

Abstract
The role of anandamide in the development of inflammatory hyperalgesia and visceral hyperreflexia was studied in the rat urinary bladder. Animals were given intraperitoneal cyclophosphamide injection, which evokes painful hemorrhagic cystitis accompanied by increased bladder reflex activity. The vanilloid receptor 1 [transient receptor potential vanilloid 1 (TRPV1)] antagonist capsazepine, applied onto the serosal surface of bladders, significantly reduced the hyperreflexia. Mass spectrometric analysis revealed that cyclophosphamide injection significantly and persistently increased the anandamide content of bladder tissues. The increase in the anandamide content paralleled the development of reflex hyperactivity. Anandamide (1-100 μm), applied onto the serosal surface of naive bladders, increased the reflex activity in a concentration-dependent manner. Repeated anandamide applications did not produce desensitization of the response. The anandamide-evoked effect was blocked by capsazepine or by instillation of resiniferatoxin, the ultrapotent TRPV1 agonist, into the bladders 24 hr before the anandamide challenge. The cannabinoid 1 receptor antagonist SR141716A [N-piperidino-5-(4-chlorophenyl)-1-(2,4-dichlorophenyl)-4-methylpyrazole-3-carboxamide] significantly increased the potency of anandamide in enhancing bladder reflex activity in naive but not in cyclophosphamide-injected animals. Application of the fatty acid amide hydrolyze inhibitor palmitoylisopropylamine onto the serosal surface of bladders also increased the reflex activity both in naive and cyclophosphamide-injected rats. This latter effect in naive animals was blocked by capsazepine and by resiniferatoxin pretreatment. Finally, intravesical instillation of anandamide (50 μm) increased c-fos expression in the spinal cord, which was reduced by capsazepine or by resiniferatoxin pretreatment.
These results suggest that anandamide, through activating TRPV1, contributes to the development of hyperreflexia and hyperalgesia during cystitis.
Introduction
The fatty acid amide anandamide activates two receptors on nociceptive primary sensory neurons, the inhibitory cannabinoid 1 (CB1) receptor and the excitatory vanilloid receptor 1 [transient receptor potential vanilloid 1 (TRPV1)] (Devane et al., 1992; Zygmunt et al., 1999; Ahluwalia et al., 2003b; Akerman et al., 2004; Evans et al., 2004). The CB1 receptor and TRPV1 are coexpressed by a significant proportion of nociceptive primary sensory neurons, which innervate both somatic and visceral tissues (Guo et al., 1999; Ahluwalia et al., 2000; Avelino et al., 2002; MacNaughton et al., 2004). The excitatory target of anandamide, TRPV1, is a nonselective cationic channel, which is responsive to various physical and chemical activators such as heat greater than ∼43°C and vanilloids, including capsaicin, the pungent agent of the chili pepper (Caterina et al., 1997). Activation of TRPV1 results in burning pain sensation and increased reflex activity of organs such as the urinary bladder (Szolcsanyi, 1977; Ishizuka et al., 1994). The lack of development of increased heat sensitivity of inflamed tissues in TRPV1-deficient mice shows that TRPV1 is indispensable for the development of inflammatory heat hyperalgesia (Caterina et al., 2000; Davis et al., 2000). The finding that intravesical instillation of vanilloids, which results in loss of TRPV1 immunoreactivity in the bladder (Avelino et al., 2002), blocks inflammation-evoked hyperactivity and spinal c-fos overexpression further suggests that TRPV1 could also be pivotal in the development of visceral inflammatory hyperreflexia and hyperalgesia (Hunt et al., 1987; Vizzard, 2000; Dinis et al., 2004).
Although anandamide acting at the CB1 receptor reduces TRPV1-mediated excitation in primary sensory neurons (Richardson et al., 1998; Ahluwalia et al., 2003b; Akerman et al., 2004), recent data suggest that in inflammatory conditions, anandamide could contribute to the development of hyperalgesia and hyperreflexia by activating TRPV1. Although in physiological conditions, the excitatory effect of anandamide on primary sensory neurons is weak, it is significantly increased when inflammatory conditions are replicated in primary sensory neuronal cultures by activating protein kinase A (PKA), protein kinase C (PKC), or phospholipase C (PLC) (Malmberg et al., 1997; Zygmunt et al., 1999; Chuang et al., 2001; Ahluwalia et al., 2003b; Zhou et al., 2003). Moreover, Ca2+ influx into capsaicin-sensitive primary sensory neurons, which occurs in inflammation (Linhart et al., 2003), results in the production of anandamide, the amount of which is comparable with that evoking TRPV1-mediated excitation of primary sensory neurons after PKA, PKC, and PLC activation in the cells (Premkumar and Ahern, 2000; Chuang et al., 2001; De Petrocellis et al., 2001; Ahluwalia et al., 2003a,b). Anandamide, through TRPV1, evokes action potential generation in capsaicin-sensitive primary afferents (Kollarik and Undem, 2004), which results in pain sensation (Schmelz et al., 2000) and release of neuropeptides, such as substance P, that are essential for the development of inflammatory hyperalgesia and hyperreflexia (Ahluwalia et al., 1994; Lecci et al., 1994a,b; Laird et al., 2000; Hunt and Mantyh, 2001).
Our aim in the present work was to determine whether anandamide contributes to the development of inflammatory hyperreflexia and hyperalgesia. Preliminary results have been published in abstract form (Avelino et al., 2003).
Materials and Methods
Animals and drugs. All of the experiments were performed in accordance with the revised National Institutes of Health Guide for the Care and Use of Laboratory Animals, with the United Kingdom Animals (Scientific Procedures) Act (1986) and associated guidelines, and with the European Communities Council Directive (86/609/EEC). The use of animals was approved by the animal subjects review board of the University of Porto, Porto, Portugal. Furthermore, all efforts were made to minimize the number of animals used in the present study.
Adult female Wistar rats (250 gm) were obtained from the Gulbenkian Foundation (Porto, Portugal). Anandamide (soy oil:water, 1:4 emulsion), capsaicin, capsazepine, and the fatty acid amide hydrolase (FAAH) inhibitor palmitoylisopropylamide (PIA) were purchased from Tocris Cookson (Bristol, UK). Resiniferatoxin (RTX) and acrolein were purchased from Sigma (Porto, Portugal). Cyclophosphamide was purchased from ASTA Medica (Radebeul, Germany). The CB1 receptor antagonist N-piperidino-5-(4-chlorophenyl)-1-(2,4-dichlorophenyl)-4-methylpyrazole-3-carboxamide (SR141716A) was a kind gift from the Chemical Synthesis and Drug Supply Program of the National Institute of Mental Health (Bethesda, MD). The stock solution of capsaicin was made in DMSO, whereas that of cyclophosphamide was diluted in saline. Capsazepine, SR141716A, RTX, and PIA stock solutions were made in absolute ethanol. All of the stock solutions were diluted to working concentrations with saline with the exception of PIA, acrolein, and cyclophosphamide for electrophysiological studies. The stock solution of PIA was diluted in 1% ethanol in saline, whereas acrolein and cyclophosphamide in the patch-clamp recordings were diluted further with the bath solution (see below). The final concentration of soy oil was between 1:48,000 and 1:1200.
