 Charles Pin-Kuang Lai1,2 Xandra Owen Breakefield1,2*
Charles Pin-Kuang Lai1,2 Xandra Owen Breakefield1,2*- 1 Department of Neurology, Neuroscience Center, Massachusetts General Hospital and Program in Neuroscience, Harvard Medical School, Boston, MA, USA
- 2 Department of Radiology, Center for Molecular Imaging Research, Massachusetts General Hospital, Boston, MA, USA
Extracellular membrane vesicles (EMVs) are nanometer sized vesicles, including exosomes and microvesicles capable of transferring DNAs, mRNAs, microRNAs, non-coding RNAs, proteins, and lipids among cells without direct cell-to-cell contact, thereby representing a novel form of intercellular communication. Many cells in the nervous system have been shown to release EMVs, implicating their active roles in development, function, and pathologies of this system. While substantial progress has been made in understanding the biogenesis, biophysical properties, and involvement of EMVs in diseases, relatively less information is known about their biological function in the normal nervous system. In addition, since EMVs are endogenous vehicles with low immunogenicity, they have also been actively investigated for the delivery of therapeutic genes/molecules in treatment of cancer and neurological diseases. The present review summarizes current knowledge about EMV functions in the nervous system under both physiological and pathological conditions, as well as emerging EMV-based therapies that could be applied to the nervous system in the foreseeable future.
Introduction
Ligand-receptor interaction and direct cell–cell contacts via specialized physical conduits, such as gap junctions and membrane nanotubes, have long been considered as the predominant means of intercellular communication (Davis and Sowinski, 2008; Goodenough and Paul, 2009). Yet, a novel method of cell-to-cell communication has recently emerged from groundbreaking discoveries in the past few years on nucleic acid content of extracellular membrane vesicles (EMVs). EMVs have been demonstrated to facilitate horizontal transfer of mRNAs, microRNAs (miRNAs), and proteins between cells without direct cell-to-cell contact (Bergsmedh et al., 2001; Ratajczak et al., 2006a; Valadi et al., 2007; Al-Nedawi et al., 2008; Skog et al., 2008; Balaj et al., 2011; Ramachandran and Palanisamy, 2011; Turchinovich et al., 2011; Chen et al., 2012). There are several EMV categories known to-date, which are included under the general terms exosomes, microvesicles (MVs), and apoptotic blebs (ABs).
Exosomes are the smallest EMVs (40–100 nm in diameter), and homogenous in shape (cup-shaped after fixation under electron microscopy with a buoyant density of 1.13–1.19 g/cm3 (Théry et al., 2001; Hristov et al., 2004). Unlike other types of EMVs that are directly shed/released from the plasma membrane, exosomes are formed by a series of processes beginning with inward invagination of clathrin-coated microdomains on the plasma membrane (Denzer et al., 2000). Once these vacuoles have entered the cell, the Endosomal Sorting Complex Required for Transport (ESCRT) facilitates the development of the invaginated vacuoles carrying ubiquitinated cargos into early endosomes. This is followed by a secondary invagination of vesicles (termed intraluminal vesicles, ILVs), into the endosomes where they accumulate with subsequent maturation of the complex into large multivesicular bodies (MVBs; Denzer et al., 2000). At this stage, MVBs may be trafficked to lysosomes for degradation (“degradative MVBs”) or, instead, fuse with the plasma membrane (“exocytic MVBs”) for the release of ILVs into the extracellular space, where upon they are referred to as exosomes (Mathivanan et al., 2010). A study on oligodendrocytes suggested that ILV release is ESCRT-independent and relies on the distribution of sphingolipid ceramide in MVBs, which directs the extracellular release of ILVs as exosomes (Trajkovic et al., 2008). Additional investigations are needed to determine if distinct MVB or ILV populations destined for degradation or exocytic release are present, as well as whether a common exosomal trafficking mechanism exists in all cell types (Mathivanan et al., 2010). Understanding the biogenesis and trafficking of exosomes will provide insight into how cells employ these extracellular organelles for intercellular communication. In some studies, release of exosomes appears to depend of Rab27 (Ostrowski et al., 2010) and Rab 35 (Hsu et al., 2010), and can be blocked with an inhibitor of neutral sphingomyelinase (Trajkovic et al., 2008). In addition, elevated [Ca2+]i, following Ca2+ and ionophore A23187 treatment was found to induce exosome and microvesicle release from erythrocytes (Allan et al., 1980; Salzer et al., 2002), further supporting a role of EMVs in response to different stimuli.
Microvesicles (MVs) are irregularly shaped, larger EMVs with a 100–1,000 nm diameter (Pilzer et al., 2005; Cocucci et al., 2009). A defined buoyant density of MVs has not yet been determined, but it may overlap that of exosomes (Théry et al., 2009; van Dommelen et al., 2011). In contrast to the endocytotic origin of exosomes, release of MVs results from outward budding at the plasma membrane followed by fission of their connecting membrane stalks (Kobayashi et al., 1984; Dolo et al., 2000; Cocucci et al., 2007; Piccin et al., 2007). While MV biogenesis remains to be defined, microdomains on the plasma membrane containing a high cholesterol level and signaling complexes, or lipid rafts, have been suggested to selectively sequester lipids for MV generation (Del Conde et al., 2005). Work by Gould and collaborators indicates that MV release may be triggered by oligomerizing proteins on the cell surface and may share mechanistic elements with release of enveloped viruses (Gould et al., 2003; Shen et al., 2011). MV production is observed in a variety of cells in a resting state, but can be significantly elevated under various stimulations, including increased [Ca2+]i, cellular stress (e.g., DNA damage), decreased cholesterol levels, cytokine exposure, and anticancer drug treatment (Salzer et al., 2002; Shedden et al., 2003; Yu et al., 2006; Llorente et al., 2007; Lehmann et al., 2008; Bianco et al., 2009). Notably, even larger EMVs (1–5 μm in diameter) are released from tumor cells, especially in response to EGF stimulation (Di Vizio et al., 2009). Altogether, these findings suggest an active physiological role of MVs under different cellular conditions.
Apoptotic blebs are 50–4,000 nm in diameter with a buoyant density of 1.16–1.28 g/cm3 (Hristov et al., 2004; Simak and Gelderman, 2006). Similar to MVs, ABs are also irregularly shaped, making them difficult to discern from one another based on their morphology. ABs, as its name suggests, are released from condensed and fragmented apoptotic cells during late stages of cell death (Henson et al., 2001; Hristov et al., 2004). ABs retain DNA fragments from the deceased cells, and can be taken up by neighboring cells for horizontal gene transfer as a form of intercellular communication (Bergsmedh et al., 2001; Holmgren, 2010).
EMV Terminology Vis-à-Vis Cell of Origin
Most cells throughout the body, including those in the nervous system are believed to release EMVs. Early pioneering studies named EMVs based on their cellular origins, and to some extent their biogenesis, such as: archaeosomes, argosomes, dexosomes, epididymosomes, prostasomes, and oncosomes (Brody et al., 1983; Quaite-Randall et al., 1995; Zitvogel et al., 1998; Greco et al., 2001; Simpson et al., 2008; Al-Nedawi et al., 2009; Krishnan and Sprott, 2008; Di Vizio et al., 2009). These EMVs include ones isolated from a variety of cells/tissues in human body, i.e., dendritic cells (DCs), prostate gland, and cancer cells, as well as other species such as Drosophila (Simpson et al., 2008). As in many rapidly expanding fields in research, standardization of nomenclature for the different categories of EMVs remains to be resolved by an official organization, and is being undertaken by the International Society for Extracellular Vesicles. As it stands now, MVs/ectosomes/microparticles generally refer to extracellular vesicles released via a direct budding/shedding from the cellular plasma membrane, whereas exosomes are those released from MVBs following their fusion with the plasma membrane. Categorizing EMVs has been intrinsically challenging due to the multiple variable characteristics of EMVs, including: (1) cellular origin and physiological state of cell; (2) biophysical properties and lipid composition; (3) nucleic acids and protein content; and (4) size. For the purpose of this review, EMVs will be used to encompass these extracellular vesicle types.
Biophysical Properties and Lipid Composition
Aside from the different biophysical properties (i.e., size, shape, buoyant density) mentioned above for exosomes, MVs, and ABs (Table 1), different types of EMVs also have different lipid compositions. By using liquid chromatography and mass spectrometry, a variety of lipid components constituting EMVs isolated from different cells have been identified, including phosphatidylcholine, phosphatidylethanolamine, phosphatidylserine (PS), lyso-bisphosphatidic acid, ceramide, cholesterol, and spingomyelin (Chu et al., 2005; Subra et al., 2007). The particular lipid composition of each EMV type likely contributes to its biophysical properties. Indeed, Parolini et al. (2009) recently reported that different lipid compositions, namely those containing sphingomyelin and N-acetylneuraminyl-galactosylglucosylceramide (GM3), are directly related to rigidity and delivery efficiency of exosomes to other cells. In addition, the level of PS exposed on the outer leaflet of exosomes appears to be lower than that of MVs and ABs (Mathivanan et al., 2011). This observation may correlate with the different biogenesis of EMV populations wherein exosomes are of endocytic origin, and MVs and ABs are derived from outward budding from the plasma membrane. PS is displayed on the outer exosome leaflet through floppase, flippase, and scramblase activities, and appears to mediate docking of proteins involved in signaling and fusion to the plasma membrane (Piccin et al., 2007). Therefore, the varying level of PS may affect communication functions of EMVs. Furthermore, ongoing studies indicate that EMVs may have a conserved glycan signature as compared to the cells from which they derive, suggesting a role of glycosylation in EMV protein sorting (Batista et al., 2011).
