 Daniel Hurley
Daniel Hurley Matthew P. McCusker
Matthew P. McCusker Séamus Fanning
Séamus Fanning Marta Martins
Marta Martins- School of Public Health, Physiotherapy and Population Science, UCD Centre for Food Safety, UCD Centre for Molecular Innovation and Drug Discovery, University College Dublin, Dublin, Ireland
Salmonella enterica (S. enterica) are Gram-negative bacteria that can invade a broad range of hosts causing both acute and chronic infections. This phenotype is related to its ability to replicate and persist within non-phagocytic host epithelial cells as well as phagocytic dendritic cells and macrophages of the innate immune system. Infection with S. enterica manifests itself through a broad range of clinical symptoms and can result in asymptomatic carriage, gastroenteritis, systemic disease such as typhoid fever and in severe cases, death (1). Exposure to S. enterica serovars Typhi and Paratyphi exhibits clinical symptoms including diarrhea, fatigue, fever, and temperature fluctuations. Other serovars such as the non-typhoidal Salmonella (NTS), of which there are over 2,500, are commonly contracted as, but not limited to, food-borne sources causing gastrointestinal symptoms, which include diarrhea and vomiting. The availability of complete genome sequences for many S. enterica serovars has facilitated research into the genetic determinants of virulence for this pathogen. This work has led to the identification of important bacterial components, including flagella, type III secretion systems, lipopolysaccharides, and Salmonella pathogenicity islands, all of which support the intracellular life cycle of S. enterica. Studies focusing on the host–pathogen interaction have provided insights into receptor activation of the innate immune system. Therefore, characterizing the host–S. enterica interaction is critical to understand the pathogenicity of the bacteria in a clinically relevant context. This review outlines salmonellosis and the clinical manifestations between typhoidal and NTS infections as well as discussing the host immune response to infection and the models that are being used to elucidate the mechanisms involved in Salmonella pathogenicity.
Introduction
Every year, thousands of cases of salmonellosis are reported worldwide. However, the actual number of infections may be very different and many times greater than expected since many milder cases are not diagnosed or reported (http://www.cdc.gov/salmonella). Salmonella infection or the disease associated with it, salmonellosis, is most often characterized by enteritis. However, host restricted serotypes tend to induce higher levels of bacteremia, while some human restricted serotypes cause a systemic disease that is characterized by mild symptoms (2). Children are the most likely group of individuals to present salmonellosis. The rate of diagnosed infections in children <5 years old is higher than the rate diagnosed in all other persons. Other groups of risk, such as the elderly and immunocompromised individuals are the most likely to present severe forms of the disease.
Persons with diarrhea usually recover completely after a few days of the initial infection, although it may be several months before their bowel habits return to normal. Contrary to this could be a small number of persons with Salmonella infections that develop pain in their joints, irritation of the eyes, and painful urination. Taken together, these symptoms indicate a disease called reactive arthritis. This disease can last for months or years, and can lead to chronic arthritis, which is extremely difficult to treat. Antibiotic treatment does not make a difference in whether or not the person develops arthritis (3). Other types of invasive infections caused by Salmonella, such as bacteremia, osteomyelitis, and meningitis, may also occur and in these cases may require antimicrobial therapy (4).
The continuous evolution of Salmonella at the genetic and genomic levels contributes to the increased virulence and resistance to multiple antibiotics, leading to a phenotype of multidrug resistance. This resistance is a significant public health concern (5). Two major changes in the epidemiology of non-typhoidal salmonellosis have occurred in the last century. These were the emergence of food-borne human infections caused by Salmonella enterica Enteriditis and by multidrug-resistant strains of Salmonella enterica Typhimurium. In this century, a concerning situation is the increased resistance that non-typhoidal Salmonella (NTS) presents to fluoroquinolones and third-generation cephalosporins. Clinical isolates showing carbapenem resistance have also being reported (4). In terms of therapy, treatment with antibiotics is not usually recommended for uncomplicated Salmonella gastroenteritis. However, recent studies indicated that a 3–5 days therapy with ceftriaxone for patients with severe gastroenteritis could lead to a faster recovery. A continuous surveillance scheme of Salmonella infections in both humans and animals is of importance. A better understanding of the mechanisms that can lead to the emergence of antimicrobial resistance in Salmonella may help develop better interventional strategies that can ultimately reduce the spread of resistant Salmonella between humans and reservoirs identified (or not) along the food chain.
Due to the importance of Salmonella in the clinical and public health setting, there has been a significant effort to deepen the knowledge about pathogenic determinants of this bacterium. The clinical relevance of the disease, associated with the advances on the molecular tools available to study Salmonella and the development of suitable animal models, have lead to the development of optimal conditions to drive the scientific community to generate a large expansion of our knowledge about the pathogenesis of Salmonella-induced enterocolitis (6). This research effort has also generated an increased amount of information on the host immune mechanisms that complements gaps that still exist in fundamental research developed in this area.
The goal of this review is to discuss salmonellosis, the clinical signs caused by Salmonella infections, and the advances in our knowledge on the innate intestinal immunity. Additionally, the interaction with the host and the models used to elucidate the mechanisms triggered by the interaction of Salmonella with the host will also be discussed.
Interactions of Salmonella with the Gut Microbiome
The intestinal microbiome, which is host to an estimated 1 × 1014 bacteria, is responsible for conferring numerous aspects of the host response against salmonellosis (7). As many as 1,000 species of bacteria inhabit this niche, with the majority being classified as Gram-positive Actinobacteria and Firmicutes as well as Gram-negative bacteroides (8). A healthy gut microbiome provides protection against epithelial cell invasion via a series of strategies including the production of toxic metabolites, which have been shown to repress the expression of Salmonella virulence genes among others. This feature assists in the clearance of pathogens from the gut lumen after NTS-induced diarrhea (7). Increased fecal shedding and establishment of carrier status is commonly associated with prolonged treatment with antimicrobial compounds as these can have adverse effects on the composition of the gut microbiome of an individual (8, 9). This depletion of the natural gut microbiome may have long lasting effects and can result in an increased susceptibility to Salmonella colonization. One such example of this scenario is S. Typhimurium, which takes advantage of the availability of ethanolamine, a nutrient present in the microbiome, to gain a significant growth advantage in the intestine during inflammation over potential competing pathogens. S. Typhimurium-encoded virulence factors have been shown to induce the production of an alternate electron acceptor by the host, which supports anaerobic respiration and enables S. Typhimurium to outcompete other fermenting gut microbes sharing the same ecological niche (10).
Salmonellosis
Salmonellosis causes significant morbidity and mortality on a global scale and occurs after the ingestion of food or water sources that have been previously contaminated by the fecal or urinary excretions of animals that can act as reservoirs of Salmonella (11). Following infection with Salmonella species, a broad range of clinical manifestations can be presented in a number of ways depending on the susceptibility of the host (12, 13). These include bacteremia, enteric fever, enterocolitis, and chronic asymptomatic carriage. Typhoid and Paratyphoid fever, collectively termed enteric fever, are contracted following infection with S. enterica serovars Typhi (S. Typhi) and Paratyphi (S. Paratyphi), respectively. In contrast, gastroenteritis is commonly associated with NTS serovars such as Typhimurium (S. Typhimurium) and Enteritidis (S. Enteritidis).
In human beings, S. Typhi and S. Paratyphi cause typhoid fever, a bacteremic illness, which presents in a unique manner when compared with other Gram-negative bacteremias (14, 15). S. Typhi has previously adapted to infect human hosts whereas other serovars have retained a broad host preference and are capable of infecting a range of animals causing enterocolitis (16). Serovars of S. enterica including Choleraesuis (S. Choleraesuis), Dublin (S. Dublin), and S. Typhimurium can successfully infect both human and animal hosts. However, the infection presents differently in each. Human infection with S. Choleraesuis and S. Dublin commonly results in bacteremia. In mice, S. Typhimurium causes symptoms similar to human typhoid fever and will disseminate throughout the body of the host causing systemic illness (17, 18). Systemic infection can result in a diverse range of clinical manifestations that include bradycardia, hepatomegaly, and splenomegaly. Bacterial emboli form skin lesions known as Rose spots that occur in approximately 30% of typhoid fever cases. NTS serovars cause self-limiting diarrhea and in rare cases, secondary bacteremia. Primary NTS bacteremia has also been reported in immunocompromised hosts (19, 20). Death from salmonellosis can be caused by perforation of the gut and necrosis of Peyer’s patches leading to peritonitis or toxic encephalopathy [H. (15)].
Salmonella Species and Subspecies
Salmonella enterica are Gram-negative facultative intracellular anaerobes that can invade a broad range of hosts causing both acute and chronic infections by means of their ability to replicate and persist within non-phagocytic epithelial cells as well as phagocytic dendritic cells and macrophages of the host innate immune system (21, 22). The genus Salmonella comprises two species, S. enterica and S. bongori (also referred to as subsp. V). The former is further divided into six subspecies (as shown in Figure 1), which are biochemically differentiated into serovars based on the composition of their carbohydrate, flagellar, and lipopolysaccharide (LPS) structures. All Salmonella serotypes can be designated by an antigenic formula based on somatic (O) and flagellar (H) antigens in addition to capsular (Vi) antigens (16).
