 Anaïs Soares
Anaïs Soares Kévin Alexandre
Kévin Alexandre Manuel Etienne
Manuel Etienne- 1GRAM 2.0, EA 2656, Normandie University, UNIROUEN, Rouen, France
- 2Infectious Diseases Department, Rouen University Hospital, Rouen, France
Pseudomonas aeruginosa biofilm-related infections are difficult to treat with antibiotics. Along the different layers of the biofilm, the P. aeruginosa population is heterogeneous, exhibiting an extreme ability to adapt his metabolic activity to the local microenvironment. At the deepest layers of the biofilm is a subset of dormant cells, called persister cells. Though antimicrobial failure might be multifactorial, it is now demonstrated that these persister cells, genetically identical to a fully susceptible strain, but phenotypically divergent, are highly tolerant to antibiotics, and contribute to antimicrobial failure. By eradicating susceptible, metabolically active cells, antibiotics bring out pre-existing persister cells. The biofilm mode of growth creates microenvironment conditions that activate stringent response mechanisms, SOS response and toxin-antitoxin systems that render the bacterial population highly tolerant to antibiotics. Using diverse, not standardized, models of biofilm infection, a large panel of antibiotic regimen has been evaluated. They demonstrated that biofilm growth had an unequal impact of antibiotic activity, colistin and meropenem being the less impacted antibiotics. Different combination and sequential antimicrobial therapies were also evaluated, and could be partially efficient, but none succeeded in eradicating persister cells, so that non-antibiotic alternative strategies are currently under development. This article reviews the molecular mechanisms involved in antibiotic tolerance and persistence in P. aeruginosa biofilm infections. A review of the antimicrobial regimen evaluated for the treatment of P. aeruginosa biofilm infection is also presented. While tremendous progress has been made in the understanding of biofilm-related infections, alternative non-antibiotic strategies are now urgently needed.
Introduction
Due to more frequent use of medical implanted devices (i.e., pacemaker, prosthesis, catheter) the burden of biofilm-related infections increased last decades (Costerton et al., 1999; Donlan, 2001; Del Pozo, 2018; Stewart and Bjarnsholt, 2020). Even when pathogens are categorized susceptible to antibiotics by routine in vitro testing, these infections are difficult to cure with antibiotics. Bacterial biofilms can survive antibiotics due to impaired antibiotic diffusion, antibiotic efflux, nutrient and oxygen limitation, expression of biofilm-specific genetic mechanisms, selection of resistant mutants, or survival of tolerant cells (Fauvart et al., 2011; Lebeaux et al., 2014; Harms et al., 2016; Hall and Mah, 2017; Olivares et al., 2020). While resistance is supported by de novo mutation or horizontal gene transfer, and is associated with an elevated antibiotic MIC, tolerance is defined by the capacity of bacteria to survive (higher minimal duration killing of 99%) despite antibiotic exposure at concentrations above MIC, without any change in the MIC (Brauner et al., 2016; Pang et al., 2019). In the stress conditions of biofilm growth, some authors consider tolerance as the predominant cause of long-term bacterial survival and antibiotic failure (Ciofu and Tolker-Nielsen, 2019).
Pseudomonas aeruginosa is an opportunistic pathogen frequently involved in biofilm-related infections. Though P. aeruginosa has largely been used as a study model to explore tolerance mechanisms and to investigate treatment strategies in biofilm, there is to date, no guidelines for the treatment of P. aeruginosa biofilm infections. Moreover, most of in vitro data on persister cells come from studies of Escherichia coli.
In this context, we reviewed, in P. aeruginosa biofilm-related models, the molecular mechanisms underlying tolerant phenotypes responsible for antibiotic failure, and the different antibiotic approaches proposed to overcome these mechanisms. Neither the non-antibiotic alternatives, nor the combinations of antibiotics and non-antibiotic compounds, to treat P. aeruginosa biofilms, were not voluntarily discussed in this article to focus on antibiotic strategies.
Why a Subset of Bacterial Cells Survives in Biofilms?
Biofilm: A Heterogeneous and Dynamic Environment
As stated by Allison et al. (2011), even when composed of a unique bacteria species, the biofilm population cannot be considered as a homogeneous entity but rather as a set of several subpopulations. Schematically, two distinct subpopulations can be described in P. aeruginosa biofilms: a stalk-forming subpopulation situated at the substratum, and a cap-forming subpopulation on top (Haagensen et al., 2017). But as the different layers of the biofilm progressively grow, a gradient of nutrients and oxygen leads to phenotypic and metabolic bacterial diversity (Borriello et al., 2004; Werner et al., 2004; Williamson et al., 2012). As a consequence, gene expression and phenotypic characteristics are likely to vary inside the biofilm, both within the structure and over time (Lenz et al., 2008; Soares et al., 2019b). Therefore, a continuum of quasi-independent bacterial populations exhibiting different phenotypes (i.e., susceptible, resistant and tolerant cells) may co-evolve in biofilm, and differences in the spatiotemporal organization of cell death and antibiotic tolerance development within the biofilm are observed (Haagensen et al., 2017). But, molecular mechanisms and pathways explaining how and why some subsets of biofilm cells have evolved and survive antibiotics while others will be rapidly killed by antibiotics, remain incompletely elucidated (Allison et al., 2011).
More Tolerant Cells Than Resistant Ones: The Key Role of Persisters
Acquired resistance, usually related to genetic modifications, appears not to be the main pathway of antibiotic failure in biofilm, while tolerance seems to play a major part (Ciofu et al., 2017; Hall and Mah, 2017; Peyrusson et al., 2020). Indeed, studies evaluating bacterial killing in biofilm models exposed to antibiotics showed that, at first, most susceptible cells die. The surviving cell fraction then reaches a plateau (Kamble and Pardesi, 2020). This subpopulation is not made of resistant mutants but of tolerant cells called persister cells, a subpopulation exhibiting a transient reversible phenotype allowing (i) to survive significant antibiotic exposure without MIC change and (ii) to regrow after antibiotic exposure (Lewis, 2010; Balaban et al., 2019). Such a persistent phenotype could be a consequence of biofilm organization. Stressful biofilm environment sometimes associated with antibiotic pressure might reveal a subpopulation of persister cells that was initially present. Indeed, transcriptomic analysis of bacterial biofilm under antibiotic pressure showed that bacteria on the top, exposed to oxygen and nutriment, have an intense gene expression, while bacteria on the bottom have reduced transcriptomic activity and enter a dormant state (Stewart and Franklin, 2008). By eradicating susceptible bacteria first, antibiotic could act as a revealer of the persister subpopulation that would pre-exist before antibiotic exposure. Thus, antibiotic treatment might be a determinant, hostile and unfavorable stressor which could eradicate susceptible non-persister cells, and turn the scale toward the metabolic pathways of persistence (Balaban, 2004). Higher antibiotic pressures (doses and exposure times) might also select susceptible cells in biofilm to lead them to a persister phenotype. The successive steps driving to persister survival in biofilms stressed by antibiotic exposure are presented in Figure 1.
