 Zhao Gao1,2†
Zhao Gao1,2† Jiao Hu1,2†Yanyan Liang1,2Qian Yang1,2Kun Yan1,2Dong Liu1,2Xiaoquan Wang1,2
Jiao Hu1,2†Yanyan Liang1,2Qian Yang1,2Kun Yan1,2Dong Liu1,2Xiaoquan Wang1,2 Min Gu1,2Xiaowen Liu1,2Shunlin Hu1,2Zenglei Hu1,2Huimou Liu1,2Wenbo Liu1,2Sujuan Chen1,2
Min Gu1,2Xiaowen Liu1,2Shunlin Hu1,2Zenglei Hu1,2Huimou Liu1,2Wenbo Liu1,2Sujuan Chen1,2 Daxin Peng1,2
Daxin Peng1,2 Xin-an Jiao3
Xin-an Jiao3 Xiufan Liu1,2*
Xiufan Liu1,2*- 1Animal Infectious Disease Laboratory, School of Veterinary Medicine, Yangzhou University, Yangzhou, China
- 2Jiangsu Co-innovation Center for Prevention and Control of Important Animal Infectious Diseases and Zoonosis, Jiangsu Key Laboratory of Zoonosis, Yangzhou University, Yangzhou, China
- 3Jiangsu Key Laboratory of Zoonosis, Jiangsu Co-Innovation Center for Prevention and Control of Important Animal Infectious Diseases and Zoonoses, Yangzhou University, Yangzhou, China
Accumulating data have identified the important roles of PA protein in replication and pathogenicity of influenza A virus (IAV). Identification of host factors that interact with the PA protein may accelerate our understanding of IAV pathogenesis. In this study, using immunoprecipitation assay combined with liquid chromatography-tandem mass spectrometry, we identified 278 human cellular proteins that might interact with PA of H5N1 IAV. Gene Ontology annotation revealed that the identified proteins are highly associated with viral translation and replication. Further KEGG pathway analysis of the interactome profile highlighted cellular pathways associated with translation, infectious disease, and signal transduction. In addition, Diseases and Functions analysis suggested that these cellular proteins are highly related with Organismal Injury and Abnormalities and Cell Death and Survival. Moreover, two cellular proteins (nucleolin and eukaryotic translation elongation factor 1-alpha 1) identified both in this study and others were further validated to interact with PA using co-immunoprecipitation and co-localization assays. Therefore, this study presented the interactome data of H5N1 IAV PA protein in human cells which may provide novel cellular target proteins for elucidating the potential molecular functions of PA in regulating the lifecycle of IAV in human cells.
Introduction
Influenza A virus (IAV) is one of the most important pathogens that causes acute respiratory disease and is responsible for relatively high morbidity and mortality every winter. In addition, IAV is a very adaptable virus and can infect a wide range of hosts (Webster et al., 1992; Olsen et al., 2006). IAV has an enveloped negative-strand RNA genome that is composed of eight viral RNA segments. IAV genome encodes at least 17 viral proteins, including 10 initially identified proteins (PB2, PB1, PA, HA, NP, NA, M1, M2, NS1, and NS2) and seven newly identified proteins PB1-F2 (Chen et al., 2001), PB1-N40 (Wise et al., 2009), PA-X (Jagger et al., 2012), PA-N155, PA-N182 (Muramoto et al., 2013), M42 (Wise et al., 2012), and NS3 (Selman et al., 2012). The PA protein is the third subunit of the trimeric polymerase complex which has multiple functions in the life cycle and pathogenesis of IAV. The N-terminal of PA possesses the endonuclease activity, cap binding, and promoter binding functions that impact the viral transcription and replication (Hara et al., 2006). Accumulating studies have also shown that adaptive mutations in PA are essential for IAV to cross host barrier (Seyer et al., 2012; Sun et al., 2014; Song et al., 2015). In addition, the PA protein is also a key player in virulence-determining of IAV (Hulse-Post et al., 2007; Song et al., 2011; Hu et al., 2013a) as well as host protein shut off (Desmet et al., 2013; Hu et al., 2013a; Llompart et al., 2014). However, the potential mechanism associated with the roles of PA in the life cycle and pathogenesis of IAV is largely unknown.
Numerous studies have demonstrated that IAV heavily relies on host cellular proteins to complete its life cycle and pathogenicity (Hale et al., 2006; Gack et al., 2009; Cho et al., 2013; Gao et al., 2015). The protein-protein interaction is a crucial manner to maintain the connection between host and IAV. Several studies have identified some host factors interacting with the influenza PA protein. HAX1, a cytoplasmic protein with anti-apoptotic function, is associated with the nuclear localization signal domain of PA (Hsu et al., 2013). In IAV-infected cells, HAX1 can impede nuclear accumulation of PA and inhibit IAV replication, indicating that host cells use HAX1-PA as a defense mechanism to limit IAV infection. In addition, hCLE is another interacting partner of PA through binding with two domains of PA (amino acid residue 493–512 and 557–574), and this interaction involves in modulating the RNA polymerase II activity (Huarte et al., 2001; Pérez-González et al., 2006). In the process of influenza virus infection, the hCLE-PA interaction contributes to the increment of viral polymerase activity, viral RNA transcription and replication, virus titer, and viral particle production (Rodriguez et al., 2011). PA also has a close interaction with MCM complex that is considered to be a host regulator of viral genome replication (Kawaguchi and Nagata, 2007). Furthermore, other host proteins, such as RanBP5, AIFMI, NPM, and RIG-I also have been identified as interacting partners of PA and influence the viral life cycle of IAV (Deng et al., 2006; Mayer et al., 2007; Bradel-Tretheway et al., 2011; Li et al., 2014). However, based on the important roles of PA in the life cycle of IAV and close interplay between PA and host machinery, other potential host factors interacting with the PA protein still need to be identified.
In this study, immunoprecipitation (IP) technique coupled with liquid chromatography-tandem mass spectrometry (LC-MS/MS) was used to characterize the host cellular proteins interacting with the PA protein of H5N1 IAV. We identified two hundred and seventy eight human cellular proteins as interacting factors with PA. Further in-depth functional analysis suggested that most of these proteins involve in viral replication, gene expression and viral infectious cycle. Moreover, Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis also demonstrated the crucial roles of these proteins in translation, infectious disease, and signal transduction. Of note, we selected two host proteins, nucleolin (NCL), and eukaryotic translation elongation factor 1-alpha 1 (eEF1A1) and confirmed their interaction with the PA protein using co-IP and co-localization tests. However, further studies are still needed to explore the potential roles of these two proteins in the life cycle of IAV.
Materials and Methods
Ethics Statement
All experiments involving live viruses were performed in negative-pressure isolators with HEPA filters in a biosafety level 3 (BSL3) facilities in accordance with the institutional biosafety manual.
Viruses and Cells
Highly pathogenic H5N1 strain A/Chicken/Jiangsu/k0402/2010(CK10) that shows high virulence in mouse was isolated from a dead chicken (Hu et al., 2013b). A549 (human type II alveolar epithelial), 293T (human embryonic kidney) and MDCK (Madin-Darby canine kidney) cells were maintained in Dulbecco's modified Eagle's medium (DMEM; Life Technologies) supplemented with 10% fetal calf serum (FCS; Life Technologies) and 50 KU/L antibiotics and were cultured at 37°C under 5% CO2.
Construction of Plasmids
The PA gene was amplified using high fidelity DNA polymerase (Invitrogen) based on CK10 cDNA and cloned into pCDNA3.1 vector to generate pCDNA-PA. The NCL and eEF1A1 genes were amplified from A549 cells. The sequences of NCL and eEF1A1 have aligned with sequences from GenBank (accession No: NM005381.2, KJ891086). The NCL and eEF1A1genes were cloned into the pires-hrGFP-1a vector (with a 3 × Flag tag in the C-terminus) to generate pires-hrGFP-1a-NCL and pires-hrGFP-1a-eEF1A1, respectively. Meanwhile, these two genes were also cloned into the pCDNA3.1-Myc-C vector (with a 3 × Myc tag in the C-terminus) to generate pCDNA-Myc-NCL and pCDNA-Myc-eEF1A1, respectively. All the positive clones were further validated by sequencing.
