004; 6: 17–32.
Qiu W, Hu M, Sridhar A, Opeskin K, Fox S, Shipitsin M et al. No evidence of clonal somatic genetic alterations in cancer-associated fibroblasts from human breast and ovarian carcinomas. Nat Genet 2008; 40: 650–655.
Mitra AK, Zillhardt M, Hua Y, Tiwari P, Murmann AE, Peter ME et al. MicroRNAs reprogram normal fibroblasts into cancer-associated fibroblasts in ovarian cancer. Cancer Discov 2012; 2: 1100–1108.
Aprelikova O, Yu X, Palla J, Wei BR, John S, Yi M et al. The role of miR-31 and its target gene SATB2 in cancer-associated fibroblasts. Cell Cycle 2010; 9: 4387–4398.
Aprelikova O, Palla J, Hibler B, Yu X, Greer YE, Yi M et al. Silencing of miR-148a in cancer-associated fibroblasts results in WNT10B-mediated stimulation of tumor cell motility. Oncogene 2013; 32: 3246–3253.
Cirri P, Chiarugi P . Cancer associated fibroblasts: the dark side of the coin. Am J Cancer Res 2011; 1: 482–497.
Hasebe T, Tamura N, Okada N, Hojo T, Akashi-Tanaka S, Shimizu C et al. p53 expression in tumor-stromal fibroblasts is closely associated with the nodal metastasis and outcome of patients with invasive ductal carcinoma who received neoadjuvant therapy. Hum Pathol 2010; 41: 262–270.
Trimboli AJ, Cantemir-Stone CZ, Li F, Wallace JA, Merchant A, Creasap N et al. Pten in stromal fibroblasts suppresses mammary epithelial tumours. Nature 2009; 461: 1084–1091.
Bronisz A, Godlewski J, Wallace JA, Merchant AS, Nowicki MO, Mathsyaraja H et al. Reprogramming of the tumour microenvironment by stromal PTEN-regulated miR-320. Nat Cell Biol 2012; 14: 159–167.
Musumeci M, Coppola V, Addario A, Patrizii M, Maugeri-Sacca M, Memeo L et al. Control of tumor and microenvironment cross-talk by miR-15a and miR-16 in prostate cancer. Oncogene 2011; 30: 4231–4242.
Calin GA, Ferracin M, Cimmino A, Di Leva G, Shimizu M, Wojcik SE et al. A microRNA signature associated with prognosis and progression in chronic lymphocytic leukemia. N Engl J Med 2005; 353: 1793–1801.
Bonci D, Coppola V, Musumeci M, Addario A, Giuffrida R, Memeo L et al. The miR-15a-miR-16-1 cluster controls prostate cancer by targeting multiple oncogenic activities. Nat Med 2008; 14: 1271–1277.
Joyce JA, Pollard JW . Microenvironmental regulation of metastasis. Nat Rev Cancer 2009; 9: 239–252.
Mundy GR . Metastasis to bone: causes, consequences and therapeutic opportunities. Nat Rev Cancer 2002; 2: 584–593.
Ell B, Mercatali L, Ibrahim T, Campbell N, Schwarzenbach H, Pantel K et al. Tumor-induced osteoclast miRNA changes as regulators and biomarkers of osteolytic bone metastasis. Cancer Cell 2013; 24: 542–556.
Ostrand-Rosenberg S . Immune surveillance: a balance between protumor and antitumor immunity. Curr Opin Genet Dev 2008; 18: 11–18.
Marigo I, Dolcetti L, Serafini P, Zanovello P, Bronte V . Tumor-induced tolerance and immune suppression by myeloid derived suppressor cells. Immunol Rev 2008; 222: 162–179.
Peranzoni E, Zilio S, Marigo I, Dolcetti L, Zanovello P, Mandruzzato S et al. Myeloid-derived suppressor cell heterogeneity and subset definition. Curr Opin Immunol 2010; 22: 238–244.
Gabrilovich DI, Ostrand-Rosenberg S, Bronte V . Coordinated regulation of myeloid cells by tumours. Nat Rev Immunol 2012; 12: 253–268.
Youn JI, Nagaraj S, Collazo M, Gabrilovich DI . Subsets of myeloid-derived suppressor cells in tumor-bearing mice. J Immunol 2008; 181: 5791–5802.