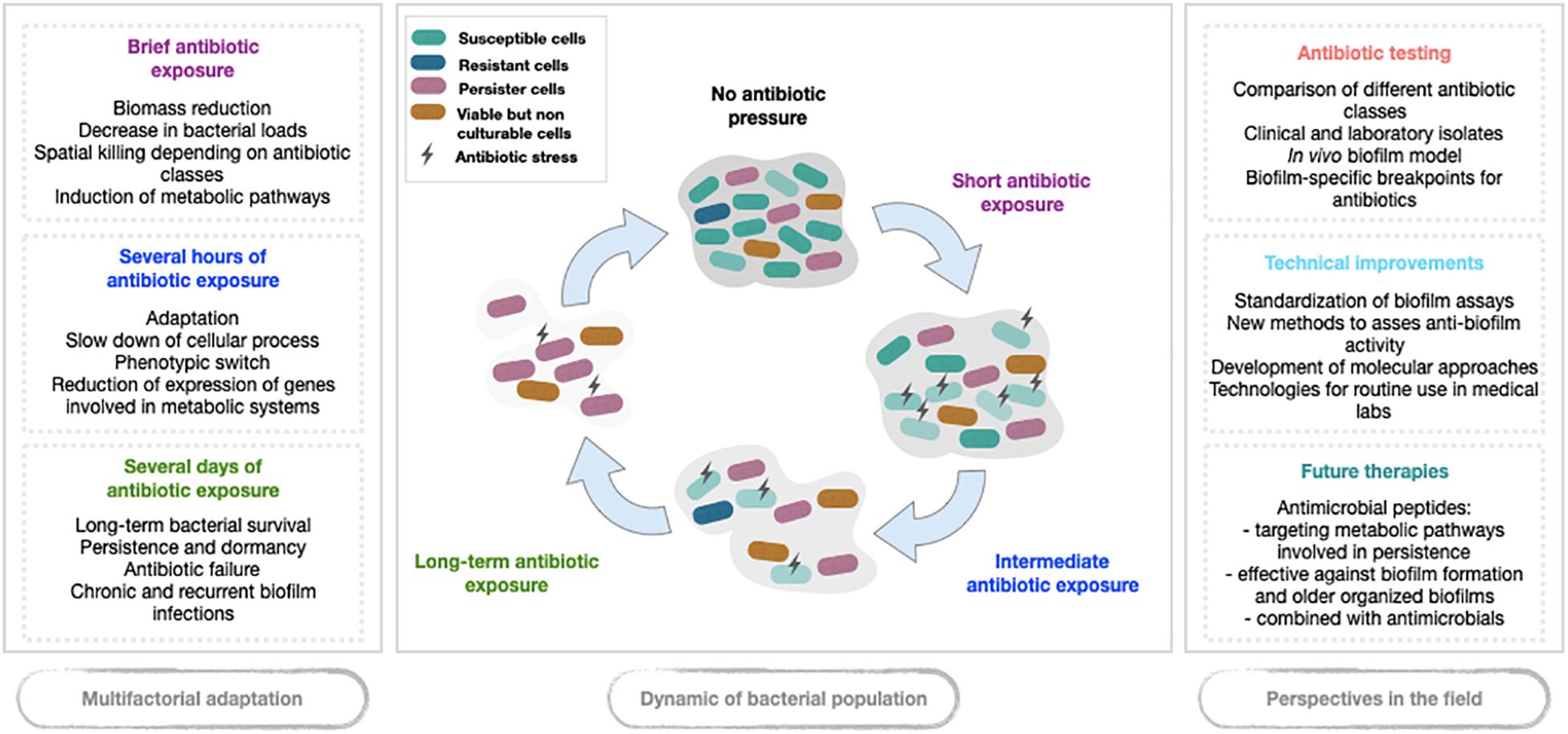
Figure 1. Schematic overview of phenotypic switch in biofilm under antibiotic pressure.
Pathways Toward Persistence in Biofilms
The Main Metabolic Pathways
Several regulatory pathways involved in P. aeruginosa persistence in biofilm have been identified through studies using mutants of genes involved in stringent or SOS response, and in Toxin/Anti-toxin (T/AT) systems (Murakami et al., 2005; Viducic et al., 2006; De Groote et al., 2009). Nutrient starvation in biofilm has been widely described as initiating the stringent response (Hauryliuk et al., 2015). Nguyen et al. (2011) infected mice with a wild-type P. aeruginosa strain or the corresponding mutant disrupted in relA and spoT, two genes encoding for two enzymes that regulate the (p)ppGpp metabolism, the key-component of stringent response. After 2 days, only 20% of mice survived, either infected by the wild-type or by the mutant strain. Interestingly, when mice were treated with ofloxacin, a 10% increase in survival was observed in mice infected by wild-type strain, while survival increased 50% in the ΔrelAΔspoT infected group. These results strongly suggest that a stringent response, regulated by RelA and SpoT, is involved in antibiotic tolerance. Biofilm environmental conditions could be a major trigger of stringent response, a mechanism contributing to the antibiotic tolerance of persister cells (Viducic et al., 2006).
The SOS response, which encompasses all the molecular mechanisms in response to DNA damages, also plays a role in bacterial persistence through different pathways for planktonic and biofilm culture conditions. In planktonic conditions, antibiotic exposure can induce the bacterial SOS response through the activation of the type I T/AT system. In planktonic cultures of Escherichia coli exposed to ciprofloxacin, knockout T/AT genes tisAB/tisR led to a decrease in persister cells level after 8 h of antibiotic exposure (Dörr et al., 2010). Dörr et al. (2009) hypothesized that SOS response might induce the expression of TisB toxin to a high level, which might cause a decrease in ATP cytosol concentrations, shutdown of the cellular metabolic and transcriptomic activity, finally leading to multidrug tolerance. It was also demonstrated in E. coli that two mutations in the hipA gene of the class II HipAB T/AT system led to a higher proportion of persister cells, particularly in stationary phase cultures (Keren et al., 2004; Schumacher et al., 2009, 2015). A very close finding was described more recently in P. aeruginosa by Li et al. (2016) in another class II HigBA T/AT system: an overexpression of higB gene increased by almost 1000 times the fraction of persister cells after exposure to sub-inhibitory ciprofloxacin concentrations in planktonic cultures. In biofilm conditions, Bernier et al. (2013) demonstrated in E. coli that ofloxacin persistence was directly related to the SOS response, and also to the maturity of the biofilm, but did not involve any of the SOS-induced toxin–antitoxin systems, as described above for planktonic conditions. To our knowledge, P. aeruginosa T/AT systems have not been explored in biofilms after antibiotic exposure at supra-MIC concentrations, and their role in antibiotic recalcitrance remains to be characterized.