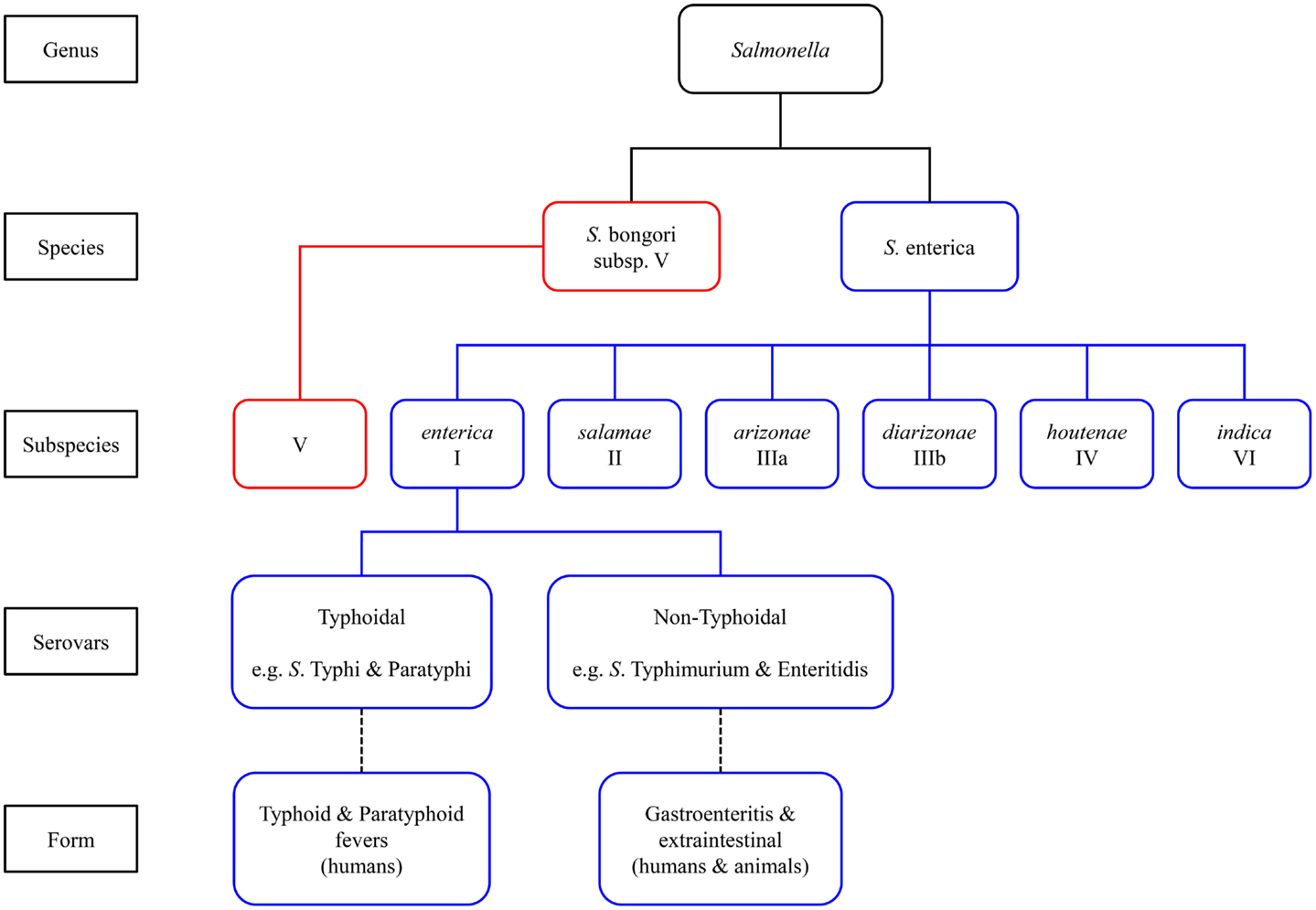
Figure 1. Classification of Salmonella species and subspecies.
Salmonella Pathogenicity Islands
Using ex vivo and in vivo animal models of infection, many virulence factors have been determined, which are responsible for inducing an inflammatory immune response in the infected host. There are two broad categories of proinflammatory stimuli that can be observed during Salmonella infection. These are pathogen-associated factors that stimulate the innate immune system of the host and virulence associated factors that exploit host processes resulting in disease pathology.
Salmonella pathogenicity islands (SPI), historically acquired through horizontal gene transfer events, include clusters of genes, which encode the mechanisms through which Salmonella acts as a virulent pathogen (23, 24). These genetic islands are located on the bacterial chromosome or on plasmids, however, not all serovars possess every known SPI. SPI-1 through SPI-5 are common among all S. enterica serovars (Table 1). To date, 23 SPI have been described although the functions of those genes contained within each island have not yet been completely elucidated (25, 26). SPI-1 and SPI-2 are of particular importance in in vivo infection (as shown in Table 1; Figure 2). The SPI encode effector proteins that are translocated directly into host cells across the plasma membrane type III secretion systems (T3SS-1 and T3SS-2) that provide Salmonella with the biochemical machinery to exploit this intracellular niche. T3SSs can also be used to secrete effector proteins into the surrounding environment to influence host cell physiology (27, 28) (Table 1).
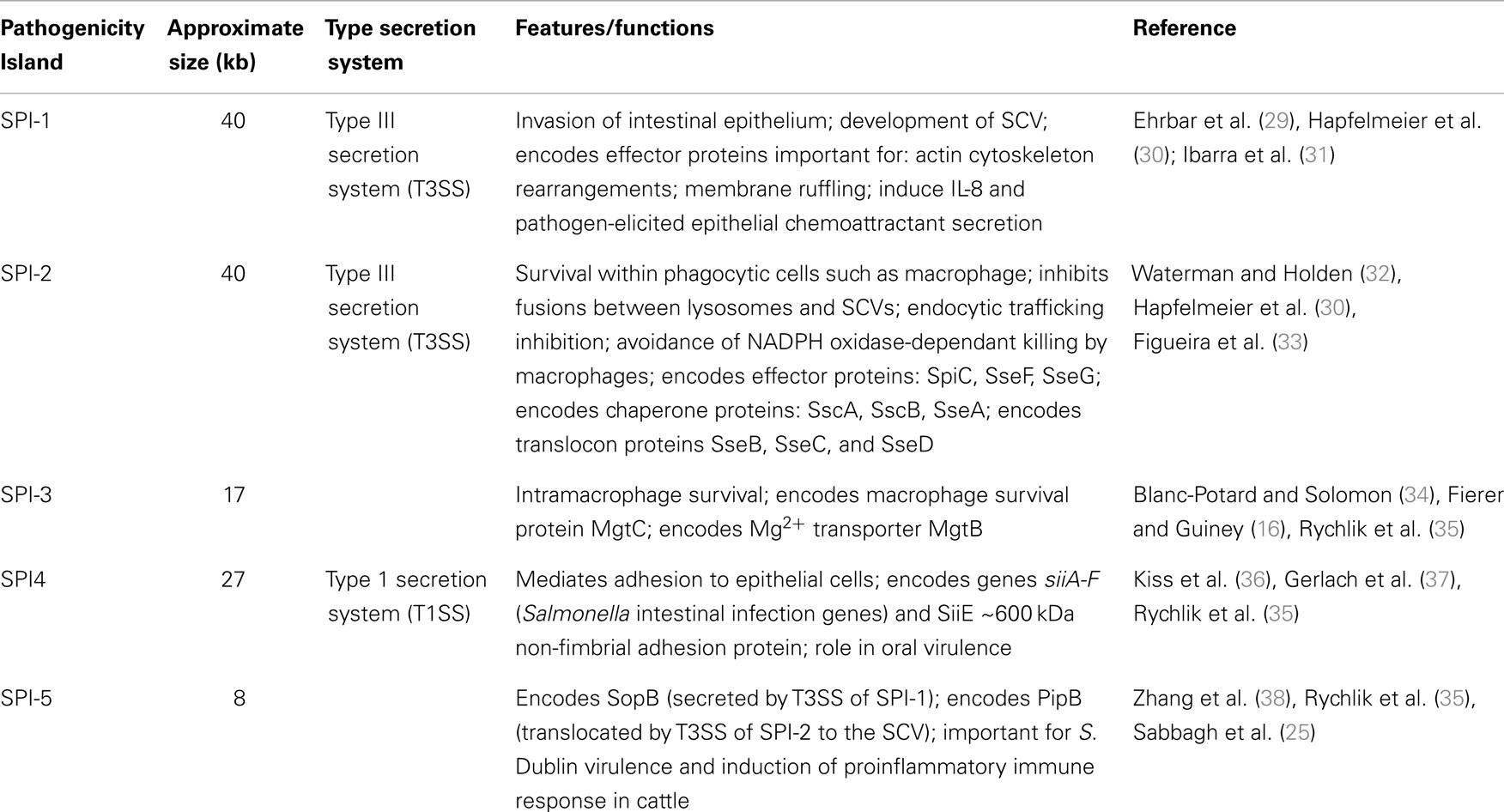
Table 1. Features and functions of SPI-1 through SPI-5 identified among all S. enterica serovars.