Western Blot Analysis
Protein samples were separated by electrophoresis on 10% (w/v) SDS-PAGE and transferred to polyvinylidene fluoride (PVDF) membranes (Bio-Rad). The membranes were then blotted with corresponding antibodies. Subsequently, the membranes were washed four times with TBST (0.05% tween-20 in Tris-buffered Saline) and incubated with horseradish peroxidase (HRP) conjugated goat-anti-mouse (Sigma-Aldrich) or goat-anti-rabbit IgG (Sigma-Aldrich). The enhanced chemiluminescence (ECL) system (Thermo) was utilized to detect the blotted proteins.
Determination of the PA Expression Level
A549 cells were seeded in 6-well plates and cultured overnight. Then the cells were infected with CK10 virus at a multiplicity of infection (MOI) of 1. CK10-infected A549 cells were harvested at 12, 24, 36, 48, and 60 h post-infection (p.i.). The protein samples were subjected to western blot assay using anti-PA antibody and anti-β-actin antibody subsequently. The protein bands were analyzed by Image J.
Statistical Analysis
Statistical analysis was performed using the SPSS statistics software. The independent samples T-test was used for data analysis. p < 0.05 was considered as significant.
IP and Co-IP
For IP, CK10-infected A549 cells were lysed in IP buffer and incubated at 4°C on a shaker for 30 min, followed by centrifugation at 12,000 rpm for 20 min. A total of 600 μl of the supernatant was incubated with anti-PA ployclonal antibody (GeneTex) or the irrespective IgG (Beyotime) at 4°C overnight. Protein A+G Agarose beads (Beyotime) were then added, and the mixture was incubated with gentle rocking at 4°C for 5 h. The beads were washed five times with cold IP buffer and eluted with Glycline-HCL (PH = 3.0) for further LC-MS/MS analysis.
For co-IP, the 293T cells were co-transfected with pCDNA-PA or pCDNA3.1 alone and pires-hrGFP-1a-NCL, pires-hrGFP-1a-eEF1A1, or pires-hrGFP-1a, respectively. The transfected cells were then lysed in IP buffer at 48 h post-transfection. The cells lysates were precipitated with appropriate antibodies in conjunction with beads as described above. The beads were then washed five times with cold IP buffer and boiled with 5 × SDS loading buffer (Cwbio) for 5 min. The immunoprecipitated proteins were then detected by western blot.
LC-MS/MS Analysis and Protein Identification
The LC-MS/MS was performed on a mass spectrometer coupled to Easy nLC (Thermo Fisher Scientific). A volume of 6 μl of each fraction was injected for nanoLC-MS/MS analysis. The peptide mixture was loaded onto a C18-reversed phase column (Thermo Scientific Easy Column, 2 cm long, 100 μm inner diameter,5 μm resin) in buffer A (0.1% Formic acid) and separated with a linear gradient of buffer B (80% acetonitrile and 0.1% Formic acid) at a flow rate of 300 nL/min controlled by IntelliFlow technology over 140 min. MS data was acquired using a data-dependent on Top10 method dynamically choosing the most abundant precursor ions from the survey scan (300–1800 m/z) for HCD fragmentation. Determination of the target value is based on predictive Automatic Gain Control (pAGC). Dynamic exclusion duration was 60 s. Survey scans were acquired at a resolution of 70,000 at m/z 200 and resolution for HCD spectra was set to 17,500 at m/z 200. Normalized collision energy was 30 eV and the underfill ratio, which specifies the minimum percentage of the target value likely to be reached at maximum fill time, was defined as 0.1%. The instrument was run with peptide recognition mode enabled. MS/MS spectra were searched using MASCOT engine (Matrix Science, version 2.2) against Universal Protein (UniProt) database.
Bioinformatics Analysis
The functional annotation and classification of all the identified proteins were performed using Gene Ontology (GO) analysis tool in the Database for Annotation Visualization and Integrated Discovery (DAVID) (version 6.7). KEGG pathway database was used for pathway analysis. The Protein-Protein interaction network was constructed using the Cytoscape software. The Diseases and Functions analysis was conducted using Ingenuity Pathway Analysis (IPA) software.
Confocal Microscopy Analysis
293T cells were seeded on coverslips in 24-well plates and cultured overnight, then co-transfected with pCDNA-PA or the empty vector (pCDNA3.1) alone and pCDNA-Myc-NCL, pCDNA-Myc-eEF1A1, or the empty vector (pCDNA-Myc-C), respectively. At 24 h post-transfection, the cells were fixed with 4% paraformaldehyde for 30 min at room temperature, and permeabilized with 0.1% Triton-X-100 for 15 min. After blocking in 5% bovine serum albumin (BSA) for 30 min, the cells were incubated for 1 h with the primary antibodies at 37°C. After washing in PBS for three times, the cells were stained with Alexa Fluor 555-conjugated goat anti-rabbit (Beyotime) and FITC-conjugated goat anti-mouse secondary antibodies (Beyotime) for 1 h at 37°C. After washing in PBS for three times, cell nuclei were stained with DAPI (Beyotime). The cells were then observed by using the Leica TCS SP8 STED 3X confocal microscope.
Results
Determination of the Peak of PA expression in the Process of IAV Infection
To determine the peak of PA expression, A549 cells that infected with CK10 virus at a MOI of 1 were harvested at 12, 24, 36, 48, and 60 h p.i. and the samples were then subjected to western blot assay by using anti-PA antibody as first antibody. At the same time, the expression patttern of β-actin protein was served as control. Three independent experiments were performed. The western blot protein bands were analyzed by Image J subsequently. And the results showed that the expression level of PA increased gradually in the process of CK10 infection, and reached a peak at 36 h p.i. The expression level of PA at 36 p.i. was significantly higher than those of other time points (Figures 1A,B). Therefore, we then collected the samples at 36 h p.i. for subsequent interactome analysis.
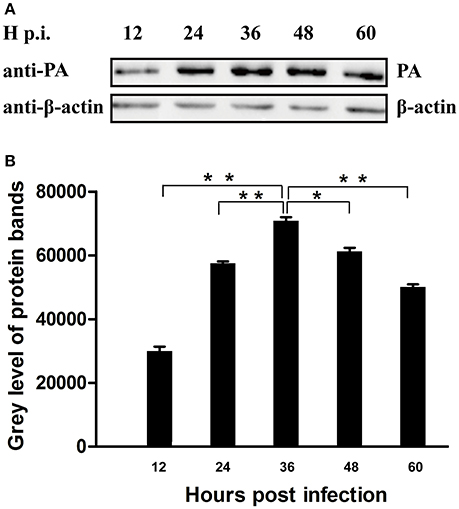
Figure 1. Determination of the peak of PA expression in the process of IAV infection. (A) Cell lysate from CK10-infected A549 cells at indicated time points were subjected to Western blot using anti-PA antibody and anti-β-actin antibody. Three independent experiments were perfomed and this is the representative data. (B) The Image J software was used to determine the expression level of the PA protein. Values shown are the gray level of the PA protein bands of western blot experiments ± SD of the results from three independent experiments. *p < 0.05 and **p < 0.01, when compared to the indicated group.
Identification of Host Cellular Proteins Interacting with the PA Protein
To efficiently precipitate the PA protein from the virus-infected cells and identify the host proteins that interact with PA, A549 cells were infected with CK10 at a MOI of 1. The cells were then harvested at 36 h p.i. and were immunopricicipitated with ployclonal antibody against PA or the irrespective antibody as control. Three independent experiments were carried out for both experimental group and control group. Subsequently, all samples were subjected to LC-MS/MS analysis. In addition, all proteins present in the negative controls were excluded and only proteins appeared at least twice in the triplicate analyses were reserved to generate interaction data. As a result, 278 proteins specifically precipitated with PA antibody were detected by LC-MS/MS analysis when compared to those precipitated with the irrespective IgG. A detailed summary of these proteins were given in Table 1 and Table S1.