Several others metabolic pathways might interact in response to antibiotic stress in biofilm. Stringent response might inhibit endogenous oxidative stress response. In the studies by Nguyen’s team (Nguyen et al., 2011; Khakimova et al., 2013), stringent response activation, in context of nutrient starvation, increased catalase activity. The production of hydroxyl radical was reduced and finally the cell killing by oxidative damages decreased. As already discussed by Hall and Mah (2017), quorum sensing might also be involved in antibiotic tolerance, as demonstrated in P. aeruginosa biofilms after tobramycin (Bjarnsholt et al., 2005; Brackman et al., 2011) and after meropenem treatments (Hazan et al., 2016). In these experiments, quorum-sensing deficient mutants had an increased susceptibility to antibiotics. Quorum-sensing, through a P. aeruginosa quorum-sensing-regulated molecule, might cause bacterial cell autolysis and DNA release. DNA subsequently would promote biofilm matrix and increase antibiotic tolerance. Nevertheless, studies on this topic are scarce and further works are required to better define the role that quorum sensing could play in biofilm tolerance to antibiotics.
Phenotypic Adaptation or Genetic Diversity?
Whether all pre-existing cells have the ability to evolve toward tolerant cells under stress conditions after a phenotypic switch, or a fraction of genetically determined cells can become predominant under antibiotic pressure is still debated. Cystic fibrosis airways exhibit very specific environmental conditions that favor biofilm bacterial growth (Sousa and Pereira, 2014). In this specific context, associated with very long-term P. aeruginosa colonization, patients usually undergo numerous antibiotic sequences that could favor genetic evolution of the bacterial biofilm population. Using a whole-genome analysis, Smith et al. (2006) demonstrated after 8 years of infection that the bacterial cells present in late P. aeruginosa infections of cystic fibrosis airways diverged from the cells originating the infections: virulence factors and DNA mismatch repair genes were mutated, resulting in the selection of “hypermutator” cells. In a shorter time-lapse, Klockgether et al. (2013) showed that genome sequences of P. aeruginosa strains from the airways of two patients with cystic fibrosis were almost identical, whereas transcriptome and metabolome were highly divergent, demonstrating the ability of genetically similar strains of P. aeruginosa to evolve and adapt to different hosts. Such finding was also demonstrated in a patient after a 2-week antipseudomonal treatment: a same strain evolved toward a more aerobic metabolism, via the downregulation of genes specifically involved in nitrogen metabolism (Fernández-Barat et al., 2017). Though not pathognomonic of persistence, similar phenotypic changes in the morphology of bacterial cells have been described in persister populations: high doses of aztreonam resulted in elongated filaments inside the biofilm (Rojo-Molinero et al., 2016); colistin persisters were elongated and had cell wall or membrane damages; DNA condensation was observed in amikacin persisters, and outer membrane vesicles were observed in ciprofloxacin persisters (Baek et al., 2020). These observations suggest that persister cells might develop through mechanisms depending on the antibiotic used, or on the environmental stress they go through.
Though the coexistence of genetic diversion and antibiotic tolerance through a phenotypic switch toward persistance has never been described, these two mechanisms should not be opposed and might even cooperate: the stressful conditions encountered in cystic fibrosis airways, the very long-term bacterial colonization of medical devices, the antibiotic pressure may favor emergence and sustainability of genetically divergent mutator strains, that might themselves promote transcriptomic changes and phenotypic diversity. To date, hypermutator strains have been described only in the very specific context of cystic fibrosis, whereas transcriptomic, metabolomic and phenotypic changes have been described in other biofilm conditions. The mechanisms underlying antibiotic tolerance might differ in these two kinds of biofilm-related infections and so the optimal antibiotic regimen might vary as well.
Antibiotic Strategies to Treat Biofilm Infections
The main studies assessing in vitro antibiotic activity in P. aeruginosa biofilms are summarized in Table 1. Very diverse in vitro biofilm models have been implemented using different culture media, biofilm incubation time, antibiotic concentrations, leading to heterogeneous results. The current lack of standardization of the methods, parameters and interpretation of results, limits the application of the obtained data to the clinical setting, including the comparison of different treatment strategies (Lebeaux et al., 2013; Macia et al., 2014). In a clinical therapeutic perspective, we focused our review on studies that exposed mature biofilms at clinically relevant concentrations.
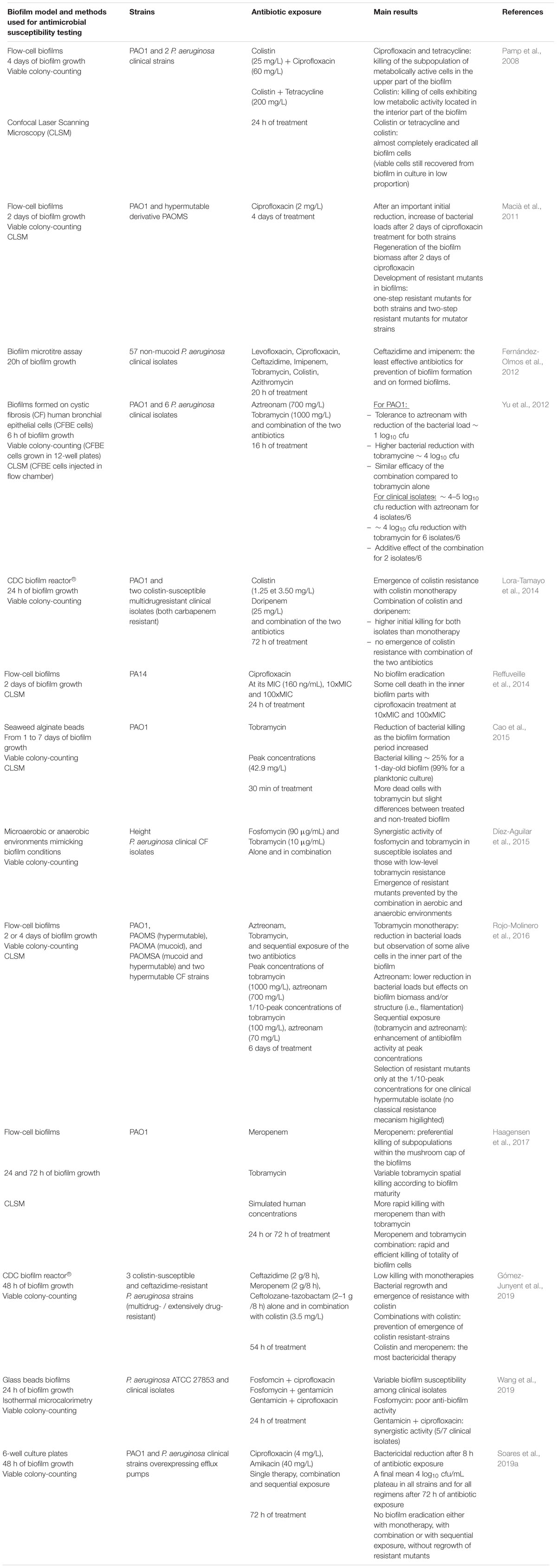
Table 1. Overview of antibiotic activity in Pseudomonas aeruginosa in vitro biofilms described in the literature.
