
Abstract
CYP24A1 and CYP27B1 are critical genes determining 1α,25(OH)2D3 concentration and impacting on carcinogenesis. A case–control study including 528 colorectal cancer (CRC) patients and 605 cancer-free controls and a follow-up study with 317 cases were conducted in northeast China. Genotypes were tested by TaqMan Genotyping Assays. Individuals carrying the GG genotype of CYP27B1 G > T (rs10877012) exhibited decreased CRC risk compared with those with the TT genotype (ORadjusted (ORadj) = 0.57, 95% Confidence Interval (CI) = 0.38–0.84). Compared with the TT genotype, a significant association between the CC genotype of CYP27B1 C > T (rs4646536) and a reduced risk of CRC was observed (ORadj = 0.59, 95% CI = 0.40–0.88). We also observed significant combined effects of the two polymorphisms in CYP27B1 with dietary factors, including the intake of cereals, overnight meal, allium vegetables, pork, canned fruit, and braised fish, on CRC risk. These associations remained significant after Bonferroni correction for multiple comparisons. The Hazard Ration (HR) of patients with the AA genotype (CYP24A1 A > G, rs4809957) was 2.38 (95% CI = 1.30–4.37) when compared with the GG genotype. Thus, our findings suggested that two polymorphisms in CYP27B1 are associated with CRC susceptibility. CYP24A1 A > G (rs4809957) polymorphism may lead to a worse prognosis of CRC.
Similar content being viewed by others


Introduction
Colorectal cancer (CRC) is a major public health issue, being the third most common cancer and the fourth most prominent cause of cancer death worldwide. The World Health Organization reported that 253,427 new cases of CRC and 139,416 deaths occurred in China in 20121,2,3. Suspected or established risk factors of CRC include red meat, alcohol drinking, obesity, physical inactivity, and smoking1, 4. In addition to the traditional dietary factors (fibre, fresh fruit and vegetables) that are protective against CRC5, 6, a number of studies have suggested that vitamin D contributes to a reduced risk of this disease7,8,9.
More than 90% of vitamin D that the human body needs is obtained from the conversion of 7-dehydrocholesterol to vitamin D3, which occurs in the skin upon UV-B radiation10. The synthesized 25(OH)D3 is hydroxylated by 1α-hydroxylase encoded by the CYP27B1 gene, and the most active metabolite of vitamin D, 1α,25-dihydroxyvitamin D3 [1α,25(OH)2D3], is yielded in this process11, 12. CYP24A1, which encodes the vitamin D-deactivating enzyme 24-α hydroxylase, is responsible for inactivating vitamin D metabolites13. CYP24A1 converts 1α,25(OH)2D3 to 1,24,25(OH)3D3, which is a less active intermediate14. Thus, both CYP24A1 and CYP27B1 are members of the cytochrome P450 superfamily, which can regulate 1α,25(OH)2D3 metabolism by repressing CYP24A1 and inducing CYP27B1 through negative feedback loops15. Aside from the classic regulatory effects on calcium and phosphate metabolism, 1α,25(OH)2D3 can also inhibit the proliferation of tumour cells16. Several in vitro studies have indicated that 1α,25(OH)2D3 would be able to reduce epithelial cell proliferation, promote differentiation in colon cells, and induce apoptosis in colorectal tumour cell lines17,18,19.
Multiple lines of evidence suggest that genetic factors such as single-nucleotide polymorphisms (SNPs) modify gene expression and consequently influence cancer risk and prognosis. MicroRNAs (miRNAs) are endogenous noncoding RNAs of ~22 nucleotides (nt) in length, which regulate genes by pairing to the 3′-untranslated regions (UTRs) of messenger RNAs (mRNAs) of target genes and specifying mRNA cleavage or repression of protein synthesis20. Some 3′-UTR polymorphisms that may be in the vicinity of a miRNA binding site have been reported to interfere with miRNA function and lead to differential gene expression. The SNPs located within miRNA binding sites could thus influence cancer risk and overall survival21,22,23,24. To date, several studies have focused on the association of polymorphisms in CYP24A1 and CYP27B1 with cancer risk25,26,27. However, very little research has addressed the role of polymorphisms in miRNA binding sites. Besides, as is well known that successfully identifying the interactions between genes and dietary factors is important to explore the aetiology of cancer. A more detailed understanding of gene–environment (diet) interaction may thus also generate the information required to develop strategies for diet modification to reduce the incidence of CRC in individuals with specific genetic variants of CYP24A1 and CYP27B1.
Against this background, we carried out this study to investigate whether polymorphisms in the target sites of miRNA in CYP24A1 and common variation in CYP27B1 are associated with the risk and prognosis of CRC. We also assessed such effects in terms of their combination and interaction with dietary factors regarding the contribution to the risk of CRC.
