
Abstract
Activation of RIPK1 controls TNF-mediated apoptosis, necroptosis and inflammatory pathways1. Cleavage of human and mouse RIPK1 after residues D324 and D325, respectively, by caspase-8 separates the RIPK1 kinase domain from the intermediate and death domains. The D325A mutation in mouse RIPK1 leads to embryonic lethality during mouse development2,3. However, the functional importance of blocking caspase-8-mediated cleavage of RIPK1 on RIPK1 activation in humans is unknown. Here we identify two families with variants in RIPK1 (D324V and D324H) that lead to distinct symptoms of recurrent fevers and lymphadenopathy in an autosomal-dominant manner. Impaired cleavage of RIPK1 D324 variants by caspase-8 sensitized patients’ peripheral blood mononuclear cells to RIPK1 activation, apoptosis and necroptosis induced by TNF. The patients showed strong RIPK1-dependent activation of inflammatory signalling pathways and overproduction of inflammatory cytokines and chemokines compared with unaffected controls. Furthermore, we show that expression of the RIPK1 mutants D325V or D325H in mouse embryonic fibroblasts confers not only increased sensitivity to RIPK1 activation-mediated apoptosis and necroptosis, but also induction of pro-inflammatory cytokines such as IL-6 and TNF. By contrast, patient-derived fibroblasts showed reduced expression of RIPK1 and downregulated production of reactive oxygen species, resulting in resistance to necroptosis and ferroptosis. Together, these data suggest that human non-cleavable RIPK1 variants promote activation of RIPK1, and lead to an autoinflammatory disease characterized by hypersensitivity to apoptosis and necroptosis and increased inflammatory response in peripheral blood mononuclear cells, as well as a compensatory mechanism to protect against several pro-death stimuli in fibroblasts.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others


References
Yuan, J., Amin, P. & Ofengeim, D. Necroptosis and RIPK1-mediated neuroinflammation in CNS diseases. Nat. Rev. Neurosci. 20, 19–33 (2019).
Zhang, X., Dowling, J. P. & Zhang, J. RIPK1 can mediate apoptosis in addition to necroptosis during embryonic development. Cell Death Dis. 10, 245 (2019).
Newton, K. et al. Cleavage of RIPK1 by caspase-8 is crucial for limiting apoptosis and necroptosis. Nature 574, 428–431 (2019).
Xu, D. et al. TBK1 suppresses RIPK1-driven apoptosis and inflammation during development and in aging. Cell 174, 1477–1491 (2018).
Meng, H. et al. Death-domain dimerization-mediated activation of RIPK1 controls necroptosis and RIPK1-dependent apoptosis. Proc. Natl Acad. Sci. USA 115, E2001–E2009 (2018).
Lin, Y., Devin, A., Rodriguez, Y. & Liu, Z. G. Cleavage of the death domain kinase RIP by caspase-8 prompts TNF-induced apoptosis. Genes Dev. 13, 2514–2526 (1999).
Degterev, A. et al. Identification of RIP1 kinase as a specific cellular target of necrostatins. Nat. Chem. Biol. 4, 313–321 (2008).
Ofengeim, D. et al. Activation of necroptosis in multiple sclerosis. Cell Reports 10, 1836–1849 (2015).
Sun, L. et al. Mixed lineage kinase domain-like protein mediates necrosis signaling downstream of RIP3 kinase. Cell 148, 213–227 (2012).
Nigro, P., Pompilio, G. & Capogrossi, M. C. Cyclophilin A: a key player for human disease. Cell Death Dis. 4, e888 (2013).
Christofferson, D. E. & Yuan, J. Cyclophilin A release as a biomarker of necrotic cell death. Cell Death Differ. 17, 1942–1943 (2010).
Zhu, K. et al. Necroptosis promotes cell-autonomous activation of proinflammatory cytokine gene expression. Cell Death Dis. 9, 500 (2018).
Henry, C. M. & Martin, S. J. Caspase-8 acts in a non-enzymatic role as a SCAFFOLD for assembly of a pro-inflammatory “FADDosome” complex upon TRAIL Stimulation. Mol. Cell 65, 715–729 (2017).