Single Therapy
Eradication of bacterial biofilms requires considerably higher antibiotics concentrations compared to the killing of planktonic bacteria. In the study by Cao et al. (2015), more than 99% of the planktonic cells were killed by tobramycin, while in biofilm, the bacterial reduction was only 25%. Similarly, ciprofloxacin showed a remarkable bactericidal activity against P. aeruginosa in planktonic conditions (MIC = 0.25 mg/L and MBC = 1 mg/L), but exhibited a much lower activity in a microtiter biofilm model (minimum biofilm eradicating concentration [MBEC; the lowest concentration of an antibiotic that prevents visible growth in the recovery medium used to collect biofilm cells] = 512 mg/L) (Supplementary Table S1) (Wang et al., 2019). Besides, antibiotic in vitro activity against bacterial biofilm vary between isolates: ciprofloxacin MBEC against numerous P. aeruginosa cystic fibrosis clinical strains ranged from 4 to >1024 mg/L independently of their respective ciprofloxacin MIC (Wang et al., 2019). In a flow-cell biofilm model, tobramycin high dose (∼1000xMIC) demonstrated efficacy against a biofilm model of P. aeruginosa (Rojo-Molinero et al., 2016). Such high antibiotic concentrations can only be reached in rare clinical situations like in the sputum of cystic fibrosis patients after aerosolized drug administration (Ruddy et al., 2013). Nevertheless, for most antibiotic/isolate couples, the MBEC values may exceed the antibiotic concentrations that can be reached in a clinical practice.
As they act by inhibiting the synthesis of peptidoglycan, ß-lactam demonstrate more activity on dividing cells (i.e., in planktonic state) than on dormant cells (Ciofu and Tolker-Nielsen, 2019). In an in vitro biofilm device model, the biofilm growth had a strong impact on ceftazidime and imipenem with a 10 to 40 times increase in their minimal biofilm inhibitory concentrations (MBIC; the lowest concentration of an antibiotic that resulted in an OD650 nm difference of ≤10% of the mean of two positive growth-control well readings), whereas ciprofloxacin and tobramycin were less affected (2 to 4 times increase in their MBIC) (Fernández-Olmos et al., 2012). Among β-lactams, and for unknown reasons, only meropenem seems to have a sustained bactericidal activity against bacterial biofilm, as described in two different studies using in vitro pharmacodynamic models (Haagensen et al., 2017; Gómez-Junyent et al., 2019). Quinolones, aminoglycosides and colistin showed concentration-dependent or dose-dependent killing in biofilms with suitable MBIC/MIC ratios, respectively, of 1, 4, and 8 (Hengzhuang et al., 2011, 2012, 2013; Macia et al., 2014). Antibiotic exposure results in bacterial killing following a biphasic survival curve. After a rapid decrease, a small subset of cells reaches a plateau and survives to long-term antibiotic regimens. Indeed, confocal laser scanning microscopy studies showed that aminoglycosides, quinolones or colistin exposure strongly reduced the biomass of P. aeruginosa biofilms (Macià et al., 2011) but did not alter the biofilm structure (Rojo-Molinero et al., 2016). Thus, even if antibiotics triggered cell death in biofilm, they did not completely eradicate bacterial biofilms (Reffuveille et al., 2014), probably, as mentioned above, related to the heterogeneous metabolic activity within cells in biofilm environment and to the selection of persisters cells (Pamp et al., 2008; Haagensen et al., 2017).
Combination of Antibiotics
Since monotherapy did not eradicate mature biofilms, combination of antibiotics appeared as a therapeutic option for the treatment of biofilm-related infections.
In numerous in vitro studies, colistin was used as the backbone of combination antibiofilm therapies. Indeed, colistin based combinations demonstrated efficacy against P. aeruginosa biofilms and seemed effective for the eradication of persister cells and to prevent the emergence of colistin resistant mutants (Pamp et al., 2008; Lora-Tamayo et al., 2014; Gómez-Junyent et al., 2019; Baek et al., 2020). For example, biofilm cells exhibiting low metabolic activity were killed by colistin whereas ciprofloxacin eradicated only the subpopulation of metabolically active biofilm cells (Pamp et al., 2008). Since, among β-lactam based combinations, meropenem showed the highest efficacy against bactrial biofilm (Haagensen et al., 2017; Gómez-Junyent et al., 2019), a colistin/meropenem combination would theoretically be a most attractive therapeutic option for biofilm-related P. aeruginosa infection. Nevertheless, due to the frequent kidney toxicity of colistin and the emergence of resistance clones to colistin (mcr gene) and/or meropenem (carbapenem-resistant strain) this combination may not be suitable for all biofilm-related infections (Biswas et al., 2012; Marchaim et al., 2019). Aminoglycosides are widely used to treat P. aeruginosa infections (Mensa et al., 2018). Some aminoglycosides-based combinations showed anti-biofilm activity. Combination of meropenem and tobramycin resulted in a rapid and efficient killing of biofilm cells, initiating from the top cell layer of the biofilm (Haagensen et al., 2017). Gentamicin and ciprofloxacin were synergistic against a biofilm of P. aeruginosa using clinical strains (Wang et al., 2019). Fosfomycin and tobramycin were also proved to be synergistic against cystic fibrosis P. aeruginosa isolates under conditions simulating microaerobic or anaerobic environments that are present in biofilm-mediated infections (Díez-Aguilar et al., 2015). In other experiments, a lack of additive effect of other combinations compared with single therapy was noticed in biofilm. In a static in vitro model of biofilm, the ciprofloxacin/amikacin combination had no benefit over monotherapy against laboratory and clinical P. aeruginosa strains in biofilm, regarding efficacy and persister eradication (Soares et al., 2019a). A lack of antibiotic efficacy was also noticed for the tobramycin/aztreonam combination therapy that exhibited an additive effect on biofilm disruption only for two clinical strains out of six (Yu et al., 2012).
Finally, though some antibiotic combinations could prevent the emergence of resistant mutants in biofilm, none has proven a total efficacy in overcoming tolerance mechanisms or in eradicating persister cells.
Therapeutic and Technical Perspectives
Some authors proposed that antibiotic sequential exposure could eradicate persister cells. In planktonic cultures, Chung and Ko (2019) demonstrated that sequential treatment of colistin and amikacin, whatever the sequence, killed Acinetobacter baumanii persister cells. In a flow-cell model of biofilm study, Rojo-Molinero et al. (2016) used tobramycin as a substrate of the efflux pump MexXY-OprM, in order to increase the susceptibility to aztreonam. A sequential treatment of tobramycin and aztreonam was administered at very high concentrations in biofilm (reproducing the concentrations in the sputum of cystic fibrosis patients after aerosolized drug administration) and was more effective than monotherapy of tobramycin or aztreonam. But in another static model of biofilm, sequential exposure to ciprofloxacin and then amikacin or to amikacin and then ciprofloxacin (at antibiotic concentrations that allowed to achieve the PK/PD objectives associated with clinical efficacy after systemic administration) did not eradicate P. aeruginosa biofilm (Soares et al., 2019a). Besides, successive antibiotic exposures could promote an increasing proportion of persister cells, which could even become tolerant to additional antibiotic class (Wiuff et al., 2005; Michiels et al., 2016). Overall, these results suggest that a first antibiotic exposure may trigger the switch to a persister phenotype, rendering the bacterial population tolerant to all other antibiotic treatments.