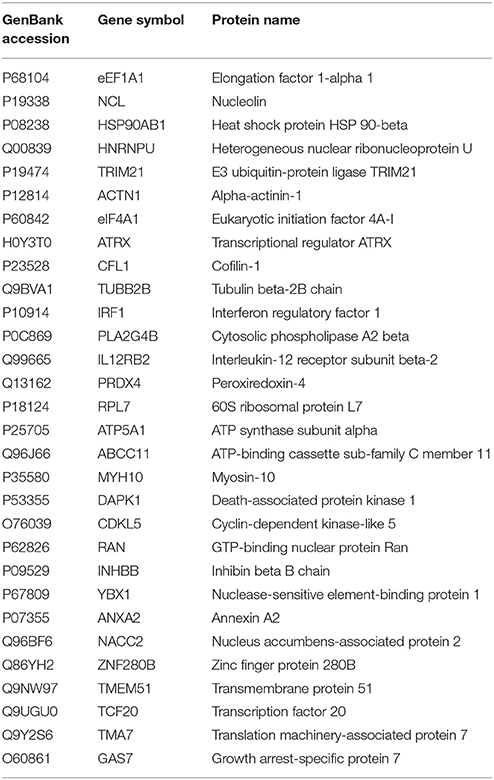
Table 1. PA-host interacting proteins in human cells (selected proteins).
Functional Analysis of All PA Interacting Proteins
To obtain an overall functional profile of the interactome associated with the PA protein, the 278 identified proteins were assigned for further bioinformatics analysis. As shown in Figure 2, three major types of annotations, biological process, cellular components, and molecular functions, were generated in the GO consortium website. In addition, we found that subclasses associated with gene expression, viral infectious cycle and viral transcription were highly enriched in biological process category (Figure 2A). As for the most enriched subclasses in cellular component, the major types, such as cytosol, integral to membrane, and ribonucleoprotein complex, were enriched (Figure 2B). Moreover, molecular function analysis showed that protein binding, ATP binding, structural constituent of ribosome and RNA binding were highly enriched (Figure 2C). A more detailed summary of GO annotation of the identified proteins was provided in Table S2.
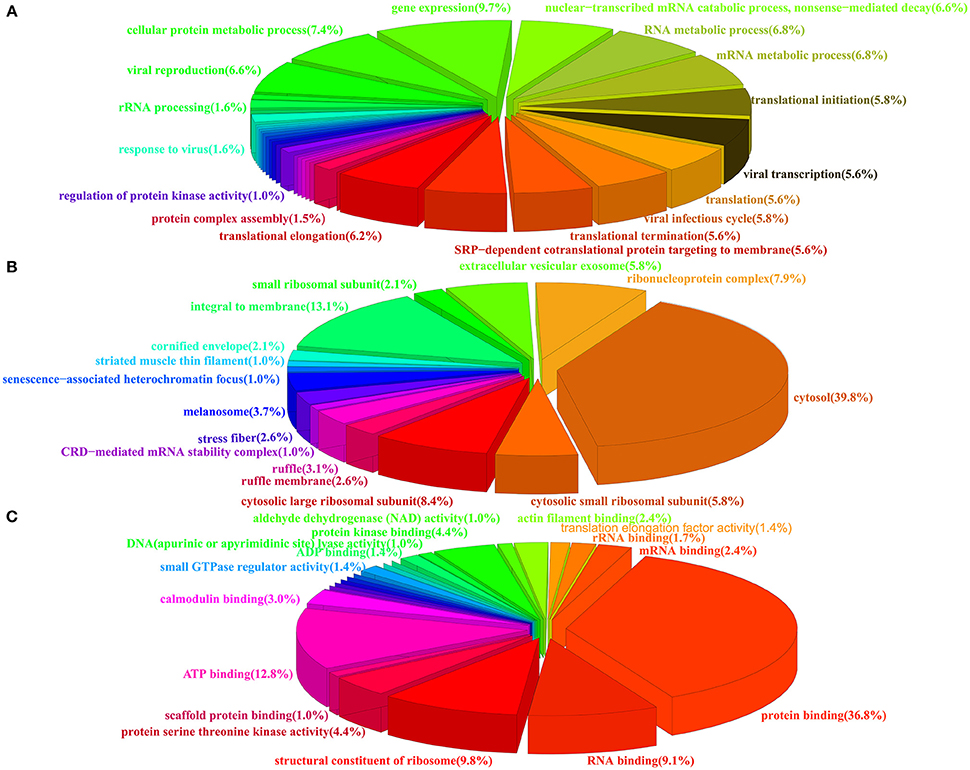
Figure 2. Pie charts showing the GO annotation of the identified cellular proteins. The GO annotation was analyzed by DAVID database and the percentage of each GO component was shown. (A) Biological process. (B) Cellular components. (C) Molecular function.
In addition, KEGG pathway analysis revealed an enrichment of 165 pathways related with 130 proteins of the infection network (Table S3). Six types of pathways were covered, including Human Diseases, Genetic Information Processing, Organismal Systems, Metabolism, Cellular Processes, and Environmental Information Processing (Figure 3A). To be noted, pathways involved in translation, infectious disease (contains infecious disease: Viral, infecious disease: Bacterial and infecious disease: Parasitic), cancers: overview and signal transduction were highly enriched in the PA-host interactome (Figure 3B, Table S3).
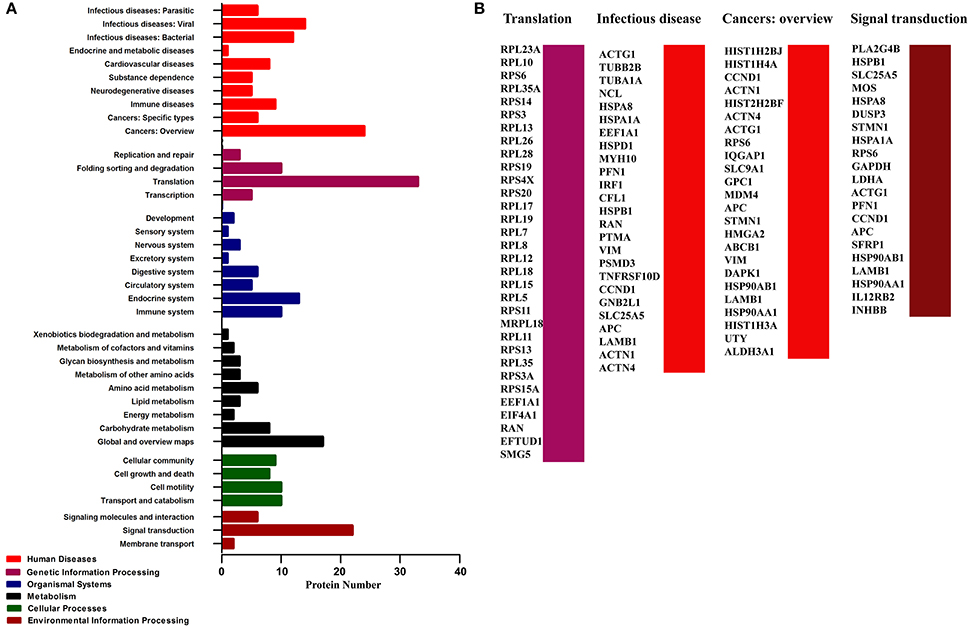
Figure 3. Pathway analysis of the cellular proteins interacting with PA based on KEGG. (A) Classification of the enriched KEGG pathways of the identified proteins. (B) Name of the identified proteins related with the top four KEGG pathways classification.
Detalied Analysis of the Two Targeted PA-Host Interacting Proteins
After detatied functional analysis, we then focused on two targeted novel cellular partners (NCL and eEF1A1) that might interact with PA, cellular proteins. As shown in Figure 4, the interactome profile indicated the important protein-protein interactions of the identified proteins associated with NCL and eEF1A1. The analysis revealed that NCL and eEF1A1 are cross-linked with several proteins with interesting interactions, including HNRNPA2B1-PTBP1-NCL-RPL5 and AHR-HSP90AA1-EEF1A1-RPL5. By GO annotation, we identified that these proteins have important functions in RNA transport, RNA splicing, apoptotic process, and translation which are closely asociated with IAV infection.