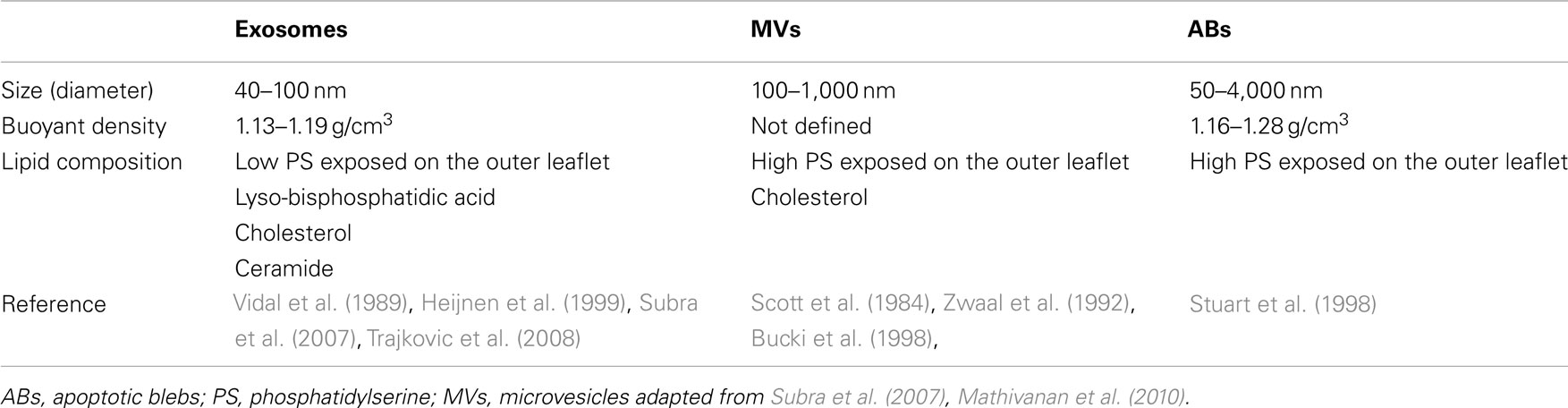
Table 1. Biophysical properties of extracellular vesicles.
Contents of EMVs
Extracellular membrane vesicles are known to contain a broad spectrum of proteins, including transcriptional factors, surface receptors, and “marker” proteins, including tetraspanins (CD63, CD81), integrins, Tsg101, Alix, heat shock protein (HSP70, 90), and flotillin-1 (Lee et al., 2011). In addition, they contain a range of nucleic acids including mRNAs, which can be translated in recipient cells, microRNA (miRNA), and non-coding RNAs (ncRNA) which can regulate gene/mRNA expression, as well as DNA of as yet unvalidated function (Valadi et al., 2007; Skog et al., 2008; Guescini et al., 2010; Balaj et al., 2011; Waldenström et al., 2012). An interactive database will be needed to accommodate these increasing findings in the field of EMVs. Efforts are indeed underway, and a manually curated, web-based community database, ExoCarta, has been introduced to record exosomal proteins, RNAs, and lipids found in different EMV studies (Mathivanan et al., 2012). In addition, an Urinary Exosome Protein Database was created, as its name suggests, to provide a database of identified proteins from urinary exosomes via protein mass spectrometry (http://dir.nhlbi.nih.gov/papers/lkem/exosome/.)
Functions of EMVs in the Nervous System
Most cells in the nervous system are believed to release EMVs. Given that EMVs are capable of transferring genetic information, proteins, and lipids without direct cell-to-cell contact, researchers are focusing on the potential role of EMVs during development of the nervous system and as part of neural functions, as well as in disease. In this review, we will cover some of the recent findings on both the physiologic and pathophysiologic roles of EMVs in the nervous system, as well as the emergence of EMV-mediated therapies which may be applicable for treatment of neurological diseases in the foreseeable future. Recent reviews by Frühbeis et al. and Bellingham also provide extended updates into EMVs’ participation in neuron-glia communication and neurodegenerative diseases, respectively (Bellingham et al., 2012; Frühbeis et al., 2012).
Normal Functions
A number of studies have implicated EMVs in neuronal development, synaptic activity, protective mechanisms, and nerve regeneration with various reports indicating EMV release by neural stem/progenitor cells (Marzesco et al., 2005), neurons (Fauré et al., 2006), astrocytes (Taylor et al., 2007), microglia (Potolicchio et al., 2005), and oligodendrocytes (Krämer-Albers et al., 2007) in the brain and Schwann cells in the peripheral nervous system (Court et al., 2008).
Development
In the developing mouse brain during early neurogenesis there is a peak (E10.5–E13.5) of EMV release into the ventricular fluid in the neural tube of small (50–80 nm) and large (600 nm) vesicles which are positive for the stem cell marker, prominin-1 (CD133), although their function is not known (Marzesco et al., 2005). Other studies suggest that these EMVs may be involved in transfer of mRNAs encoding pluripotent transcription factors which can reprogram phenotypes of other cells (Ratajczak et al., 2006a). EMVs have the capacity to participate in the spatial and temporal gradients critical in development. Consistent with this role, non-neuronal floor plate cells in the ventral midline of mouse embryos are able to transfer β-galactosidase to neighboring axons, suggesting EMV transfer as an aspect of axonal path finding (Campbell and Peterson, 1993; Figure 1A). In a temporal patterning motif, oligodendrocytes appear to release EMVs as a means of auto-inhibiting myelination until appropriate signals are released from neurons during development, indicating that neuronal maturation is complete (Bakhti et al., 2011). Then as myelination commences EMVs are released from oligodendrocytes in a ceramide-triggered cascade that may be critical for transfer of the major myelin protein, proteolipoprotein (Trajkovic et al., 2008). EMVs may also participate in the genetic changes in the genome (genomic plasticity) of embryonic cells by supporting novel retrotransposon integrations. Both neural progenitor cells in normal embryos (Coufal et al., 2009) and brain tumor cells (Balaj et al., 2011) have high levels of retrotransposon expression, indicating a broadly transcriptionally active genome with the potential for retrotransposon integration events creating novel genotype/phenotypes. At least in the case of tumors, these EMVs have high levels of retrotransposon sequences and reverse transcriptase which may allow cell-to-cell transfer of this genomic plasticity leading to changes in gene expression, with pro-proliferative events being selected for.
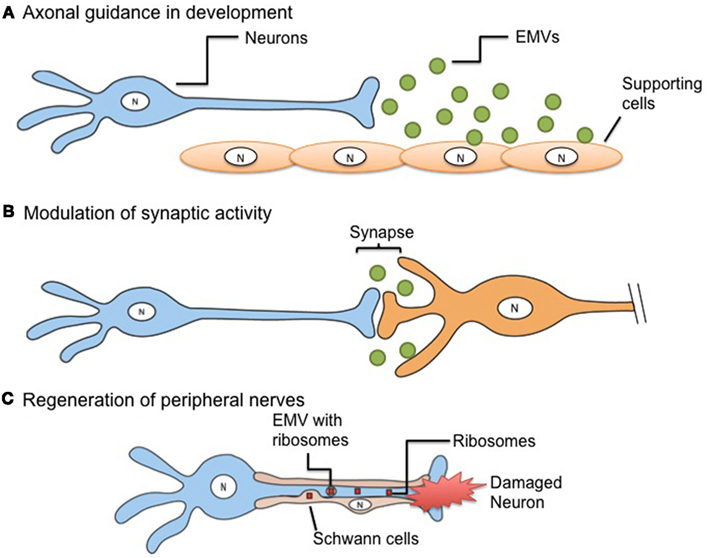
Figure 1. Extracellular membrane vesicles-mediated mechanisms in neurons. (A) A gradient of EMVs in the developing nervous system can serve as a directional guide to axonal growth. (B) EMVs released from presynaptic nerve terminals and taken up by their postsynaptic partners can carry informational content which can modulate the strength of synaptic activity. (C) Regeneration of peripheral nerves in enhanced by the EMV transfer of ribosomes and mRNA directly from surrounding Schwann cells into the injured nerve to promote protein synthesis.