Azithromycin showed potential inhibiting effects on P. aeruginosa biofilm with reducing of bacteria virulence factors, adhesion abilities and thus biofilm formation by inhibiting the cell-cell communication system, quorum sensing. Azithromycin seems to be of particular interest in case of cystic fibrosis to decrease long-term lung inflammation. The potential of this antibiotic is less described, and seems less appropriate, in case of biofilm device-related infection, diagnosed when biofilm is already mature (Nalca et al., 2006; Lutz et al., 2012; Olivares et al., 2020).
As a result, the non-antibiotic therapeutic perspectives, which target the metabolic pathways potentially involved in biofilm persistence, could be more relevant strategies (de la Fuente-Núñez et al., 2014; Pletzer et al., 2016). In the study by Reffuveille et al. (2014), an antimicrobial peptide, which acts by binding to the key component (p)ppGpp, could both inhibit biofilm formation and kill viable cells within a mature P. aeruginosa biofilm, in combination with ciprofloxacin. Such targeted approaches, though promising, have long been in the pipe-line and are still to be developed. Recent reviews (Koo et al., 2017; Defraine et al., 2018; Roy et al., 2018) offer comprehensive overviews of potential anti-persistence targets and strategies.
Nevertheless, there is a lack of high-quality clinical studies in the field of biofilm infections, both for cystic fibrosis patients and for tissue and device-related biofilm infections.
For further investigation of anti-biofilm activity of future components, it would be necessary to improve in vitro and in vivo biofilm models to monitor treatment efficacy. There is a real need for standardization of in vitro biofilm assays to reduce the variability in experimental results and to allow more direct comparisons between studies. New biofilm methods are required to assess the activity of future therapies but also to determine PK/PD parameters of existing antibiotics on young and old biofilms including combination antibiotic therapy (Figure 1).
Concluding Remarks
Finally, though efficient therapeutics are still far from current practice in the medical treatment of biofilm-related infections, tremendous progress has been made during the past decade regarding the knowledge on molecular mechanisms responsible for antibiotic failure. All recent studies confirm that persister cells, exhibiting a high level of tolerance to antibiotics, play a key-role in the ability of biofilms to escape antibiotics. The molecular mechanisms involved in persistence are still partially described, but dormancy, stringent and SOS response, are major contributors to persistence. The mechanisms involved in bacterial persistence seem to differ according to the bacterial strain, and to the nature of the biofilm, hence the therapeutic approach might also need to be adapted to the specificities of the biofilm considered. When it comes to treating P. aeruginosa biofilm infection, we still lack standardized in vitro methods that would be representative of in vivo infections and that would allow a comparison of different regimens. Moreover, despite intense in vitro studies on biofilm tolerance, we are disappointed by the lack of therapeutic translation. To date, no antibiotic regimen has proven efficacy to overcome persistence mechanisms in P. aeruginosa biofilms, and non-antibiotic alternatives are highly expected.
Author Contributions
AS reviewed the literature and wrote the manuscript. KA and ME reviewed the manuscript. All authors listed approved the work for publication.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2020.02057/full#supplementary-material
References
Allison, K. R., Brynildsen, M. P., and Collins, J. J. (2011). Heterogeneous bacterial persisters and engineering approaches to eliminate them. Curr. Opin. Microbiol. 14, 593–598. doi: 10.1016/j.mib.2011.09.002
Baek, M. S., Chung, E. S., Jung, D. S., and Ko, K. S. (2020). Effect of colistin-based antibiotic combinations on the eradication of persister cells in Pseudomonas aeruginosa. J. Antimicrob. Chemother. 75, 917–924. doi: 10.1093/jac/dkz552
Balaban, N. Q. (2004). Bacterial persistence as a phenotypic switch. Science 305, 1622–1625. doi: 10.1126/science.1099390
Balaban, N. Q., Helaine, S., Lewis, K., Ackermann, M., Aldridge, B., Andersson, D. I., et al. (2019). Definitions and guidelines for research on antibiotic persistence. Nat. Rev. Microbiol. 17, 441–448. doi: 10.1038/s41579-019-0196-3
Bernier, S. P., Lebeaux, D., DeFrancesco, A. S., Valomon, A., Soubigou, G., Coppée, J.-Y., et al. (2013). Starvation, together with the SOS response, mediates high biofilm-specific tolerance to the fluoroquinolone ofloxacin. PLoS Genet. 9:e1003144. doi: 10.1371/journal.pgen.1003144
Biswas, S., Brunel, J.-M., Dubus, J.-C., Reynaud-Gaubert, M., and Rolain, J.-M. (2012). Colistin: an update on the antibiotic of the 21st century. Expert Rev. Anti Infect. Ther. 10, 917–934. doi: 10.1586/eri.12.78
Bjarnsholt, T., Jensen, P. Ø, Burmølle, M., Hentzer, M., Haagensen, J. A. J., Hougen, H. P., et al. (2005). Pseudomonas aeruginosa tolerance to tobramycin, hydrogen peroxide and polymorphonuclear leukocytes is quorum-sensing dependent. Microbiol. Read. Engl. 151, 373–383. doi: 10.1099/mic.0.27463-0
Borriello, G., Werner, E., Roe, F., Kim, A. M., Ehrlich, G. D., and Stewart, P. S. (2004). Oxygen limitation contributes to antibiotic tolerance of Pseudomonas aeruginosa in Biofilms. Antimicrob. Agents Chemother. 48, 2659–2664. doi: 10.1128/AAC.48.7.2659-2664.2004
Brackman, G., Cos, P., Maes, L., Nelis, H. J., and Coenye, T. (2011). Quorum sensing inhibitors increase the susceptibility of bacterial biofilms to antibiotics In Vitro and In Vivo. Antimicrob. Agents Chemother. 55, 2655–2661. doi: 10.1128/AAC.00045-11
Brauner, A., Fridman, O., Gefen, O., and Balaban, N. Q. (2016). Distinguishing between resistance, tolerance and persistence to antibiotic treatment. Nat. Rev. Microbiol. 14, 320–330. doi: 10.1038/nrmicro.2016.34
Cao, B., Christophersen, L., Thomsen, K., Sonderholm, M., Bjarnsholt, T., Jensen, P. O., et al. (2015). Antibiotic penetration and bacterial killing in a Pseudomonas aeruginosa biofilm model. J. Antimicrob. Chemother. 70, 2057–2063. doi: 10.1093/jac/dkv058
Chung, E. S., and Ko, K. S. (2019). Eradication of persister cells of Acinetobacter baumannii through combination of colistin and amikacin antibiotics. J. Antimicrob. Chemother. 74, 1277–1283. doi: 10.1093/jac/dkz034
Ciofu, O., Rojo-Molinero, E., Macià, M. D., and Oliver, A. (2017). Antibiotic treatment of biofilm infections. APMIS 125, 304–319. doi: 10.1111/apm.12673
Ciofu, O., and Tolker-Nielsen, T. (2019). Tolerance and resistance of Pseudomonas aeruginosa Biofilms to antimicrobial agents-how P. aeruginosa can escape antibiotics. Front. Microbiol. 10:913. doi: 10.3389/fmicb.2019.00913
Costerton, J. W., Stewart, P. S., and Greenberg, E. P. (1999). Bacterial biofilms: a common cause of persistent infections. Science 284, 1318–1322.