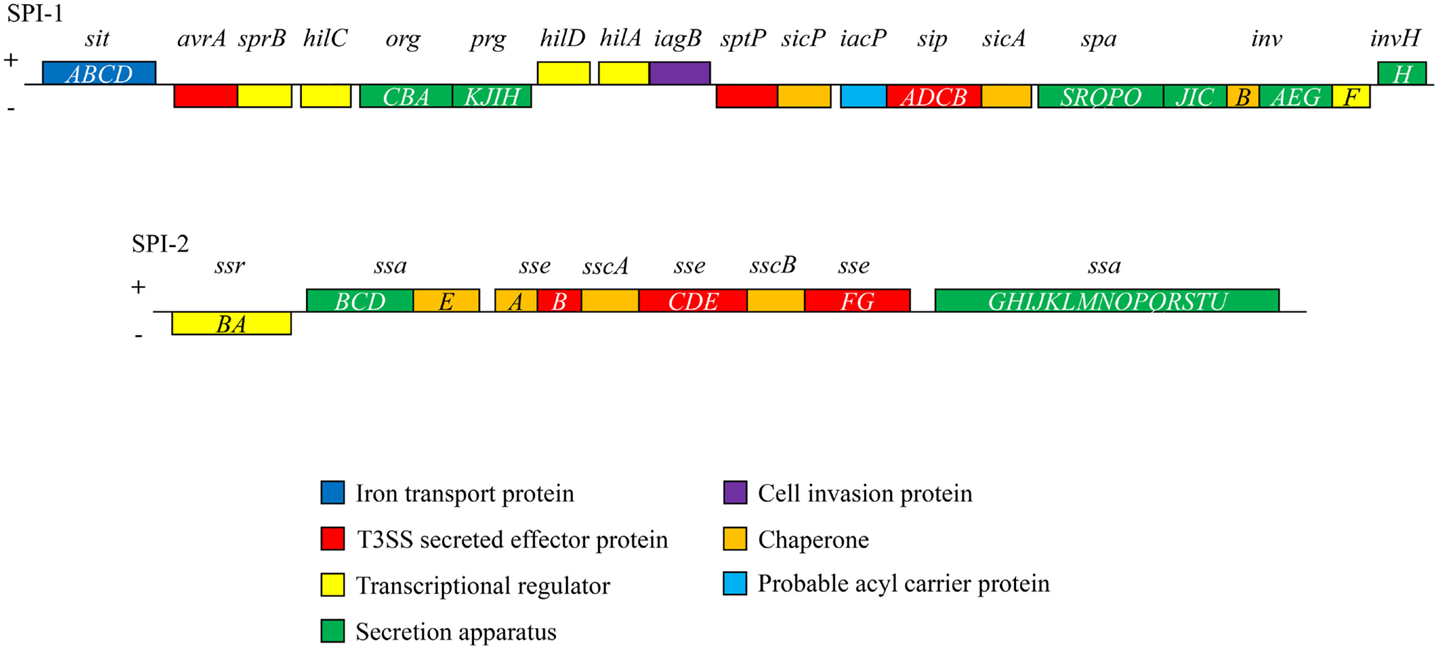
Figure 2. Schematic illustration of the genes of SPI-1 and SPI-2 indicating their functional categories is shown. In Salmonella, SPI-1 and SPI-2 encode a range of effector proteins, secretion apparatus, and transcriptional regulators in addition to T3SS-1 and T3SS-2.
Salmonella pathogenicity islands-1 was originally thought to be important as an invasion-related cluster of genes required for oral virulence (39). More recently, additional functions have been described for this locus. SPI-1-induced activation of the host innate immune system results in inflammation and the recruitment of polymorphonuclear (PMN) cells across the intestinal epithelial barrier following the secretion of the effector protein SipA by Salmonella. The latter protein is required in conjunction with the cytokine, IL-8, and pathogen-elicited epithelial chemoattractant (PEEC) to recruit neutrophils as has been reported in cultured epithelial monolayers (40). The production of PEEC can be induced by SipA secretion or by direct addition of SipA to cultured intestinal epithelial monolayers leading to the recruitment of basolateral neutrophils to the apical epithelial membrane (41, 42). SPI-1 effector secretion also leads to NF-κB signaling- and caspase-1-mediated IL-1β/IL-18 activation (43). SipB, an SPI-1 encoded effector protein, which is translocated across the host cell membrane by T3SS-1, is critical for inflammatory disease in vivo (38) and is responsible for pyroptotic cell death, a rapid form of programed cell death associated with antimicrobial responses during inflammation that possesses both apoptotic and necrotic features (44, 45). SipB binds caspase-1 (IL-1β converting enzyme) in the cell cytosol resulting in the maturation of proinflammatory cytokines IL-1β and IL-18 into active peptides (46). Further studies have revealed that both caspase-1 and Ipaf deficient mice exhibit an increased susceptibility to typhoid fever, thereby demonstrating the protective proinflammatory role played by caspase-1 (47).
The proinflammatory activity of SPI-2 while less characterized has been shown to be important for intracellular persistence and systemic virulence in murine typhoid fever in addition to evading host phagosome oxidation mechanisms (48). T3SS-2 plays an important role in inflammatory disease, highlighting the involvement of SPI-2 in the onset of enterocolitis. SPI-2 functions by enabling the translocation of effectors across the membrane of the Salmonella-containing vacuole (SCV) in infected host cells. The genes encoding T3SS-2 are controlled by two-component regulatory systems such as OmpR–EnvZ and the SPI-2 encoded SsrA–SsrB. As many as 28 SPI-2 encoded effectors have been identified to date with many of these currently of unknown function such as SseK1-3 and SteA–B, D–E. SseF is involved in SCV localization and Salmonella-induced filament (Sif) formation. PipB2 is responsible for kinesin-1 recruitment to the SCV and Sif extension, whereas SspH2 and SteC are recruited to and involved in the formation of the SCV-associated F-actin meshwork, respectively (49). The Toll-like receptors (TLR) adapter, myeloid differentiation primary response gene (MyD88) is required for SPI-1 independent intestinal inflammation in mice (30).
The Interaction of Salmonella with the Host
Salmonella invades both phagocytic and non-phagocytic cells including mononuclear phagocytic cells present in the lymphoid follicles, liver, and spleen. Epithelial cells and phagocytic cells such as dendritic cells, neutrophils, and macrophages identify specific pathogen-associated molecular pattern (PAMP) motifs and endogenous danger-associated molecular pattern molecules (DAMPs) present in the bacteria. Pattern-recognition receptors (PRRs), which include NOD-like receptors (NLRs) and TLRs, comprise the early components of the immune system that function to detect invading pathogens through PAMPs and DAMPs and signal to recruit and activate phagocytic cells such as neutrophils and macrophages (50, 51). These receptors trigger an immune response and are key to establishing an important network between the innate and adaptive immune systems. Bacterial DNA, flagella, and LPS are examples of PAMPs, which activate TLR4, TLR5, and TLR9 signaling in the host. LPS-induced TLR4 activation is important for triggering the inflammatory responses of the host. It also plays an important role in mounting an inflammatory response to intravenously administered LPS. Mice with mutations in TLR4-encoding genes exhibit an increased susceptibility to Salmonella infection irrespective of other Salmonella resistance loci (52, 53). Additionally, LPS plays an important role in the onset of sepsis during systemic infection as observed by its role in inducing inflammation in macrophages (54).
The immune system can be divided into two main parts: the innate or non-specific and the adaptive or specific components. The innate immune system is the first host challenge presented to invading pathogens whereas the adaptive immune system provides further protection in addition to an immunological memory, which enables a faster response upon repeat exposure to the same pathogen or antigen. In addition to cellular components such as phagocytic cells, there are humoral elements such as the complement system that make up the innate immune system. Additionally, anatomical features like the mammalian skin layer act as physical barriers to infection. The interplay between the innate and adaptive immune systems, including different types of cells and molecules such as cytokines and antibodies, form the totality of the host immunity.
Leukocytes of the innate immune system include phagocytic cells, namely dendritic cells, macrophages, and neutrophils, which can engulf foreign antigens, particles, or pathogens. These phagocytic cells are recruited following the release of specific cytokine signals. These cells serve an important role in the activation of the adaptive immunity, which usually assumes the presence of lymphocytes (55). Other cells, such as basophils, eosinophils, and mast cells are also part of the host innate immune system that contributes to the innate immunity.