Results
Characteristics of study subjects
The demographic characteristics of all subjects in this study are summarized in Table 1. The ages (Mean ± SD) of cases and controls were 60.31 ± 11.30 and 57.14 ± 11.21, respectively (P < 0.001). Compared with controls, cases presented a lower body mass index (23.26 ± 3.37, P < 0.001). Consequently, age and BMI were adjusted in the following multivariate analyses. No significant differences were observed between cases and controls for sex (P = 0.685), occupation (P = 0.267) and family history of cancer (P = 0.102).
Polymorphisms of CYP24A1 and CYP27B1 and CRC risk
The genotype distributions of all of the four polymorphisms in controls were in accordance with Hardy–Weinberg equilibrium (P > 0.05). For rs10877012, GG genotype carriers showed a lower risk of CRC than those with the TT genotype (ORadjusted (ORadj) = 0.57, 95% Confidence Interval (CI) = 0.38–0.84) with P-values of 0.005 and 0.020 before and after Bonferroni correction, respectively. The recessive model and additive model showed significant results with P-values of 0.009 and 0.008, respectively; the recessive model (P = 0.036) and additive model (P = 0.032) remained significant after multiple testing correction. For rs4646536, the CC genotype reduced the risk of CRC compared with the TT genotype (ORadj = 0.59, 95% CI = 0.40–0.88) with P-values of 0.010 and 0.040 before and after Bonferroni correction, respectively; all three genetic models showed significant results with P-values of 0.040, 0.023, and 0.010, but only the additive model remained significant after multiple testing correction. We did not observe any noteworthy associations between rs4809957 and rs2762934 in CYP24A1 and CRC risk (Table 2). Based on the Akaike information criterion (AIC) values, the dominant model was chosen for rs4809957 and the recessive model was chosen for the other three SNPs for use in crossover analysis and multivariate regression analysis.
Subgroup analyses
Table 3 displays the results of subgroup analyses for the associations between rs10877012 and rs4646536 polymorphisms in CYP27B1 and CRC risk. Compared with the wild-type genotype (TT), homozygote variant genotypes (GG of rs10877012; CC of rs4646536) reduced the risk of CRC significantly in the elderly (>60 years) (ORadj = 0.39, 95% CI = 0.21–0.71; ORadjusted = 0.42, 95% CI = 0.23–0.78, respectively), and in females (ORadj = 0.43, 95% CI = 0.23–0.79; ORadj = 0.44, 95% CI = 0.23–0.84, respectively). After adjustment for multiple comparisons, all of these results remained significant. Additionally, no significant result was found in the subgroup analysis by occupation. Only GG of rs10877012 was observed with significant reduced risk of colon cancer (ORadj = 0.42, 95% CI = 0.23–0.78). However, we did not find any significant associations between CYP24A1 polymorphisms and CRC risk in the subgroup analyses (data not shown).
Haplotypes of CYP27B1 and CYP24A1 and CRC risk
We constructed haplotypes and estimated haplotype frequencies in cases and controls for the four SNPs. The haplotypes with frequencies ≥3% are shown in Supplemental Table 2. Complete linked dimorphism (D′ = 1) was detected for rs4809957 and rs2762934 in CYP24A1, while D′ = 0.96 was detected for rs10877012 and rs4646536 in CYP27B1. The most common haplotype in CYP24A1 in cases (64.01%) and controls (63.93%) was G-G. Meanwhile, the most common haplotype in CYP27B1 in cases (68.16%) and controls (62.18%) was T-C. Compared with those carrying all other types of haplotype, individuals carrying the G-T haplotype showed a decreased CRC risk (OR = 0.80, 95% CI = 0.66–0.97). In contrast, compared with those carrying all other types of haplotype, individuals carrying the T-C haplotype showed an increased CRC risk (OR = 1.25, 95% CI = 1.04–1.51). However, after the Bonferroni correction, no significant association of any haplotype with CRC risk remained.
Combined and interactive effects of polymorphisms and dietary factors on the risk of CRC
Based on univariate and multivariate analyses of the associations between dietary factors and CRC risk, we found statistically significant results for the consumption of cereals, vegetables, pork, braised fish, soybean, milk, allium vegetables, canned fruit, and overnight meal (Supplemental Table 3). Tables 4 and 5 shows the results of combined and interactive effects of dietary factors and polymorphisms in CYP24A1 and CYP27B1, respectively. For CYP24A1, a significant combined effect was only observed for GA + AA genotype carriers in the dominant genetic model combined with the consumption of cereals (≥50 g/week) [ORgenetic&dietary (ORgd) = 0.41, 95% CI = 0.28–0.60] after Bonferroni correction. No significant interactive effect was observed between dietary factors and polymorphisms in CYP24A1.