Ingold, I. et al. Selenium utilization by GPX4 Is required to prevent hydroperoxide-induced ferroptosis. Cell 172, 409–422 (2018).
Stockwell, B. R. et al. Ferroptosis: a regulated cell death nexus linking metabolism, redox biology, and disease. Cell 171, 273–285 (2017).
Dixon, S. J. & Stockwell, B. R. The role of iron and reactive oxygen species in cell death. Nat. Chem. Biol. 10, 9–17 (2014).
Zhang, D.-W. et al. RIP3, an energy metabolism regulator that switches TNF-induced cell death from apoptosis to necrosis. Science 325, 332–336 (2009).
Zhang, Y. et al. RIP1 autophosphorylation is promoted by mitochondrial ROS and is essential for RIP3 recruitment into necrosome. Nat. Commun. 8, 14329 (2017).
Cuchet-Lourenço, D. et al. Biallelic RIPK1 mutations in humans cause severe immunodeficiency, arthritis, and intestinal inflammation. Science 361, 810–813 (2018).
Li, Y. et al. Human RIPK1 deficiency causes combined immunodeficiency and inflammatory bowel diseases. Proc. Natl Acad. Sci. USA 116, 970–975 (2019).
Ito, Y. et al. RIPK1 mediates axonal degeneration by promoting inflammation and necroptosis in ALS. Science 353, 603–608 (2016).
Ofengeim, D. et al. RIPK1 mediates a disease-associated microglial response in Alzheimer’s disease. Proc. Natl Acad. Sci. USA 114, E8788–E8797 (2017).
Zhou, Q. et al. Loss-of-function mutations in TNFAIP3 leading to A20 haploinsufficiency cause an early-onset autoinflammatory disease. Nat. Genet. 48, 67–73 (2016).
Zhou, Q. et al. Early-onset stroke and vasculopathy associated with mutations in ADA2. N. Engl. J. Med. 370, 911–920 (2014).
Zhou, Q. et al. Biallelic hypomorphic mutations in a linear deubiquitinase define otulipenia, an early-onset autoinflammatory disease. Proc. Natl Acad. Sci. USA 113, 10127–10132 (2016).
Retterer, K. et al. Clinical application of whole-exome sequencing across clinical indications. Genet. Med. 18, 696–704 (2016).
Wu, Z. et al. Chaperone-mediated autophagy is involved in the execution of ferroptosis. Proc. Natl Acad. Sci. USA 116, 2996–3005 (2019).
Ding, Y. et al. Reference values for peripheral blood lymphocyte subsets of healthy children in China. J. Allergy Clin. Immunol. 142, 970–973 (2018).
Acknowledgements
We thank the patients, their families and the unaffected controls for their support during this research study. We thank J. Zhang for Ripk1+/+Ripk3−/− and Ripk1D325A/D325ARipk3−/− MEFs. We thank L. Shen, M. Shi, M. Yu and R. Wang for help. Q.Z. received the grants 2018YFC1004903 from National Key Research and Development Project, 31771548 and 81971528 from The National Natural Science Foundation of China, LR19H100001 from Zhejiang Provincial Natural Science Foundation of China and 2018QN81009 from the Fundamental Research Funds for the Central Universities. X.W. received the grant 81373221 from The National Natural Science Foundation of China. The work of Z.W., H.P. and X.H. was supported by the National Key R&D Program of China (2016YFA0501900) and the China National Natural Science Foundation (31530041). The work of H.L. was supported by Ningbo Health Branding Subject Fund (PPXK2018-06).