De Groote, V. N., Verstraeten, N., Fauvart, M., Kint, C. I., Verbeeck, A. M., Beullens, S., et al. (2009). Novel persistence genes in Pseudomonas aeruginosa identified by high-throughput screening. FEMS Microbiol. Lett. 297, 73–79. doi: 10.1111/j.1574-6968.2009.01657.x
de la Fuente-Núñez, C., Reffuveille, F., Haney, E. F., Straus, S. K., and Hancock, R. E. W. (2014). Broad-spectrum anti-biofilm peptide that targets a cellular stress response. PLoS Pathog. 10:e1004152. doi: 10.1371/journal.ppat.1004152
Defraine, V., Fauvart, M., and Michiels, J. (2018). Fighting bacterial persistence: current and emerging anti-persister strategies and therapeutics. Drug Resist. Updat. 38, 12–26. doi: 10.1016/j.drup.2018.03.002
Del Pozo, J. L. (2018). Biofilm-related disease. Expert Rev. Anti Infect. Ther. 16, 51–65. doi: 10.1080/14787210.2018.1417036
Díez-Aguilar, M., Morosini, M. I., Tedim, A. P., Rodríguez, I., Aktaş, Z., and Cantón, R. (2015). Antimicrobial activity of fosfomycin-tobramycin combination against Pseudomonas aeruginosa Isolates assessed by time-kill assays and mutant prevention concentrations. Antimicrob. Agents Chemother. 59, 6039–6045. doi: 10.1128/AAC.00822-15
Donlan, R. M. (2001). Biofilm formation: a clinically relevant microbiological process. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 33, 1387–1392. doi: 10.1086/322972
Dörr, T., Lewis, K., and Vulić, M. (2009). SOS response induces persistence to fluoroquinolones in Escherichia coli. PLoS Genet. 5:e1000760. doi: 10.1371/journal.pgen.1000760
Dörr, T., Vuliæ, M., and Lewis, K. (2010). Ciprofloxacin causes persister formation by inducing the TisB toxin in Escherichia coli. PLoS Biol. 8:e1000317. doi: 10.1371/journal.pbio.1000317
Fauvart, M., De Groote, V. N., and Michiels, J. (2011). Role of persister cells in chronic infections: clinical relevance and perspectives on anti-persister therapies. J. Med. Microbiol. 60, 699–709. doi: 10.1099/jmm.0.030932-0
Fernández-Barat, L., Ciofu, O., Kragh, K. N., Pressler, T., Johansen, U., Motos, A., et al. (2017). Phenotypic shift in Pseudomonas aeruginosa populations from cystic fibrosis lungs after 2-week antipseudomonal treatment. J. Cyst. Fibros. 16, 222–229. doi: 10.1016/j.jcf.2016.08.005
Fernández-Olmos, A., García-Castillo, M., Maiz, L., Lamas, A., Baquero, F., and Cantón, R. (2012). In vitro prevention of Pseudomonas aeruginosa early biofilm formation with antibiotics used in cystic fibrosis patients. Int. J. Antimicrob. Agents 40, 173–176. doi: 10.1016/j.ijantimicag.2012.04.006
Gómez-Junyent, J., Benavent, E., Sierra, Y., El Haj, C., Soldevila, L., Torrejón, B., et al. (2019). Efficacy of ceftolozane/tazobactam, alone and in combination with colistin, against multidrug-resistant Pseudomonas aeruginosa in an in vitro biofilm pharmacodynamic model. Int. J. Antimicrob. Agents 53, 612–619. doi: 10.1016/j.ijantimicag.2019.01.010
Haagensen, J., Verotta, D., Huang, L., Engel, J., Spormann, A. M., and Yang, K. (2017). Spatiotemporal pharmacodynamics of meropenem- and tobramycin-treated Pseudomonas aeruginosa biofilms. J. Antimicrob. Chemother. 72, 3357–3365. doi: 10.1093/jac/dkx288
Hall, C. W., and Mah, T.-F. (2017). Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 41, 276–301. doi: 10.1093/femsre/fux010
Harms, A., Maisonneuve, E., and Gerdes, K. (2016). Mechanisms of bacterial persistence during stress and antibiotic exposure. Science 354:aaf4268. doi: 10.1126/science.aaf4268
Hauryliuk, V., Atkinson, G. C., Murakami, K. S., Tenson, T., and Gerdes, K. (2015). Recent functional insights into the role of (p)ppGpp in bacterial physiology. Nat. Rev. Microbiol. 13, 298–309. doi: 10.1038/nrmicro3448
Hazan, R., Que, Y. A., Maura, D., Strobel, B., Majcherczyk, P. A., Hopper, L. R., et al. (2016). Auto poisoning of the respiratory chain by a quorum-sensing-regulated molecule favors biofilm formation and antibiotic tolerance. Curr. Biol. 26, 195–206. doi: 10.1016/j.cub.2015.11.056
Hengzhuang, W., Ciofu, O., Yang, L., Wu, H., Song, Z., Oliver, A., et al. (2013). High β-Lactamase levels change the pharmacodynamics of β-Lactam antibiotics in Pseudomonas aeruginosa biofilms. Antimicrob. Agents Chemother. 57, 196–204. doi: 10.1128/AAC.01393-12
Hengzhuang, W., Wu, H., Ciofu, O., Song, Z., and Høiby, N. (2011). Pharmacokinetics/pharmacodynamics of colistin and imipenem on mucoid and nonmucoid Pseudomonas aeruginosa biofilms. Antimicrob. Agents Chemother. 55, 4469–4474. doi: 10.1128/AAC.00126-11
Hengzhuang, W., Wu, H., Ciofu, O., Song, Z., and Høiby, N. (2012). In Vivo pharmacokinetics/pharmacodynamics of colistin and imipenem in Pseudomonas aeruginosa biofilm infection. Antimicrob. Agents Chemother. 56, 2683–2690. doi: 10.1128/AAC.06486-11
Kamble, E., and Pardesi, K. (2020). Antibiotic tolerance in biofilm and stationary-phase planktonic cells of Staphylococcus aureus. Microb. Drug Resist. doi: 10.1089/mdr.2019.0425 [Epub ahead of print].