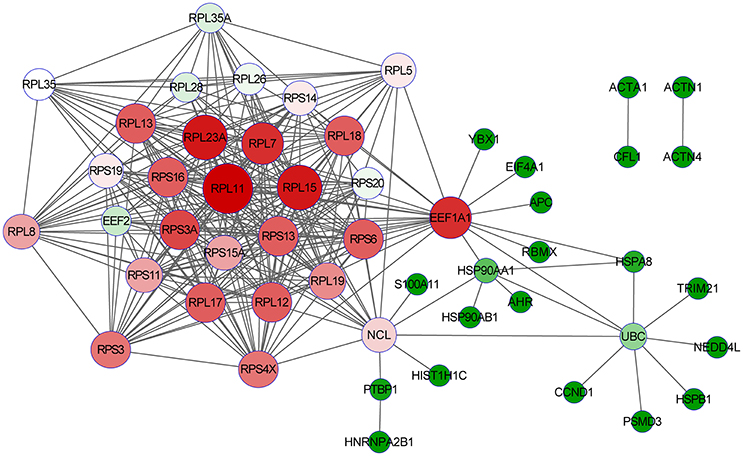
Figure 4. The interaction network of the identified proteins with NCL and eEF1A1. The open source Bioinformatics software Cytoscape 3.2.0 (http://www.cytoscape.org/) was used to visualize protein-protein interactions.
Meanwhile, the top Diseases and Functions analysis of the identified 278 cellular proteins was also performed using IPA software (Figures 5A,B, Figure S1). Surprisingly, we found that both NCL and eEF1A1 were related with Organismal Injury and Abnormalities. In addition, NCL was also involved in Cell Death and Survival. It has been demonstrated that immune injury and cell death contribute to the pathogenicity of IAV (Iwai et al., 2013; Duan and Thomas, 2016; Sridhar, 2016). Therefore, these resluts indicated that NCL and eEF1A1 might contribute to the virulence of IAV.
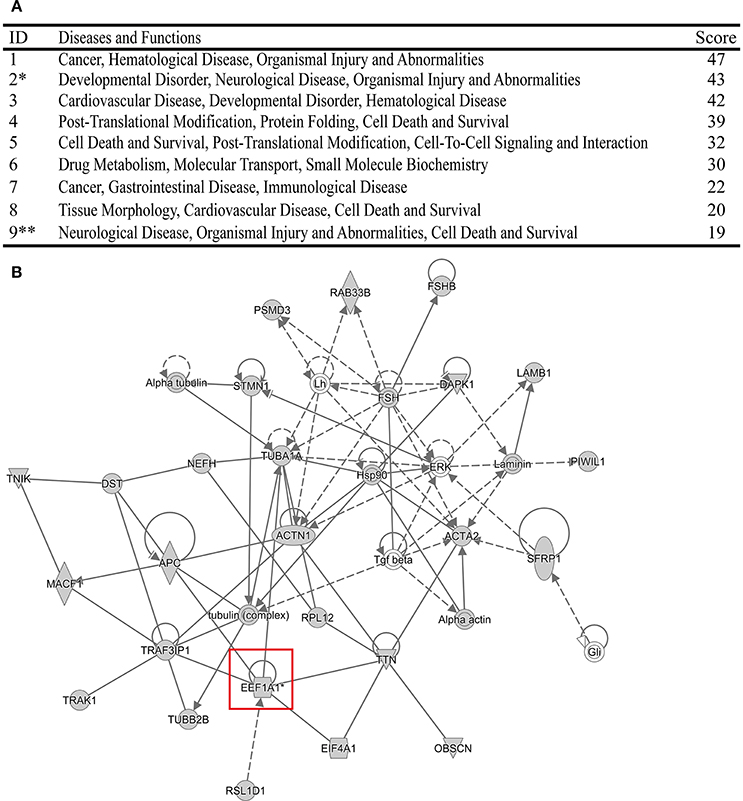
Figure 5. The top Diseases and Functions of the identified cellular proteins analyzed using IPA program. (A) *Stands for that the Diseases and Functions related with eEF1A1. **Stands for that the Diseases and Functions related with NCL. (B) Network of the Diseases and Functions related with eEF1A1.
Comparison of the Identifed PA-Host Interacting Proteins with the Available Interactome Data
In order to systematically analyze the screened PA-interacting host proteins, we then made a detailed comparison of the interactome data associated with PA from different studies. Using affinity purification-mass spectrometry (AP-MS), Wang et al. identified 134 H5N1 IAV PA interacting proteins in chicken cells (Wang et al., 2016). Watanabe et al. also characterized 304 proteins that interact with the PA protein of the H1N1 influenza virus in human cells using LC/MS (Watanabe et al., 2014). In the present study, 278 human proteins were found to interact with H5N1 PA protein using LC-MS/MS in A549 cells. As a result, our dataset shared 50 and 10 host proteins with data from H1N1-Human study and H5N1-Chicken study respectively (Figure 6A; Table S4). It is worth noting that NCL and eEF1A1 are identifed as PA-interacting factors both in our study and H1N1 study (Figure 6B). These resluts further indicated that these two proteins may play an important role in influenza virus infection.
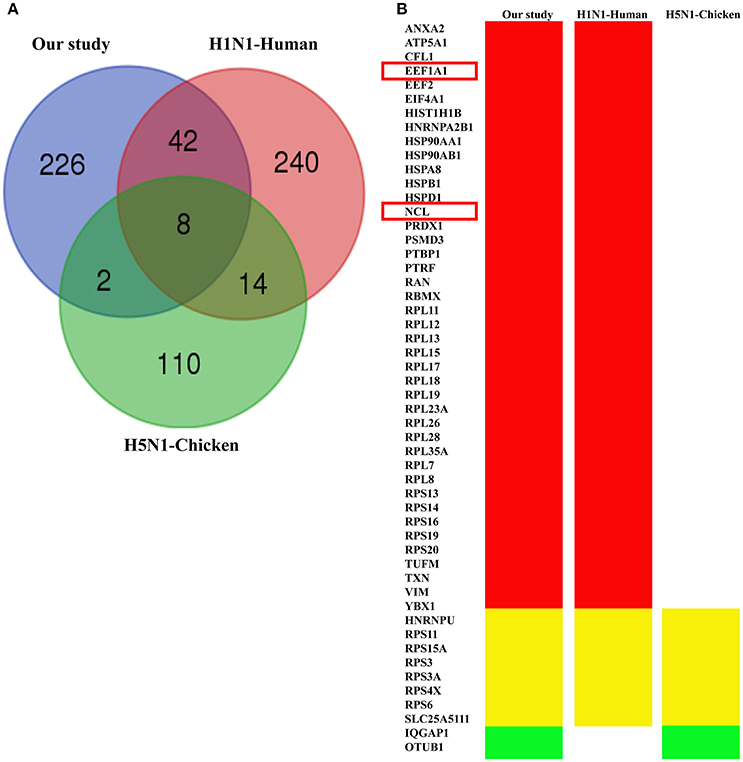
Figure 6. Shared host interacting factors of the PA protein across different hosts and IAV strains. (A) Venn diagram showing shared host interacting proteins with PA among our study, PA associated proteins of H1N1 IAV in the A549 cell line, and of H5N1 IAV in the DF1 cell line. (B) Name of the proteins shared by three different studies. Red stands for that the proteins only shared by our study and H1N1-Human study. Yellow represents the proteins shared by these three studies. Green stands for the proteins only shared by our study and H5N1-Chicken study. The NCL and eEF1A1 (highlighted in the red box) were found both in our study and H1N1-Human study.