Author information
Authors and Affiliations
Contributions
P.T., J.S., Z.W., S.W., J.W. and W.L. contributed equally. H.P., R.B., J.Z., Y.W. and P.Y.L. contributed equally. Q.Z., J.Y., X.W. and X.Y. designed the study, directed and supervised the research. P.T., J.S., Z.W., S.W., J.W., W.L. and H.P. performed experiments and analysed most of the data. J.Z., Y.W., P.Y.L., R.F., H.H., Z.Y., X.H. and Y.Z. performed experiments. J.S., R.B., W.Y., Q.Z., J.H., W.W., B.S., M.Y., D.L., H.L., D.D., K.H., K.M., C.R., J.M., R.M.L. and X.W. enrolled the patients, collected and interpreted clinical information. N.D. provided comments. Q.Z., J.Y., X.Y. and P.T. wrote the manuscript, with input from others. I.A. provided valuable advice and edited the manuscript. All authors contributed to the review and approval of the manuscript.
Corresponding authors
Ethics declarations
Competing interests
J.Y. is a consultant of Denali Therapeutics. R.B., K.H. and K.M. are employees of GeneDx, Inc. The rest of authors declare no competing financial interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Peer review information Nature thanks Min Ae Lee-Kirsch, Egil Lien and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Extended data figures and tables
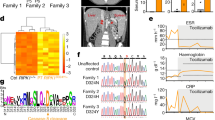
Extended Data Fig. 1 Identification of RIPK1 variants and their effects.
a, Schematic of the WES data-filtering approach under the assumption of de novo inheritance in family 1, leading to the identification of a novel RIPK1 variant. INDEL, frameshift or non-frameshift insertions and deletions; SNP, single nucleotide polymorphisms including missense, splice-site and stop-codon variants. b, Exome sequencing reads covering the D324V variant in family 1, displayed by the integrative genomics viewer. c, Confirmation of RIPK1 variants at residue Asp324 for patients P1–P5 by Sanger sequencing. d, Evolutionary conservation of the caspase-8 cleavage site D324 in RIPK1. Amino acid sequence of RIPK1 flanking D324 was aligned by ClustalW across various species. e, Schematic domain structure of RIPK1. The position of identified variants leading to defective caspase-8 cleavage is indicated. f, In silico analysis of novel RIPK1 variants at D324. cDNA positions are determined according to the reference sequence NM_003804. Four predictions including SIFT, PolyPhen-2, LRT and Mutation Taster annotated by ANNOVAR were included in the analysis. D indicates damaging or deleterious variant. The gnomAD database includes 123,136 exomes and 15,496 genomes. The Kaviar database includes 77,238 exomes and genomes. g, RIPK1 cleavage site variants caused defective cleavage in vitro. HEK293T cells were transiently transfected with wild-type or mutant RIPK1 plasmids followed by immunoblotting of cell lysates. EV, empty vector; H1, H2, different cloning plasmids of RIPK1(D324H) variant; V1, V2, different cloning plasmids of RIPK1(D324V) variant; WT, wild-type RIPK1 plasmid. For gel source data, see Supplementary Fig. 1. Results are representative of three independent experiments. h, The degradation of wild-type and D325A mutant RIPK1 protein was analysed by CHX chase assay. Top, Ripk1D325A/D325ARipk3−/− and Ripk1+/+Ripk3−/− MEFs were incubated with 50 µg ml−1 CHX for the indicated period of time and collected for western blot. Results are representative of three independent experiments. The western blot was quantified by Image J. Bottom, the relative RIPK1 protein level was normalized to CHX-untreated cells. Data are mean ± s.d., n = 3. Circles correspond to each independent experiment. For gel source data, see Supplementary Fig. 1. i, Neither mutant disrupted the recruitment of RIPK1 and caspase-8 into the FADDosome. Ripk1-knockout MEFs complemented with wild-type RIPK1, or D325V or D325H mutant were treated as indicated for 1 or 3 h. T/Z, 50 ng ml−1 TNF, 50 μM Z-VAD-FMK. ‘+’ denotes 20 μM Nec-1s. Lysates were immunoprecipitated with anti-FADD, and analysed by immunoblotting using the indicated antibodies. For gel source data, see Supplementary Fig. 1. Results are representative of three independent experiments. j, Unlabelled in vitro transcription and translation of wild-type and mutant RIPK1 constructs (D324V, D324H and D324K) were performed in the TNT T7 Quick Coupled Transcription/Translation System followed by incubation with purified recombinant caspase-8 protein for 3 h and then analysed by immunoblotting of RIPK1. CL, cleaved RIPK1; FL, full-length RIPK1. For gel source data, see Supplementary Fig. 1. Results are representative of three independent experiments. k, The D324V variant disrupted the RIPK1 cleavage by caspase-8. Fibroblasts from patient P1 and an unaffected control were treated with TNF and CHX for the indicated amount of time followed by immunoblotting analysis. For gel source data, see Supplementary Fig. 1. Results are representative of three independent experiments.