Several studies in Drosophila have begun to tease out the role of EMVs in different developmental processes. EMV-like vesicles, termed argosomes (Greco et al., 2001) are used to transport a morphogenic Wnt signaling protein along spatial and temporal gradients in wing development, and may also carry Hedgehog, Notch, decapentaplegic (dpp), and Wingless (Wg) signaling proteins involved in setting up developmental gradients in other tissues (for review see Cadigan, 2002; Lakkaraju and Rodriguez-Boulan, 2008). Recently, EMVs were found to be involved in transfer of Wnt-1/Wg at the neuromuscular junction in Drosophila both during development and in mature neurons (Korkut et al., 2009). In this case, a multipass transmembrane protein, Evi, assists in presynaptic trafficking of Wnt-1/Wg into vesicles within the synaptic cleft, as well as in subsequent interactions of this signaling ligand with its receptor in postsynaptic cells. When Evi was rendered non-functioning in evi mutant Drosophila model, Wnt signaling across synapses is disrupted (Korkut et al., 2009).
Synaptic activity
In pioneering studies, Fauré et al. (2006) showed that undifferentiated cortical neurons in culture released EMVs containing L1, a neuronal cell adhesion protein, and GluR2/3 subunits of glutamate AMPA receptors, with release of EMVs being stimulated by depolarization, thus suggesting a role in synaptic function. Subsequent studies confirmed this phenomenon in fully differentiated cortical neurons in culture, and in addition showed that release was stimulated by a calcium ionophore, as well as by an antagonist of GABAA receptors, both of which result in increased spontaneous neuronal activity (Lachenal et al., 2011). Further, EMVs were found to incorporate the neuronal specific heavy chain of tetanus toxin and reasoned that the presence of GluR2 subunits in EMVs and increased release associated with depolarization could modulate synaptic activity (Smalheiser, 2007; Figure 1B).
Injury and regeneration
In general, studies to-date indicate that EMVs primarily serve a protective role in the nervous system. For example, oligodendrocytes release exosomes containing myelin and stress-protective proteins, which serve in the trophic support of neurons (Krämer-Albers et al., 2007). Although synapsin I is usually thought of as a neuronal specific protein associated with synaptic vesicles, it is also produced in lower amounts by astrocytes and released from them within EMVs in response to stress conditions (Wang et al., 2011). These glia-derived EMVs promote neurite outgrowth and increase neuron survival through association between synapsin I and oligomannosidic glycans in response to depolarization and toxic insults to neurons. Brain injury leads to accumulation of toxic proteins in neurons with survival promoted by expression of Ndfip1, an interacting protein with Nedd4 ubiquitin ligases which mediate protein degradation (Sang et al., 2006). Both Ndfip1 and Nedd4 proteins are found in EMVs released by neurons and are hypothesized to serve as a means of rapid removal of toxic proteins after injury (Putz et al., 2008). In a case where EMVs can cause damage to nervous tissue, during brain injury increase in extracellular ATP leads to release of EMVs from microglia and astrocytes through a sphingomyelinase-dependent process (Bianco et al., 2009). These EMVs contain and release IL-1β, a cytokine which inducing inflammatory responses that are damaging to brain tissue.
In a variation on the EMV release mechanism, Schwann cells surrounding a degenerating or damaged peripheral nerve, translocate vesicles surrounded by two membranes containing polyribosomes into the axon where the contents are released (Court et al., 2008; Twiss and Fainzilber, 2009; Figure 1C). This serves as a means of delivering mRNA and ribosomes to injured neurons to promote local protein synthesis needed for regeneration, with recent studies in vivo showing that labeled ribosomes in the nerve are derived from Schwann cells (Court et al., 2011).
Pathology
Neurodegeneration
Extracellular membrane vesicles have been implicated in the spread of toxic proteins within the nervous system in a number of neurodegenerative diseases, including transmissible spongiform encephalopathies, Alzheimer’s disease (AD), Parkinson’s disease (PD), tauopathies, and amyotrophic lateral sclerosis (ALS; for review see Vingtdeux et al., 2007; Vella et al., 2008; Guest et al., 2011; Frühbeis et al., 2012). In all these diseases mutated or “misfolded” proteins serve as templates for formation of protein oligomers that accumulate and interfere with neuronal function, eventually leading to cell death. Possibly in an attempt to rid themselves of these proteins, neurons process them through the endosomal pathway leading either to degradation in lysosomes or to incorporation into MVBs, with the latter serving as a release hatch into the extracellular space within EMVs. In early studies of the role of EMVs in this process, two groups described the incorporation of both the normal prion protein (PrP) and the misfolded pathogenic prion protein (PrPsc) into EMVs (Ecroyd et al., 2004; Fevrier et al., 2004; Février et al., 2005). Further studies showed that EMV-associated PrPsc was transmitted to normal cells with initiation of prion propagation involving PrP in those cells (Vella et al., 2007). This concept of the tendency of some proteins to seed their own aggregation with “infectious” delivery via EMVs has been implicated in a number of neurodegenerative diseases. Interestingly, in many of these diseases there is a spatiotemporal propagation of the pathology suggesting cell-to-cell spread (Guest et al., 2011), which for non-secreted proteins could be mediated by EMV transfer or nanotubes (Rustom et al., 2004; Gousset and Zurzolo, 2009).
Alzheimer’s disease
The concept of EMV-mediated transfer of aggregation-prone proteins has been the most studied for the amyloid-β (Aβ) peptide associated with AD. Although the neuropathologic plaques characteristic of the disease are extracellular, it is currently believed that the plaques may represent a “disposal dump” and it is really the soluble oligomeric fibrils of the Aβ peptide which are neurotoxic (Lublin and Gandy, 2010), as they can serve as a “seeding” center for AD pathology in naive mice (Eisele et al., 2010). These peptides are generated when the amyloid precursor protein is proteolytically processed at the plasma membrane with peptides being re-taken up into endosomes where they can enter MVBs and be released from the cell in EMVs (Rajendran et al., 2006). The possibility that Aβ EMVs can serve as infectious agents is supported by orally transmitted amyloid A1 (AA) amyloidosis among cheetahs (Zhang et al., 2008) with EMVs present in saliva and fecal matter – although, to-date, the role of EMVs in either disease process has not been confirmed. EMVs can also serve as a means of degradation of toxic Aβ when taken up by microglia, but when that clearance pathway is overwhelmed pathologic accumulation of Aβ neurons commences (Yuyama et al., 2012).
Other neurodegenerative conditions
In other neurodegenerative diseases, proteins capable of seeding pathology have also been found in EMVs and shown to have spatiotemporal spread within the nervous system. These include the microtubule-associated tau protein which aggregates in a number of dementia states (Saman et al., 2012), the mutant SOD1 protein underlying some familial forms of ALS (Gomes et al., 2007) and α-synuclein which plays a central role in PD pathogenesis (Emmanouilidou et al., 2010). Studies in PD mouse models show that grafted cells containing aggregated alpha α-synuclein can transfer this aggregate to host cells in the brain (Hansen et al., 2011). These aggregates appear to be passed between cells through the extracellular space as independent entities or via EMVs or nanotubes (Hansen and Li, 2012) with the relative contribution of each of these pathways in the brain still undetermined. It remains an intriguing possibility that EMVs can act as “infectious” agents to spread toxic oligomerizing proteins not only within an individual, but between individuals through fluid contacts, as in amyloidosis in cheetahs (Zhang et al., 2008).
Other disease states
Roles are emerging for EMVs in a number of neurologic disease states. They are capable of transmitting death signals, for example, incorporation of caspase-1 into EMVs produced by monocytes and with transfer to surrounding cells can be the “kiss of death” (Sakar et al., 2009). Apoptotic bodies which form during cell death are also caspase-containing EMVs that can deliver contents to other cells (Simpson et al., 2008). EMVs have also been implicated in autoimmune diseases, such as multiple sclerosis, with those derived from DCs leading to activation of inflammatory NF-κB in microglia and recruitment of major histocompatibility complex (MHC) class II for presentation of self-antigens (Teo and Wong, 2010). In contrast, EMVs can also be protective, with those shed from endothelial cells and astrocytes containing nucleoside triphosphate diphosphohydrolases which can degrade toxic levels of ATP released during breach of the blood brain barrier (BBB) in ischemia (Ceruti et al., 2011). As an apparently common form of intercellular communication, EMVs are undoubtedly a critical player in many different events in the nervous system – providing protection from neurodegeneration, as well as propagation of toxic influences.