Validation of Interaction between NCL, eEF1A1, and PA Using Co-IP and Co-Localization
Considering the NCL and eEF1A1 were shared in this study and the H1N1 study, we then verifed their interactions with the PA protein. 293T cells were co-transfected with pCDNA-PA or the empty vector (pCDNA3.1) alone with the Flag-tagged NCL expression plasmid (pires-hrGFP-1a-NCL), Flag-tagged eEF1A1 expression plasmid (pires-hrGFP-1a-eEF1A1) or the empty vector (pires-hrGFP-1a) respectively. After 48 h post-transfection, the co-IP assay was performed using anti-PA or anti-FLAG antibody. The immune-complexes were detected in western blot assay using anti-FLAG or anti-PA antibody subsequently. The results showed that both NCL and eEF1A1 were detected only in the presence of PA, but not in the presence of empty vector (Figures 7A,B). In addition, the PA protein was detected only in the presence of NCL and eEF1A1, but not in the presence of empty vector (Figures 7C,D).
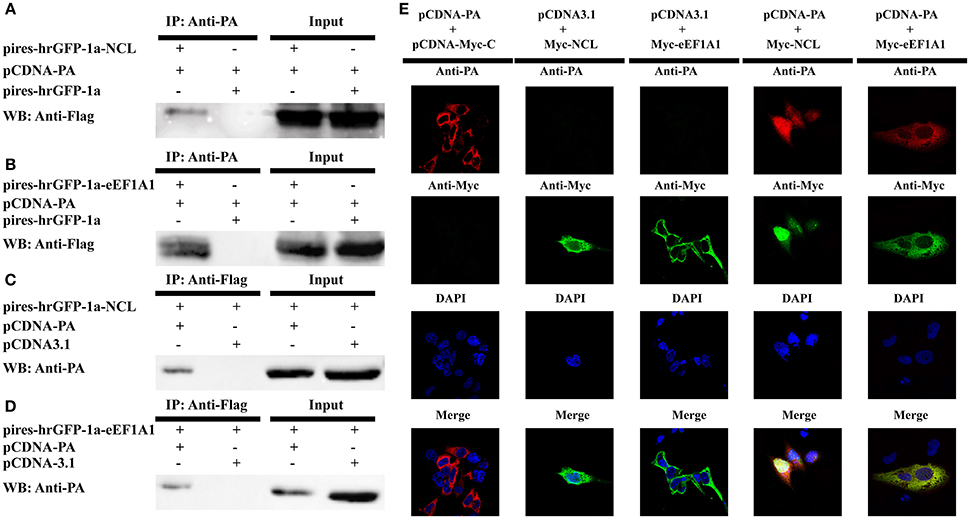
Figure 7. Confirmation of the interaction of IAV PA with NCL and eEF1A1. 293T cells were transiently co-transfected with the PA-expressing plasmid or the empty vector and the tagged NCL, eEF1A1 or the empty vector for co-immunoprecipitation and co-localization studies. (A–D) Cell lysates were prepared at 48 h post-transfection and the proteins were immunoprecipitated with anti-PA or anti-FLAG antibodies. Proteins in cell lysates (input) and immunoprecipitated samples were detected with the antibodies against FLAG or PA in Western blot. (A) Co-precipitation of about 110 kDa NCL protein with recombinant viral PA in cell lysate. (B) Co-precipitation of about 50 kDa eEF1A1 protein with PA in cell lysate. (C) Co-precipitation (reverse IP) of about 85 kDa PA with NCL. (D) Co-precipitation (reverse IP) of about 85 kDa PA with eEF1A1. (E) Confocal microscopy analysis was carried out for demonstrating colocalization of PA and NCL or eEF1A1. 293T cells were transiently co-transfected with PA expressing vector or empty vector and Myc-tagged NCL expressing vector, eEF1A1 expressing vector or empty vector, respectively. 24 h later, cells were fixed, and yellow regions are the areas of PA and NCL or eEF1A1 co-localization. Nuclei were stained using DAPI (blue).
To demonstrate the co-localization of NCL and PA or eEF1A1 and PA, three plasmid-pCDNA-Myc-NCL, pCDNA-Myc-eEF1A1, and pCDNA-PA were constructed. Cells grown on coverslips were co-transfected with pCDNA-PA or the empty vector (pCDNA3.1) alone with pCDNA-Myc-NCL, pCDNA-Myc-eEF1A1, or the empty vector (pCDNA-Myc-C), respectively. The cells were fixed at 24 h post-transfection. Then the cells were subjected to confocal microscopy analysis by using primary antibodies of Myc-tag, PA and secondary antibodies with two different fluorophores. Co-localization of two proteins in cells was visualized by confocal microscope. As shown in Figure 7E, NCL and PA were co-localized both in the nucleus and cytoplasm, while eEF1A1 and PA were only detected in the cytoplasm of the cell. Thus, these results clearly confirmed that the PA protein interacts well with the cellular proteins NCL or eEF1A1.
Discussion
Since the first report in Asia in 1997, H5N1 influenza viruses have spread to many countries on different continents and caused considerable loss to both poultry industry and human health (Webster and Govorkova, 2006). In addition, IAV can evade the host immune response and cause persistent infection (Quinones-Parra et al., 2014). However, until now, the mechanism of IAV infection is not fully elucidated. It is well-known that virus can infect hosts and survive in host cells through interacting with the host cellular factors, exploiting the cellular pathways, and subverting defense systems inhibiting viral propagation established by host cells (Lamkanfi and Dixit, 2010; Watanabe et al., 2010). PA, a subunit of RNA polymerase of IAV, possesses pleiotropic effects: (1) plays important roles in viral transcription and replication (Kawaguchi et al., 2005; Regan et al., 2006). (2) contributes to the adaption of IAV in mammalian host (Gabriel et al., 2005; Sakabe et al., 2011; Seyer et al., 2012). (3) increases the virulence of IAV (Hulse-Post et al., 2007; Song et al., 2011; Hu et al., 2013a). (4) involves in host protein shut off (Rodriguez et al., 2007; Desmet et al., 2013; Llompart et al., 2014). (5) participates in immune regulation in the process of IAV infection (Huang et al., 2013; Sakabe et al., 2013). Furthermore, three novel ORFs derived from the PA gene encoding PA-X (Jagger et al., 2012), PA-N155, and PA-N182 (Muramoto et al., 2013) have been identified. Especially, PA-X plays important roles in viral life cycle and pathogenesis (Hu et al., 2015). Therefore, illustrating the PA-host interactions that might contribute to viral pathogenesis of IAV is crucial for understanding the potential mechanisms of IAV infection and development of new anti-viral drugs.
Many technologies have been employed to characterize the protein interaction network, such as AP-MS and yeast two-hybrid (Y2H). Currently, compared with Y2H, AP-MS is more widely-used because it could reveal interactions that mimic the actual physiological condition. In this study, IP coupled LC-MS/MS technique was applied to explore the host factors in IAV-infected A549 cells that interact with PA of the H5N1 influenza virus. Compared with IP assay using PA plasmid-transfected cells, interactome data obtained from virus-infected cells could provide insights for this method not only conserved the native protein conformation during virus replication, but also able to explore the cellular proteins that might interact indirectly with PA protein.
In this study, 278 human cellular proteins were identified to interact with PA of H5N1 influenza virus (Table 1, Table S1). To further explore the biological significance of the interaction between PA and host cellular proteins, bioinformatics analysis was utilized to comprehensively evaluate and characterize the bio-functions of the identified host proteins. Results of sub-cellular locations and functional classes analysis based on GO analysis demonstrated that the identified proteins were highly associated with gene expression, viral translation, and replication (Figure 2).