Extended Data Fig. 2 Increased pro-inflammatory signalling in patient PBMCs.
a, Intracellular cytokine staining of patient PBMCs showed expression of IL-6, TNF and IL-8 in CD14+ monocytes at baseline (unstimulated, UNS) (top) and after LPS stimulation for 6 h (bottom) compared to one representative (left) and four paediatric unaffected controls (right). Data are mean ± s.e.m. Circles correspond to each tested individual. b, Intracellular cytokine staining of patient PBMCs showed increased expression of IL-6 in patient CD3+CD4+ T cells after LPS stimulation (1 μg ml−1) for 6 h compared to 1 representative (left) and 4 paediatric unaffected controls (right). Data are mean ± s.e.m. Circles correspond to each tested individual. c, Basal phosphorylation of STAT3 (top) and phosphorylation of p38 after LPS stimulation (bottom) of patient monocytes (CD14+), B cells (CD19+), and T cells (CD3+) compared to one representative (left) and four paediatric unaffected controls (right) as determined by flow cytometry analysis. Data are mean ± s.e.m. Circles correspond to each tested individual. d, Single-cell RNA sequencing revealed a higher percentage of monocytes in P1 compared with an age- and sex-matched unaffected control (C1). e, f, Single-cell RNA sequencing revealed that the NF-κB (e) and type I IFN (f) signalling pathways were upregulated in patient CD14+ and CD16+ monocytes compared with an age- and sex-matched unaffected control (C1) and an adult control (AC). The adult control data were downloaded from 10X Genomics. Analysis of patient sample was performed in duplicate. For gene names, see Supplementary Fig. 2. g, Violin plots showing the distribution of gene expression of selected genes in different cell clusters for P1 and an age- and sex-matched unaffected control. n = 1,007 cells for patient and n = 4,340 cells for control CD4 T cell cluster. n = 1,868 cells for patient and n = 1,427 cells for control B cell cluster. n = 422 cells for patient and n = 1,478 cells for control CD8 T cell cluster. n = 302 cells for patient and n = 1,101 cells for control NK cell cluster. n = 1,125 cells for patient and n = 241 cells for control CD14 monocytes cluster. n = 1,184 cells for patient and n = 78 cells for control CD16 monocytes cluster. n = 371 cells for patient and n = 563 cells for control T memory cell cluster. n = 333 cells for patient and n = 524 cells for control γδ T cell cluster. n = 249 cells for patient and n = 597 cells for control NK cell/T doublets cluster. n = 182 cells for patient and n = 228 cells for control B activated cell cluster. n = 357 cells for patient and n = 20 cells for control dendritic cell cluster. n = 117 cells for patient and n = 217 cells for control naive B cell cluster. n = 279 cells for patient and n = 35 cells for control erythrocytes cluster. n = 29 cells for patient and n = 86 cells for control megakaryocyte cell cluster. n = 78 cells for patient and n = 23 cells for control plasma cell cluster. n = 33 cells for patient and n = 34 cells for control plasmacytoid dendritic cell cluster. The PBMCs from P1 for in a–g were obtained during a fever episode.