Movahedi K, Guilliams M, Van den Bossche J, Van den Bergh R, Gysemans C, Beschin A et al. Identification of discrete tumor-induced myeloid-derived suppressor cell subpopulations with distinct T cell-suppressive activity. Blood 2008; 111: 4233–4244.
Liu Y, Lai L, Chen Q, Song Y, Xu S, Ma F et al. MicroRNA-494 is required for the accumulation and functions of tumor-expanded myeloid-derived suppressor cells via targeting of PTEN. J Immunol 2012; 188: 5500–5510.
Li L, Zhang J, Diao W, Wang D, Wei Y, Zhang CY et al. MicroRNA-155 and MicroRNA-21 promote the expansion of functional myeloid-derived suppressor cells. J Immunol 2014; 192: 1034–1043.
Marigo I, Bosio E, Solito S, Mesa C, Fernandez A, Dolcetti L et al. Tumor-induced tolerance and immune suppression depend on the C/EBPbeta transcription factor. Immunity 2010; 32: 790–802.
Zhang M, Liu Q, Mi S, Liang X, Zhang Z, Su X et al. Both miR-17-5p and miR-20a alleviate suppressive potential of myeloid-derived suppressor cells by modulating STAT3 expression. J Immunol 2011; 186: 4716–4724.
Squadrito ML, Etzrodt M, De Palma M, Pittet MJ . MicroRNA-mediated control of macrophages and its implications for cancer. Trends Immunol 2013; 34: 350–359.
Squadrito ML, Pucci F, Magri L, Moi D, Gilfillan GD, Ranghetti A et al. miR-511-3p modulates genetic programs of tumor-associated macrophages. Cell Rep 2012; 1: 141–154.
Cai X, Yin Y, Li N, Zhu D, Zhang J, Zhang CY et al. Re-polarization of tumor-associated macrophages to pro-inflammatory M1 macrophages by microRNA-155. J Mol Cell Biol 2012; 4: 341–343.
Cui TX, Kryczek I, Zhao L, Zhao E, Kuick R, Roh MH et al. Myeloid-derived suppressor cells enhance stemness of cancer cells by inducing microRNA101 and suppressing the corepressor CtBP2. Immunity 2013; 39: 611–621.
Ishimoto T, Sugihara H, Watanabe M, Sawayama H, Iwatsuki M, Baba Y et al. Macrophage-derived reactive oxygen species suppress miR-328 targeting CD44 in cancer cells and promote redox adaptation. Carcinogenesis 2013; 35: 1003–1011.
Sugihara H, Ishimoto T, Watanabe M, Sawayama H, Iwatsuki M, Baba Y et al. Identification of miR-30e* regulation of Bmi1 expression mediated by tumor-associated macrophages in gastrointestinal cancer. PLoS One 2013; 8: e81839.
Dangi-Garimella S, Strouch MJ, Grippo PJ, Bentrem DJ, Munshi HG . Collagen regulation of let-7 in pancreatic cancer involves TGF-beta1-mediated membrane type 1-matrix metalloproteinase expression. Oncogene 2011; 30: 1002–1008.
Valadi H, Ekstrom K, Bossios A, Sjostrand M, Lee JJ, Lotvall JO . Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol 2007; 9: 654–659.
Zernecke A, Bidzhekov K, Noels H, Shagdarsuren E, Gan L, Denecke B et al. Delivery of microRNA-126 by apoptotic bodies induces CXCL12-dependent vascular protection. Sci Signal 2009; 2: ra81.
Kosaka N, Iguchi H, Yoshioka Y, Takeshita F, Matsuki Y, Ochiya T . Secretory mechanisms and intercellular transfer of microRNAs in living cells. J Biol Chem 2010; 285: 17442–17452.
Kroh EM, Parkin RK, Mitchell PS, Tewari M . Analysis of circulating microRNA biomarkers in plasma and serum using quantitative reverse transcription-PCR (qRT-PCR). Methods 2010; 50: 298–301.
Vickers KC, Palmisano BT, Shoucri BM, Shamburek RD, Remaley AT . MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat Cell Biol 2011; 13: 423–433.
Gould SJ, Raposo G . As we wait: coping with an imperfect nomenclature for extracellular vesicles. J Extracell Vesicles 2013; 2: 20389.