For rs10877012, significant combined effects were observed for GG genotype carriers in the recessive genetic model combined with the intake of cereals (≥50 g/week, ORgd = 0.31, 95% CI = 0.19–0.52, P = 0.000004), overnight meal (≤3 times/week, ORgd = 0.45, 95% CI = 0.28–0.73, P = 0.000642), allium vegetables (≥1 times/week, ORgd = 0.40, 95% CI = 0.24–0.68, P = 0.000596), pork (<250 g/week, ORgd = 0.34, 95% CI = 0.20–0.57, P = 0.00065), canned fruit (No, ORgd = 0.32, 95% CI = 0.17–0.61, P = 0.000474), and braised fish (<1 time/week, ORgd = 0.34, 95% CI = 0.21–0.57, P = 0.00062). Similar combined effects were found for rs4646536 in CYP27B1 and dietary factors (as shown in Table 5). However, no significant interactive effects were observed between dietary factors and polymorphisms in CYP27B1.
Polymorphisms, clinical characteristics, and CRC prognosis
The associations of polymorphisms with clinical characteristics in CRC patients are summarized in Supplementary Tables 4 and 5. Significant correlations of rs2765934 in CYP24A1 with histological classification (P = 0.015) and metastasis (P = 0.040) were found, as well as of rs10877012 (P = 0.036) and rs4646536 (P = 0.020) in CYP27B1 with metastasis.
As shown in Table 6, compared with the GG genotype, AA genotype carriers of rs4809957 polymorphism in CYP24A1 had worse prognosis (HR = 2.38, 95% CI = 1.30–4.37). Further analysis of the polymorphisms and the prognosis of CRC depending on different sites were conducted. The prognosis of AA (rs4809957) genotype carriers was worse than that of GG + GA carriers in CRC, colon cancer, and rectal cancer (log-rank tests P < 0.01, P < 0.01, P = 0.02, respectively) (Figure 1A–C). For rs4646536, patients who carried the TT genotype had worse prognosis than those with the CC + CT genotypes (log-rank test P = 0.01) only in colon cancer (Figure 1D).

Kaplan-Meier curves of overall survival in CRC (colon and rectal cancer). for CYP24A1 rs480957 and CYP27B1 rs4646536 polymorphisms ((A). CYP24A1 rs480957 in CRC, (B) CYP24A1 rs480957 in colon cancer, (C) CYP24A1 rs480957 in rectal cancer, (D) CYP27B1 rs4646536 in colon cancer).
Discussion
There is growing evidence that vitamin D reduces the incidence of CRC. At the molecular level, vitamin D suppresses CRC development and growth by affecting cell proliferation, differentiation, apoptosis, and angiogenesis28. Polymorphisms located within miRNA binding sites and other gene regions have been reported to play an important role in gene regulation29. To date, no study has been conducted on the association between polymorphisms in genes related to vitamin D metabolism (CYP24A1, CYP27B1) and the risk of CRC in the Chinese population. In our study, it is notable that the GG genotype of the rs10877012 polymorphism decreased CRC risk by about 44%, compared with the TT genotype. Our data also indicated that, compared with the TT genotype, the CC genotype of the rs4646536 polymorphism decreased this risk by about 43%.
Vitamin D insufficiency is common in the elderly. The reduced capacity of the skin to manufacture cholecalciferol30, 31 has been evidenced in relation to increased CRC risk in this age group7, 9. Based on the subgroup results in our study, compared with the TT genotype, the GG genotype of rs10877012 showed a significant protective role only in elderly participants (>60 years), but not in younger ones (≤60). This difference emphasized the significance of genetic variation on the risk of CRC in the elderly. It indicated that, among the elderly population, individuals carrying the GG genotype exhibit lower activity of CYP24A1 and a higher level of 1α,25(OH)2D3, associated with a reduced CRC risk compared with that of individuals with the TT genotype. However, this genetic effect did not significantly modify the CRC risk in younger adults (≤60) because of sufficient cholecalciferol production. Similarly, oestrogen can promote the formation of activated vitamin D by stimulating the secretion of parathyroid hormone32. In the total of 433 females in this study, 362 were postmenopausal or on the brink of menopause (aged more than 50); thus, their oestrogen levels had declined significantly. The protective effects of variant genotypes were also highlighted with a decreased risk of CRC only in the female subgroup. Therefore, considering the genetic effects, it should be recommended that the elderly (>60) and postmenopausal women with the TT genotype of rs10877012 and rs4646536 should have more vitamin D supplementation than individuals with GG (rs10877012) and CC (rs4646536) genotypes to obtain the same level of susceptibility to CRC. In addition, solar UV-B radiation is responsible for converting the precursor of vitamin D3 (7-dehydrochlolesterol, provitamin D) in the skin into vitamin D3 33. Most humans obtain their required amount of vitamin D (90%–95%) from exposure to sunlight34. Previous epidemiological studies demonstrated that exposure to solar UV-B radiation was associated with a decreased risk of colon cancer35. Physical worker is considered to have more chance of exposure to solar UV-B because of lots of outdoor work. However, in this study, we did not observe significant results in subgroup analysis by occupation.