During the initial stages of an inflammatory response, neutrophils and macrophages are recruited to the site of infection. Neutrophils phagocytose the invading pathogens and kill them intracellularly. Similarly, macrophages and newly recruited monocytes, which will differentiate into macrophages following signaling or chemical stimulation, also function by phagocytosing and killing the pathogens at the intracellular level. Furthermore, macrophages are capable of killing infected or self-target cells and can also induce further downstream immune responses through the presentation of surface antigens to signal and recruit other cells and cell types (56).
A common feature of salmonellosis is the notable inflammatory response elicited by the host innate immune system. Both the host and pathogen have evolved defense mechanisms that result in a complex cross-talk that culminates with the induction of the host immune response.
Salmonella species can cross the epithelial barrier by passive transport facilitated by dendritic cells, which extend pseudopods between local epithelial cells, or by active invasion. Upon reaching the lower intestine, the bacteria will adhere to the mucosal membranes and invade epithelial cells (57). One such site where this occurs is the microfold (M) cells of Peyer’s patches that are located in the small intestine where the bacteria will translocate across the epithelial barrier to the underlying follicles and mesenteric lymph nodes of the lymphoid tissue (58) (Figures 3A,B). During sustained bacteremia, secondary infections can occur due to the dissemination of the bacteria to other organs such as the gall bladder, liver, and spleen. The gall bladder serves as a reservoir in chronic cases of S. Typhi and S. Typhimurium infection (59, 60). Infection by invading bacteria can originate from both the blood and/or retrograde bile. Biofilm formation on gallstones is a reported avenue through which chronic carriage and shedding of Salmonella species can be established. These events set in motion a cycle of infection wherein bacteria basolaterally reinvade epithelial cells of the intestinal wall or are shed in feces. In time, the symptoms of salmonellosis will resolve. However, asymptomatic carriage of the bacteria can occur in patients for months or years with the potential to relapse in the future.
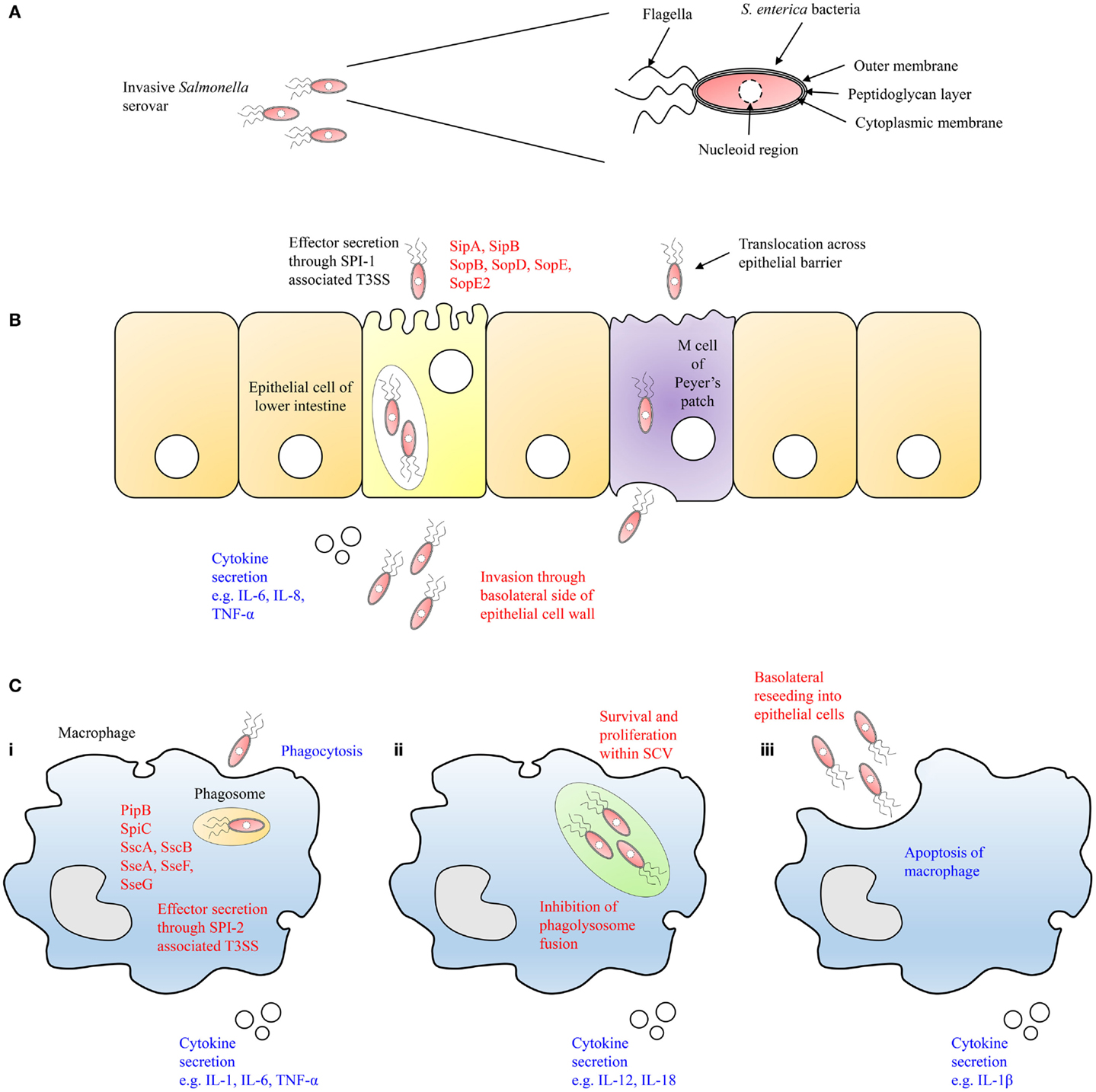
Figure 3. Schematic illustration of the infection of epithelial cells of the lower intestine and macrophages by Salmonella is shown. (A) The complex membrane structure of Salmonella allows it to survive until reaching the epithelial cell wall of the host in the lower intestine. (B) Salmonella then translocate across M cells of Peyer’s patches or actively invade epithelial cells by the secretion of effector proteins through the SPI-1 encoded T3SS-1. (C) (i) After crossing the epithelial barrier, Salmonella are engulfed by proximal macrophages that will secrete effector proteins into the cytosol of the cell via the SPI-2 encoded T3SS-2 and prevent fusion of the phagosome with the lysosome. (ii) Within the SCV, Salmonella will proliferate resulting in cytokine secretion by the macrophage. (iii) Finally, the macrophage will undergo apoptosis, and Salmonella will escape the cell to basolaterally reinvade epithelial cells or other phagocytic cells of the host innate immune system.
Transmission of Infection
Following the ingestion of contaminated food, these bacteria will colonize the intestines by invading dendritic cells and enterocytes of the intestinal epithelium barrier. Salmonella species, which are successful in passing this barrier are confronted by proximal macrophages and may be phagocytosed, or actively invade the macrophages, using T3SS-1 and fimbriae, among other bacterial surface adhesins [H. (15)] (Figure 3Ci).
After being internalized by macrophages, Salmonella then reside within a membrane bound compartment distinct from the phagosome and lysosome known as the SCV. In this cellular compartment, Salmonella can survive and replicate in the absence of host antimicrobial defense mechanisms, thereby evading endosomal fusion with the NADPH oxidase complex (61) (Figure 3Cii). From within the SCVs, SPI-2 genes are expressed encoding T3SS-2, which enables Salmonella to translocate a range of effector proteins into the cytoplasm of the host cell including SigD/SopB, SipA, SipC, SodC-1, SopE2, and SptP leading to the rearrangement of the actin cytoskeleton. T3SS-2 has been described as necessary for systemic virulence in murine models and survival within macrophages (62). In contrast, systemic translocation of S. Dublin in cattle requires T3SS-1 but not T3SS-2 (63).
Cytokine Responses and Signaling
Proinflammatory cytokines including the interleukins (IL-1β and IL-6), interferons (IFN-γ), and tumor necrosis factor (TNF-α) are synthesized and these act to promote systemic inflammation (64–67). IFN-γ, also known as macrophage activating factor (MAF), plays an important role in persistent infection as it influences the duration of macrophage activation. Secretion of IFN-γ is dependent on IL-18, also known as interferon gamma inducing factor, and is essential for establishing an early host resistance to infection with Salmonella (65, 68).
Macrophages are involved in both the innate and adaptive immune responses. Following exposure to specific cytokines, they undergo either classical (Th1) or alternative (Th2) activation. Classical activation by bacterial LPS or IFN-γ leads to alteration in the secretory profile of the cells through production of organic nitrate compounds such as nitric oxide (NO). Alternative activation by IL-4, IL-10, or IL-13 leads to the production of polyamines and proline inducing proliferation and collagen production, respectively. The presence of Salmonella within these cells leads to cytokine secretion and an inflammatory reaction or programed cell death through apoptosis (69, 70) (Figure 3Ciii).