Induction of inflammation. Previous findings suggest that TRPV1 plays a pivotal role in the development of inflammatory hyperreflexia and hyperalgesia in cystitis evoked by the anticancer drug cyclophosphamide (Vizzard, 2000; Dinis et al. 2004). Thus, in the present study, we used the well established model of cyclophosphamide-induced bladder inflammation (Cox, 1979) to find out whether anandamide is responsible for the activation of TRPV1 and contributes to the development of increased bladder reflex activity and pain. Two schedules of intraperitoneal injections were used to produce cystitis: either a single injection of 200 mg/kg or three injections of 75 mg/kg each on days 1, 4, and 7. The single-injection model was used to assess bladder reflex activity 4 hr after the injection and to measure the anandamide content of the bladders 4, 24, and 72 hr after the injection. The repeated injection model was used to measure the anandamide content of the bladders 8 d after the first injection. Both treatments produced the same visual signs of inflammation. The bladders were red, swollen, and in some cases, covered by abundant serosal pethechial suffusions. In some animals, the urine contained blood. Control animals received intraperitoneal saline injection(s). These animals had healthy bladders containing normal urine.
Induced desensitization of capsaicin-sensitive bladder afferents. To induce desensitization of TRPV1-expressing bladder afferents, rats were anesthetized with isofluorane (4% for induction and 2% for maintenance), and a 26 gauge catheter was inserted into the bladder through the urethra. RTX (0.5 ml of 10 nm), which has been shown to desensitize capsaicin-sensitive afferents (Avelino et al., 1999), was then instilled into the bladder through the catheter, and the RTX was left in place for 30 min. After emptying the bladder and rinsing it with saline, animals were left to recover from anesthesia and used 24 hr later for subsequent experiments.
Cystometry and drug application. Bladder reflex activity was studied by cystometry performed under anesthesia with subcutaneous injection of 1.2 mg/kg urethane. The body temperature was monitored continuously by a rectal thermometer and maintained at 36-37°C with a heating pad. Bladders were exposed through a low midline abdominal incision, and a 21 gauge needle was inserted into the bladder dome. Bladders were then left to stabilize for 30 min. Continuous infusion of saline (6 ml/hr) at 37°C was then started while intravesical pressure was continuously monitored with a pressure transducer connected to a personal computer running the Acquire software package (Gould Instruments, Valley View, OH). Bladder contractions could expel the infused saline freely through the urethra.
A silicone plastic tube (inner diameter, 1 mm) was placed near the bladder dome, and saline at 37°C was continuously applied onto the bladder serosa at a rate of 0.5 ml/min. The number of bladder contractions occurring during a 10 min saline application was taken as the basal bladder reflex activity. Bladder reflex activity was then studied during the application of different drug solutions at 37°C, with a 0.5 ml/min rate, onto the serosal surface of the bladders. Drugs were applied for 10 min except for PIA, which was superfused for 30 min, and SR141716A, which was superfused onto inflamed bladders for 20 min. The excess fluid around the bladders was removed by constant and careful aspiration. Different concentrations of anandamide (1, 5, 10, 50, and 100 μm) were applied onto the bladders successively in a cumulative manner.
Anandamide detection in intact and inflamed bladders. Four hours (n = 4), 24 hr (n = 4), and 3 d (n = 4) after single cyclophosphamide injections, and 24 hr after the last repeated cyclophosphamide injections on the seventh day of the experiment (i.e., on day 8; n = 4), rats were anesthetized, decapitated, and the bladders were harvested immediately through laparotomy. Bladders of four naive rats and of four animals injected 4 hr previously with saline were also harvested. After removal, bladders were immediately frozen on dry ice and stored at -80°C. Frozen bladders were homogenized in water (1:5, w/v). The homogenate was extracted with 1 ml of ethyl acetate and then centrifuged at 3000 × g for 10 min. The organic phase was removed, dried under vacuum at -40°C, weighed, and redissolved in 100 μl of methanol.
Chromatographic separation of anandamide was performed at room temperature in an HPLC mass spectrometer system (HP1100 MSD; Hewlett-Packard, Palo Alto, CA) equipped with a vacuum degasser, autosampler (Hewlett-Packard), and a Phenomenex (Cheshire, UK) C18 reversed-phase column (3 μm; 300 Å; 30 × 4.6 mm). Ten microliters of the sample were injected into the system, and the flow rate was held constant at 1 ml/min. Under these conditions, the retention time of anandamide was 6.173 min. The column efflux was directly introduced into the ion source of the HP1100 MSD detector. Quantitative analysis was performed by selected ion recording over the respective protonated molecular ions. The peak area was chosen as the chromatographic signal for quantification and integrated automatically using the WinNT Chem-Station (Hewlett-Packard) software package.
Standard curves were prepared by spiking bladder homogenate samples originating from control (vehicle-treated) animals with six different concentrations of anandamide as external standard. These were then processed in the same way as the original samples. The calibration set was prepared on the day of analysis, and the standards were run before and after each analytical batch. Samples were compared with the standard curve covering the concentration range of the samples. The standard curves after extraction were linear. The unknown sample concentrations were calculated from the linear regression parameters of the standard curves and expressed in picomoles per gram of dry weight of bladders.
Whole-cell voltage-clamp recordings. Electrophysiological recordings were made from Chinese hamster ovarian (CHO) cells heterologously expressing rat TRPV1. The cell line was generated as described previously (McIntyre et al., 2001). Briefly, a 3.3 kb ratVR1 cDNA insert was prepared by removing it from the pBK-CMV plasmid with EcoR1 and NotI digestion and ligating it into pIRESneo (Clontech, Cambridge, UK). The VR1-pIRESneo construct was then transfected into a CHO host cell line by Lipofectamine Plus (Invitrogen, San Diego, CA). Cells were maintained in MEM α medium supplemented with 100 U/ml penicillin, 100 μg/ml streptomycin, 2 mm l-glutamine, 10% fetal calf serum, and 700 μg/ml geneticin.
Recordings were made using conventional whole-cell voltage-clamp technique with an Axoclamp 200B amplifier using the pClamp7 software package (Axon Instruments, Union City, CA). The composition of the bath and pipette solutions were (in mm): 140 NaCl, 5 KCl, 1 MgCl2, 2 CaCl2, 10 HEPES, 10 glucose, pH 7.4 and 140 KCl, 1 CaCl2, 2 MgATP, 10 HEPES, 10 EGTA, pH 7.4, respectively. All of the recordings were made at room temperature at a holding membrane potential of -60 mV. Cells were initially tested for capsaicin sensitivity by brief (5 sec) application of capsaicin (1 μm). Cyclophosphamide (1 nm, 1 μm, or 1 mm) or acrolein (1 nm, 1 μm, or 1 mm) was superfused to the cells in a cumulative manner. Each drug was applied for 5 sec with a 1 min interval between the different drugs-concentrations. One minute after the application of the highest concentration of acrolein or cyclophosphamide, capsaicin (1 μm) was superfused onto the cells again to see whether acrolein or cyclophosphamide had any long-lasting effect on the capsaicin-evoked response.