Several early studies revealed that CRC could be prevented by dietary fibre36, 37. Dietary fibre is a complex carbohydrate derived from plants that escapes digestion in the small intestine and thus reaches the colon. An analysis of data from Europe, North America, and Australia has shown that fibre-rich foods such as cereals are strongly protective against CRC, as are vegetables, while fruit is neutral in this regard38. This study produced results consistent with these previous findings, with cereals showing the strongest protective role against CRC. Moreover, significant combined effects were found for cereal intake and SNPs in CYP24A1 and CYP27B1, with ORs ranging from 0.31 to 0.41.
In view of their identified functions associated with adenoma- or carcinoma-related gut microbes, dietary factors have been recognized as major causes of CRC. As a kind of allium vegetables, garlic contains oil-soluble organosulfur compounds such as ajoene, diallyl sulphide, diallyl disulphide, and diallyl trisulphide, whereas onion mainly contains S-propenyl-cysteine sulphoxide, but also other sulphoxides39. Researchers have found that diallyl sulphide can penetrate bacterial membranes. Thus, researchers have suggested that the organosulphur compounds in allium vegetables have the potential to be used as antimicrobial agents. In this study, a significantly reduced risk was observed as a combined effect of genetic variants in CYP27B1 and the consumption of allium vegetables more than once a week. Moreover, bacterial toxins in overnight meal could also cause destruction of the normal gut microbiota and induce chronic gastroenteritis40. This is reasonable given the significant protective effects of genetic variants in CYP27B1 and fewer than three overnight meals per week.
Additionally, a report from Duke Medicine Health News suggested that red meat increases the risk of CRC. It was also reported that diet and lifestyle changes could prevent 64,000 cases of CRC per year in the USA41. Another systematic review also indicated that red meat intake is associated with an elevated risk of developing CRC42. In this study, compared with the group with the combination of the TT + TG genotype and ≥250 g/week pork intake, the significant combined protective effect of the GG genotype and <250 g/week intake was observed. Especially, this protective effect (OR = 0.34, 95% CI = 0.20–0.57) is much greater than the single main effect of pork (<250 g/week vs. ≥250 g/week, OR = 0.64, 95% CI = 0.49–0.85). It indicated that individuals with the TT + TG genotype should reduce their intake of pork to modify their susceptibility to CRC. Similarly, we found significant combined effects of the GG genotype with no canned fruit intake and with braised fish intake less than once a week. As shown in Table 5, similar combined effects were observed between the two polymorphisms in CYP27B1 and dietary factors.
The emergence of the field of Molecular pathologic epidemiology (MPE) has emerged as an integrative analysis of exposures, host factors (genetic variants) and dysfunction of cells or organ units43, 44. A major value of MPE lies in the provision of a better understanding of heterogeneity in the carcinogenic process and the influences of exogenous and endogenous factors, which should contribute further to personalized prevention and treatment strategies. Based on further validation of the combined effects that we identified in this study, guidelines for dietary intake for individuals with a specific genetic background should be addressed.
There is a paucity of information on the associations of clinical characteristics and 25-OHD level or polymorphisms in genes related to vitamin D metabolism. Four published studies on patients with CRC reported better overall survival among those with higher 25(OH)D3 levels than those with lower levels45,46,47,48. miRNAs are involved in post-transcriptional regulation and can regulate the expression of genes by targeting messenger RNA (mRNA) to degrade or suppress the translation of mRNA49, 50. CYP24A1 catalyses an irreversible and rate-limiting step in the degradation of 1α,25(OH)2D3 27. To our knowledge, this is the first study to reveal a significant effect of rs4809957 polymorphism, which is in a miRNA binding site of the CYP24A1 gene, on the prognosis of CRC patients.
Our study is associated with several potential limitations. First, recall bias may be inevitable when collecting information on dietary factors, although we did our best to minimize it. Second, the frequencies of dietary factors in our questionnaire limited our ability to quantify food intake or calculate the precise intake of vitamins and nutrients. Third, the relatively small size of this study probably reduced the statistical power in the subgroup. Additionally, Bonferroni correction is considered to be a conservative procedure to counteract the problem of multiple comparisons; some ‘real’ effects may have gone undetected in our analysis.
Conclusions
In conclusion, our study suggested that two polymorphisms in the CYP27B1 gene are associated with the risk of CRC, particular in the subgroups of the elderly, women, and non-mental workers. Although the interactions had no significance, we observed combined effects of the polymorphisms in CYP27B1 with dietary factors regarding CRC risk. Moreover, the rs4809957 polymorphism in the CYP24A1 gene may be an independent predictor of survival in CRC among the population in northeast China. Further epidemiological studies with a large scale and more polymorphisms of genes related to vitamin D metabolism are needed to confirm these findings.