Tumors
Early on investigators noticed that glioblastoma (GBM) cells were covered with “microparticles” (EMVs; Tani et al., 1978). During transformation and progression to malignancy brain tumor cells appear to increase the number and types of EMVs released, as for example when EMV content is compared in conditioned media from GBM cells with normal cells in culture (Balaj et al., 2011). EMVs are believed to be used by tumor cells to modify normal cells in their vicinity so as to promote tumor growth, with most studies carried out in non-nervous system tumors. Pro-active mechanisms include suppression of immune responses to the tumor, opening up of the extracellular space to facilitate tumor cell invasion, stimulation of angiogenesis and modulation of cellular phenotypes (for review see Ichim et al., 2008; Al-Nedawi et al., 2009; Muralidharan-Chari et al., 2010). In addition to proteins within EMVs which modulate these responses, EMVs from GBM cells (as well as other tumor cell types) are also enriched in RNAs associated with proliferation, invasion and immune repression (Skog et al., 2008). Other tumor-related aspects of EMVs include their ability to expel chemotherapeutic drugs from tumor cells (Shedden et al., 2003) and to carry tissue factor leading to hypercoagulation in cancer patients (Zwicker et al., 2009). It seems likely that most of these tumor-enhancing functions of EMVs are associated with tumors of the nervous system as in tumors in other tissues.
Immune responses
There is an extensive literature on the role of EMVs both in suppression and enhancement of immune responses in cancer, most of which have been studied in peripheral tumors. In general, EMVs released by tumor cells serve to suppress the immune response to tumor antigens, including acting as decoys in the tumor environs, switching off T cell responses, eliminating antitumor effector cells, and preventing differentiation of immature DCs into antigen-presenting cells (for review see Taylor and Gerçel-Taylor, 2005; Iero et al., 2008; Théry et al., 2009).
Angiogenesis and invasion
Extracellular membrane vesicles from GBM cells in culture promote angiogenesis of human brain microvascular endothelial cells, which is mediated in part by their relatively high concentrations of angiogenic factors – VEGF, IL-8, TIMP-1, IL-6, and angiogenin as compared to the tumor cells of origin (Skog et al., 2008; Graner et al., 2009). Another EMV protein in this angiogenic cascade includes delta-like 4 Zigand (D114), a recently identified partner for the Notch receptor, that inhibits Notch signaling thereby increasing vessel density and branching in glioma xenograft tumors in vivo (Sheldon et al., 2010). Other components of EMVs which promote angiogenesis include sphingomyelin (Kim et al., 2002) and CD147/extracellular matrix metalloprotease inducer (EMMPRIN; Millimaggi et al., 2007). In addition, the acidic environment of the tumor can cause lysis of EMVs and release of vesicular proteins, such as VEGF (Taraboletti et al., 2006). Several studies using other tumor cell types have also indicated possible transfer of functional miRNAs via EMVs (e.g., Collino et al., 2010; Kosaka et al., 2010; Zhang et al., 2010; Mittelbrunn et al., 2011; Yang et al., 2011). miRNA-296 is known to be elevated in brain microvascular endothelial cells as part of an angiogenic response to the presence of glioma cells (Würdinger et al., 2008) and this miRNA is contained in GBM EMVs (J. Skog, unpublished data), so it seems likely that glioma EMVs may contribute to elevated miRNA-296 in endothelial cells. Tumor-derived EMVs also express matrix metalloproteinases (MMPs) and an extracellular MMP inducer on their surface to degrade the extracellular matrix and thereby facilitate invasion of tumor cells into surrounding normal brain tissue (e.g., Castellana et al., 2009).
Phenotypic modification
Extracellular membrane vesicles released by brain tumors alter the phenotype of surrounding cells, presumably through a complex of factors including transcriptional regulators (both proteins and ncRNA), miRNAs, mRNAs, and surface receptors (for review see van der Vos et al., 2011). An early example of this was the demonstration that glioma cells expressing the mutant epidermal growth factor receptor variant III (EGFRvIII) on their plasma membrane pass this onto the membrane of EMVs from where is incorporated into recipient cell membranes, leading to an increased transformative phenotype of the recipient cells (Al-Nedawi et al., 2008). Parallel studies showed that the mRNA for EGFRvIII is also present in EMVs from mutant-positive tumor cells and can be detected in serum EMVs from patients who harbor the corresponding mutation in their tumors, thereby providing a biomarker for genetic status of the tumors (Skog et al., 2008). Others have shown that oligodendroglioma cells send out EMVs containing the apoptotic protein, TRAIL, which leads to death of normal astrocytes and neurons (Lo Cicero et al., 2011).
Genotypic modification
Interestingly, a number of tumors release retroviral-like particles contained within the EMV pool that contain the RNA from human endogenous retroviral (HERV) sequences and reverse transcriptase (Lavie et al., 2005; Contreras-Galindo et al., 2008; Balaj et al., 2011), and may thus mediate abortive infections which disrupt the recipient cell genome. GBM and medulloblastoma brain tumor cells have been found to harbor high levels of HERV RNAs, as well as reverse transcriptase activity (Balaj et al., 2011). It seems likely that a component of increased EMV production by these tumor cells comprises retroviral-like particles, which though non-replicative may still have the capacity to “infect” other cells and integrate into the recipient cell genome, thereby causing potential mutagenesis and oncogene activation. Further, HERVs encode fusogenic proteins which may increase the ability of tumor-derived EMVs to fuse with and enter recipient cells (Duelli and Lazebnik, 2007).
Other studies have shown that during tumor cell death, ABs which fractionate with EMVs, contain oncogene DNA for H-ras and c-Myc which are taken up by other cells (Holmgren et al., 1999; Bergsmedh et al., 2001). Even non-dying tumors cells with amplified c-Myc release this DNA into EMVs (Balaj et al., 2011). It remains to be determined if these EMV-transferred oncogenes can be integrated into the genome of recipient cells.
Viruses
Three viruses associated with disruption of brain functions have been found to use EMVs to promote infection and avoid immune rejection by the host, as well as in some cases to confer resistance to infection (for review see Meckes and Raab-Traub, 2011; Wurdinger et al., 2012). These three viruses are – herpes simplex virus type 1 (HSV-1), which in immune compromised patients can cause viral encephalitis (Steiner, 2011); the tumorigenic herpes virus, Epstein–Barr virus (EBV), which can cause central nervous system (CNS) lymphomas (Gerstner and Batchelor, 2010); and human immunodeficiency virus (HIV), which can lead to neurocognitive deficits, dementia, and premature brain aging (Gannon et al., 2011).
Herpes simplex virus type 1
Early during HSV-1 replicative infection/activation, prior to production of infectious virions, EMVs are released from infected cells and serve to prime surrounding cells for productive infection and to reduce immune rejection of the virus (McLauchlan et al., 1992). These EMVs contain viral tegument proteins, some of which serve as immediate early transcription factors to “jump start” secondary infection (Dargan and Subak-Sharpe, 1997). A viral glycoprotein contained in the EMVs, glycoprotein B (gB) also acts on MHCII molecules to prevent presentation of viral peptide antigens to the immune system (Temme et al., 2010).
Epstein–Barr virus-infected cells also use EMVs to reduce the immune response through incorporation of immune suppressive proteins, LMP1 (Flanagan et al., 2003), and galectin-9 (Klibi et al., 2009). In addition, EBV transfers viral miRNAs via EMVs to repress translation of cell proteins which promote resistance to infection (Pegtel et al., 2010; Meckes et al., 2010). EMVs produced by different cell types may have different effects, for example those released by B cells containing glycoprotein 350 can block EBV infection of other cells (Vallhov et al., 2011).
Human immunodeficiency virus retroviral particles bud from the plasma membrane in a similar manner to MVs (Gould et al., 2003; Jouvenet et al., 2011). EMVs released from cells harboring HIV can confer increased infectivity to other cells through transfer of CCR5 co-receptors (Mack et al., 2000) and CXCR4, a chemokine receptor that interacts with CD4 on the cell surface to facilitate HIV entry (Rozmyslowicz et al., 2003). EMVs derived from infected cells also contain the HIV protein, Nef, which can induce apoptosis of CD4+ cells (Lenassi et al., 2010), thus suppressing the immune response. In a bit of a turnaround these EMVs from HIV infected cells also contain an anti-viral cytidine deaminase which can inhibit viral replication (Khatua et al., 2009).
EMVs as Therapeutic Delivery Vehicles
Before the recent landmark discoveries of EMVs as a new conduit for cell-to-cell genetic communication (Ratajczak et al., 2006b; Valadi et al., 2007; Ramachandran and Palanisamy, 2011; Turchinovich et al., 2011; Chen et al., 2012), the artificial counterpart of EMVs, liposomes, had been well studied as a nanodelivery system over past decades. Liposomes are spherical vesicles composed of one or multiple natural and/or synthetic lipid bilayers with an aqueous core and a diameter ranging from 50 nm to 5 μm. By taking advantage of these structural properties, liposomes have been investigated as a means to “load and deliver” pharmaceutical drugs and peptides (reviewed in Malam et al., 2009). However, a major conundrum in liposome-mediated delivery lies in its biocompatibility and biodegradability properties, such that ideal liposomes should evade detection by the immune system and have a longer half-life in the circulation for therapeutic cargo delivery (reviewed in Immordino et al., 2006). While remarkable advances have been made in reducing immunogenicity of liposomes, described as “stealth liposomes” and in increasing their half-life in the circulation by coating them with poly-(ethylene glycol, PEG; Allen et al., 1989, 1991), researchers continue to seek endogenous nanodelivery systems to overcome the obstacles faced by artificial vesicles. With emerging understanding of their biological functions, EMVs have been suggested as an ideal candidate to fulfill this role as “physiologic liposomes.”