Spinal c-fos expression. Activation of nociceptive primary sensory neurons resulting in pain-related behavior in animals evokes enhanced c-fos expression in the spinal cord (Hunt et al., 1987). Thus, to find out whether anandamide induces nociceptive transmission to spinal dorsal horn neurons, we studied spinal c-fos expression. To avoid laparotomy-induced spinal c-fos expression, anandamide in this experiment was instilled into the bladder through the urethra. Twelve rats were anesthetized with intraperitoneal 35% aqueous chloral hydrate (0.1 ml/100 gm), and their bladders were catheterized through the urethra with a 26 gauge catheter and emptied. The bladders of four animals were instilled with 0.5 ml of a 50 μm anandamide solution, whereas another four rats received 0.5 ml of saline. To ascertain the role of TRPV1 on the anandamide-evoked effect, bladders of four animals were instilled with 0.5 ml of saline containing 50 μm anandamide and 50 μm capsazepine. In addition, four rats previously desensitized with intravesical RTX also received 0.5 ml of 50 μm anandamide. Solutions were left in place for 30 min and then withdrawn, and the bladders were rinsed with saline.
Two hours after the beginning of the drug application, rats were perfused through the ascending aorta with 100 ml of 0.1 m PBS followed by 1000 ml of 4% paraformaldehyde in 0.1 m phosphate buffer, pH 7.4. The L6 spinal segments were removed and immersed for 2 hr into the same fixative followed by immersion overnight in 30% sucrose in phosphate buffer. Spinal segments were cut by a freezing microtome into 40 μm transverse sections, and one in every three consecutive sections was immersed and stored in a cryoprotective solution at -20°C (Lu and Haber, 1992). After sectioning of spinal cords from all 16 animals, sections were washed with PBS and incubated in the same bath with a polyclonal antiserum against the Fos protein raised in rabbit (diluted at 1:6000; Oncogene Sciences, Uniondale, NY). The immunoreaction was visualized by the ABC-HRP method (Vector Elite kit; Vector Laboratories, Burlingame, CA). Fos-immunoreactive cells in the dorsal horn in 10 sections taken randomly from the spinal cord of each animal were counted.
Data presentation and statistical analysis. Bladder reflex activity was evaluated by calculating the average number of bladder contractions per minute during a 10 min saline or drug application. The relevant data obtained from four to nine animals were averaged. Mean values of the anandamide concentrations in bladders were calculated from appropriate data of four animals. Spinal c-fos expression was analyzed by calculating the mean of the number of Fos-immunoreactive cells found in 10 sections of each animal. The mean number of Fos-immunopositive cells found in the appropriate four animals was then calculated.
Differences between the effects of various drugs were studied by ANOVA. The significance of the differences between relevant data were established by the post hoc Bonferroni test. Differences were considered significant at p < 0.05. Data are expressed as mean ± SEM, and n refers to the number of animals.
Results
The effect of capsazepine on bladder reflex activity
The baseline bladder reflex activity of naive rats was 0.48 ± 0.01 (n = 4) contractions per minute (Fig. 1A,C). Capsazepine applied onto the bladders did not produce any change in the frequency of bladder contractions either at 10 μm (data not shown) or 50 μm concentrations (Fig. 1A,C).

Capsazepine (CPZ) reduces the reflex activity of inflamed bladders only. A, Typical cystometrogram from a naive bladder. The basal reflex activity was determined during a 10 min period by intravesical perfusion of saline (37°C) and superfusing 37°C saline onto the serosal surface of the bladder. Note that 50 μm capsazepine did not change the reflex activity of the noninflamed bladder. B, Typical cystometrogram from an inflamed bladder 4 hr after intraperitoneal cyclophosphamide injection. Note that the basal reflex activity measured during saline superfusion is significantly higher than that in the naive bladder shown in A. In contrast to naive bladders, capsazepine (10 or 50 μm) superfusion onto the serosal surface did reduce the reflex activity significantly. However, 50 μm capsazepine did not have greater effect than 10 μm. C, Average number of bladder contractions in naive bladders (n = 4) during saline (empty bar) and during capsazepine (50 μm; solid bar) superfusion onto the serosal surface. D, Average number of contractions in bladders from animals that received intraperitoneal cyclophosphamide injection 4 hr before the recordings (empty bar; n = 5). Both 10 μm (solid gray bar) and 50 μm (solid black bar) capsazepine reduced the enhanced reflex activity significantly. *p < 0.05. Error bars indicate SEM.
Four hours after a single cyclophosphamide injection, the frequency of bladder contractions was significantly higher than that in naive rats (1.31 ± 0.1; p = 0.0004; n = 5) (Fig. 1B,D). In contrast to the lack of any effect in naive rats, capsazepine produced a significant reduction in the bladder reflex activity in the cyclophosphamide-injected animals both at 10 μm (to 0.9 ± 0.05; p = 0.004; n = 5) and 50 μm (to 0.84 ± 0.09; p = 0.003; n = 5) (Fig. 1B,D). Although the effect of 50 μm capsazepine was somewhat greater than that produced by 10 μm capsazepine, the difference was not significant. These findings were in agreement with previous results (Vizzard 2000; Dinis et al., 2004) and indicated that TRPV1 activity contributed to the development of hyperreflexia in cyclophosphamide-induced cystitis. Furthermore, because capsazepine has been shown to compete for the capsaicin-binding site of TRPV1 (Bevan et al., 1992), these findings suggested that TRPV1 might be activated by a ligand(s) binding to that domain of the receptor.
Effect of cyclophosphamide and acrolein on TRPV1
Although neither cyclophosphamide nor its metabolite acrolein, which is excreted by urine (Cox, 1979), has structural similarity to any of the known vanilloid agonists, it is possible that these agents activate TRPV1 on bladder afferents. Thus, during a search for a probable TRPV1 activator, we examined whether cyclophosphamide or acrolein has any effect on TRPV1. Application of capsaicin (1 μm) evoked inward current (A = -0.26 nA to -6.33 nA) in all the cells tested (n = 39) (Fig. 2). In contrast, neither cyclophosphamide (n = 12) nor acrolein (n = 13) evoked any current between 1 nm and 1 mm (Fig. 2). Application of capsaicin after an acrolein or cyclophosphamide challenge evoked inward currents in all cells. In some cells, the response to capsaicin after acrolein or cyclophosphamide application was larger than that seen with the first capsaicin application. However, similar increases were observed without acrolein or cyclophosphamide application (n = 14; data not shown); therefore, these increases were unrelated to any drug treatment. These results indicated that TRPV1 in bladder afferents was probably activated not by cyclophosphamide or acrolein but by an endogenous ligand.