Vader P, Breakefield XO, Wood MJ . Extracellular vesicles: emerging targets for cancer therapy. Trends Mol Med 2014; 20: 385–393.
Kosaka N, Ochiya T . Unraveling the mystery of cancer by secretory microRNA: horizontal microRNA transfer between living cells. Front Genet 2011; 2: 97.
Turchinovich A, Samatov TR, Tonevitsky AG, Burwinkel B . Circulating miRNAs: cell-cell communication function? Front Genet 2013; 4: 119.
Kosaka N, Iguchi H, Hagiwara K, Yoshioka Y, Takeshita F, Ochiya T . Neutral sphingomyelinase 2 (nSMase2)-dependent exosomal transfer of angiogenic microRNAs regulate cancer cell metastasis. J Biol Chem 2013; 288: 10849–10859.
Yang M, Chen J, Su F, Yu B, Su F, Lin L et al. Microvesicles secreted by macrophages shuttle invasion-potentiating microRNAs into breast cancer cells. Mol Cancer 2011; 10: 117.
Aucher A, Rudnicka D, Davis DM . MicroRNAs transfer from human macrophages to hepato-carcinoma cells and inhibit proliferation. J Immunol 2013; 191: 6250–6260.
Bronisz A, Wang Y, Nowicki MO, Peruzzi P, Ansari KI, Ogawa D et al. Extracellular vesicles modulate the glioblastoma microenvironment via a tumor suppression signaling network directed by miR-1. Cancer Res 2014; 74: 738–750.
Shen G, Li X, Jia YF, Piazza GA, Xi Y . Hypoxia-regulated microRNAs in human cancer. Acta Pharmacol Sin 2013; 34: 336–341.
Kulshreshtha R, Ferracin M, Wojcik SE, Garzon R, Alder H, Agosto-Perez FJ et al. A microRNA signature of hypoxia. Mol Cell Biol 2007; 27: 1859–1867.
Huang X, Ding L, Bennewith KL, Tong RT, Welford SM, Ang KK et al. Hypoxia-inducible mir-210 regulates normoxic gene expression involved in tumor initiation. Mol Cell 2009; 35: 856–867.
Bruning U, Cerone L, Neufeld Z, Fitzpatrick SF, Cheong A, Scholz CC et al. MicroRNA-155 promotes resolution of hypoxia-inducible factor 1alpha activity during prolonged hypoxia. Mol Cell Biol 2011; 31: 4087–4096.
Loayza-Puch F, Yoshida Y, Matsuzaki T, Takahashi C, Kitayama H, Noda M . Hypoxia and RAS-signaling pathways converge on, and cooperatively downregulate, the RECK tumor-suppressor protein through microRNAs. Oncogene 2010; 29: 2638–2648.
Chan YC, Khanna S, Roy S, Sen CK . miR-200b targets Ets-1 and is down-regulated by hypoxia to induce angiogenic response of endothelial cells. J Biol Chem 2011; 286: 2047–2056.
Crosby ME, Kulshreshtha R, Ivan M, Glazer PM . MicroRNA regulation of DNA repair gene expression in hypoxic stress. Cancer Res 2009; 69: 1221–1229.
Huang X, Le QT, Giaccia AJ . MiR-210—micromanager of the hypoxia pathway. Trends Mol Med 2010; 16: 230–237.
Ivan M, Huang X . miR-210: fine-tuning the hypoxic response. Adv Exp Med Biol 2014; 772: 205–227.
Jafarifar F, Yao P, Eswarappa SM, Fox PL . Repression of VEGFA by CA-rich element-binding microRNAs is modulated by hnRNP L. EMBO J 2011; 30: 1324–1334.
Sempere LF, Christensen M, Silahtaroglu A, Bak M, Heath CV, Schwartz G et al. Altered microRNA expression confined to specific epithelial cell subpopulations in breast cancer. Cancer Res 2007; 67: 11612–11620.
Sempere LF, Preis M, Yezefski T, Ouyang H, Suriawinata AA, Silahtaroglu A et al. Fluorescence-based codetection with protein markers reveals distinct cellular compartments for altered microRNA expression in solid tumors. Clin Cancer Res 2010; 16: 4246–4255.
Sempere LF . Integrating contextual miRNA and protein signatures for diagnostic and treatment decisions in cancer. Expert Rev Mol Diagn 2011; 11: 813–827.