Investigators have made significant progress in the use of EMVs for therapy by taking advantage of their low immunogenicity and unique delivering capability (Zhang et al., 2007; Sun et al., 2009; Alvarez-Erviti et al., 2011; Zhuang et al., 2011; Bolukbasi et al., 2012). By genetic engineering of EMV producer cells or direct modification of EMVs, they can be used to transport therapeutic molecules and agents via insertion into the lipid bilayer and/or loading into their aqueous core (Figure 2). Analogous to liposomes, EMVs also serve as an excellent means of protection of “therapeutic cargoes” wherein packaged mRNA, small interfering RNAs (siRNA), proteins, and drugs are better preserved from degradation when compared to their unshielded counterparts. Using these advantages, EMV-mediated therapy is being actively studied in three main fields: immunotherapy, RNA-interference (RNAi) and drug delivery.
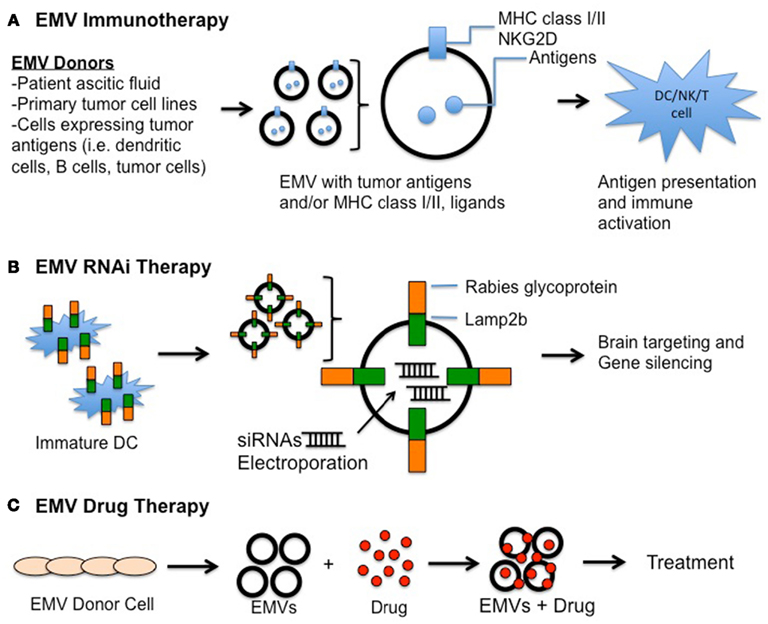
Figure 2. Extracellular membrane vesicles-based therapies. (A) EMV immunotherapy. EMVs containing tumor-antigen within and/or on the membrane surface are isolated from different sources and introduced in vivo to elicit targeted immune responses. (B) EMV RNAi therapy. EMVs derived from immature dendritic cells (DCs) expressing Rabies glycoprotein-Lamp2b fusion protein were electroporated with siRNAs for targeting against neurons, microglia, and oligodendrocytes for subsequent gene silencing (Alvarez-Erviti et al., 2011). (C) EMV drug therapy. Therapeutic compounds can be packaged into/onto EMVs isolated from donor cells to minimize degradation and increase delivery to intended sites.
EMVs in cancer immunotherapy
Immunotherapy represents one of the most investigated aspects in EMV-mediated therapy. In immunotherapy, an ideal cancer vaccine serves as an antigen-presenting medium to prime the immune system to recognize tumor-specific antigens, thereby eliciting immune responses against the tumor cells while leaving normal cells unharmed (Trumpfheller et al., 2012). A study by Raposo et al. (1996) first showed B lymphocyte-secreted EMVs contained MHC class II by immunoelectron microscopy and could induce MHC class II-dependent CD4+ T cell responses in vitro. Since DCs are the most potent antigen-presenting cells in the adaptive immune system, numerous studies ensued focusing primarily on the immuno-modulating effect of EMVs on DCs in search of more effective cancer vaccines (see review in Tan et al., 2010). Briefly, researchers have used EMVs isolated from various sources to pulse DCs with antigens (André et al., 2004; Chaput et al., 2004; Cho et al., 2005; Hao et al., 2007; Taïeb et al., 2006; Bai et al., 2007; Beauvillain et al., 2007; Guo et al., 2008; Temchura et al., 2008; Viaud et al., 2009; Bu et al., 2011) in order to activate immune cells (i.e., T lymphocytes and natural killer cells) against tumor cells, including those in the ascitic fluid of cancer patients (André et al., 2002), and wild-type or engineered cancer cells in vitro (Wolfers et al., 2001; Hegmans et al., 2005; Chen et al., 2006; Yang et al., 2007; Xiu et al., 2007; Ristorcelli et al., 2008, 2009; Cho et al., 2009; Xie et al., 2010; Rountree et al., 2011; Zeelenberg et al., 2011). Furthermore, Viaud et al. (2011) recently reported that highly immunogenic, clinical grade EMVs isolated from interferon-γ treated monocyte-derived DCs express CD40, CD80, CD86, and ICAM-1 on their membranes and can prime CD8+ T cells in a peptide-dependent manner (i.e., MART1, melanoma antigen recognized by T cells) both in vitro and in vivo.
Despite the recent progress made in developing EMVs as vaccines against different types of cancer, a brain cancer-specific EMV vaccine has not yet been reported. This is not surprising since the CNS is considered an immune-privileged site as it lacks a lymphatic system, has low numbers of circulating T lymphocytes, and possesses a BBB consisting of endothelial cells joined by tight junctions, which restricts passage of larger molecules and cells. In addition, high grade gliomas induce immunosuppression in patients, constituting a further challenge to CNS tumor immunotherapy (Bodmer et al., 1989; Misra et al., 2003; Fecci et al., 2006; Avril et al., 2010; Gustafson et al., 2010). To counter this phenomenon, active immunotherapy using autologous DCs pulsed with autologous tumor antigens, as well as GBM-specific antigens including EGFRvIII peptides have been shown to prolong survival in some patients with primary or recurrent GBMs (reviewed in Thomas et al., 2012). Given that EMVs are capable of ferrying antigens that can subsequently pulse DCs, they may serve as a means to administer active immunotherapy against brain cancers. Notably, a recent paper by Alvarez-Erviti et al. (2011) reported brain-targeting EMVs, which cross the BBB following systemic injection into mice with EMVs derived from syngeneic DCs engineered to express a targeting ligand (Alvarez-Erviti et al., 2011; discussed below in EMVs in RNAi therapy). Altogether, these encouraging findings warrant future investigation to examine if EMVs can be employed as an effective CNS cancer vaccine vehicle, thus overcoming immune-privileged properties of the CNS.
EMVs in RNAi therapy
RNA-interference therapies have been actively investigated in the past few decades to target various human diseases, including genetic disorders, HIV infection, and cancers (Burnett and Rossi, 2012). While RNA-based therapies can involve the use of ribozymes, aptamers, and siRNAs, this section will focus on the emerging application of EMVs as a delivery vehicle for therapeutic siRNAs.
Small interfering RNAs are short (∼21–23 nt), single-stranded RNA molecules that target mRNAs with perfect or near-perfect Watson–Crick base-pairing to initiate posttranscriptional gene silencing. In brief, siRNAs can be produced from exogenously introduced double-stranded RNAs (dsRNAs) or short hairpin RNAs (shRNAs) expressed in cells. Upon binding to a pre-RNA-Induced Silencing Complex (RISC) containing Dicer and TAR RNA-binding protein, dsRNAs and shRNAs are processed into siRNAs (passenger or guide) and loaded into RISC complex for mRNA targeting via Watson–Crick based-pairing by the guide strand (Bernstein et al., 2001; Martinez et al., 2002). Argonaute 2 (AGO2), an endonuclease, of the RISC complex then cleaves the target mRNA to inhibit gene expression (Matranga et al., 2005; Rand et al., 2005). Importantly, since the guide strand remains protected from degradation within the RISC complex, it can be used repeatedly to degrade other targeted mRNAs (Matranga et al., 2005; Rand et al., 2005). With these properties, siRNA has been considered as an ideal candidate for RNAi therapy (Burnett and Rossi, 2012).
However, aside from its off-target effects, one of the major challenges confronted by siRNA therapy lies in its delivery formulation in vivo. siRNAs can be immunogenic and are inherently prone to degradation due to ribonucleases present in the extracellular space, serum and cells (Whitehead et al., 2011). While strategies such as chemical modification of siRNAs to counter degradation have been developed to minimize these drawbacks, a vehicle to deliver and shelter siRNAs from external environments, as well as to mediate targeting to specific cells has long been sought-after (Castanotto and Rossi, 2009). Vehicles such as liposomes and nanoparticles have been recruited to serve these functions but are faced by immunogenicity issues (see EMVs as therapeutic delivery vehicles) and difficulty in transversing the BBB in the CNS, making siRNA delivery to the brain particularly difficult (Shim and Kwon, 2010; see EMVs in cancer immunotherapy).