Whole-cell voltage-clamp recordings from TRPV1-expressing CHO cells challenged by capsaicin (caps), cyclophosphamide (top trace), and acrolein (bottom trace). Recordings were done with physiological pipette and bath solutions at Em = -60 mV, and drugs were applied for 5 sec with a 1 min interval between the applications. Capsaicin (1 μm) evoked inward current in all cells tested. However, neither cyclophosphamide nor acrolein at 1 nm, 1 μm, or 1 mm concentration produced any detectable currents. Furthermore, because 1 μm capsaicin applied after the cyclophosphamide or acrolein challenge produced similar responses to the first capsaicin application, neither cyclophosphamide nor acrolein seem to produce sensitization or desensitization of TRPV1-mediated responses. Solid bars above current traces indicate drug applications.
Anandamide content of intact and inflamed bladders
Among the known endogenous TRPV1 activators, anandamide binds to TRPV1 at the capsaicin binding site (Jordt and Julius, 2002; Gavva et al., 2004). Thus, we next investigated whether cyclophosphamide injection increased the anandamide content of the urinary bladder. Measurements were done in bladders from naive and saline-injected rats and in bladders from cyclophosphamide-injected animals 4, 24, and 72 hr after a single injection or 8 d after first of the multiple injections. The anandamide concentrations in the bladders of the naive and the saline-injected animals were similar (34.6 ± 3.9 pmol/gm of dry tissue, n = 4; 32.1 ± 2.8 pmol/gm of dry tissue, n = 4, respectively). Therefore, data from these animals were pooled together and used as control data (33.4 ± 1.4 pmol/gm of dry tissue; n = 8) (Fig. 3D). Four hours after single (200 mg/kg) cyclophosphamide injections, the anandamide level of the bladders increased significantly (45.0 ± 3.4 pmol/gm of dry tissue; p = 0.02; n = 4) (Fig. 3D). At 24 hr, the anandamide level showed no additional increase. At 72 hr, the anandamide content of the bladders reached its highest concentration (59.28 ± 7.8 pmol/gm of dry tissue; n = 4) (Fig. 3D). The anandamide level in the bladders from the animals injected repeatedly on days 1, 4, and 7 was 49.18 ± 6 pmol/gm of dry tissue (n = 4) at 24 hr after the last injection. This value was also significantly higher (p = 0.04) than that in the control, untreated bladders (Fig. 3D). These findings showed that the anandamide concentration of the bladders was persistently elevated in cyclophosphamide-injected rats.

Intraperitoneal cyclophosphamide injection increases the anandamide content of bladder tissues. A, A liquid chromatography-mass spectrometry chromatogram of a spiked control sample. The scan was set between m/z = 200 and m/z = 600. A major peak elutes at 6.173 min, giving a mass ion of 348.3, which represents the protonated molecular ion with 100% abundance (arrow). The molecular weight of anandamide is 347.3. B, A liquid chromatography-mass spectrometry chromatogram of a sample from a naive bladder. Note that the size of the peak representing anandamide is negligible (arrow). C, A liquid chromatography-mass spectrometry chromatogram of a sample from an inflamed bladder. Note that the peak representing anandamide (arrow) is significantly larger that that in B. D, Average anandamide content of control and inflamed bladders at different time points after intraperitoneal cyclophosphamide injection. Note that the anandamide content of inflamed bladders is significantly increased at either of the time points in comparison with that of naive bladders. *p < 0.05. Error bars indicate SEM.
The effect of exogenous anandamide on the micturition reflex activity
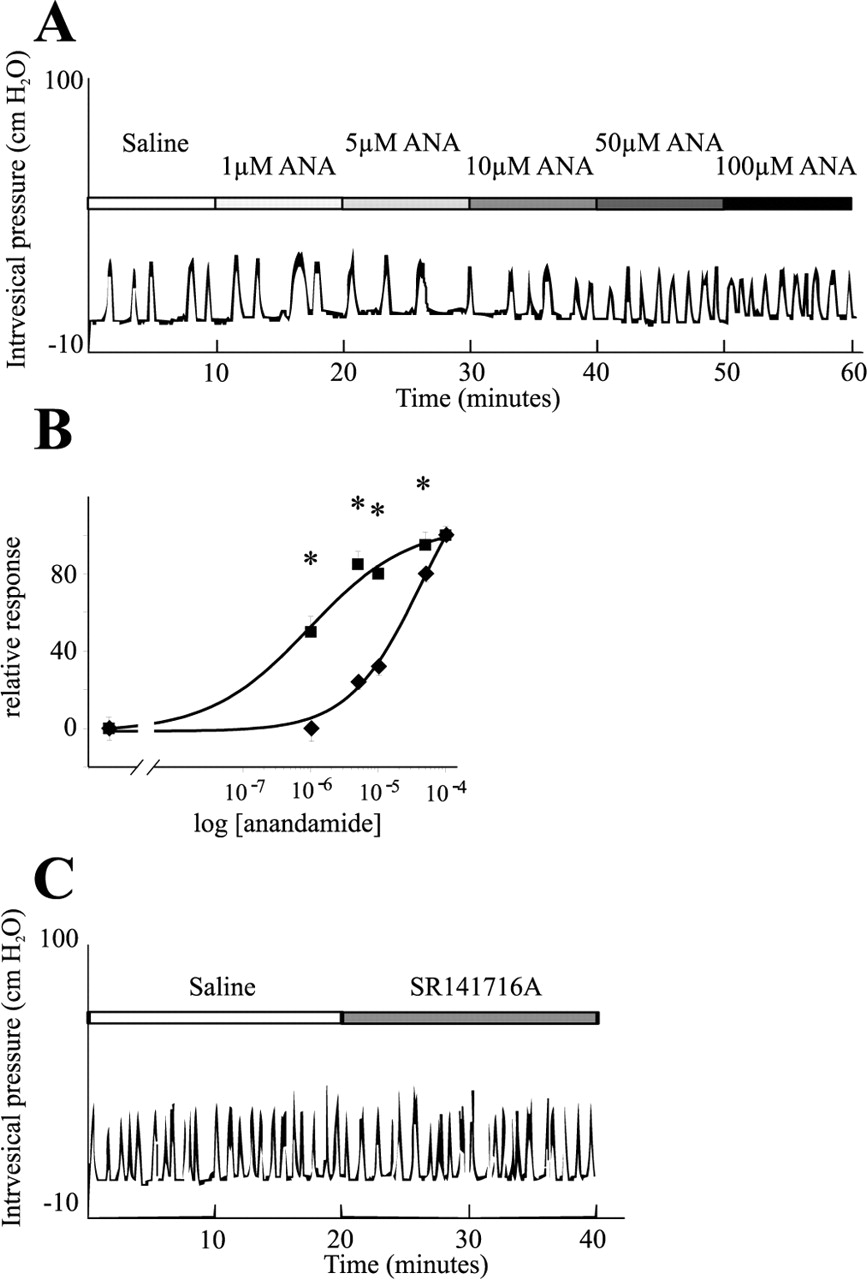
To find out whether the increased anandamide content could be responsible for the increased reflex activity of the bladders observed during cystitis, exogenous anandamide was applied onto the serosal surface of naive bladders, while the intravesical pressure was monitored by cystometry. The basal bladder reflex activity during a 10 min saline application was 0.51 ± 0.09 contractions per minute (n = 8). Application of the anandamide vehicle (soy oil in saline between 1:1200 and 1:48,000 dilutions) did not produce any effect on the bladder reflex activity (data not shown). Application of anandamide (1100 μm), however, produced a concentration-dependent increase in the number of contractions (Fig. 4A,B). The lowest concentration of anandamide producing a significant increase in the frequency of contractions was 50 μm (0.73 ± 0.03 contractions/min; p = 0.0001; n = 8). One hundred micromolar anandamide produced an additional increase in the activity (0.79 ± 0.03 contractions/min; p = 0.000002; n = 8). This value, however, was not significantly different from that produced by 50 μm anandamide. The EC50 of anandamide was 17.8 ± 2 μm (n = 8). The effect of anandamide on the reflex activity could be observed within 1 min after the drug reached the surface of the bladders, and the activity remained constant during the 10 min application.