Extended Data Fig. 3 Increased expression of genes in NF-κB and type-I IFN pathways in patient PBMCs.
a, Visualization of expression of genes involved in NF-κB pathway (SOCS3, CD40, TLR2, IL1RN, IL6, CD38, TAP1, IL15, ICOS, CD83, TNF, IFNG, CXCL2, FCGRT, TNFSF10, ICAM1, IGHG1, TNFRSF1B, CCL3, IGHG4 and TREM1) (coloured single cells) on UMAP plot projecting PBMCs from patient P1 (n = 7,936 cells) and an age- and sex-matched unaffected control (n = 10,992 cells). b, Visualization of expression of genes involved in type-I IFN signalling pathway (IFI6, IFI30, OAS1, GBP1, IFI27, ISG15, OAS2, SOCS1, IFI44, IRF7, OAS3, LY6E, IFI44L, MX1, OASL, RSAD2, IFIT3, HERC5, USP18, IFIT2, HERC6 and EPSTI1) (coloured single cells) on UMAP plot projecting PBMCs from patient P1 (n = 7,936 cells) and an age- and sex-matched unaffected control (n = 10,992 cells). The PBMCs from P1 in a and b were obtained during a fever episode.
Extended Data Fig. 4 Patient PBMCs are sensitive to TNF-induced cell death but not to ferroptosis.
a, qPCR of PBMCs confirmed comparative expression levels of cytokine and chemokine-related genes in P1, after 4 months of tocilizumab treatment, compared to 8 paediatric unaffected controls. Data are mean ± s.e.m. Circles correspond to each tested individual. Analysis of each sample was performed in triplicate. b, Patient PBMCs were hypersensitive to TNF-induced cell death. PBMCs from 8 age-matched unaffected controls and patients P1 and P3 were treated as indicated for 24 h. N, 20 μM Nec-1s; S, 100 nM SM-164; T, 100 ng ml−1 TNF; Z, 100 μM Z-VAD-FMK. Cell death was measured by ToxiLight assay. Data are mean ± s.e.m. Circles correspond to each tested individual. Analysis of each sample was performed in triplicate. c, Induction of necroptosis and apoptosis by TNF in the patient PBMCs. PBMCs from patient P5 and a paediatric unaffected control were treated with indicated stimulation for 24 h before cell lysates were analysed by immunoblotting. For gel source data, see Supplementary Fig. 1. Results are representative of two independent experiments. d, Patient PBMCs stimulated with TNF alone exhibited upregulated gene expression of inflammatory signals, which was reduced by Nec-1s. PBMCs of patient and 2 unaffected controls were treated with 100 ng ml−1 TNF or TNF plus 20 μM Nec-1s for 24 h before being analysed by NanoString. e, The transcription levels of RIPK1 in PBMCs from patient and unaffected controls measured by qPCR. Data are mean ± s.e.m. Circles correspond to each tested individual. Analysis of each sample was performed in triplicate. f, PBMCs of patient and two paediatric unaffected controls showed similar responses to RSL3-induced ferroptosis. Circles correspond to each tested individual. Analysis of each sample was performed in triplicate. g, RNA sequencing of patient PBMCs indicated no difference in expression patterns of genes involved in ferroptosis and antioxidant when compared to three paediatric unaffected controls. Analysis of each sample was performed in duplicate. h, Single-cell RNA sequencing did not reveal distinct expression patterns of genes involved in ferroptosis and antioxidant in patient CD14+ and CD16+ monocytes compared with an age- and sex-matched unaffected control (C1) and an adult control (AC). The adult control data were downloaded from 10X Genomics. i, GSH concentration in PBMCs from P1 was similar to three paediatric unaffected controls. Data are mean ± s.e.m. Circles correspond to each tested individual. Analysis of each sample was performed in triplicate. The PBMCs for a, b, d, f and i were obtained during remission. The PBMCs for c, e, g and h were obtained during a fever episode.