KEGG pathway analysis showed that 130 of 278 identified host proteins involved in 165 pathways. Notably, a significant proportion of the enriched KEGG pathways, including 4 enriched KEGG pathways and 33 proteins, were shown to be associated with translation (Figure 3B, Table S3). Therefore, we surmised that the translation-related pathways enriched by PA-host interactions might play an important role in IAV infection. Regarding IAV life cycle, after entry into the host cells, the viral ribonucleoprotein complexes and viral RNA (vRNPs) are transported into the nucleus where replication occurs. In the nucleus, the polymerase also allows the transcription of the genome into mRNA, which is then transported back to the cytoplasm and translated into viral proteins. Then, the NP, PB1, PB2, and PA proteins will re-enter the nucleus to form the RNP complex with viral RNA (Berri et al., 2013). Therefore, our functional analysis data of the PA-host interaction proteins further confirmed the important role of PA in viral replication. The second highest proportion of enriched KEGG pathways was associated with infectious disease, including 16 enriched KEGG pathways and 25 proteins (Figure 3B, Table S3). Among the 25 proteins, ACTG1, HSPA8, TNFRSF10D, and HSPA1A are highly related with the pathway of IAV (Table S3). Therefore, we speculated that the remaining 21 proteins associated with infectious disease may play a shared role in influenza virus infection and other infectious pathogens. Thus, we surmised that studies in terms of other etiologies may offer good ways to investigate the pathogenesis of IAV.
Among the 278 identified host proteins, NCL and eEF1A1 were confirmed to interact with PA using co-IP and co-localization (Figure 7). NCL is a multi-functional protein that predominantly locates in the cell nucleolus. Although NCL is also found on the cell surface, it does not possess a transmembrane domain and therefore it may not act as a typical membrane protein (Hovanessian et al., 2000). Previous studies have demonstrated that NS1, HA, and NP proteins of IAV can also interact with NCL. The NS1 protein interacts with NCL in nucleoplasm and nucleolus mainly via its C-terminal NLS2/NoLS and N-terminal NLS1 domains (Melén et al., 2012). Chan et al. revealed that HA protein is also associated with NCL (Chan et al., 2016). Both inhibiting cell surface NCL and depleting endogenous NCL can substantially reduce influenza virus internalization. In addition, Kumar et al. identified the interaction between NCL and NP. Down-regulation of the host NCL expression facilitates the viral gene transcription, and the viral replication (Kumar et al., 2016). Moreover, it has been demonstrated that NCL also plays a significant role in the internalization of parainfluenza virus type 3 (Bose et al., 2004). Furthermore, surface NCL is also shown to be involved in the entry of several viruses, including respiratory syncytial virus (RSV), CrimeanCongo hemorrhagic fever virus (CCHFV), and human immunodeficiency virus (HIV) (Nisole et al., 2002; Tayyari et al., 2011; Xiao et al., 2011). However, the potential roles of the NCL in the PA-associated functions are currently unknown and need to be further confirmed in the future.
eEF1A1 is not only a translation factor but also a pleiotropic protein, including cytoskeleton modulation, chaperone-like activity, and regulation of cell proliferation and cell death (Abbas et al., 2015). Previous studies have indicated that eEF1A1 is a p53-interacting protein and has anti-apoptotic property in p53 family signaling pathway (Blanch et al., 2013). P53 also interplays with influenza virus, and down-regulation of p53 expression results in resisting host innate and adaptive immune system against IAV (Muñoz-Fontela et al., 2011; Terrier et al., 2011, 2012). However, the mechanism underlying this activity is still poorly understood. Therefore, we surmised that the p53 interacting protein eEF1A1 identified in this study might act as a p53 partner and facilitate the IAV infection. However, further studies are needed to verify this assumption.
In summary, in this study, 278 cellular proteins were identified as interacting partners for PA of H5N1 IAV. GO and KEGG pathway analysis revealed that these proteins played important roles in translation and viral replication. Further IPA analysis demonstrated that these host factors are associated with Organismal Injury and Abnormalities, Post-Translational Modification, Protein Folding, Cell Death, and Survival. Moreover, two targeted host proteins, NCL and eEF1A1, associated with Organismal Injury and Abnormalities and Cell Death and Survival, were further confirmed to interact with PA both using co-IP and co-localization analysis. Therefore, our findings provide new targets for further research of IAV infection and pathogenesis.
Author Contributions
Conceived and designed the experiments: ZG, JH and XFL. Performed the experiments: ZG, YYL, QY, KY and JH. Analyzed the data: ZG and JH. Contributed reagents/materials/analysis tools: DL, XQW, MG, ZLH, SLH, SJC, XWL, HML, DXP, XAJ and WBL. Wrote the paper: ZG, JH and XFL.
Funding
This work was supported by the National Natural Science Foundation of China (31502076), by the Jiangsu Provincial Natural Science Foundation of China (BK20150444), by the Natural Science Foundation of the Higher Education Institutions of Jiangsu Province, China (15KJB230006), by the Special Financial Grant from the China Postdoctoral Science Foundation (2016T90515), by the National key research and development project of China (2016YFD0501601 and 2016YFD0500202), by the National Key Technologies R&D Program of China (2015BAD12B01-3), by the earmarked fund for Modern Agro-industry Technology Research System (nycytx-41-G07) and by A Project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions, (PAPD).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2017.00739/full#supplementary-material
References
Abbas, W., Kumar, A., and Herbein, G. (2015). The eEF1A proteins: at the crossroads of oncogenesis, apoptosis, and viral infections. Front. Oncol. 5:75. doi: 10.3389/fonc.2015.00075
Berri, F., Lê, V. B., Jandrot-Perrus, M., Lina, B., and Riteau, B. (2013). Switch from protective to adverse inflammation during influenza: viral determinants and hemostasis are caught as culprits. Cell. Mol. Life Sci. 71, 885–898. doi: 10.1007/s00018-013-1479-x
Blanch, A., Robinson, F., Watson, I. R., Cheng, L. S., and Irwin, M. S. (2013). Eukaryotic translation elongation factor 1-alpha 1 inhibits p53 and p73 dependent apoptosis and chemotherapy sensitivity. PLoS ONE 8:e66436. doi: 10.1371/journal.pone.0066436
Bose, S., Basu, M., and Banerjee, A. K. (2004). Role of nucleolin in human parainfluenza virus type 3 infection of human lung epithelial cells. J. Virol. 78, 8146–8158. doi: 10.1128/JVI.78.15.8146-8158.2004
Bradel-Tretheway, B. G., Mattiacio, J. L., Krasnoselsky, A., Stevenson, C., Purdy, D., Dewhurst, S., et al. (2011). Comprehensive proteomic analysis of influenza virus polymerase complex reveals a novel association with mitochondrial proteins and RNA polymerase accessory factors. J. Virol. 85, 8569–8581. doi: 10.1128/JVI.00496-11
Chan, C. M., Chu, H., Zhang, A. J., Leung, L. H., Sze, K. H., Kao, R. Y., et al. (2016). Hemagglutinin of influenza A virus binds specifically to cell surface nucleolin and plays a role in virus internalization. Virology 494, 78–88. doi: 10.1016/j.virol.2016.04.008
Chen, W., Calvo, P. A., Malide, D., Gibbs, J., Schubert, U., Bacik, I., et al. (2001). A novel influenza A virus mitochondrial protein that induces cell death. Nat. Med. 7, 1306–1312. doi: 10.1038/nm1201-1306
Cho, H., Ahn, S. H., Kim, K. M., and Kim, Y. K. (2013). Non-structural protein 1 of influenza viruses inhibits rapid mRNA degradation mediated by double-stranded RNA-binding protein, staufen1. FEBS Lett. 587, 2118–2124. doi: 10.1016/j.febslet.2013.05.029
Deng, T., Engelhardt, O. G., Thomas, B., Akoulitchev, A. V., Brownlee, G. G., and Fodor, E. (2006). Role of ran binding protein 5 in nuclear import and assembly of the influenza virus RNA polymerase complex. J. Virol. 80, 11911–11919. doi: 10.1128/JVI.01565-06