Extracellular membrane vesicles, on the other hand, have been shown to preserve mRNAs and miRNAs within their “aqueous” proteinaceous core even under external RNase treatment, and subsequently to deliver functional RNAs to recipient cells (Valadi et al., 2007; Skog et al., 2008; Zomer et al., 2010; Mathivanan et al., 2011). Remarkably, Alvarez-Erviti et al. (2011) recently reported an exciting strategy targeting EMVs to the brain via systemic injection in mice. By fusing neuron-targeting rabies viral glycoprotein (RVG) peptides to the N-terminus of Lamp2b, a murine exosomal membrane protein, and expressing it in immature DC derived from mouse bone marrow, this team successfully isolated brain-targeting EMVs (Alvarez-Erviti et al., 2011). The EMVs were loaded with siRNAs by electroporation which targeted either GAPDH or BACE1 mRNAs, the latter being a therapeutic target in AD. These EMVs were injected intravenously into syngeneic mice, resulting in a significant knock-down of expression of the targeted mRNAs in the brain when compared to other body tissues (Alvarez-Erviti et al., 2011). Furthermore, two studies have recently described a “zipcode”-like sequence in the 3′ untranslated region (3′UTR) of enriched mRNAs in EMVs derived from human primary GBM and melanoma cells (Batagov et al., 2011; Bolukbasi et al., 2012). This suggests that mRNAs, and perhaps therapeutic siRNAs, can be specifically targeted for EMV packaging in cells. While more studies are required to address the use of EMVs in clinical trials, these novel findings shed light on the promising potential of EMV-mediated RNAi therapy.
EMVs in drug therapy
Like other therapeutic strategies, in vivo delivery of conventional therapeutic drugs has also faced similar challenges, including targeted delivery to tissues/cells, poor drug stability and rapid metabolic degradation. To explore EMV’s potential as a drug delivery vehicle, Sun et al. (2009) first reported successful loading of curcumin, a polyphenol anti-inflammatory compound, into EMVs (“exosomal curcumin”). “Exosomal curcumin” exhibited higher solubility and bioavailability than curcumin alone, and “exosomal curcumin” significantly decreased lipopolysaccharide (LPS)-induced inflammatory activity both in vitro and in vivo more effectively than curcumin itself. It is worth noting, however, that unlike “typical exosomes” which band between 30 and 45% sucrose after gradient density centrifugation, “exosomal curcumin” banded between 45 and 60% sucrose (Sun et al., 2009), suggesting that “exosomal curcumin” may comprise a subpopulation of EMVs and/or the added molecular weight from curcumin loads. Following these encouraging findings, the same group reported successful delivery of “exosomal curcumin” and “exosomal JSI124,” a signal transducer and activator of transcription 3 (Stat3) inhibitor, to the rodent brain via intranasal injection, thereby bypassing the BBB (Zhuang et al., 2011). Remarkably, EMV-mediated delivery of curcumin significantly suppressed LPS-induced inflammation, as well as myelin oligodendrocyte glycoprotein-induced experimental autoimmune encephalomyelitis (Zhuang et al., 2011), an animal model for human CNS demyelinating diseases such as multiple sclerosis and acute disseminated encephalomyelitis (Miller and Karpus, 2007). Additionally, “exosomal JSI124” delivered via intranasal administration suppressed GL26 glioma growth in the brain (Zhuang et al., 2011). Although the authors reported no apparent toxicity or aberrant behavior in the animals during and after treatment, more detailed studies are required to establish the safety parameters of intranasal administration of EMVs, such as dosage and potential immunogenicity. Encouraging findings from these studies support a new means for drug delivery and warrant upcoming investigations to test EMV packaging of other therapeutic compounds, EMV immunogenicity, as well as route of delivery across the BBB for future clinical considerations.
Conclusion
With the emergence of EMVs as a de novo extracellular organelle for cell-to-cell communication, researchers have gathered and studied the role of EMVs under both physiological and pathological conditions, as well as their applications for therapies. In the present review, we have focused on recent discoveries of EMVs’ involvement in the nervous system and EMV-mediated therapies developed to-date. While an impressive number of exciting findings have been made in the past few years, many questions still remain to be answered with respect to different aspects of EMV biology. Due to the different cellular origins and biogenesis of EMVs, standardized nomenclature and isolation protocols for EMVs need to be established by the research community for better advancement of ongoing EMV research. Meanwhile, although a majority of the studies to-date has focused on their involvement in diseases, relatively few have reported on EMV’s physiological role during development and adult functions in the nervous system. Understanding EMVs’ half-life, circulation, and release of cargoes in vivo will also be needed to illuminate this intricate intercellular communication system within the body. Altogether, future investigations and exciting findings in EMVs should further reveal how multiple cellular populations communicate and interact, as well as how EMVs can be employed in therapies.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We wish to thank Ms. Suzanne McDavitt for skilled editorial assistance. This review was supported by grants NIH/NCI CA141150 and CA069246 (Xandra Owen Breakefield) and the Canadian Institutes of Health Research (Charles Pin-Kuang Lai).
References
Allan, D., Thomas, P., and Limbrick, A. R. (1980). The isolation and characterization of 60 nm vesicles (“nanovesicles”) produced during ionophore A23187-induced budding of human erythrocytes. Biochem. J. 188, 881.
Allen, T. M., Hansen, C., Martin, F., Redemann, C., and Yau-Young, A. (1991). Liposomes containing synthetic lipid derivatives of poly(ethylene glycol) show prolonged circulation half-lives in vivo. Biochim. Biophys. Acta 1066, 29–36.
Allen, T. M., Hansen, C., and Rutledge, J. (1989). Liposomes with prolonged circulation times: factors affecting uptake by reticuloendothelial and other tissues. Biochim. Biophys. Acta 981, 27–35.
Al-Nedawi, K., Meehan, B., Micallef, J., Lhotak, V., May, L., Guha, A., and Rak, J. (2008). Intercellular transfer of the oncogenic receptor EGFRvIII by microvesicles derived from tumor cells. Nat. Cell Biol. 10, 619–624.
Al-Nedawi, K., Meehan, B., and Rak, J. (2009). Microvesicles: messengers and mediators of tumor progression. Cell Cycle 8, 2014–2018.
Alvarez-Erviti, L., Seow, Y., Yin, H., Betts, C., Lakhal, S., and Wood, M. J. (2011). Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat. Biotechnol. 29, 341–345.
André, F., Chaput, N., Schartz, N. E. C., Flament, C., Aubert, N., Bernard, J., Lemonnier, F., Raposo, G., Escudier, B., Hsu, D.-H., Tursz, T., Amigorena, S., Angevin, E., and Zitvogel, L. (2004). Exosomes as potent cell-free peptide-based vaccine. I. Dendritic cell-derived exosomes transfer functional MHC class I/peptide complexes to dendritic cells. J. Immunol. 172, 2126–2136.
André, F., Schartz, N. E. C., Movassagh, M., Flament, C., Pautier, P., Morice, P., Pomel, C., Lhomme, C., Escudier, B., Le Chevalier, T., Tursz, T., Amigorena, S., Raposo, G., Angevin, E., and Zitvogel, L. (2002). Malignant effusions and immunogenic tumor-derived exosomes. Lancet 360, 295–305.
Avril, T., Saikali, S., Vauleon, E., Jary, A., Hamlat, A., De Tayrac, M., Mosser, J., and Quillien, V. (2010). Distinct effects of human glioblastoma immunoregulatory molecules programmed cell death ligand-1 (PDL-1) and indoleamine 2,3-dioxygenase (IDO) on tumor-specific T cell functions. J. Neuroimmunol. 225, 22–33.
Bai, O., Li, F., Yuan, J., Laferte, S., and Xiang, J. (2007). Mature dendritic cells pulsed with exosomes stimulate efficient cytotoxic T-lymphocyte responses and antitumor immunity. Immunology 120, 90–102.
Bakhti, M., Winter, C., and Simons, M. (2011). Inhibition of myelin membrane sheath formation by oligodendrocyte-derived exosome-like vesicles. J. Biol. Chem. 286, 787–796.
Balaj, L., Lessard, R., Dai, L., Cho, Y.-J., Pomeroy, S. L., Breakefield, X. O., and Skog, J. (2011). Tumor microvesicles contain retrotransposon elements and amplified oncogene sequences. Nat. Commun. 2, 180.
Batagov, A. O., Kuznetsov, V. A., and Kurochkin, I. V. (2011). Identification of nucleotide patterns enriched in secreted RNAs as putative cis-acting elements targeting them to exosome nano-vesicles. BMC Genomics 12(Suppl. 3), S18. doi:10.1186/1471-2164-12-S3-S18
Batista, B. S., Eng, W. S., Pilobello, K. T., Hendricks-Muñoz, K. D., and Mahal, L. K. (2011). Identification of a conserved glycan signature for microvesicles. J. Proteome Res. 10, 4624–4633.