Extended Data Fig. 5 RIPK1 cleavage site variants in MEFs promote cell death and inflammatory response but have no protective effect against ferroptosis.
a, b, RIPK1 cleavage site variants in MEFs promote cell death induced by TNF. Ripk1−/− MEFs were complemented with: GFP; wild-type RIPK1; or D325V or D325H mutant, and treated for 12 h (a) or as indicated (b). N, 20 μM Nec-1s; S, 20 nM SM-164; T, 50 ng ml−1 TNF; Z, 50 μM Z-VAD-FMK. Cell viability and cell death were measured by CellTiter-Glo assay (a) and ToxiLight assay (b), respectively. Data are mean ± s.e.m, n = 3. Circles correspond to each independent experiment. P values were determined by unpaired, two-tailed t-test (shown if P < 0.05). c, Western blots illustrating increased levels of p-S166-RIPK1, p-S345-MLKL and cleaved caspase-3 after stimulation with TNF and SM-164, which were inhibited by Nec-1s. Ripk1−/− MEFs complemented with: GFP; wild-type RIPK1; or D325V or D325H mutant, were treated as indicated. Cell lysates were analysed by immunoblotting using indicated antibodies. For gel source data, see Supplementary Fig. 1. Results are representative of three independent experiments. d, e, D325A knock-in Ripk1 mutation sensitizes Ripk3−/− MEFs to TNF-induced RIPK1-dependent apoptosis. Ripk1D325A/D325ARipk3−/− and Ripk1+/+Ripk3−/− MEFs were simulated with TNF only, TNF plus Nec-1s (d) or a combination of TNF, SM-164 and Nec-1s (e) as indicated (concentrations as in a). Cell death was measured by the SYTOX Green Nucleic Acid Stain assay. Data are mean ± s.e.m., n = 4. Circles correspond to each independent experiment. P values determined by unpaired two-tailed t-test, and indicate the comparison between Ripk1D325A/D325ARipk3−/− and Ripk1+/+Ripk3−/− MEFs after TNF or TNF plus SM-164 stimulation for indicated amount of time. f, RIPK1 cleavage site variants in MEFs sensitize TRAIL-induced cell death. Ripk1−/− MEFs complemented with: GFP, wild-type RIPK1, or D325V or D325H mutant were treated with TRAIL (100 ng ml−1) or TRAIL plus Nec-1s (20 μM) for 36 h. Data are mean ± s.e.m., n = 3. Circles correspond to each independent experiment. P values determined by unpaired two-tailed t-test. g, TRAIL stimulation of Ripk1−/− MEFs complemented with D325V or D325H mutant promotes RIPK1 activation, which was inhibited by Nec-1s (20 μM). TRAIL (100 ng ml−1); TNF (50 ng ml−1) for 12 h. For gel source data, see Supplementary Fig. 1. Results are representative of three independent experiments. h, Ripk1-knockout MEFs complemented with wild-type RIPK1, or D325V or D325H mutant plasmid showed similar responses to erastin- or RSL3-induced ferroptosis. Data are mean ± s.e.m., n = 3. Circles correspond to each independent experiment. i, Western blots of proteins involved in ferroptosis in Ripk1−/− MEFs complemented with wild-type RIPK1, or D325V or D325H mutant. Cells were treated with erastin for 5 or 10 h, followed by immunoblotting of cell lysates. For gel source data, see Supplementary Fig. 1. Results are representative of three independent experiments. j, RNA sequencing of Ripk1−/− MEFs complemented with wild-type RIPK1, D325V or D325H mutant indicated no difference in expression patterns of genes involved in ferroptosis and antioxidant. Analysis of each sample was performed in duplicate. k, GSH concentration of Ripk1−/− MEFs complemented with wild-type RIPK1, D325V or D325H mutant was similar both at baseline and after erastin or glutamate stimulation for 8 h. Data are mean ± s.e.m., n = 3. Circles correspond to each independent experiment. l, Immunofluorescence showed similar levels of cytosolic ROS after erastin stimulation in Ripk1−/− MEFs complemented with wild-type RIPK1, D325V or D325H mutant. Cells were treated by erastin for 8 h before incubation with the cytosolic ROS sensor carboxy-H2DCFDA. Green foci indicate cytosolic ROS. Scale bar, 150 μm. Results are representative of two independent experiments.