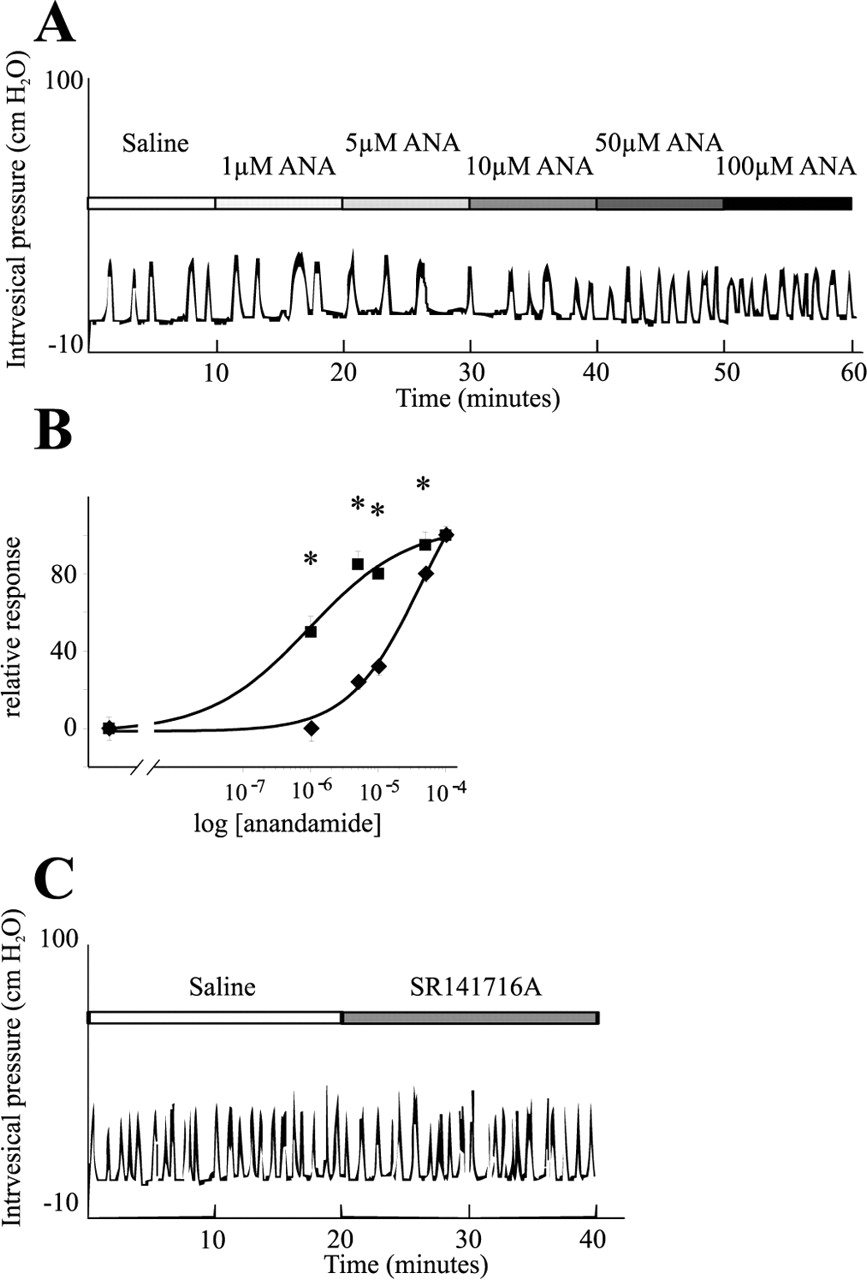
Effect of exogenous anandamide (ANA) on the reflex activity of naive bladders. A, A typical cystometrogram during cumulative application of anandamide (1-100 μm) onto the serosal surface of a bladder. Note that only 50 and 100 μm anandamide increased the bladder reflex activity. B, Dose-response relationship of anandamide on naive bladders in the absence (♦) or in the presence (▪) of SR141716A, a cannabinoid 1 receptor antagonist. Note that the CB1 receptor antagonist significantly increased the potency of anandamide on the bladder. C, Whereas exogenous anandamide, in addition to TRPV1, seems to activate the CB1 receptor in naive bladders, endogenous anandamide during inflammation apparently does not induce any activity of the inhibitory receptor; 20 min superfusion of the CB1 receptor antagonist failed to increase the reflex activity of bladders from cyclophosphamide-injected rats. *p < 0.05.
Prolonged application of TRPV1 agonists induces desensitization of TRPV1 (Szallasi, 2002). As shown in Figure 5A, the first application of 50 nm RTX onto the bladder surface induced a significant increase in the number of contractions (0.5 contractions/min before and 0.9 contractions/min during RTX application), whereas the second RTX application failed to increase the bladder activity (0.3 contractions/min during the 10 min wash before the second RTX application and 0.3 contractions/min during the second RTX application).

Whereas vanilloids, such as capsaicin or RTX, do induce desensitization, anandamide (ANA) does not induce desensitization of the reflex activity of the urinary bladder. A, First application of RTX onto the serosal surface of naive bladders increases the reflex activity. However, the second RTX activation fails to increase the number of contractions. B, A typical cystometrogram on a naive bladder. Both the first and the second anandamide superfusion enhanced the reflex activity significantly. Note that the increase in the reflex activity was similar during the first and second anandamide application. C, Average response of naive bladders to 50 μm anandamide (solid bars; n = 4). Empty bars represent the number of contractions per minute during saline superfusion onto the serosal surface of bladders. *p < 0.05. Error bars indicate SEM.