Keren, I., Shah, D., Spoering, A., Kaldalu, N., and Lewis, K. (2004). Specialized persister cells and the mechanism of multidrug tolerance in Escherichia coli. J. Bacteriol. 186, 8172–8180. doi: 10.1128/JB.186.24.8172-8180.2004
Khakimova, M., Ahlgren, H. G., Harrison, J. J., English, A. M., and Nguyen, D. (2013). The stringent response controls catalases in Pseudomonas aeruginosa and is required for hydrogen peroxide and antibiotic tolerance. J. Bacteriol. 195, 2011–2020. doi: 10.1128/JB.02061-12
Klockgether, J., Miethke, N., Kubesch, P., Bohn, Y.-S., Brockhausen, I., Cramer, N., et al. (2013). Intraclonal diversity of the Pseudomonas aeruginosa cystic fibrosis airway isolates TBCF10839 and TBCF121838: distinct signatures of transcriptome, proteome, metabolome, adherence and pathogenicity despite an almost identical genome sequence: genome and phenome diversity of P. aeruginosa clone TB. Environ. Microbiol. 15, 191–210. doi: 10.1111/j.1462-2920.2012.02842.x
Koo, H., Allan, R. N., Howlin, R. P., Stoodley, P., and Hall-Stoodley, L. (2017). Targeting microbial biofilms: current and prospective therapeutic strategies. Nat. Rev. Microbiol. 15, 740–755. doi: 10.1038/nrmicro.2017.99
Lebeaux, D., Chauhan, A., Rendueles, O., and Beloin, C. (2013). From in vitro to in vivo models of bacterial biofilm-related infections. Pathogens 2, 288–356. doi: 10.3390/pathogens2020288
Lebeaux, D., Ghigo, J.-M., and Beloin, C. (2014). Biofilm-related infections: bridging the gap between clinical management and fundamental aspects of recalcitrance toward antibiotics. Microbiol. Mol. Biol. Rev. 78, 510–543. doi: 10.1128/MMBR.00013-14
Lenz, A. P., Williamson, K. S., Pitts, B., Stewart, P. S., and Franklin, M. J. (2008). Localized gene expression in Pseudomonas aeruginosa biofilms. Appl. Environ. Microbiol. 74, 4463–4471. doi: 10.1128/AEM.00710-08
Lewis, K. (2010). Persister cells. Annu. Rev. Microbiol. 64, 357–372. doi: 10.1146/annurev.micro.112408.134306
Li, M., Long, Y., Liu, Y., Liu, Y., Chen, R., Shi, J., et al. (2016). HigB of Pseudomonas aeruginosa Enhances killing of phagocytes by up-regulating the Type III secretion system in ciprofloxacin induced persister cells. Front. Cell. Infect. Microbiol. 6:125. doi: 10.3389/fcimb.2016.00125
Lora-Tamayo, J., Murillo, O., Bergen, P. J., Nation, R. L., Poudyal, A., Luo, X., et al. (2014). Activity of colistin combined with doripenem at clinically relevant concentrations against multidrug-resistant Pseudomonas aeruginosa in an in vitro dynamic biofilm model. J. Antimicrob. Chemother. 69, 2434–2442. doi: 10.1093/jac/dku151
Lutz, L., Pereira, D., Paiva, R., Zavascki, A., and Barth, A. (2012). Macrolides decrease the minimal inhibitory concentration of anti-pseudomonal agents against Pseudomonas aeruginosa from cystic fibrosis patients in biofilm. BMC Microbiol. 12:196. doi: 10.1186/1471-2180-12-196
Macià, M. D., Pérez, J. L., Molin, S., and Oliver, A. (2011). Dynamics of mutator and antibiotic-resistant populations in a pharmacokinetic/pharmacodynamic model of Pseudomonas aeruginosa biofilm treatment. Antimicrob. Agents Chemother. 55, 5230–5237. doi: 10.1128/AAC.00617-11
Macia, M. D., Rojo-Molinero, E., and Oliver, A. (2014). Antimicrobial susceptibility testing in biofilm-growing bacteria. Clin. Microbiol. Infect. 20, 981–990. doi: 10.1111/1469-0691.12651
Marchaim, D., Kaye, D., and Kaye, K. S. (2019). Use of colistin in critically Ill patients. Adv. Exp. Med. Biol. 1145, 155–179. doi: 10.1007/978-3-030-16373-0_12
Mensa, J., Barberán, J., Soriano, A., Llinares, P., Marco, F., Cantón, R., et al. (2018). Antibiotic selection in the treatment of acute invasive infections by Pseudomonas aeruginosa: guidelines by the Spanish society of chemotherapy. Rev. Espanola Quimioter. Publ. Soc. Espanola Quimioter. 31, 78–100.
Michiels, J. E., Van den Bergh, B., Verstraeten, N., Fauvart, M., and Michiels, J. (2016). In Vitro emergence of high persistence upon periodic aminoglycoside challenge in the ESKAPE pathogens. Antimicrob. Agents Chemother. 60, 4630–4637. doi: 10.1128/AAC.00757-16
Murakami, K., Ono, T., Viducic, D., Kayama, S., Mori, M., Hirota, K., et al. (2005). Role for rpoS gene of Pseudomonas aeruginosa in antibiotic tolerance. FEMS Microbiol. Lett. 242, 161–167. doi: 10.1016/j.femsle.2004.11.005
Nalca, Y., Jänsch, L., Bredenbruch, F., Geffers, R., Buer, J., and Häussler, S. (2006). Quorum-sensing antagonistic activities of azithromycin in Pseudomonas aeruginosa PAO1: a global approach. Antimicrob. Agents Chemother. 50, 1680–1688. doi: 10.1128/AAC.50.5.1680-1688.2006
Nguyen, D., Joshi-Datar, A., Lepine, F., Bauerle, E., Olakanmi, O., Beer, K., et al. (2011). Active starvation responses mediate antibiotic tolerance in biofilms and nutrient-limited bacteria. Science 334, 982–986. doi: 10.1126/science.1211037
Olivares, E., Badel-Berchoux, S., Provot, C., Prévost, G., Bernardi, T., and Jehl, F. (2020). Clinical impact of antibiotics for the Treatment of Pseudomonas aeruginosa biofilm infections. Front. Microbiol. 10:2894. doi: 10.3389/fmicb.2019.02894
Pamp, S. J., Gjermansen, M., Johansen, H. K., and Tolker-Nielsen, T. (2008). Tolerance to the antimicrobial peptide colistin in Pseudomonas aeruginosa biofilms is linked to metabolically active cells, and depends on the pmr and mexAB-oprM genes. Mol. Microbiol. 68, 223–240. doi: 10.1111/j.1365-2958.2008.06152.x
Pang, Z., Raudonis, R., Glick, B. R., Lin, T.-J., and Cheng, Z. (2019). Antibiotic resistance in Pseudomonas aeruginosa: mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 37, 177–192. doi: 10.1016/j.biotechadv.2018.11.013
Peyrusson, F., Varet, H., Nguyen, T. K., Legendre, R., Sismeiro, O., Coppée, J.-Y., et al. (2020). Intracellular Staphylococcus aureus persisters upon antibiotic exposure. Nat. Commun. 11:2200. doi: 10.1038/s41467-020-15966-7