Beauvillain, C., Ruiz, S., Guiton, R., Bout, D., and Dimier-Poisson, I. (2007). A vaccine based on exosomes secreted by a dendritic cell line confers protection against T. gondii infection in syngeneic and allogeneic mice. Microbes Infect. 9, 1614–1622.
Bellingham, S. A., Guo, B. B., Coleman, B. M., and Hill, A. F. (2012). Exosomes: vehicles for the transfer of toxic proteins associated with neurodegenerative diseases? Front. Physiol. 3:124. doi:10.3389/fphys.2012.00124
Bergsmedh, A., Szeles, A., Henriksson, M., Bratt, A., Folkman, M. J., Spetz, A. L., and Holmgren, L. (2001). Horizontal transfer of oncogenes by uptake of apoptotic bodies. Proc. Natl. Acad. Sci. U.S.A. 98, 6407–6411.
Bernstein, E., Caudy, A. A., Hammond, S. M., and Hannon, G. J. (2001). Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature 409, 363–366.
Bianco, F., Perrotta, C., Novellino, L., Francolini, M., Riganti, L., Menna, E., Saglietti, L., Schuchman, E. H., Furlan, R., Clementi, E., Matteoli, M., and Verderio, C. (2009). Acid sphingomyelinase activity triggers microparticle release from glial cells. EMBO J. 28, 1043–1054.
Bodmer, S., Strommer, K., Frei, K., and Siepl, C. (1989). Immunosuppression and transforming growth factor-beta in glioblastoma. Preferential production of transforming growth factor-beta 2. J. Immunol. 143, 3222–3229.
Bolukbasi, M. F., Mizrak, A., Ozdener, G. B., Madlener, S., Ströbel, T., Erkan, E. P., Fan, J.-B., Breakefield, X. O., and Saydam, O. (2012). miR-1289 and “Zipcode-”like sequence enrich mRNAs in microvesicles. Mol. Ther. Nucleic Acids 1, e10.
Brody, I., Ronquist, G., and Gottfries, A. (1983). Ultrastructural localization of the prostasome – an organelle in human seminal plasma. Ups. J. Med. Sci. 88, 63–80.
Bu, N., Wu, H., Sun, B., Zhang, G., Zhan, S., Zhang, R., and Zhou, L. (2011). Exosome-loaded dendritic cells elicit tumor-specific CD8 + cytotoxic T cells in patients with glioma. J. Neurooncol. 104, 659–667.
Bucki, R., Bachelot-Loza, C., Zachowski, A., Giraud, F., and Sulpice, J. C. (1998). Calcium induces phospholipid redistribution and microvesicle release in human erythrocyte membranes by independent pathways. Biochemistry 37, 15383–15391.
Burnett, J. C., and Rossi, J. J. (2012). RNA-based therapeutics: current progress and future prospects. Chem. Biol. 19, 60–71.
Cadigan, K. M. (2002). Regulating morphogen gradients in the Drosophila wing. Semin. Cell Dev. Biol. 13, 83–90.
Campbell, R. M., and Peterson, A. C. (1993). Expression of a lacZ transgene reveals floor plate cell morphology and macromolecular transfer to commissural axons. Development 119, 1217–1228.
Castanotto, D., and Rossi, J. J. (2009). The promises and pitfalls of RNA-interference-based therapeutics. Nature 457, 426–433.
Castellana, D., Zobairi, F., Martinez, M. C., Panaro, M. A., Mitolo, V., Freyssinet, J. M., and Kunzelmann, C. (2009). Membrane microvesicles as actors in the establishment of a favorable prostatic tumoral niche: a role for activated fibroblasts and CX3CL1-CX3CR1 axis. Cancer Res. 69, 785–793.
Ceruti, S., Colombo, L., Magni, G., Viganò, F., Boccazzi, M., Deli, M. A., Sperlágh, B., Abbracchio, M. P., and Kittel, A. (2011). Oxygen-glucose deprivation increases the enzymatic activity and the microvesicle-mediated release of ectonucleotidases in the cells composing the blood-brain barrier. Neurochem. Int. 59, 259–271.
Chaput, N., Schartz, N., André, F., Taïeb, J., Novault, S., Bonnaventure, P., Aubert, N., Bernard, J., Lemonnier, F., Merad, M., Adema, G., Adams, M., Ferrantini, M., Carpentier, A. F., Escudier, B., Tursz, T., Angevin, E., Zitvogel L. (2004). Exosomes as potent cell-free peptide-based vaccine. II. Exosomes in CpG adjuvants efficiently prime naive Tc1 lymphocytes leading to tumor rejection. J. Immunol. 172, 2137–2146.
Chen, W., Wang, J., Shao, C., Liu, S., Yu, Y., Wang, Q., and Cao, X. (2006). Efficient induction of antitumor T cell immunity by exosomes derived from heat-shocked lymphoma cells. Eur. J. Immunol. 36, 1598–1607.
Chen, X., Liang, H., Zhang, J., Zen, K., and Zhang, C.-Y. (2012). Horizontal transfer of microRNAs: molecular mechanisms and clinical applications. Protein Cell 3, 28–37.
Cho, J., Lee, Y., Kim, S., and Ko, J. (2009). MHC independent anti-tumor immune responses induced by Hsp70-enriched exosomes generate tumor regression in murine models. Cancer Lett. 275, 256–265.
Cho, J. A., Yeo, D.-J., Son, H.-Y., Kim, H.-W., Jung, D.-S., Ko, J.-K., Koh, J. S., Kim, Y.-N., and Kim, C. W. (2005). Exosomes: a new delivery system for tumor antigens in cancer immunotherapy. Int. J. Cancer 114, 613–622.
Chu, Z., Witte, D. P., and Qi, X. (2005). Saposin C-LBPA interaction in late-endosomes/lysosomes. Exp. Cell Res. 303, 300–307.
Cocucci, E., Racchetti, G., and Meldolesi, J. (2009). Shedding microvesicles: artefacts no more. Trends Cell Biol. 19, 43–51.
Cocucci, E., Racchetti, G., Podini, P., and Meldolesi, J. (2007). Enlargeosome traffic: exocytosis triggered by various signals is followed by endocytosis, membrane shedding or both. Traffic 8, 742–757.
Collino, F., Deregibus, M. C., Bruno, S., Sterpone, L., Aghemo, G., Viltono, L., Tetta, C., and Camussi, G. (2010). Microvesicles derived from adult human bone marrow and tissue specific mesenchymal stem cells shuttle selected pattern of miRNAs. PLoS ONE 5, e11803. doi:10.1371/journal.pone.0011803
Contreras-Galindo, R., Kaplan, M. H., Leissner, P., Verjat, T., Ferlenghi, I., Bagnoli, F., Giusti, F., Dosik, M. H., Hayes, D. F., Gitlin, S. D., and Markovitz, D. M. (2008). Human endogenous retrovirus K (HML-2) elements in the plasma of people with lymphoma and breast cancer. J. Virol. 82, 9329–9336.
Coufal, N. G., Garcia-Perez, J. L., Peng, G. E., Yeo, G. W., Mu, Y., Lovci, M. T., Morell, M., O’Shea, K. S., Moran, J. V., and Gage, F. H. (2009). L1 retrotransposition in human neural progenitor cells. Nature 460, 1127–1131.
Court, F. A., Hendriks, W. T., MacGillavry, H. D., Alvarez, J., and van Minnen, J. (2008). Schwann cell to axon transfer of ribosomes: toward a novel understanding of the role of glia in the nervous system. J. Neurosci. 28, 11024–11029.
Court, F. A., Midha, R., Cisterna, B. A., Grochmal, J., Shakhbazau, A., Hendriks, W. T., and Van Minnen, J. (2011). Morphological evidence for a transport of ribosomes from Schwann cells to regenerating axons. Glia 59, 1529–1539.
Dargan, D. J., and Subak-Sharpe, J. H. (1997). The effect of herpes simplex virus type 1 L-particles on virus entry, replication, and the infectivity of naked herpesvirus DNA. Virology 239, 378–388.
Davis, D. M., and Sowinski, S. (2008). Membrane nanotubes: dynamic long-distance connections between animal cells. Nat. Rev. Mol. Cell Biol. 9, 431–436.
Del Conde, I., Shrimpton, C. N., Thiagarajan, P., and López, J. A. (2005). Tissue-factor-bearing microvesicles arise from lipid rafts and fuse with activated platelets to initiate coagulation. Blood 106, 1604–1611.
Denzer, K., Kleijmeer, M., and Heijnen, H. (2000). Exosome: from internal vesicle of the multivesicular body to intercellular signaling device. J. Cell Sci. 19, 3365–3374.
Di Vizio, D., Kim, J., Hager, M. H., Morello, M., Yang, W., Lafargue, C. J., True, L. D., Rubin, M. A., Adam, R. M., Beroukhim, R., Demichelis, F., and Freeman, M. R. (2009). Oncosome formation in prostate cancer: association with a region of frequent chromosomal deletion in metastatic disease. Cancer Res. 69, 5601–5609.