Methods
Study subjects
We carried out this study after obtaining written informed consent from the subjects and approval from the Human Research and Ethics Committee of Harbin Medical University. All experiments including all relevant details were performed in accordance with relevant guidelines and regulations.
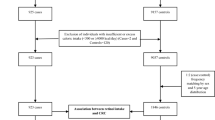
A case–control study was designed to assess the roles of genetic polymorphisms in CYP24A1 and CYP27B1 as well as dietary factors in the risk of CRC. The subjects in this study consisted of 528 patients with primary sporadic colorectal cancer and 605 cancer-free controls (520 hospital-based and 85 community-based). Cases were recruited from the Second and Third Affiliated Hospitals of Harbin Medical University before surgery. Patients were excluded if they suffered from the following diseases: neuroendocrine carcinoma, malignant melanoma, non-Hodgkin’s lymphoma, gastrointestinal stromal tumours, Lynch syndrome, or familial adenomatous polyposis. Controls were recruited from the Second Affiliated Hospital of Harbin Medical University and Hong Qi community of Harbin. Controls with any kinds of cancer or gastrointestinal diseases were excluded. All subjects were enrolled from June 2004 to January 2008. Approximately 5 ml of peripheral venous blood was obtained from all cases either before surgery for the patients or at enrolment for the controls.
A cohort study was also proposed to explore the potential factors associated with the prognosis of CRC patients. For this purpose, among the 528 CRC patients, 317 were followed up from November 2004 to March 2015 via telephone interviews.
Data collection
All subjects were interviewed face-to-face by trained interviewers using the same questionnaire comprising questions on both demographic characteristics and dietary factors. Demographic data included age, sex, height, weight, nationality, education, marital status, occupation, and family history of cancer. Detailed dietary status in the last year before the diagnosis of disease included the consumption of cereals, allium vegetables, pork, milk, fruit, braised fish, soybean, canned fruit, and overnight meals.
All of the CRC patients were followed up from November 2004 to March 2015 by a telephone call once a year. Clinical information (tumour size and location, general classification, histological and pathological types, Duke stage, degree of differentiation, lymph node metastasis, chemotherapy) was collected from the medical records. Overall survival time was calculated from the date at which patients were diagnosed to the date of their death from any cause; patients who were lost to follow-up, suffered from recurrence, or were still alive at the end of the follow-up were measured as censored data.
SNP selection and genotyping
We analysed the SNPs in the miRNA binding sites within the 3′-UTR of CYP24A1 and CYP27B1 by an extensive search in dbSMR (http://miracle.igib.res.in/polyreg/). Using RNA hybrid (http://bibiserv.techfak.uni-bielefeld.de/rnahybrid/submission.html), the Gibbs free energy [DG, expressed in kilojoules per mole (kJ/mol)] for both wild-type and variant alleles of each SNP was determined; the difference of DG between the two alleles (wild-type allele DG − variant allele DG) was calculated as DDG. The sum of all |DDG|s for each SNP (|DDG tot|) was calculated for predicting the biological impact of the polymorphisms. Three polymorphisms (rs4809957, rs2762934, rs16999067) in target sites of miRNA in CYP24A1 were predicted, but one (rs16999067) has no allele frequency in the Chinese population; thus, two polymorphic sites (rs4809957 and rs2762934) were selected in this study. There are no polymorphisms in miRNA target sites in CYP27B1, but two common polymorphisms (rs10877012 and rs4646536) with a minor allele frequency of more than 5% in the Chinese population were also selected based on previous in vitro studies. Detailed information on these four genetic polymorphisms is provided in Supplemental Table 1.
DNA was extracted from leukocytes using the QIAamp DNA Blood Mini Kit, in accordance with the manufacturer’s protocol (Qiagen, Valencia, CA, USA). DNA samples were genotyped using fluorogenic 5′-nuclease assay (TaqMan SNP Genotyping Assay; Applied Biosystems, Foster City, CA, USA) on a Lightcycler® 480|| (Roche, Applied Biosystems) platform. The assay IDs of the probes were C_3120981_30, C_3120982_30, C_25623453_10, and AHCTA6I. A reaction mix of 25 μl contained 10 ng of DNA, 12.5 μl of Universal PCR Master Mix, and 0.625 μl of probe/primer mix. Polymerase chain reaction (PCR) amplification conditions were as follows: an initial step of 95 °C for 10 min, followed by 40 cycles of 92 °C for 15 s and 60 °C for 1 min. We repeated the genotyping for 10% of samples at random.