To find out whether anandamide had a similar desensitizing effect, a 10 min saline and 10 min anandamide (50 μm) application was followed by a 30 min wash with saline and another 10 min 50 μm anandamide application. The first anandamide application increased the number of bladder contractions from 0.55 ± 0.06 to 0.85 ± 0.1 contractions/min (n = 4) (Fig. 5B,C). This value was similar to that found when 50 μm anandamide was applied in a cumulative manner and was significantly different from the basal value (p = 0.002). The frequency of the bladder reflex contractions decreased gradually during the saline wash period and by 20 min reached a stable value (0.4 ± 0.14 contractions/min; n = 4) (Fig. 5B,C), which was not significantly different from the basal activity of the bladders (p = 0.1). During the second anandamide application, the frequency of bladder contractions increased to 0.90 ± 0.14 contractions/min (n = 4) (Fig. 5B,C), which was significantly higher than the basal value (p = 0.005) and similar to that observed during the first anandamide application (p = 0.1).
To find out whether the anandamide-evoked increase in the bladder reflex activity was mediated through TRPV1, the basal bladder activity was determined in four rats, and 50 μm anandamide was applied onto the bladders for 10 min. Anandamide application increased the bladder activity from 0.48 ± 0.01 to 0.82 ± 0.02 contractions/min (p = 0.00003; n = 4) (Fig. 6A). This application was followed by saline wash for 30 min, which reduced the number of bladder contractions to the control values (0.47 ± 0.16 contractions/min). Next, bladders were exposed to 50 μm capsazepine for 30 min. In agreement with our previous observations, capsazepine did not change the bladder reflex activity (data not shown). Then, 50 μm anandamide together with capsazepine (50 μm) was applied onto the bladders. In the presence of capsazepine, anandamide failed to produce any increase in the frequency of bladder contractions (0.41 ± 0.03 contractions/min; p = 0.1; n = 4) (Fig. 6A).

Anandamide (ANA) induces reflex activity in the bladder through TRPV1. A, Average response of naive bladders to anandamide (50 μm) in the absence (solid black bar) or presence (50 μm; solid gray bar) of capsazepine (CPZ). Capsazepine completely blocked the anandamide-induced reflex activity (n = 4). B, Desensitization of capsaicin-sensitive bladder afferents by intravesical RTX instillation also completely blocked the 50 μm anandamide-evoked increase in bladder reflex activity (solid bar; n = 4). Empty bars in both A and B represent the number of contractions during saline application. **p < 0.05. Error bars indicate SEM.
To further establish the involvement of TRPV1 in anandamide-induced bladder hyperreflexia, we applied 50 μm anandamide onto bladders that had been desensitized with intravesical RTX instillation. The basal activity of the RTX-pretreated bladders (0.5 ± 0.07 contractions/min; n = 4) (Fig. 6B) was not different from that of the naive animals. In contrast to the results with naive rats, 50 μm anandamide failed to produce any increase in the number of reflex contractions of these bladders (0.48 ± 0.07 contractions/min; n = 4) (Fig. 6B). Together, these findings indicated that exogenous anandamide increased the bladder reflex activity through TRPV1 and that anandamide did not produce long-term desensitization.
The effect of the CB1 receptor antagonist SR1411716A on the exogenous anandamide-evoked increase of reflex activity in naive bladders
Previous data show that TRPV1 and the CB1 receptor are coexpressed in some primary sensory neurons and visceral afferents (Ahluwalia et al., 2000, 2003b; MacNaughton et al., 2004). To find out whether in addition to TRPV1 activation, anandamide also activated the CB1 receptor in bladder afferents, the CB1 receptor antagonist SR141716A (50 μm) was applied onto the bladders for 10 min after determining the basal bladder reflex activity. Anandamide (1-100 μm) was then applied together with SR141716A (50 μm) in a cumulative manner. The baseline bladder reflex activity in these experiments was 0.58 ± 0.03 contractions/min (n = 8) (Fig. 4B). Administration of SR141716A alone had no effect on the bladder reflex activity (0.6 ± 0.04 contractions/min; n = 8). However, the anandamide-evoked bladder activity was significantly enhanced by the CB1 receptor antagonist (Fig. 4B). The lowest concentration of anandamide producing a significant increase in the reflex activity in the presence of SR141716A was 1 μm (0.69 ± 0.05 contractions/min; p = 0.006; n = 8) (Fig. 4B). Five micromolar anandamide produced an additional increase in the bladder reflex activity (0.77 ± 0.05; p = 0.0003; n = 8) (Fig. 4B). This value, however, was not significantly different from that produced by 1 μm (p = 0.1). No additional significant increase in the reflex activity could be evoked with 10, 50, or 100 μm anandamide (Fig. 4B). Comparison of the anandamide concentration-response curves on the bladder reflex activity in the absence or presence of the CB1 receptor antagonist revealed that SR141716A reduced the EC50 of anandamide from 17.8 ± 2 to 1.8 ± 0.6 μm (n = 8). The difference between the EC50 values was significant (p = 0.001). These data clearly demonstrated that in addition to an action on TRPV1, anandamide also activated the CB1 receptor in the naive bladder, which reduces contractility.
The effect of the CB1 receptor antagonist SR1411716A on reflex activity in inflamed bladders
To find out whether anandamide produced during inflammation, or any other endogenous CB1 receptor ligand activated the CB1 receptor in inflamed bladders, we also superfused SR141716A to the bladders of three cyclophosphamide-injected rats. The average reflex activity of the bladders during saline application was 0.95 ± 0.06 contractions/min (n = 3) (Fig. 4C). Twenty minute superfusion of SR141716A did not change the reflex activity of the bladder significantly (0.93 ± 0.05 contractions/min; n = 3) (Fig. 4C). These findings indicated that the CB1 receptor was not activated by endogenous anandamide or other CB1 receptor agonist in inflamed bladders.
The effect of inhibition of anandamide hydrolysis on reflex activity in naive bladders
To find out whether endogenous anandamide could also increase the number of bladder reflex contractions, the FAAH inhibitor PIA (30 μm) was applied onto the serosal surface of naive bladders. After establishing the basal reflex activity in five animals (0.42 ± 0.04 contractions/min) (Fig. 7A,B), PIA was applied for 30 min. Fifteen minutes after the beginning of PIA application, the frequency of bladder reflex contractions increased to 0.66 ± 0.04 contractions/min (p = 0.002; n = 6) (Fig. 7A,B), which remained unchanged until the end of the drug administration. Washing the surface of the bladder with saline reduced the frequency of the reflex contractions to the basal value (data not shown).