Extended Data Fig. 6 Patient fibroblasts were resistant to both necroptosis and ferroptosis.
a, Patient fibroblasts were resistant to necroptosis induced by SM-164, Z-VAD-FMK and TNF or LPS. Fibroblasts from P1 and seven paediatric unaffected controls were treated as indicated for 24 h. LPS, 1 μg ml−1; N, 10 μM Nec-1s; S, 50 nM SM-164; T, 50 ng ml−1 TNF; Z, 50 μM Z-VAD-FMK. Cell death was measured by ToxiLight assay. Data are mean ± s.e.m. Circles correspond to each tested individual. Analysis of each sample was performed in triplicate. b, Patient fibroblasts showed reduced necroptosis signals after SM-164, Z-VAD-FMK and LPS stimulation compared to six paediatric unaffected controls. Patient and control fibroblasts were treated with indicated stimulation for 6 h (concentrations as in a). Cells were lysed and analysed by immunoblotting with indicated antibodies. For gel source data, see Supplementary Fig. 1. Results are representative of three independent experiments. c, Patient fibroblasts showed reduced necroptosis signals after SM-164, Z-VAD-FMK and TNF stimulation compared with a paediatric unaffected control. Patient and control fibroblasts were treated as indicated for 6 h (concentrations as in a). Cells were lysed and analysed by immunoblotting with indicated antibodies. For gel source data, see Supplementary Fig. 1. Results are representative of three independent experiments. d, The reduction of RIPK1 was rescued by Nec-1s in patient fibroblasts. Fibroblasts were treated as indicated for 24 h (concentrations as in a). NSA, 0.5 μM necrosulfonamide. Cell lysates were analysed by immunoblotting using indicated antibodies. For gel source data, see Supplementary Fig. 1. Results are representative of three independent experiments. e, Patient fibroblast showed reduced transcription levels of RIPK1 compared to five paediatric unaffected controls. The mRNA levels of RIPK1 were measured by qPCR. Data are mean ± s.e.m. Circles correspond to each tested individual. Analysis of each sample was performed in triplicate. f, Patient fibroblasts exhibited reduced TNFR1 expression at baseline compared to five paediatric unaffected controls. For gel source data, see Supplementary Fig. 1. Results are representative of three independent experiments. g, Patient fibroblasts displayed downregulation of genes involved in cell death compared with three paediatric unaffected controls. Analysis of each sample was performed in duplicate. h, Patient fibroblasts were resistant to erastin- or RSL3-induced ferroptosis compared with three paediatric unaffected controls. Cell death was measured by ToxiLight assay. Data are mean ± s.e.m. Circles correspond to each tested individual. Analysis of each sample was performed in triplicate.
Supplementary information
Supplementary Figures
This file contains Supplementary Figure 1 (Uncropped images of Western blots used in the indicated figures) and Supplementary Figure 2 (Heatmaps with gene information).
Supplementary Table 1 |
Rare variants (allele frequency < 5%) of genes associated with autoinflammatory diseases in family 1.
Supplementary Table 2 |
Rare variants (allele frequency < 5%) of genes associated with autoinflammatory diseases in family 2.
Rights and permissions
About this article

Cite this article
Tao, P., Sun, J., Wu, Z. et al. A dominant autoinflammatory disease caused by non-cleavable variants of RIPK1. Nature 577, 109–114 (2020). https://doi.org/10.1038/s41586-019-1830-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-019-1830-y
This article is cited by
-
Cellular heterogeneity in TNF/TNFR1 signalling: live cell imaging of cell fate decisions in single cells
Cell Death & Disease (2024)
-
Immunogenic cell death in cancer: targeting necroptosis to induce antitumour immunity
Nature Reviews Cancer (2024)
-
ZBP1 and TRIF trigger lethal necroptosis in mice lacking caspase-8 and TNFR1
Cell Death & Differentiation (2024)
-
RIPK3 cleavage is dispensable for necroptosis inhibition but restricts NLRP3 inflammasome activation
Cell Death & Differentiation (2024)
-
Caspase cleavage of RIPK3 after Asp333 is dispensable for mouse embryogenesis
Cell Death & Differentiation (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