Cytokine signaling, induced by the interaction of the host cells and bacteria, is crucial to the development and progression of salmonellosis. Cytokines are responsible for regulating both the innate and adaptive host immune responses. The equilibrium between pro- and anti-inflammatory cytokines controls the infection preventing damage to the host from prolonged inflammation. In vitro cell culture of bone marrow derived macrophages and primary cell lines have shown that Salmonella promotes chemokine and cytokine synthesis in both dendritic and epithelial cells as well as macrophages (69, 71, 72). Cytokines have a broad range of effects upon the host cell during infection. Chemokine C–C motif ligand (CCL2), IFN-γ, IL-12, IL-18, TNF-α, and transforming growth factor (TGF-β) confer protection during infection (73). Conversely, IL-4 and IL-10 interfere with the host defense mechanisms (74).
Environment Adaptation
Salmonella adapt to the intracellular environment of phagocytic cells during infection. The transition from extracellular to intravacuolar environments involves global modulation of bacterial gene expression. The complete transcriptional landscape of intracellular S. Typhimurium following macrophage infection has been previously reported (75, 76). During replication in murine J774 macrophages, 919 of 4,451 S. Typhimurium genes are differentially expressed. Many of the in vivo-regulated genes are of unknown function suggesting novel macrophage-associated functions for intracellular growth (77).
It has been shown previously that S. Typhimurium requires glycolysis for infection of mice and macrophages and that glucose transport is required for replication within macrophages. During systemic infection of mice, S. Typhimurium replicates in macrophages within the SCV. Mutation of the pfkAB-encoded phosphofructokinase, the rate-limiting step in glycolysis, severely attenuates replication and survival within RAW 264.7 macrophages. Mutants with perturbed phosphoenolpyruvate:carbohydrate phosphotransferase systems or those unable to catabolize glucose exhibit reduced replication within RAW 264.7 macrophages (78).
Salmonella upregulates RpoS-dependent stress responses as well as other response mechanisms when challenged to grow in sublethal concentrations of the bile salt sodium deoxycholate (DOC). The latter is known to disrupt membranes, denature proteins, and damage DNA (79). It has been previously shown that Salmonella can pre-adapt to several stresses in order to survive the adverse conditions encountered, such as those encountered in a contaminated food matrix and any associated food production processes. Similarly, the subsequent ingestion of the bacterium by the host presents an array of challenges to the organism including acid, cold, osmotic, and peroxide stress (80).
Pathological Symptoms
Prolonged activation of the innate immune system can have adverse effects, which include intravascular coagulation, systemic inflammation, and tissue injury. In severe cases, these symptoms can lead to death. An aggressive proinflammatory response to infection with Salmonella is not a common occurrence and it arises rarely in patients with typhoid fever. Unusual cases leading to intravascular coagulation do not present with readily recognizable clinical signs (81, 82). In these cases, the blood serum levels of IL-1β and TNF-α are lower when compared to that of patients infected with other Gram-negative bacteria (83).
Individuals suffering from typhoid fever exhibit a distinct peripheral blood metabolite profile, which has been elucidated by both microarray and transcriptional profiling techniques (66, 84). This profile diminishes following treatment and upon recovery the majority of individuals exhibit a peripheral blood profile similar to that of uninfected controls. Those who do not develop a typical peripheral blood profile following treatment may possess genetic mutations that render them incapable of mounting an appropriate immune response. These patients have been shown to be prone to relapse, reinfection, and in some cases become carriers (66).
Immunodeficiency
There has been no evidence to support a correlation with susceptibility to typhoid fever and primary or acquired immunodeficiency. This is in contrast to infection with NTS serovars where infection causes high levels of morbidity and mortality in patients with primary or acquired immunodeficiencies such as HIV infection. It has been proposed that this difference is attributed to the manner in which signaling occurs via the PRRs. The production of IL-17 by T-helper 17 cells (Th17) among other cytokines (IL-21, IL-22, and IL-26) is important for the dissemination of NTS serovars but not S. Typhi (85, 86).
Models of Infection
S. Typhi is a host-adapted pathogen, which infects humans causing typhoid fever. Investigating the interactions of this pathogen with the host has proved challenging as there are few animal models for typhoid fever that are of direct relevance to their human infection counterpart. This problem has been partially alleviated by the establishment of the murine S. Typhimurium infection model, which has been used to study typhoid fever. The immune responses and subsequent inflammation mounted by mice following an S. Typhimurium infection mimics those observed in human patients with typhoid fever as well as the subsequent intestinal pathology (87). Mice are inoculated orally or systemically by intravenous or intraperitoneal injection in addition to optional streptomycin pre-treatment (88). S. Typhimurium induced colitis in streptomycin-pre-treated mice is reminiscent of many symptoms of the human infection counterpart including epithelial ulceration and infiltration of PMN/CD18(+) cells (89). A comparison between streptomycin-pre-treated and untreated mice highlighted the drastic influence of streptomycin on resistance to colonization by S. Typhimurium whereby 100% of treated and none of the untreated mice excreted the bacterium in their feces (90). A disease with features reminiscent of typhoid fever can be observed in BALB/c or C57BL/6 mice when inoculated with S. Typhimurium due to a mutation in the SLC11A1 gene, which encodes natural resistance-associated macrophage protein one (Nramp1). In contrast to this, chronic and persistent carrier states of infection can be studied using Nramp+/+ mice as they are resistant to infection with S. Typhimurium (25, 88). However, there has been no correlation identified in humans between Nramp alleles and susceptibility to typhoid fever as S. Typhimurium causes less severe disease symptoms in humans to that of S. Typhi. As a result, conclusions drawn from animal experiments must be interpreted carefully (91). Furthermore, it has been reported that tlr11−/−mice are more susceptible to infection by S. Typhimurium and can be infected with S. Typhi, which typically does not cause infection as TRL11 is normally expressed in mice but not in humans (92). Recently, an alternative S. Typhi murine model, which resembles human typhoid fever, was established using non-obese diabetic (NOD)-SCID IL2rγnull mice, which have been humanized by engrafting human hematopoietic stem cells (hu–SRC–SCID mice). This model results in lethal infection with inflammatory and pathological responses, which mimic human typhoid fever (93).
As well as murine models of infection, the larvae of the wax moth Galleria mellonella (G. mellonella) have been used to study host–pathogen interactions with Salmonella species. Isogenic mutant strains of S. Typhimurium lacking known virulence determinants were tested to identify their role in pathogenicity. Interestingly, mutants depleted of either or both SPI encoded T3SS-1 and T3SS-2 exhibited no alterations in their virulence phenotype. Attenuation of the PhoPQ two-component signal transduction system resulted in reduced pathogenicity due to the lack of phoQ (94). As reported in murine models, mutations in the hfq gene, which encodes the chaperone protein Hfq that plays an important role in the binding of regulatory sRNA transcripts to their antisense targets attenuated the pathogenicity of S. Typhimurium in G. mellonella. Endoribonuclease RNase E and RNase III mutants show an attenuated virulence phenotype including impairment in motility and reduced proliferation inside G. mellonella (95).
Recently, zebrafish (Danio rerio) models have provided a unique opportunity to study the function of phagocytic cells such as neutrophils and macrophages. Transgenic zebrafish lines with fluorescently labeled leukocyte populations enable non-invasive imaging of the mechanisms by which different pathogens interact with macrophages and evade the host innate immunity (96). Similarly, 28 h old zebrafish embryos infected with DsRed labeled S. Typhimurium allowed for the precise location of the pathogen to be determined in a living host over a 3 day time course using multidimensional digital imaging microscopy. Lethal infection with S. Typhimurium residing and proliferating in both the endothelium layer of blood vessels and macrophages was observed (97).
Future Perspectives
To date, there have been many studies elucidating the complex Salmonella–host interactome. Our understanding of the virulence determinants of Salmonella species and their mechanisms of action has been extended by the utilization of murine, G. mellonella, and zebrafish models of S. Typhimurium infection in addition to ex vivo cell culture methods. Despite this, further work is needed to determine the specific contribution of many of these regulators and virulence factors for which clear functions and roles have yet to be defined. Characterizing the pathogenesis of salmonellosis will be crucial to the development and implementation of future therapeutic strategies to treat this illness. The importance of which has been recently highlighted in reports on the emergence of antimicrobial resistance in Salmonella and many other bacterial pathogens (98).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Daniel Hurley’s research is supported by the Wellcome Trust Computational Infection Biology Ph.D. Programme (Grant reference: 099837/Z/12/Z).