Statistical analysis
Categorical variables were tested by chi-square test and continuous variables were tested by two-sample t-test between cases and controls. The genotype distributions in controls were tested for Hardy–Weinberg equilibrium. The AIC statistic was applied to determine the goodness of model fitting. The model with the lowest AIC was considered as the best model and was analysed in subsequent crossover analysis and interaction analysis. To correct for multiple testing, we used the Bonferroni-corrected P-value. Haplotypes were evaluated using SHEsis software. We used D′ to assess the extent of linkage disequilibrium (LD) of SNPs in the genome. Univariate and multivariate logistic regression analyses were used to calculate crude and adjusted ORs and 95% CIs. The combined and interactive effects between genetic polymorphisms and dietary factors were estimated by crossover analysis and multivariate logistic regression51.
Kaplan–Meier curve and log-rank test were used to assess the influence of genetic variants on overall survival. HRs and corresponding 95% CIs were computed using univariate and multivariate Cox proportional hazard models. Statistical analyses were carried out using SAS, version 9.2 (SAS Institute, Cary, NC, USA). All reported P-values were two-sided, and P ≤ 0.05 was considered to represent a significant difference.
References
Potter, J. D. Risk factors for colon neoplasia–epidemiology and biology. Eur J Cancer 31A, 1033–1038 (1995).
Jemal, A. et al. Global cancer statistics. CA Cancer J Clin 61, 69–90, doi:10.3322/caac.20107 (2011).
Siegel, R., Desantis, C. & Jemal, A. Colorectal cancer statistics, 2014. CA Cancer J Clin 64, 104–117, doi:10.3322/caac.21220 (2014).
Jain, M. et al. A case-control study of diet and colo-rectal cancer. Int J Cancer 26, 757–768 (1980).
Potter, J. D. & McMichael, A. J. Diet and cancer of the colon and rectum: a case-control study. J Natl Cancer Inst 76, 557–569 (1986).
Potter, J. D. Nutrition and colorectal cancer. Cancer Causes Control 7, 127–146 (1996).
McCullough, M. L. et al. Calcium, vitamin D, dairy products, and risk of colorectal cancer in the Cancer Prevention Study II Nutrition Cohort (United States). Cancer Causes Control 14, 1–12 (2003).
Garland, C. F. et al. Serum 25-hydroxyvitamin D and colon cancer: eight-year prospective study. Lancet 2, 1176–1178 (1989).
Wu, K., Willett, W. C., Fuchs, C. S., Colditz, G. A. & Giovannucci, E. L. Calcium intake and risk of colon cancer in women and men. J Natl Cancer Inst 94, 437–446 (2002).
Deeb, K. K., Trump, D. L. & Johnson, C. S. Vitamin D signalling pathways in cancer: potential for anticancer therapeutics. Nat Rev Cancer 7, 684–700, doi:10.1038/nrc2196 (2007).
Evans, S. R., Schwartz, A. M., Shchepotin, E. I., Uskokovic, M. & Shchepotin, I. B. Growth inhibitory effects of 1,25-dihydroxyvitamin D3 and its synthetic analogue, 1alpha,25-dihydroxy-16-ene-23yne-26,27-hexafluoro-19-nor-cholecalcifero l (Ro 25-6760), on a human colon cancer xenograft. Clin Cancer Res 4, 2869–2876 (1998).
Jacobs, E. T. et al. CYP24A1 and CYP27B1 polymorphisms modulate vitamin D metabolism in colon cancer cells. Cancer Res 73, 2563–2573, doi:10.1158/0008-5472.CAN-12-4134 (2013).
Hamamoto, H. et al. Structure-function analysis of vitamin D 24-hydroxylase (CYP24A1) by site-directed mutagenesis: amino acid residues responsible for species-based difference of CYP24A1 between humans and rats. Mol Pharmacol 70, 120–128, doi:10.1124/mol.106.023275 (2006).
Jones, G., Prosser, D. E. & Kaufmann, M. 25-Hydroxyvitamin D-24-hydroxylase (CYP24A1): its important role in the degradation of vitamin D. Arch Biochem Biophys 523, 9–18, doi:10.1016/j.abb.2011.11.003 (2012).
Ordonez-Moran, P. et al. Vitamin D and cancer: an update of in vitro and in vivo data. Front Biosci 10, 2723–2749 (2005).
Haussler, M. R. et al. The nuclear vitamin D receptor: biological and molecular regulatory properties revealed. J Bone Miner Res 13, 325–349, doi:10.1359/jbmr.1998.13.3.325 (1998).
Ylikomi, T. et al. Antiproliferative action of vitamin D. Vitam Horm 64, 357–406 (2002).
Diaz, G. D., Paraskeva, C., Thomas, M. G., Binderup, L. & Hague, A. Apoptosis is induced by the active metabolite of vitamin D3 and its analogue EB1089 in colorectal adenoma and carcinoma cells: possible implications for prevention and therapy. Cancer Res 60, 2304–2312 (2000).