Pletzer, D., Coleman, S. R., and Hancock, R. E. (2016). Anti-biofilm peptides as a new weapon in antimicrobial warfare. Curr. Opin. Microbiol. 33, 35–40. doi: 10.1016/j.mib.2016.05.016
Reffuveille, F., de la Fuente-Núñez, C., Mansour, S., and Hancock, R. E. W. (2014). A broad-spectrum antibiofilm peptide enhances antibiotic action against bacterial biofilms. Antimicrob. Agents Chemother. 58, 5363–5371. doi: 10.1128/AAC.03163-14
Rojo-Molinero, E., Macià, M. D., Rubio, R., Moyà, B., Cabot, G., López-Causapé, C., et al. (2016). Sequential treatment of biofilms with aztreonam and tobramycin is a novel strategy for combating Pseudomonas aeruginosa chronic respiratory infections. Antimicrob. Agents Chemother. 60, 2912–2922. doi: 10.1128/AAC.00196-16
Roy, R., Tiwari, M., Donelli, G., and Tiwari, V. (2018). Strategies for combating bacterial biofilms: a focus on anti-biofilm agents and their mechanisms of action. Virulence 9, 522–554. doi: 10.1080/21505594.2017.1313372
Ruddy, J., Emerson, J., Moss, R., Genatossio, A., McNamara, S., Burns, J. L., et al. (2013). Sputum tobramycin concentrations in cystic fibrosis patients with repeated administration of inhaled tobramycin. J. Aerosol. Med. Pulm. Drug Deliv. 26, 69–75. doi: 10.1089/jamp.2011.0942
Schumacher, M. A., Balani, P., Min, J., Chinnam, N. B., Hansen, S., Vulić, M., et al. (2015). HipBA-promoter structures reveal the basis of heritable multidrug tolerance. Nature 524, 59–64. doi: 10.1038/nature14662
Schumacher, M. A., Piro, K. M., Xu, W., Hansen, S., Lewis, K., and Brennan, R. G. (2009). Molecular mechanisms of HipA-mediated multidrug tolerance and its neutralization by HipB. Science 323, 396–401. doi: 10.1126/science.1163806
Smith, E. E., Buckley, D. G., Wu, Z., Saenphimmachak, C., Hoffman, L. R., D’Argenio, D. A., et al. (2006). Genetic adaptation by Pseudomonas aeruginosa to the airways of cystic fibrosis patients. Proc. Natl. Acad. Sci. U.S.A. 103, 8487–8492. doi: 10.1073/pnas.0602138103
Soares, A., Alexandre, K., Lamoureux, F., Lemée, L., Caron, F., Pestel-Caron, M., et al. (2019a). Efficacy of a ciprofloxacin/amikacin combination against planktonic and biofilm cultures of susceptible and low-level resistant Pseudomonas aeruginosa. J. Antimicrob. Chemother. 74, 3252–3259. doi: 10.1093/jac/dkz355
Soares, A., Roussel, V., Pestel-Caron, M., Barreau, M., Caron, F., Bouffartigues, E., et al. (2019b). Understanding ciprofloxacin failure in pseudomonas aeruginosa biofilm: persister cells survive matrix disruption. Front. Microbiol. 10:2603. doi: 10.3389/fmicb.2019.02603
Sousa, A., and Pereira, M. (2014). Pseudomonas aeruginosa diversification during infection development in cystic fibrosis lungs—a review. Pathogens 3, 680–703. doi: 10.3390/pathogens3030680
Stewart, P. S., and Bjarnsholt, T. (2020). Risk factors for chronic biofilm-related infection associated with implanted medical devices. Clin. Microbiol. Infect. 26, 1034–1038. doi: 10.1016/j.cmi.2020.02.027
Stewart, P. S., and Franklin, M. J. (2008). Physiological heterogeneity in biofilms. Nat. Rev. Microbiol. 6, 199–210. doi: 10.1038/nrmicro1838
Viducic, D., Ono, T., Murakami, K., Susilowati, H., Kayama, S., Hirota, K., et al. (2006). Functional analysis of spoT, relA and dksA genes on quinolone tolerance in Pseudomonas aeruginosa under nongrowing condition. Microbiol. Immunol. 50, 349–357.
Wang, L., Di Luca, M., Tkhilaishvili, T., Trampuz, A., and Gonzalez Moreno, M. (2019). Synergistic activity of fosfomycin, ciprofloxacin, and gentamicin against Escherichia coli and Pseudomonas aeruginosa biofilms. Front. Microbiol. 10:2522. doi: 10.3389/fmicb.2019.02522
Werner, E., Roe, F., Bugnicourt, A., Franklin, M. J., Heydorn, A., Molin, S., et al. (2004). Stratified growth in Pseudomonas aeruginosa biofilms. Appl. Environ. Microbiol. 70, 6188–6196. doi: 10.1128/AEM.70.10.6188-6196.2004
Williamson, K. S., Richards, L. A., Perez-Osorio, A. C., Pitts, B., McInnerney, K., Stewart, P. S., et al. (2012). Heterogeneity in Pseudomonas aeruginosa biofilms includes expression of ribosome hibernation factors in the antibiotic-tolerant subpopulation and hypoxia-induced stress response in the metabolically active population. J. Bacteriol. 194, 2062–2073. doi: 10.1128/JB.00022-12
Wiuff, C., Zappala, R. M., Regoes, R. R., Garner, K. N., Baquero, F., and Levin, B. R. (2005). Phenotypic tolerance: antibiotic enrichment of noninherited resistance in bacterial populations. Antimicrob. Agents Chemother. 49, 1483–1494. doi: 10.1128/AAC.49.4.1483-1494.2005
Yu, Q., Griffin, E. F., Moreau-Marquis, S., Schwartzman, J. D., Stanton, B. A., and O’Toole, G. A. (2012). In vitro evaluation of tobramycin and aztreonam versus Pseudomonas aeruginosa biofilms on cystic fibrosis-derived human airway epithelial cells. J. Antimicrob. Chemother. 67, 2673–2681. doi: 10.1093/jac/dks296
Keywords: biofilm, Pseudomonas aeruginosa, antibiotic treatment, tolerance, persister cells, adaptation
Citation: Soares A, Alexandre K and Etienne M (2020) Tolerance and Persistence of Pseudomonas aeruginosa in Biofilms Exposed to Antibiotics: Molecular Mechanisms, Antibiotic Strategies and Therapeutic Perspectives. Front. Microbiol. 11:2057. doi: 10.3389/fmicb.2020.02057
Received: 16 June 2020; Accepted: 05 August 2020;
Published: 27 August 2020.
Edited by:
Maria Tomas, A Coruña University Hospital Complex (CHUAC), SpainReviewed by:
David Lebeaux, Assistance Publique Hopitaux De Paris, FranceRosalia Cavaliere, University of Technology Sydney, Australia
Copyright © 2020 Soares, Alexandre and Etienne. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Anaïs Soares, anais.so@free.fr; Manuel Etienne, manuel.etienne@chu-rouen.fr