Blocking of anandamide hydrolysis with the fatty acid amide hydrolyze inhibitor PIA enhances the reflex activity both of naive bladders and inflamed bladders. A, A typical cystometrogram showing the effect of PIA on a naive bladder. Note that by about the 20th minute of PIA application, the reflex activity is increased and became stable. B, Average responses of naive bladders to PIA application (solid bar; n = 4). The PIA-evoked increase in the reflex activity was significant. C, Capsazepine (CPZ; 50 μm) prevented the PIA-induced increase in bladder reflex activity (solid bar). Empty bars in both B and C show the number of contractions during saline application. D, PIA application also increased the activity of two of four inflamed bladders. One bladder became tetanic during PIA application. **p < 0.05. Error bars indicate SEM.
To find out whether the PIA-evoked effect was mediated by TRPV1, 50 μm capsazepine was applied onto the bladders for 10 min followed by a 30 min coapplication of 50 μm capsazepine and 30 μm PIA. The basal activity of these bladders was 0.38 ± 0.03 contractions/min (n = 5) (Figure 7C). This value was not changed by capsazepine application (data not shown). In the presence of the TRPV1 antagonist, PIA failed to increase the number of reflex contractions of the bladder (0.35 ± 0.04 contractions/min; p = 0.1; n = 5) (Fig. 7C).
The effect of inhibition of anandamide hydrolysis on reflex activity in inflamed bladders
To find out whether elevating the level of endogenous anandamide by inhibiting its hydrolysis increases the reflex activity in inflamed bladders, we also applied PIA onto the serosal surface of bladders from four cyclophosphamide-injected rats. Soon after the beginning of PIA superfusion, the elevated activity seen in two of the four bladders (0.86 and 1.1 contractions/min) was further increased. One of these two bladders became tetanic during PIA application (from 1.1 contractions/min) (Fig. 7D), whereas the activity of the other increased from 0.86 to 1.2 contractions/min. The frequency of contractions in the other two bladders before PIA application was already 1.3 contractions/min, which seems to be the highest number of contractions possible in the model we used. PIA did not evoke any change in the activity of these bladders.
The effect of anandamide instillation on spinal c-fos expression
Immunohistochemical staining of the L6 spinal cord revealed that saline instillation into the bladder induced c-fos expression in 51.5 ± 7.8 cells per spinal cord section (n = 4) (Fig. 8A,C). Positive cells were distributed by three main areas, lamina I, intermediolateral gray matter, and dorsal commissure. Instillation of 50 μm anandamide significantly increased the number of Fos-positive neurons to 79.5 ± 4.7 cells per section (p = 0.003; n = 4) (Fig. 8B,C). The spinal distribution was similar to that found in saline-instilled animals (Fig. 8B). Instillation of capsazepine (50 μm) together with anandamide (50 μm) completely prevented the anandamide-evoked increase in c-fos expression (51.2 ± 5.3; p = 0.5; n = 4) (Fig. 8C). Anandamide also failed to increase the number of Fos-immunoreactive neurons when it was instilled into the bladders of rats pretreated with RTX (Fig. 8C).

Intravesical instillation of anandamide (ANA) significantly increases spinal c-fos expression at the L6 segment. A, Representative section obtained 2 hr after instillation of saline. B, Representative section obtained 2 hr after 50 μm anandamide. C, The anandamide-evoked effect was mediated by TRPV1 because either coapplication of 50 μm capsazepine (CPZ) or intravesical RTX instillation 24 hr before the anandamide instillation prevented the anandamide-evoked increase in the number of Fos-immunopositive cells. Scale bar, 100 μm. *p < 0.05. Error bars indicate SEM.
Discussion
In agreement with previous results, we found in the present study that intraperitoneal cyclophosphamide injection evoked hemorrhagic cystitis, accompanied by increased bladder reflex activity and spinal c-fos expression (Lecci et al., 1994b; Vizzard, 2000; Dinis et al., 2004). Our recent data that desensitization of capsaicin-sensitive bladder afferents by intravesical RTX instillation blocked both the increased spinal c-fos expression and the enhanced bladder activity in cyclophosphamide-injected rats (Dinis et al., 2004) indicate that capsaicin-sensitive bladder afferents are involved in the development of cyclophosphamide-evoked bladder hyperactivity and hyperalgesia. The present finding that capsazepine significantly reduced the cyclophosphamide-evoked bladder hyperreflexia indicates that TRPV1 activity contributes to the activation of capsaicin-sensitive bladder afferents in this cystitis model.
Capsazepine competes for the capsaicin binding site of TRPV1 (Bevan et al. 1992). Thus, during cyclophosphamide-induced cystitis, a compound(s) binding to that TRPV1 domain could be present in the bladder. Cyclophosphamide applied systemically induces cystitis through its metabolite, acrolein, that is excreted and thereby accumulates in the urine (Cox, 1979). Thus, TRPV1 could be activated by either of these compounds. However, our recordings show that neither cyclophosphamide nor acrolein evokes any current in TRPV1-expressing cells. The alternative possibility for TRPV1 activation after cyclophosphamide injection is that cyclophosphamide and/or acrolein induce the production of an endogenous TRPV1 ligand(s), which binds to the capsaicin-binding site of bladder afferents. Because previous findings suggested that anandamide could be produced in inflammatory conditions (Ahluwalia et al., 2003a; Linhart et al., 2003) and that anandamide seems to bind to the capsaicin-binding site (Jordt and Julius, 2002; Gavva et al., 2004), we studied the amount of this compound in bladder tissues. The finding that both the anandamide content of the tissues and the bladder reflex activity was significantly increased 4 hr after the cyclophosphamide injection suggests that anandamide and bladder hyperreflexia could be causally related in cystitis. Although we did not measure bladder reflex activity at time points other than 4 hr after injection, the findings that the anandamide content of the bladder tissues remained elevated up to 8 d after the first of the repeated cyclophosphamide injections and that prolonged cyclophosphamide-induced cystitis is also accompanied by bladder hyperreflexia (Lecci et al.,1994b; Dinis et al., 2004) support the assumed causal relationship between anandamide and inflammatory hyperreflexia.
If anandamide were responsible for the activation of TRPV1 in cyclophosphamide-induced cystitis, then it should replicate the hyperactivity and pain occurring in inflammation. Exogenous anandamide indeed increased the reflex activity and enhanced the spinal c-fos expression that has been used to indicate the transmission of nociceptive information to dorsal horn neurons (Hunt et al., 1987). However, the magnitude of the anandamide-evoked increase in the reflex activity and c-fos expression failed to reach those observed in cyclophosphamide-induced cystitis. The lowest concentration of anandamide that increased the bladder reflex activity significantly was 50 μm, which was higher than the minimal concentration of this compound activating TRPV1 in vitro (Zygmunt et al., 1999; Smart et al., 2000; Roberts et al., 2002; Ahluwalia et al., 2003b; Price et al., 2004). However, because of the diluting effects of the tissue fluids, direct comparisons of the concentrations used in vitro and in the present study are impossible. Nevertheless, the finding that both capsazepine and RTX pretreatment blocked the anandamide-evoked increase both in the bladder activity and spinal c-fos expression suggests that both of these effects were mediated through TRPV1. The possibility that anandamide activated capsaicin-sensitive fibers through other mechanisms, such as activation of 5-hydroxytryptamine or NMDA receptors (Hampson et al., 1998; Kimura et al., 1998), seems unlikely because capsazepine probably cannot bind to these receptors (Bevan et al., 1992; Walpole et al., 1994).