Tangpricha, V. et al. 25-hydroxyvitamin D-1alpha-hydroxylase in normal and malignant colon tissue. Lancet 357, 1673–1674, doi:10.1016/S0140-6736(00)04831-5 (2001).
Bartel, D. P. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116, 281–297, doi:10.1016/S0092-8674(04)00045-5 (2004).
Langevin, S. M. & Christensen, B. C. Let-7 microRNA-binding-site polymorphism in the 3′UTR of KRAS and colorectal cancer outcome: a systematic review and meta-analysis. Cancer medicine 3, 1385–1395, doi:10.1002/cam4.279 (2014).
Saridaki, Z. et al. A let-7 microRNA-binding site polymorphism in KRAS predicts improved outcome in patients with metastatic colorectal cancer treated with salvage cetuximab/panitumumab monotherapy. Clin Cancer Res 20, 4499–4510, doi:10.1158/1078-0432.CCR-14-0348 (2014).
Yuan, Y. et al. A functional variant at the miRNA binding site in E2F1 gene is associated with risk and tumor HPV16 status of oropharynx squamous cell carcinoma. Molecular carcinogenesis 56, 1100–1106, doi:10.1002/mc.22576 (2017).
Lee, A. R., Park, J., Jung, K. J., Jee, S. H. & Kim-Yoon, S. Genetic variation rs7930 in the miR-4273-5p target site is associated with a risk of colorectal cancer. OncoTargets and therapy 9, 6885–6895, doi:10.2147/OTT.S108787 (2016).
Vidigal, V. M. et al. Genetic polymorphisms of vitamin D receptor (VDR), CYP27B1 and CYP24A1 genes and the risk of colorectal cancer. The International journal of biological markers 32, e224–e230, doi:10.5301/jbm.5000248 (2017).
Zeljic, K. et al. Vitamin D receptor, CYP27B1 and CYP24A1 genes polymorphisms association with oral cancer risk and survival. Journal of oral pathology & medicine: official publication of the International Association of Oral Pathologists and the American Academy of Oral Pathology 41, 779–787, doi:10.1111/j.1600-0714.2012.01164.x (2012).
Fuhrman, B. J. et al. Sunlight, polymorphisms of vitamin D-related genes and risk of breast cancer. Anticancer Res 33, 543–551 (2013).
Rheem, D. S., Baylink, D. J., Olafsson, S., Jackson, C. S. & Walter, M. H. Prevention of colorectal cancer with vitamin D. Scandinavian journal of gastroenterology 45, 775–784, doi:10.3109/00365521003734125 (2010).
Vosa, U., Esko, T., Kasela, S. & Annilo, T. Altered Gene Expression Associated with microRNA Binding Site Polymorphisms. PloS one 10, e0141351, doi:10.1371/journal.pone.0141351 (2015).
Matsuoka, L. Y., Ide, L., Wortsman, J., MacLaughlin, J. A. & Holick, M. F. Sunscreens suppress cutaneous vitamin D3 synthesis. J Clin Endocrinol Metab 64, 1165–1168, doi:10.1210/jcem-64-6-1165 (1987).
MacLaughlin, J. & Holick, M. F. Aging decreases the capacity of human skin to produce vitamin D3. J Clin Invest 76, 1536–1538, doi:10.1172/JCI112134 (1985).
Chattar-Cora, D., Onime, G. D., Coppa, G. F., Valentine, I. S. & Rivera, L. Anatomic, age, and sex distribution of colorectal cancer in a New York City Hispanic population. J Natl Med Assoc 90, 19–24 (1998).
Holick, M. F. Vitamin D: importance in the prevention of cancers, type 1 diabetes, heart disease, and osteoporosis. Am J Clin Nutr 79, 362–371 (2004).
Holick, M. F. Sunlight and vitamin D for bone health and prevention of autoimmune diseases, cancers, and cardiovascular disease. Am J Clin Nutr 80, 1678S–1688S (2004).
Jongbloet, P. H. Do sunlight and vitamin D reduce the likelihood of colon cancer? Time for a paradigm shift? International journal of epidemiology 35, 1359–1360, doi:10.1093/ije/dyl206 (2006).
Park, Y. et al. Dietary fiber intake and risk of colorectal cancer: a pooled analysis of prospective cohort studies. Jama 294, 2849–2857, doi:10.1001/jama.294.22.2849 (2005).
Uchida, K. et al. Dietary fiber, source foods and colorectal cancer risk: the Fukuoka Colorectal Cancer Study. Scandinavian journal of gastroenterology 45, 1223–1231, doi:10.3109/00365521.2010.492528 (2010).
Hill, M. J. Cereals, cereal fibre and colorectal cancer risk: a review of the epidemiological literature. European journal of cancer prevention: the official journal of the European Cancer Prevention Organisation (ECP) 6, 219–225 (1997).