References
1. Gunn JS, Marshall JM, Baker S, Dongol S, Charles RC, Ryan ET. Salmonella chronic carriage: epidemiology, diagnosis, and gallbladder persistence. Trends Microbiol (2014). doi: 10.1016/j.tim.2014.06.007
2. Gal-Mor O, Boyle EC, Grassl GA. Same species, different diseases: how and why typhoidal and non-typhoidal Salmonella enterica serovars differ. Front Microbiol (2014) 5:391. doi:10.3389/fmicb.2014.00391
3. Batz MB, Henke E, Kowalcyk B. Long-term consequences of foodborne infections. Infect Dis Clin North Am (2013) 27(3):599–616. doi:10.1016/j.idc.2013.05.003
4. Chen HM, Wang Y, Su LH, Chiu CH. Nontyphoid Salmonella infection: microbiology, clinical features, and antimicrobial therapy. Pediatr Neonatol (2013) 54(3):147–52. doi:10.1016/j.pedneo.2013.01.010
5. Fàbrega A, Vila J. Salmonella enterica serovar Typhimurium skills to succeed in the host: virulence and regulation. Clin Microbiol Rev (2013) 26(2):308–41. doi:10.1128/CMR.00066-12
6. Santos RL. Pathobiology of salmonella, intestinal microbiota, and the host innate immune response. Front Immunol (2014) 26(5):252. doi:10.3389/fimmu.2014.00252
7. Ruby T, McLaughlin L, Gopinath S, Monack D. Salmonella’s long-term relationship with its host. FEMS Microbiol Rev (2012) 36(3):600–15. doi:10.1111/j.1574-6976.2012.00332.x
8. Croswell A, Amir E, Teggatz P, Barman M, Salzman NH. Prolonged impact of antibiotics on intestinal microbial ecology and susceptibility to enteric Salmonella infection. Infect Immun (2009) 77(7):2741–53. doi:10.1128/IAI.00006-09
9. Gopinath S, Carden S, Monack D. Shedding light on Salmonella carriers. Trends Microbiol (2012) 20(7):320–7. doi:10.1016/j.tim.2012.04.004
10. Thiennimitr P, Winter SE, Bäumler AJ. Salmonella, the host and its microbiota. Curr Opin Microbiol (2012) 15(1):108–14. doi:10.1016/j.mib.2011.10.002
11. Raffatellu M, Wilson RP, Winter SE, Bäumler AJ. Clinical pathogenesis of typhoid fever. J Infect Dev Ctries (2008) 2(4):260–6. doi:10.3855/jidc.219
12. Crump JA, Mintz ED. Global trends in typhoid and paratyphoid fever. Clin Infect Dis (2010) 50(2):241–6. doi:10.1086/649541
13. Majowicz SE, Musto J, Scallan E, Angulo FJ, Kirk M, O’Brien SJ, et al. The global burden of nontyphoidal Salmonella gastroenteritis. Clin Infect Dis (2010) 50(6):882–9. doi:10.1086/650733
14. Coburn B, Grassl GA, Finlay BB. Salmonella, the host and disease: a brief review. Immunol Cell Biol (2006) 85(2):112–8. doi:10.1038/sj.icb.7100007
15. de Jong HK, Parry CM, van der Poll T, Wiersinga WJ. Host–pathogen interaction in invasive Salmonellosis. PLoS Pathog (2012) 8(10):e1002933. doi:10.1371/journal.ppat.1002933
16. Fierer J, Guiney DG. Diverse virulence traits underlying different clinical outcomes of Salmonella infection. J Clin Invest (2001) 107(7):775–80. doi:10.1172/JCI12561
17. Bäumler AJ, Tsolis RM, Ficht TA, Adams LG. Evolution of host adaptation in Salmonella enterica. Infect Immun (1998) 66(10):4579–87.
18. Stecher B, Macpherson AJ, Hapfelmeier S, Kremer M, Stallmach T, Hardt W-D. Comparison of Salmonella enterica serovar Typhimurium colitis in germfree mice and mice pretreated with streptomycin. Infect Immun (2005) 73(6):3228–41. doi:10.1128/IAI.73.6.3228-3241.2005
19. Gordon MA. Salmonella infections in immunocompromised adults. J Infect (2008) 56(6):413–22. doi:10.1016/j.jinf.2008.03.012
20. Dhanoa A, Fatt QK. Non-typhoidal Salmonella bacteraemia: epidemiology, clinical characteristics and its’ association with severe immunosuppression. Ann Clin Microbiol Antimicrob (2009) 8:15. doi:10.1186/1476-0711-8-15
21. Richter-Dahlfors A, Buchan AMJ, Finlay BB. Murine Salmonellosis studied by confocal microscopy: Salmonella Typhimurium resides intracellularly inside macrophages and exerts a cytotoxic effect on phagocytes in vivo. J Exp Med (1997) 186(4):569–80. doi:10.1084/jem.186.4.569
22. Yrlid U, Svensson M, Håkansson A, Chambers BJ, Ljunggren HG, Wick MJ. In vivo activation of dendritic cells and T cells during Salmonella enterica serovar Typhimurium infection. Infect Immun (2001) 69(9):5726–35. doi:10.1128/IAI.69.9.5726-5735.2001
23. Knodler LA, Finlay BB, Steele-Mortimer O. The Salmonella effector protein SopB protects epithelial cells from apoptosis by sustained activation of Akt. J Biol Chem (2005) 280(10):9058–64. doi:10.1074/jbc.M412588200
24. Ibarra JA, Steele-Mortimer O. Salmonella – the ultimate insider. Salmonella virulence factors that modulate intracellular survival. Cell Microbiol (2009) 11(11):1579–86. doi:10.1111/j.1462-5822.2009.01368.x
25. Sabbagh SC, Forest CG, Lepage C, Leclerc J-M, Daigle F. So similar, yet so different: uncovering distinctive features in the genomes of Salmonella enterica serovars Typhimurium and Typhi. FEMS Microbiol Lett (2010) 305(1):1–13. doi:10.1111/j.1574-6968.2010.01904.x
26. Hayward MR, Jansen VAA, Woodward MJ. Comparative genomics of Salmonella enterica serovars Derby and Mbandaka, two prevalent serovars associated with different livestock species in the UK. BMC Genomics (2013) 14(1):365. doi:10.1186/1471-2164-14-365
27. Galán JE. Interaction of Salmonella with host cells through the centisome 63 Type III secretion system. Curr Opin Microbiol (1999) 2(1):46–50. doi:10.1016/S1369-5274(99)80008-3
28. Hensel M, Shea JE, Waterman SR, Mundy R, Nikolaus T, Banks G, et al. Genes encoding putative effector proteins of the type III secretion system of Salmonella pathogenicity island 2 are required for bacterial virulence and proliferation in macrophages. Mol Microbiol (1998) 30(1):163–74. doi:10.1046/j.1365-2958.1998.01047.x
29. Ehrbar K, Friebel A, Miller SI, Hardt WD. Role of the Salmonella pathogenicity island 1 (spi-1) protein InvB in type III secretion of SopE and SopE2, two Salmonella effector proteins encoded outside of SPI-1. J Bacteriol (2003) 185(23):6950–67. doi:10.1128/JB.185.23.6950-6967.2003
30. Hapfelmeier S, Stecher B, Barthel M, Kremer M, Müller AJ, Heikenwalder M, et al. The Salmonella pathogenicity island (SPI)-2 and SPI-1 type III secretion systems allow Salmonella serovar typhimurium to trigger colitis via MyD88-dependent and MyD88-independent mechanisms. J Immunol (2005) 174(3):1675–85. doi:10.4049/jimmunol.174.3.1675
31. Ibarra JA, Knodler LA, Sturdevant DE, Virtaneva K, Carmody AB, Fischer ER, et al. Induction of Salmonella pathogenicity island 1 under different growth conditions can affect Salmonella-host cell interactions in vitro. Microbiology (2010) 156(Pt 4):1120–33. doi:10.1099/mic.0.032896-0
32. Waterman SR, Holden DW. Functions and effectors of the Salmonella pathogenicity island 2 type III secretion system. Cell Microbiol (2003) 5:501–11. doi:10.1046/j.1462-5822.2003.00294.x
33. Figueira R, Watson KG, Holden DW, Helaine S. Identification of Salmonella pathogenicity island-2 type III secretion system effectors involved in intramacrophage replication of S. enterica serovar Typhimurium: implications for rational vaccine design. MBio (2013) 4(2):e00065–13. doi:10.1128/mBio.00065-13
34. Blanc-Potard AB, Solomon F, Kayser J, Groisman EA. The SPI-3 pathogenicity island of Salmonella enterica. J Bacteriol (1999) 181(3):998–1004.