Nouraee N, Van Roosbroeck K, Vasei M, Semnani S, Samaei NM, Naghshvar F et al. Expression, tissue distribution and function of miR-21 in esophageal squamous cell carcinoma. PLoS One 2013; 8: e73009.
Carroll AP, Tooney PA, Cairns MJ . Context-specific microRNA function in developmental complexity. J Mol Cell Biol 2013; 5: 73–84.
Huang JC, Babak T, Corson TW, Chua G, Khan S, Gallie BL et al. Using expression profiling data to identify human microRNA targets. Nat Methods 2007; 4: 1045–1049.
Ulitsky I, Laurent LC, Shamir R . Towards computational prediction of microRNA function and activity. Nucleic Acids Res 2010; 38: e160.
Liu B, Liu L, Tsykin A, Goodall GJ, Green JE, Zhu M et al. Identifying functional miRNA-mRNA regulatory modules with correspondence latent dirichlet allocation. Bioinformatics 2010; 26: 3105–3111.
Bossel Ben-Moshe N, Avraham R, Kedmi M, Zeisel A, Yitzhaky A, Yarden Y et al. Context-specific microRNA analysis: identification of functional microRNAs and their mRNA targets. Nucleic Acids Res 2012; 40: 10614–10627.
Suzuki HI, Mihira H, Watabe T, Sugimoto K, Miyazono K . Widespread inference of weighted microRNA-mediated gene regulation in cancer transcriptome analysis. Nucleic Acids Res 2013; 41: e62.
Muramatsu F, Kidoya H, Naito H, Sakimoto S, Takakura N . microRNA-125b inhibits tube formation of blood vessels through translational suppression of VE-cadherin. Oncogene 2013; 32: 414–421.
Cubillos-Ruiz JR, Baird JR, Tesone AJ, Rutkowski MR, Scarlett UK, Camposeco-Jacobs AL et al. Reprogramming tumor-associated dendritic cells in vivo using miRNA mimetics triggers protective immunity against ovarian cancer. Cancer Res 2012; 72: 1683–1693.
Acknowledgements
We thank members of the Department of Molecular Pathology, the University of Tokyo, Tokyo, Japan. This work was supported by KAKENHI (Grants-in-Aid for Young Scientists (A) (No. 24689018, to HIS) and for Scientific Research on Innovative Areas ‘RNA regulation’ (No. 23112702, to HIS) and ‘Integrative research on cancer microenvironment network’(No. 22112002, to KM)) from the MEXT of Japan, the Cell Science Research Foundation and JSPS Postdoctoral Fellowship for Research Abroad. We apologize to colleagues whose work was not cited because of space limitations.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
Works by Dr Suzuki and Dr Miyazono have been funded by the Ministry of Education, Culture, Sports, Science, and Technology (MEXT) of Japan. Dr Suzuki is also supported by a fund from the Cell Science Research Foundation. Dr Miyazono has received compensation as a member of the scientific advisory board of Isarna Therapeutics GmbH. He also owns stock in Immunofuture, Inc. Mr Matsuyama is currently an employee of the Otsuka Maryland Medicinal Laboratories, Inc. Dr Katsura declares no potential conflict of interest.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/4.0/
About this article

Cite this article
Suzuki, H., Katsura, A., Matsuyama, H. et al. MicroRNA regulons in tumor microenvironment. Oncogene 34, 3085–3094 (2015). https://doi.org/10.1038/onc.2014.254
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/onc.2014.254
This article is cited by
-
SEVs-mediated miR-6750 transfer inhibits pre-metastatic niche formation in nasopharyngeal carcinoma by targeting M6PR
Cell Death Discovery (2023)
-
The role of microRNAs in the modulation of cancer-associated fibroblasts activity during pancreatic cancer pathogenesis
Journal of Physiology and Biochemistry (2023)
-
MicroRNA in lung cancer—a novel potential way for early diagnosis and therapy
Journal of Applied Genetics (2023)
-
Liquid biopsy at the frontier of detection, prognosis and progression monitoring in colorectal cancer
Molecular Cancer (2022)
-
Tumor-derived miR-130b-3p induces cancer-associated fibroblast activation by targeting SPIN90 in luminal A breast cancer
Oncogenesis (2022)