Desmet, E. A., Bussey, K. A., Stone, R., and Takimoto, T. (2013). Identification of the N-terminal domain of the influenza virus PA responsible for the suppression of host protein synthesis. J. Virol. 87, 3108–3118. doi: 10.1128/JVI.02826-12
Duan, S., and Thomas, P. G. (2016). Balancing immune protection and immune pathology by CD8(+) T-cell responses to influenza infection. Front. Immunol. 7:25. doi: 10.3389/fimmu.2016.00025
Gabriel, G., Dauber, B., Wolff, T., Planz, O., Klenk, H. D., and Stech, J. (2005). The viral polymerase mediates adaptation of an avian influenza virus to a mammalian host. Proc. Natl. Acad. Sci. U.S.A. 102, 18590–18595. doi: 10.1073/pnas.0507415102
Gack, M. U., Albrecht, R. A., Urano, T., Inn, K. S., Huang, I. C., Carnero, E., et al. (2009). Influenza A virus NS1 targets the ubiquitin ligase TRIM25 to evade recognition by the host viral RNA sensor RIG-I. Cell Host Microbe 5, 439–449. doi: 10.1016/j.chom.2009.04.006
Gao, S., Wu, J., Liu, R. Y., Li, J., Song, L., Teng, Y., et al. (2015). Interaction of NS2 with AIMP2 facilitates the switch from ubiquitination to SUMOylation of M1 in influenza A virus-infected cells. J. Virol. 89, 300–311. doi: 10.1128/JVI.02170-14
Hale, B. G., Jackson, D., Chen, Y. H., Lamb, R. A., and Randall, R. E. (2006). Influenza A virus NS1 protein binds p85beta and activates phosphatidylinositol-3-kinase signaling. Proc. Natl. Acad. Sci. U.S.A. 103, 14194–14199. doi: 10.1073/pnas.0606109103
Hara, K., Schmidt, F. I., Crow, M., and Brownlee, G. G. (2006). Amino acid residues in the N-terminal region of the PA subunit of influenza A virus RNA polymerase play a critical role in protein stability, endonuclease activity, cap binding, and virion RNA promoter binding. J. Virol. 80, 7789–7798. doi: 10.1128/JVI.00600-06
Hovanessian, A. G., Puvion-Dutilleul, F., Nisole, S., Svab, J., Perret, E., Deng, J. S., et al. (2000). The cell-surface-expressed nucleolin is associated with the actin cytoskeleton. Exp. Cell Res. 261, 312–328. doi: 10.1006/excr.2000.5071
Hsu, W. B., Shih, J. L., Shih, J. R., Du, J. L., Teng, S. C., Huang, L. M., et al. (2013). Cellular protein HAX1 interacts with the influenza A virus PA polymerase subunit and impedes its nuclear translocation. J. Virol. 87, 110–123. doi: 10.1128/JVI.00939-12
Hu, J., Hu, Z., Mo, Y., Wu, Q., Cui, Z., Duan, Z., et al. (2013a). The PA and HA gene-mediated high viral load and intense innate immune response in the brain contribute to the high pathogenicity of H5N1 avian influenza virus in mallard ducks. J. Virol. 87, 11063–11075. doi: 10.1128/JVI.00760-13
Hu, J., Mo, Y., Wang, X., Gu, M., Hu, Z., Zhong, L., et al. (2015). PA-X decreases the pathogenicity of highly pathogenic H5N1 influenza A virus in avian species by inhibiting virus replication and host response. J. Virol. 89, 4126–4142. doi: 10.1128/JVI.02132-14
Hu, J., Zhao, K., Liu, X., Wang, X., Chen, Z., and Liu, X. (2013b). Two highly pathogenic avian influenza H5N1 viruses of clade 2.3.2.1 with similar genetic background but with different pathogenicity in mice and ducks. Transbound. Emerg. Dis. 60, 127–139. doi: 10.1111/j.1865-1682.2012.01325.x
Huang, C. H., Chen, C. J., Yen, C. T., Yu, C. P., Huang, P. N., Kuo, R. L., et al. (2013). Caspase-1 deficient mice are more susceptible to influenza A virus infection with PA variation. J. Infect. Dis. 208, 1898–1905. doi: 10.1093/infdis/jit381
Huarte, M., Sanz-Ezquerro, J. J., Roncal, F., Ortin, J., and Nieto, A. (2001). PA subunit from influenza virus polymerase complex interacts with a cellular protein with homology to a family of transcriptional activators. J. Virol. 75, 8597–8604. doi: 10.1128/JVI.75.18.8597-8604.2001
Hulse-Post, D. J., Franks, J., Boyd, K., Salomon, R., Hoffmann, E., Yen, H. L., et al. (2007). Molecular changes in the polymerase genes (PA and PB1) associated with high pathogenicity of H5N1 influenza virus in mallard ducks. J. Virol. 81, 8515–8524. doi: 10.1128/JVI.00435-07
Iwai, A., Shiozaki, T., and Miyazaki, T. (2013). Relevance of signaling molecules for apoptosis induction on influenza A virus replication. Biochem. Biophys. Res. Commun. 441, 531–537. doi: 10.1016/j.bbrc.2013.10.100
Jagger, B. W., Wise, H. M., Kash, J. C., Walters, K. A., Wills, N. M., Xiao, Y. L., et al. (2012). An overlapping protein-coding region in influenza A virus segment 3 modulates the host response. Science 337, 199–204. doi: 10.1126/science.1222213
Kawaguchi, A., and Nagata, K. (2007). De novo replication of the influenza virus RNA genome is regulated by DNA replicative helicase, MCM. EMBO J. 26, 4566–4575. doi: 10.1038/sj.emboj.7601881
Kawaguchi, A., Naito, T., and Nagata, K. (2005). Involvement of influenza virus PA subunit in assembly of functional RNA polymerase complexes. J. Virol. 79, 732–744. doi: 10.1128/JVI.79.2.732-744.2005
Melén, K., Tynell, J., Fagerlund, R., Roussel, P., Hernandez-Verdun, D., and Julkunen, I. (2012). Influenza A H3N2 subtype virus NS1 protein targets into the nucleus and binds primarily via its C-terminal NLS2/NoLS to nucleolin and fibrillarin. Virol. J. 9:167 doi: 10.1186/1743-422X-9-167
Kumar, D., Broor, S., and Rajala, M. S. (2016). Interaction of host nucleolin with influenza A virus nucleoprotein in the early phase of infection limits the late viral gene expression. PLoS ONE 11:e0164146. doi: 10.1371/journal.pone.0164146
Lamkanfi, M., and Dixit, V. M. (2010). Manipulation of host cell death pathways during microbial infections. Cell Host Microbe 8, 44–54. doi: 10.1016/j.chom.2010.06.007
Li, W., Chen, H., Sutton, T., Obadan, A., and Perez, D. R. (2014). Interactions between the influenza A virus RNA polymerase components and retinoic acid-inducible gene I. J. Virol. 88, 10432–10447. doi: 10.1128/JVI.01383-14
Llompart, C. M., Nieto, A., and Rodriguez-Frandsen, A. (2014). Specific residues of PB2 and PA influenza virus polymerase subunits confer the ability for RNA polymerase II degradation and virus pathogenicity in mice. J. Virol. 88, 3455–3463. doi: 10.1128/JVI.02263-13
Mayer, D., Molawi, K., Martinez-Sobrido, L., Ghanem, A., Thomas, S., Baginsky, S., et al. (2007). Identification of cellular interaction partners of the influenza virus ribonucleoprotein complex and polymerase complex using proteomic-based approaches. J. Proteome Res. 6, 672–682. doi: 10.1021/pr060432u
Muñoz-Fontela, C., Pazos, M., Delgado, I., Murk, W., Mungamuri, S. K., Lee, S. W., et al. (2011). p53 serves as a host antiviral factor that enhances innate and adaptive immune responses to influenza A virus. J. Immunol. 187, 6428–6436. doi: 10.4049/jimmunol.1101459
Muramoto, Y., Noda, T., Kawakami, E., Akkina, R., and Kawaoka, Y. (2013). Identification of novel influenza A virus proteins translated from PA mRNA. J. Virol. 87, 2455–2462. doi: 10.1128/JVI.02656-12
Nisole, S., Krust, B., and Hovanessian, A. G. (2002). Anchorage of HIV on permissive cells leads to coaggregation of viral particles with surface nucleolin at membrane raft microdomains. Exp. Cell Res. 276, 155–173. doi: 10.1006/excr.2002.5522
Olsen, B., Munster, V. J., Wallensten, A., Waldenström, J., Osterhaus, A. D., and Fouchier, R. A. (2006). Global patterns of influenza A virus in wild birds. Science 312, 384–388. doi: 10.1126/science.1122438
Pérez-González, A., Rodriguez, A., Huarte, M., Salanueva, I. J., and Nieto, A. (2006). hCLE/CGI-99, a human protein that interacts with theinfluenza virus polymerase, is a mRNA transcriptionmodulator. J. Mol. Biol. 362, 887–900. doi: 10.1016/j.jmb.2006.07.085
Quinones-Parra, S., Loh, L., Brown, L. E., Kedzierska, K., and Valkenburg, S. A. (2014). Universal immunity to influenza must outwit immune evasion. Front. Microbiol. 5:285. doi: 10.3389/fmicb.2014.00285
Regan, J. F., Liang, Y., and Parslow, T. G. (2006). Defective assembly of influenza A virus due to a mutation in the polymerase subunit PA. J. Virol. 80, 252–261. doi: 10.1128/JVI.80.1.252-261.2006
Webster, R. G., Bean, W. J., Gorman, O. T., Chambers, T. M., and Kawaoka, Y. (1992). Evolution and ecology of influenza A viruses. Microbiol. Rev. 56, 152–179.