Block, E. The chemistry of garlic and onions. Scientific American 252, 114–119 (1985).
Candela, M. et al. Inflammation and colorectal cancer, when microbiota-host mutualism breaks. World J Gastroenterol, doi:10.3748/wjg.v20.i4.908. (2014).
Colorectal cancer: red meat increases risk, fiber decreases it. Diet and lifestyle changes could prevent 64,000 cases per year in US DukeMedicine healthnews 17, 5–6 (2011).
Smolinska, K. & Paluszkiewicz, P. Risk of colorectal cancer in relation to frequency and total amount of red meat consumption. Systematic review and meta-analysis. Archives of medical science: AMS 6, 605–610, doi:10.5114/aoms.2010.14475 (2010).
Ogino, S., Chan, A. T., Fuchs, C. S. & Giovannucci, E. Molecular pathological epidemiology of colorectal neoplasia: an emerging transdisciplinary and interdisciplinary field. Gut 60, 397–411, doi:10.1136/gut.2010.217182 (2011).
Ogino, S. & Stampfer, M. Lifestyle factors and microsatellite instability in colorectal cancer: the evolving field of molecular pathological epidemiology. J Natl Cancer Inst 102, 365–367, doi:10.1093/jnci/djq031 (2010).
Fedirko, V. et al. Prediagnostic 25-hydroxyvitamin D, VDR and CASR polymorphisms, and survival in patients with colorectal cancer in western European ppulations. Cancer epidemiology, biomarkers & prevention: a publication of the American Association for Cancer Research, cosponsored by the American Society of Preventive Oncology 21, 582–593, doi:10.1158/1055-9965.EPI-11-1065 (2012).
Mezawa, H. et al. Serum vitamin D levels and survival of patients with colorectal cancer: post-hoc analysis of a prospective cohort study. BMC cancer 10, 347, doi:10.1186/1471-2407-10-347 (2010).
Ng, K. et al. Circulating 25-hydroxyvitamin d levels and survival in patients with colorectal cancer. J Clin Oncol 26, 2984–2991, doi:10.1200/JCO.2007.15.1027 (2008).
Ng, K. et al. Prospective study of predictors of vitamin D status and survival in patients with colorectal cancer. Br J Cancer 101, 916–923, doi:10.1038/sj.bjc.6605262 (2009).
Reddy, S. D., Gajula, R. P., Pakala, S. B. & Kumar, R. MicroRNAs and cancer therapy: the next wave or here to stay? Cancer Biol Ther 9, 479–482 (2010).
Landi, D. et al. Polymorphisms within micro-RNA-binding sites and risk of sporadic colorectal cancer. Carcinogenesis 29, 579–584, doi:10.1093/carcin/bgm304 (2008).
Wu, Y. Z. et al. Application of Crossover Analysis-logistic Regression in the Assessment of Gene- environmental Interactions for Colorectal Cancer. Asian Pacific journal of cancer prevention: APJCP 13, 2031–2037 (2012).
Acknowledgements
This study was supported by grants from Postdoctoral Science Foundation of China (No. 20100481026), Doctoral Program Foundation of State Education Ministry (20122307120005) and Dr. Wu Lien-teh Science Foundation of Harbin Medical University (WLD-QN1106).
Author information
Authors and Affiliations
Contributions
F.W. and Y.Z. contributed to the study design, data interpretation, study supervision, drafting the manuscript, and the acquisition of funding. Z.L and B.C. contributed to critical revision of the manuscript for important intellectual content. C.G. and Y.Y contributed to DNA extraction, genotyping, data analysis. L.Z., J.L. and H.Y. contributed to data analysis and manuscript draft. Q.C. and D.P. contributed to questionnaire, sample collection, and DNA extraction. S.L. and J.T. contributed to manuscript preparation. All authors contributed to review and revision of the manuscript.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gong, C., Long, Z., Yu, Y. et al. Dietary factors and polymorphisms in vitamin D metabolism genes: the risk and prognosis of colorectal cancer in northeast China. Sci Rep 7, 8827 (2017). https://doi.org/10.1038/s41598-017-09356-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-09356-1
This article is cited by
-
Genetic polymorphisms of CYP24A1 gene and cancer susceptibility: a meta-analysis including 40640 subjects
World Journal of Surgical Oncology (2023)
-
Vitamin D is inversely associated with Monocyte to HDL-C ratio among medical staff in Chengdu, China
BMC Endocrine Disorders (2023)
-
Association of CYP24A1 with survival and drug resistance in clinical cancer patients: a meta-analysis
BMC Cancer (2022)
-
Mendelian randomization analysis of vitamin D in the secondary prevention of hypertensive-diabetic subjects: role of facilitating blood pressure control
Genes & Nutrition (2022)
-
The role of vitamin D in ovarian cancer: epidemiology, molecular mechanism and prevention
Journal of Ovarian Research (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