Palmitoylisopropylamine also increased the reflex activity of naive bladders. This effect was inhibited by capsazepine, indicating that it was mediated by an endovanilloid(s) binding to the capsaicin-binding domain of TRPV1. Although we did not measure the anandamide content of bladders after PIA application, the finding that PIA inhibits the anandamide-hydrolyzing enzyme FAAH (Jonsson et al., 2001) suggests that that endovanilloid was anandamide. In agreement with the assumed role of anandamide in the development of inflammatory bladder hyperreflexia and the inhibitory action of PIA on FAAH, PIA also increased the activity of two of four inflamed bladders. The lack of effect of PIA on the other two inflamed bladders could be attributable to the apparent maximal activity of bladders before PIA application.
Repeated or long-lasting exposure of TRPV1 to vanilloids evokes TRPV1 desensitization, which results in unresponsiveness of bladders to repeated vanilloid agonist challenge (Szallasi, 2002). Anandamide can be responsible for the development of TRPV1-mediated inflammatory hyperalgesia and hyperreflexia only if it does not desensitize TRPV1. The findings that successive anandamide application increased the frequency of the reflex contractions to the same level and that the increase in frequency was maintained with prolonged anandamide application are in agreement with previous findings (Premkumar and Ahern, 2000; Ahluwalia et al., 2003b) that anandamide lacks desensitizing effects on TRPV1. This difference between capsaicin or RTX and anandamide might be attributable to different allosteric modifications in TRPV1 by the different ligands.
The CB1 receptor antagonist SR141716A significantly enhanced the excitatory potency of anandamide in naive bladders, indicating that in addition to TRPV1-mediated excitation, anandamide also evokes CB1 receptor-mediated inhibition in the bladder. The underlying mechanism of the SR141716A-evoked effect is not clear. However, it is an appealing possibility that the SR141716A-evoked effect was produced by blocking the CB1 receptor activation-evoked inhibition on bladder afferents coexpressing the CB1 receptor and TRPV1. This assumption is supported by previous data that a significant proportion of TRPV1-expressing primary sensory neurons express the CB1 receptor and that anandamide evokes both TRPV1-mediated excitation and CB1 receptor-mediated inhibition in those cells (Ahluwalia et al., 2000, 2003b). The recent data of MacNaughton et al. (2004) that TRPV1 and the CB1 receptor are coexpressed by paravascular nerves and nerve fibers in submucosal ganglia in the ileum also support this assumption.
Interestingly, however, the CB1 receptor antagonist had no effect on inflamed bladders, suggesting that inflammatory processes might inhibit the effects of CB1 receptor agonists. Previous data support this assumption, because both PKA and PKC, which are activated in inflammation (Malmberg et al., 1997; Zhou et al., 2003), induce phosphorylation of the CB1 receptor, which results in reduced CB1 receptor activity (Garcia et al., 1998; Huang et al., 2002). Because both PKA and PKC increase both the potency and efficacy of anandamide on TRPV1 (Premkumar and Ahern, 2000; De Petrocellis et al., 2001), anandamide could excite capsaicin-sensitive bladder afferents in inflammatory conditions at a significantly lower concentration than we observed in the present study on naive bladders.
Recent findings that Ca2+ influx into capsaicin-sensitive primary sensory neurons results in anandamide production indicate that in addition to macrophages and vascular endothelial cells, capsaicin-sensitive bladder afferents could also be the source of anandamide activating TRPV1 in cystitis (Di Marzo et al., 1996; Kuwae et al., 1999; Zygmunt et al., 1999; Ahluwalia et al., 2003a). Acrolein evokes the production of inflammatory mediators in the bladder (Ahluwalia et al., 1994), which induce Ca2+ influx into primary sensory neurons (Linhart et al., 2003) and thus probably stimulate anandamide production in bladder afferents. However, increased anandamide concentration in the bladder could also be attributable to decreased anandamide hydrolysis by FAAH in bladder tissues. Whatever the underlying mechanism, the inflammation-induced increase in the anandamide content of the whole bladder was modest. Nevertheless, as mentioned, the concentration of anandamide produced by capsaicin-sensitive primary sensory neurons after Ca2+ influx is comparable with that evoking TRPV1-mediated excitation when inflammatory conditions are simulated in cultures (Ahluwalia et al., 2003a,b). Thus, if anandamide production in capsaicin-sensitive bladder afferents were responsible for the increased anandamide content in bladder tissues in cystitis, the local concentration of anandamide at TRPV1 could be high enough to open the ion channel.
In summary, our present findings show that (1) the anandamide content is increased in the bladder during cyclophosphamide-induced cystitis, (2) exogenous anandamide increases bladder reflex activity and spinal c-fos expression in a TRPV1-dependent manner, (3) anandamide does not induce desensitization of TRPV1, (4) endogenous anandamide enhances bladder reflex activity in a TRPV1-dependent manner, and (5) both the exogenous and endogenous anandamide-evoked effects resemble to those occurring in cystitis. These findings suggest that anandamide contributes to TRPV1 activation and to the development of pain and hyperactivity of the bladder in inflammation. Because at present no drug is available to either block the synthesis (Schmid, 2000) or increase the degradation of endogenous anandamide (Giang and Cravatt, 1997), ultimate proof for the role of anandamide in the development of inflammatory bladder pain and hyperreflexia cannot be provided. Nevertheless, recent findings show that ileitis is also accompanied by increased anandamide production and that exogenous anandamide evokes ileitis and plasma extravasation in the intestine by a TRPV1-dependent mechanism (McVey et al., 2003). Although, McVey et al. (2003) did not study the effect of anandamide on intestinal motility and hyperalgesia, these findings strongly support the putative role of anandamide in the development of visceral inflammatory hyperreflexia and hyperalgesia.
Footnotes
This work was funded by the Fundação Ciência e Tecnologia Project Programa Operacional Ciência, Tecnologia, Inovação 32466/Neurociências e Sistemas Endócrinos 2000 and by the Fonds Européen de Développement Régional.
Correspondence should be addressed to Prof. Francisco Cruz, Department of Urology, Hospital São João, Alameda Hernāni Monteiro, 4200-319 Porto, Portugal. E-mail: cruzfjmr{at}med.up.pt.
Copyright © 2004 Society for Neuroscience 0270-6474/04/2411253-11$15.00/0
↵* P.D. and A.C. contributed equally to this work.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}