Rodriguez, A., Perez-Gonzalez, A., and Nieto, A. (2007). Influenza virus infection causes specific degradation of the largest subunit of cellular RNA polymerase II. J. Virol. 81, 5315–5324. doi: 10.1128/JVI.02129-06
Rodriguez, A., Perez-Gonzalez, A., and Nieto, A. (2011). Cellular human CLE/C14orf166 protein interacts with influenza virus polymerase and is required for viral replication. J. Virol. 85, 12062–12066. doi: 10.1128/JVI.00684-11
Sakabe, S., Ozawa, M., Takano, R., Iwastuki-Horimoto, K., and Kawaoka, Y. (2011). Mutations in PA, NP, and HA of a pandemic (H1N1) 2009 influenza virus contribute to its adaptation to mice. Virus Res. 158, 124–129. doi: 10.1016/j.virusres.2011.03.022
Sakabe, S., Takano, R., Nagamura-Inoue, T., Yamashita, N., Nidom, C. A., Quynh Le, M., et al. (2013). Differences in cytokine production in human macrophages and in virulence in mice are attributable to the acidic polymerase protein of highly pathogenic influenza A virus subtype H5N1. J. Infect. Dis. 207, 262–271. doi: 10.1093/infdis/jis523
Selman, M., Dankar, S. K., Forbes, N. E., Jia, J. J., and Brown, E. G. (2012). Adaptive mutation in influenza A virus non-structural gene is linked to host switching and induces a novel protein by alternative splicing. Emerg. Microbes Infect. 1:e42. doi: 10.1038/emi.2012.38
Seyer, R., Hrincius, E. R., Ritzel, D., Abt, M., Mellmann, A., Marjuki, H., et al. (2012). Synergistic adaptive mutations in the hemagglutinin and polymerase acidic protein lead to increased virulence of pandemic 2009 H1N1 influenza A virus in mice. J. Infect. Dis. 205, 262–271. doi: 10.1093/infdis/jir716
Song, J., Feng, H., Xu, J., Zhao, D., Shi, J., Li, Y., et al. (2011). The PA protein directly contributes to the virulence of H5N1 avian influenza viruses in domestic ducks. J. Virol. 85, 2180–2188. doi: 10.1128/JVI.01975-10
Song, J., Xu, J., Shi, J., Li, Y., and Chen, H. (2015). Synergistic effect of S224P and N383D substitutions in the PA of H5N1 Avian influenza virus contributes to mammalian adaptation. Sci. Rep. 5:10510. doi: 10.1038/srep10510
Sridhar, S. (2016). Heterosubtypic T-cell immunity to influenza in humans: challenges for universal T-cell influenza vaccines. Front. Immunol. 7:195. doi: 10.3389/fimmu.2016.00195
Sun, Y., Xu, Q., Shen, Y., Liu, L., Wei, K., Sun, H., et al. (2014). Naturally occurring mutations in the PA gene are key contributors to increased virulence of pandemic H1N1/09 influenza virus in mice. J. Virol. 88, 4600–4604. doi: 10.1128/JVI.03158-13
Tayyari, F., Marchant, D., Moraes, T. J., Duan, W., Mastrangelo, P., and Hegele, R. G. (2011). Identification of nucleolin as a cellular receptor for human respiratory syncytial virus. Nat. Med. 17, 1132–1135. doi: 10.1038/nm.2444
Terrier, O., Josset, L., Textoris, J., Marcel, V., Cartet, G., Ferraris, O., et al. (2011). Cellular transcriptional profiling in human lung epithelial cells infected by different subtypes of influenza A viruses reveals an overall down-regulation of the host p53 pathway. Virol. J. 8:285. doi: 10.1186/1743-422X-8-285
Terrier, O., Marcel, V., Cartet, G., Lane, D. P., Lina, B., Rosa-Calatrava, M., et al. (2012). Influenza A viruses control expression of proviral human p53 Isoforms p53β and Δ133p53α. J. Virol. 86, 8452–8460. doi: 10.1128/J.V.I.07143-11
Wang, Q., Li, Q., Liu, R., Zheng, M., Wen, J., and Zhao, G. (2016). Host cell interactome of PA protein of H5N1 influenza A virus in chicken cells. J. Proteomics 136, 48–54. doi: 10.1016/j.jprot.2016.01.018
Watanabe, T., Kawakami, E., Shoemaker, J. E., Lopes, T. J., Matsuoka, Y., Tomita, Y., et al. (2014). Influenza virus-host interactome screen as a platform for antiviral drug development. Cell Host Microbe 16, 795–805. doi: 10.1016/j.chom.2014.11.002
Watanabe, T., Watanabe, S., and Kawaoka, Y. (2010). Cellular networks involved in the influenza virus life cycle. Cell Host Microbe 7, 427–439. doi: 10.1016/j.chom.2010.05.008
Webster, R. G., and Govorkova, E. A. (2006). H5N1 influenza–continuing evolution and spread. N. Engl. J. Med. 355, 2174–2177. doi: 10.1056/NEJMp068205
Wise, H. M., Foeglein, A., Sun, J., Dalton, R. M., Patel, S., Howard, W., et al. (2009). A complicated message: identification of a novel PB1-related protein translated from influenza A virus segment 2 mRNA. J. Virol. 83, 8021–8031. doi: 10.1128/JVI.00826-09
Wise, H. M., Hutchinson, E. C., Jagger, B. W., Stuart, A. D., Kang, Z. H., Robb, N., et al. (2012). Identification of a novel splice variant form of the influenza A virus M2 ion channel with an antigenically distinct ectodomain. PLoS Pathog. 8:e1002998. doi: 10.1371/journal.ppat.1002998
Keywords: H5N1 IAV, PA protein, virus-host interaction, pathogenesis, mammalian adaptation
Citation: Gao Z, Hu J, Liang Y, Yang Q, Yan K, Liu D, Wang X, Gu M, Liu X, Hu S, Hu Z, Liu H, Liu W, Chen S, Peng D, Jiao X-a and Liu X (2017) Generation and Comprehensive Analysis of Host Cell Interactome of the PA Protein of the Highly Pathogenic H5N1 Avian Influenza Virus in Mammalian Cells. Front. Microbiol. 8:739. doi: 10.3389/fmicb.2017.00739
Received: 07 February 2017; Accepted: 10 April 2017;
Published: 28 April 2017.
Edited by:
Dongsheng Zhou, Beijing Institute of Microbiology and Epidemiology, ChinaReviewed by:
Stephen Kwok-Wing Tsui, The Chinese University of Hong Kong, Hong KongMeilin Jin, Huazhong Agricultural University, China
Jinhua Liu, China Agricultural University, China
Copyright © 2017 Gao, Hu, Liang, Yang, Yan, Liu, Wang, Gu, Liu, Hu, Hu, Liu, Liu, Chen, Peng, Jiao and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiufan Liu, xfliu@yzu.edu.cn
†These authors have contributed equally to this work.