35. Rychlik I, Karasova D, Sebkova A. Virulence potential of five major pathogenicity islands (SPI-1 to SPI-5) of Salmonella enterica serovar enteritidis for chickens. BMC Microbiol (2009) 9:268. doi:10.1186/1471-2180-9-268
36. Kiss T, Morgan E, Nagy G. Contribution of SPI-4 genes to the virulence of Salmonella enterica. FEMS Microbiol Lett (2007) 275(1):153–9. doi:10.1111/j.1574-6968.2007.00871.x
37. Gerlach RG, Jäckel D, Stecher B. Salmonella pathogenicity island 4 encodes a giant non-fimbrial adhesin and the cognate type 1 secretion system. Cell Microbiol (2007) 9(7):1834–50. doi:10.1111/j.1462-5822.2007.00919.x
38. Zhang S, Santos RL, Tsolis RM, Stender S, Hardt W-D, Bäumler AJ, et al. The Salmonella enterica serotype Typhimurium effector proteins SipA, SopA, SopB, SopD, and SopE2 Act in concert to induce diarrhea in calves. Infect Immun (2002) 70(7):3843–55. doi:10.1128/IAI.70.7.3843-3855.2002
39. Marcus SL, Brumell JH, Pfeifer CG, Finlay BB. Salmonella pathogenicity islands: big virulence in small packages. Microbes Infect (2000) 2(2):145–56. doi:10.1016/S1286-4579(00)00273-2
40. McCormick BA, Colgan SP, Delp-Archer C, Miller SI, Madara JL. Salmonella typhimurium attachment to human intestinal epithelial monolayers: transcellular signalling to subepithelial neutrophils. J Cell Biol (1993) 123(4):895–907. doi:10.1083/jcb.123.4.895
41. Gewirtz AT, Siber AM, Madara JL, McCormick BA. Orchestration of neutrophil movement by intestinal epithelial cells in response to Salmonella Typhimurium can be uncoupled from bacterial internalization. Infect Immun (1999) 67(2):608–17.
42. Lee CA, Silva M, Siber AM, Kelly AJ, Galyov E, McCormick BA. A secreted Salmonella protein induces a proinflammatory response in epithelial cells, which promotes neutrophil migration. Proc Natl Acad Sci USA (2000) 97(22):12283–8. doi:10.1073/pnas.97.22.12283
43. Hobbie S, Chen LM, Davis RJ, Galán JE. Involvement of mitogen-activated protein kinase pathways in the nuclear responses and cytokine production induced by Salmonella Typhimurium in cultured intestinal epithelial cells. J Immunol (1997) 159(11):5550–9.
44. Hernandez LD, Pypaert M, Flavell RA, Galán JE. A Salmonella protein causes macrophage cell death by inducing autophagy. J Cell Biol (2003) 163(5):1123–31. doi:10.1083/jcb.200309161
45. Fink SL, Cookson BT. Apoptosis, pyroptosis, and necrosis: mechanistic description of dead and dying eukaryotic cells. Infect Immun (2005) 73(4):1907–16. doi:10.1128/IAI.73.4.1907-1916.2005
46. Hersh D, Monack DM, Smith MR, Ghori N, Falkow S, Zychlinsky A. The Salmonella invasin SipB induces macrophage apoptosis by binding to caspase-1. Proc Natl Acad Sci USA (1999) 96(5):2396–401. doi:10.1073/pnas.96.5.2396
47. Lara-Tejero M, Sutterwala FS, Ogura Y, Grant EP, Bertin J, Coyle AJ, et al. Role of the caspase-1 inflammasome in Salmonella Typhimurium pathogenesis. J Exp Med (2006) 203(6):1407–12. doi:10.1084/jem.20060206
48. Coburn B, Li Y, Owen D, Vallance BA, Finlay BB. Salmonella enterica serovar Typhimurium pathogenicity island 2 is necessary for complete virulence in a mouse model of infectious enterocolitis. Infect Immun (2005) 73(6):3219–27. doi:10.1128/IAI.73.6.3219-3227.2005
49. Figueira R, Holden DW. Functions of the Salmonella pathogenicity island 2 (SPI-2) type III secretion system effectors. Microbiology (2012) 158(Pt 5):1147–61. doi:10.1099/mic.0.058115-0
50. Kawai T, Akira S. The role of pattern-recognition receptors in innate immunity: update on toll-like receptors. Nat Immunol (2010) 11(5):373–84. doi:10.1038/ni.1863
51. Schroder K, Tschopp J. The inflammasomes. Cell (2010) 140(6):821–32. doi:10.1016/j.cell.2010.01.040
52. Vazquez-Torres A, Vallance BA, Bergman MA, Finlay BB, Cookson BT, Jones-Carson J, et al. Toll-like receptor 4 dependence of innate and adaptive immunity to Salmonella: importance of the Kupffer cell network. J Immunol (2004) 172(10):6202–8. doi:10.4049/jimmunol.172.10.6202
53. Wilson RP, Raffatellu M, Chessa D, Winter SE, Tükel C, Bäumler AJ. The Vi-capsule prevents toll-like receptor 4 recognition of Salmonella. Cell Microbiol (2008) 10(4):876–90. doi:10.1111/j.1462-5822.2007.01090.x
54. O’Brien GC, Wang JH, Redmond HP. Bacterial lipoprotein induces resistance to Gram-negative sepsis in TLR4-deficient mice via enhanced bacterial clearance. J Immunol (2005) 174(2):1020–6. doi:10.4049/jimmunol.174.2.1020
55. Janeway CA Jr., Travers P, Walport M, Shlomchik MJ. Immunobiology. New York: Garland Science (2001).
56. Delves PJ, Martin SJ, Burton DR, Roitt IM. Essential Immunology. New Jersey: Wiley-Blackwell (2011).
57. Rescigno M, Urbano M, Valzasina B, Francolini M, Rotta G, Bonasio R, et al. Dendritic cells express tight junction proteins and penetrate gut epithelial monolayers to sample bacteria. Nat Immunol (2001) 2(4):361–7. doi:10.1038/86373
58. Jones BD, Ghori N, Falkow S. Salmonella Typhimurium initiates murine infection by penetrating and destroying the specialized epithelial M cells of the Peyer’s patches. J Exp Med (1994) 180(1):15–23. doi:10.1084/jem.180.1.15
59. Crawford RW, Rosales-Reyes R, Ramírez-Aguilar Mde L, Chapa-Azuela O, Alpuche-Aranda C, Gunn JS. Gallstones play a significant role in Salmonella spp. gall bladder colonization and carriage. Proc Natl Acad Sci USA (2010) 107(9):4353–8. doi:10.1073/pnas.1000862107
60. Gonzalez-Escobedo G, Marshall JM, Gunn JS. Chronic and acute infection of the gall bladder by Salmonella Typhi: understanding the carrier state. Nat Rev Microbiol (2011) 9(1):9–14. doi:10.1038/nrmicro2490
61. Gorvel JP, Méresse S. Maturation steps of the Salmonella-containing vacuole. Microbes Infect (2001) 3(14–15):1299–303. doi:10.1016/S1286-4579(01)01490-3
62. Coombes BK, Coburn BA, Potter AA, Gomis S, Mirakhur K, Li Y, et al. Analysis of the contribution of Salmonella pathogenicity islands 1 and 2 to enteric disease progression using a novel bovine ileal loop model and a murine model of infectious enterocolitis. Infect Immun (2005) 73(11):7161–9. doi:10.1128/IAI.73.11.7161-7169.2005
63. Pullinger GD, Paulin SM, Charleston B, Watson PR, Bowen AJ, Dziva F, et al. Systemic translocation of Salmonella enterica serovar Dublin in cattle occurs predominantly via efferent lymphatics in a cell-free niche and requires type III secretion system 1 (T3SS-1) but not T3SS-2. Infect Immun (2007) 75(11):5191–9. doi:10.1128/IAI.00784-07
64. Butler T, Ho M, Acharya G, Tiwari M, Gallati H. Interleukin-6, gamma interferon, and tumor necrosis factor receptors in typhoid fever related to outcome of antimicrobial therapy. Antimicrob Agents Chemother (1993) 37(11):2418–21. doi:10.1128/AAC.37.11.2418
65. Monack DM, Bouley DM, Falkow S. Salmonella typhimurium persists within macrophages in the mesenteric lymph nodes of chronically infected Nramp1+/+ Mice and can be reactivated by IFNgamma neutralization. J Exp Med (2004) 199(2):231–41. doi:10.1084/jem.20031319
66. Thompson LJ, Dunstan SJ, Dolecek C, Perkins T, House D, Dougan G, et al. Transcriptional response in the peripheral blood of patients infected with Salmonella enterica serovar typhi. Proc Natl Acad Sci USA (2009) 106(52):22433–8. doi:10.1073/pnas.0912386106
67. Gal-Mor O, Suez J, Elhadad D, Porwollik S, Leshem E, Valinsky L, et al. Molecular and cellular characterization of a Salmonella enterica serovar Paratyphi A outbreak strain and the human immune response to infection. Clin Vaccine Immunol (2012) 19(2):146–56. doi:10.1128/CVI.05468-11
68. Mastroeni P, Clare S, Khan S, Harrison JA, Hormaeche CE, Okamura H, et al. Interleukin 18 contributes to host resistance and gamma interferon production in mice infected with virulent Salmonella Typhimurium. Infect Immun (1999) 67(2):478–83.
69. Svensson M, Johansson C, Wick MJ. Salmonella Typhimurium-induced cytokine production and surface molecule expression by murine macrophages. Microb Pathog (2001) 31(2):91–102. doi:10.1006/mpat.2001.0448
70. Rosenberger CM, Pollard AJ, Finlay BB. Gene array technology to determine host responses to Salmonella. Microbes Infect (2001) 3(14–15):1353–60. doi:10.1016/S1286-4579(01)01497-6
71. Jung HC, Eckmann L, Yang SK, Panja A, Fierer J, Morzycka-Wroblewska E, et al. A distinct array of proinflammatory cytokines is expressed in human colon epithelial cells in response to bacterial invasion. J Clin Invest (1995) 95(1):55–65. doi:10.1172/JCI117676
72. Pietilä TE, Veckman V, Kyllönen P, Lähteenmäki K, Korhonen TK, Julkunen I. Activation, cytokine production, and intracellular survival of bacteria in Salmonella-infected human monocyte-derived macrophages and dendritic cells. J Leukoc Biol (2005) 78(4):909–20. doi:10.1189/jlb.1204721
73. Depaolo RW, Lathan R, Rollins BJ, Karpus WJ. The chemokine CCL2 is required for control of murine gastric Salmonella enterica infection. Infect Immun (2005) 73(10):6514–22. doi:10.1128/IAI.73.10.6514-6522.2005
74. Uchiya K-I, Groisman EA, Nikai T. Involvement of Salmonella pathogenicity island 2 in the up-regulation of interleukin-10 expression in macrophages: role of protein kinase A signal pathway. Infect Immun (2004) 72(4):1964–73. doi:10.1128/IAI.72.4.1964-1973.2004
75. Kröger C, Dillon SC, Cameron AD, Papenfort K, Sivasankaran SK, Hokamp K, et al. The transcriptional landscape and small RNAs of Salmonella enterica serovar Typhimurium. Proc Natl Acad Sci USA (2012) 109(20):E1277–86. doi:10.1073/pnas.1201061109
76. Kröger C, Colgan A, Srikumar S, Händler K, Sivasankaran SK, Hammarlöf DL, et al. An infection-relevant transcriptomic compendium for Salmonella enterica serovar Typhimurium. Cell Host Microbe (2013) 14(6):683–95. doi:10.1016/j.chom.2013.11.010
77. Eriksson S, Lucchini S, Thompson A, Rhen M, Hinton JC. Unravelling the biology of macrophage infection by gene expression profiling of intracellular Salmonella enterica. Mol Microbiol (2003) 47(1):103–18. doi:10.1046/j.1365-2958.2003.03313.x
78. Bowden SD, Rowley G, Hinton JC, Thompson A. Glucose and glycolysis are required for the successful infection of macrophages and mice by Salmonella enterica serovar Typhimurium. Infect Immun (2009) 77(7):3117–26. doi:10.1128/IAI.00093-09
79. Hernández SB, Cota I, Ducret A, Aussel L, Casadesús J. Adaptation and preadaptation of Salmonella enterica to bile. PLoS Genet (2012) 8(1):e1002459. doi:10.1371/journal.pgen.1002459
80. Shah J, Desai PT, Chen D, Stevens JR, Weimer BC. Preadaptation to cold stress in Salmonella enterica serovar Typhimurium increases survival during subsequent acid stress exposure. Appl Environ Microbiol (2013) 79(23):7281–9. doi:10.1128/AEM.02621-13
81. Van der Poll T, Opal SM. Host-pathogen interactions in sepsis. Lancet Infect Dis (2008) 8(1):32–43. doi:10.1016/S1473-3099(07)70265-7
82. de Jong HK, van der Poll T, Wiersinga WJ. The systemic pro-inflammatory response in sepsis. J Innate Immun (2010) 2(5):422–30. doi:10.1159/000316286
83. Raffatellu M, Chessa D, Wilson RP, Tükel C, Akçelik M, Bäumler AJ. Capsule-mediated immune evasion: a new hypothesis explaining aspects of typhoid fever pathogenesis. Infect Immun (2006) 74(1):19–27. doi:10.1128/IAI.74.1.19-27.2006
84. Näsström E, Vu Thieu NT, Dongol S, Karkey A, Voong Vinh P, Ha Thanh T, et al. Salmonella Typhi and Salmonella Paratyphi A elaborate distinct systemic metabolite signatures during enteric fever. ELife (2014) 5:3. doi:10.7554/eLife.03100
85. Raffatellu M, Santos RL, Verhoeven DE, George MD, Wilson RP, Winter SE, et al. Simian immunodeficiency virus-induced mucosal interleukin-17 deficiency promotes Salmonella dissemination from the gut. Nat Med (2008) 14(4):421–8. doi:10.1038/nm1743
86. Gordon MA, Kankwatira AM, Mwafulirwa G, Walsh AL, Hopkins MJ, Parry CM, et al. Invasive non-typhoid Salmonellae establish systemic intracellular infection in HIV-infected adults: an emerging disease pathogenesis. Clin Infect Dis (2010) 50(7):953–62. doi:10.1086/651080
87. Santos RL, Zhang S, Tsolis RM, Kingsley RA, Adams LG, Bäumler AJ. Animal models of Salmonella infections: enteritis versus typhoid fever. Microbes Infect (2001) 3(14–15):1335–44. doi:10.1016/S1286-4579(01)01495-2
88. Kaiser P, Diard M, Stecher B, Hardt WD. The streptomycin mouse model for Salmonella diarrhea: functional analysis of the microbiota, the pathogen’s virulence factors, and the host’s mucosal immune response. Immunol Rev (2012) 245(1):56–83. doi:10.1111/j.1600-065X.2011.01070.x
89. Barthel M, Hapfelmeier S, Quintanilla-Martínez L, Kremer M, Rohde M, Hogardt M, et al. Pretreatment of mice with streptomycin provides a Salmonella enterica serovar Typhimurium colitis model that allows analysis of both pathogen and host. Infect Immun (2003) 71(5):2839–58. doi:10.1128/IAI.71.5.2839-2858.2003
90. Que JU, Hentges DJ. Effect of streptomycin administration on colonization resistance to Salmonella typhimurium in mice. Infect Immun (1985) 48(1):169–74.
91. Song J, Willinger T, Rongvaux A, Eynon EE, Stevens S, Manz MG, et al. A mouse model for the human pathogen Salmonella Typhi. Cell Host Microbe (2010) 8(4):369–76. doi:10.1016/j.chom.2010.09.003
92. Mathur R, Oh H, Zhang D, Park S-G, Seo J, Koblansky A, et al. A mouse model of Salmonella Typhi infection. Cell (2012) 151(3):590–602. doi:10.1016/j.cell.2012.08.042
93. Libby SJ, Brehm MA, Greiner DL, Shultz LD, McClelland M, Smith KD, et al. Humanized nonobese diabetic-SCID IL2rgammanull mice are susceptible to lethal Salmonella Typhi infection. Proc Natl Acad Sci USA (2010) 107(35):15589–94. doi:10.1073/pnas.1005566107
94. Bender JK, Wille T, Blank K, Lange A, Gerlach RG. LPS structure and PhoQ activity are important for Salmonella Typhimurium virulence in the Galleria mellonella infection model. PLoS One (2013) 8(10):e73287. doi:10.1371/journal.pone.0073287
95. Viegas SC, Mil-Homens D, Fialho AM, Arraiano CM. The virulence of Salmonella enterica serovar Typhimurium in the insect model Galleria mellonella is impaired by mutations in RNase E and RNase III. Appl Environ Microbiol (2013) 79(19):6124–33. doi:10.1128/AEM.02044-13
96. Torraca V, Masud S, Spaink HP, Meijer AH. Macrophage-pathogen interactions in infectious diseases: new therapeutic insights from the zebrafish host model. Dis Model Mech (2014) 7(7):785–97. doi:10.1242/dmm.015594
Keywords: gastroenteritis, host innate immunity, macrophages, NTS, pathogenicity islands, salmonellosis
Citation: Hurley D, McCusker MP, Fanning S and Martins M (2014) Salmonella–host interactions – modulation of the host innate immune system. Front. Immunol. 5:481. doi: 10.3389/fimmu.2014.00481
Received: 07 July 2014; Accepted: 20 September 2014;
Published online: 07 October 2014.
Edited by:
Constantino López-Macías, Mexican Social Security Institute, Mexico; University of Oxford, UKReviewed by:
Vida A. Dennis, Alabama State University, USAGuntram A. Grassl, Christian-Albrechts-University of Kiel, Germany
Edmundo Calva, Universidad Nacional Autónoma de México, Mexico
Copyright: © 2014 Hurley, McCusker, Fanning and Martins. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marta Martins, School of Public Health, Physiotherapy and Population Science, UCD Centre for Food Safety, UCD Centre for Molecular Innovation and Drug Discovery, University College Dublin, Room S1.06, Science Centre South, Belfield, Dublin 4, Ireland e-mail: marta.martins@ucd.ie