Dolo, V., Li, R., Dillinger, M., Flati, S., Manela, J., Taylor, B. J., Pavan, A., and Ladisch, S. (2000). Enrichment and localization of ganglioside G(D3) and caveolin-1 in shed tumor cell membrane vesicles. Biochim. Biophys. Acta 1486, 265–274.
Duelli, D., and Lazebnik, Y. (2007). Cell-to-cell fusion as a link between viruses and cancer. Nat. Rev. Cancer 7, 968–976.
Ecroyd, H., Sarradin, P., Dacheux, J. L., and Gatti, J. L. (2004). Compartmentalization of prion isoforms within the reproductive tract of the ram. Biol. Reprod. 71, 993–1001.
Eisele, Y. S., Obermüller, U., Heilbronner, G., Baumann, F., Kaeser, S. A., Wolburg, H., Walker, L. C., Staufenbiel, M., Heikenwalder, M., and Jucker, M. (2010). Peripherally applied Abeta-containing inoculates induce cerebral beta-amyloidosis. Science 330, 980–982.
Emmanouilidou, E., Melachroinou, K., Roumeliotis, T., Garbis, S. D., Ntzouni, M., Margaritis, L. H., Stefanis, L., and Vekrellis, K. (2010). Cell-produced alpha-synuclein is secreted in a calcium-dependent manner by exosomes and impacts neuronal survival. J. Neurosci. 30, 6838–6851.
Fauré, J., Lachenal, G., Court, M., Hirrlinger, J., Chatellard-Causse, C., Blot, B., Grange, J., Schoehn, G., Goldberg, Y., Boyer, V., Kirchhoff, F., Raposo, G., Garin, J., and Sadoul, R. (2006). Exosomes are released by cultured cortical neurones. Mol. Cell. Neurosci. 31, 642–648.
Fecci, P. E., Mitchell, D. A., Whitesides, J. F., Xie, W., Friedman, A. H., Archer, G. E., Herndon, J. E., Bigner, D. D., Dranoff, G., and Sampson, J. H. (2006). Increased regulatory T-cell fraction amidst a diminished CD4 compartment explains cellular immune defects in patients with malignant glioma. Cancer Res. 66, 3294–3302.
Fevrier, B., Vilette, D., Archer, F., Loew, D., Faigle, W., Vidal, M., Laude, H., and Raposo, G. (2004). Cells release prions in association with exosomes. Proc. Natl. Acad. Sci. U.S.A. 101, 9683–9688.
Février, B., Vilette, D., Laude, H., and Raposo, G. (2005). Exosomes: a bubble ride for prions? Traffic 6, 10–17.
Flanagan, J., Middeldorp, J., and Sculley, T. (2003). Localization of the Epstein-Barr virus protein LMP 1 to exosomes. J. Gen. Virol. 84, 1871–1879.
Frühbeis, C., Fröhlich, D., and Krämer-Albers, E. M. (2012). Emerging roles of exosomes in neuron-glia communication. Front. Physiol. 3:119. doi:10.3389/fphys.2012.00119
Gannon, P., Khan, M. Z., and Kolson, D. L. (2011). Current understanding of HIV-associated neurocognitive disorders pathogenesis. Curr. Opin. Neurol. 24, 275–283.
Gerstner, E. R., and Batchelor, T. T. (2010). Primary central nervous system lymphoma. Arch. Neurol. 67, 291–297.
Gomes, C., Keller, S., Altevogt, P., and Costa, J. (2007). Evidence for secretion of Cu,Zn superoxide dismutase via exosomes from a cell model of amyotrophic lateral sclerosis. Neurosci. Lett. 428, 43–46.
Goodenough, D. A., and Paul, D. L. (2009). Gap junctions. Cold Spring Harb. Perspect. Biol. 1, a002576.
Gould, S. J., Booth, A. M., and Hildreth, J. E. (2003). The Trojan exosome hypothesis. Proc. Natl. Acad. Sci. U.S.A. 100, 10592–10597.
Gousset, K., and Zurzolo, C. (2009). Tunnelling nanotubes: a highway for prion spreading? Prion 3, 94–98.
Graner, M. W., Alzate, O., Dechkovskaia, A. M., Keene, J. D., Sampson, J. H., Mitchell, D. A., and Bigner, D. D. (2009). Proteomic and immunologic analyses of brain tumor exosomes. FASEB J. 23, 1541–1557.
Greco, V., Hannus, M., and Eaton, S. (2001). Argosomes: a potential vehicle for the spread of morphogens through epithelia. Cell 106, 633–645.
Guescini, M., Genedani, S., Stocchi, V., and Agnati, L. F. (2010). Astrocytes and Glioblastoma cells release exosomes carrying mtDNA. J. Neural Transm. 117, 1–4.
Guest, W. C., Silverman, J. M., Pokrishevsky, E., O’Neill, M. A., Grad, L. I., and Cashman, N. R. (2011). Generalization of the prion hypothesis to other neurodegenerative diseases: an imperfect fit. J. Toxicol. Environ. Health 74, 1433–1459.
Guo, F., Chang, C. K., Fan, H. H., Nie, X. X., Ren, Y. N., Liu, Y. Y., and Zhao, L. H. (2008). Anti-tumor effects of exosomes in combination with cyclophosphamide and polyinosinic-polycytidylic acid. J. Int. Med. Res. 36, 1342–1353.
Gustafson, M., Lin, Y., and New, K. (2010). Systemic immune suppression in glioblastoma: the interplay between CD14 + HLA-DRlo/neg monocytes, tumor factors, and dexamethasone. Neuro-oncol. 12, 631–644.
Hansen, C., Angot, E., Bergström, A. L., Steiner, J. A., Pieri, L., Paul, G., Outeiro, T. F., Melki, R., Kallunki, P., Fog, K., Li, J. Y., and Brundin, P. (2011). α-Synuclein propagates from mouse brain to grafted dopaminergic neurons and seeds aggregation in cultured human cells. J. Clin. Invest. 121, 715–727.
Hansen, C., and Li, J. Y. (2012). Beyond α-synuclein transfer: pathology propagation in Parkinson’s disease. Trends Mol. Med. 18, 248–255.
Hao, S., Bai, O., Li, F., Yuan, J., Laferte, S., and Xiang, J. (2007). Mature dendritic cells pulsed with exosomes stimulate efficient-cytotoxic T-lymphocyte responses and antitumor immunity. Immunology 120, 90–102.
Hegmans, J., Hemmes, A., and Aerts, J. (2005). Immunotherapy of murine malignant mesothelioma using tumor lysate-pulsed dendritic cells. Am. J. Respir. Crit. Care Med. 171, 1168–1177.
Heijnen, H. F., Schiel, A. E., Fijnheer, R., Geuze, H. J., and Sixma, J. J. (1999). Activated platelets release two types of membrane vesicles: microvesicles by surface shedding and exosomes derived from exocytosis of multivesicular bodies and alpha-granules. Blood 94, 3791–3799.
Henson, P. M., Bratton, D. L., and Fadok, V. A. (2001). Apoptotic cell removal. Curr. Biol. 11, R795–R805.
Holmgren, L. (2010). Horizontal gene transfer: you are what you eat. Biochem. Biophys. Res. Commun. 396, 147–151.
Holmgren, L., Szeles, A., Rajnavölgyi, E., Folkman, J., Klein, G., Ernberg, I., and Falk, K. I. (1999). Horizontal transfer of DNA by the uptake of apoptotic bodies. Blood 93, 3956–3963.
Hristov, M., Erl, W., Linder, S., and Weber, P. C. (2004). Apoptotic bodies from endothelial cells enhance the number and initiate the differentiation of human endothelial progenitor cells in vitro. Blood 104, 2761–2766.
Hsu, C., Morohashi, Y., Yoshimura, S., Manrique-Hoyos, N., Jung, S., Lauterbach, M. A., Bakhti, M., Grønborg, M., Möbius, W., Rhee, J., Barr, F. A., and Simons, M. (2010). Regulation of exosome secretion by Rab35 and its GTPase-activating proteins TBC1D10A-C. J. Cell Biol. 189, 223–232.
Ichim, T. E., Zhong, Z., Kaushal, S., Zheng, X., Ren, X., Hao, X., Joyce, J. A., Hanley, H. H., Riordan, N. H., Koropatnick, J., Bogin, V., Minev, B. R., Min, W. P., and Tullis, R. H. (2008). Exosomes as a tumor immune escape mechanism: possible therapeutic implications. J. Transl. Med. 6, 37.
Iero, M., Valenti, R., Huber, V., Filipazzi, P., Parmiani, G., Fais, S., and Rivoltini, L. (2008). Tumor-released exosomes and their implications in cancer immunity. Cell Death Differ. 15, 80–88.
Immordino, M. L., Dosio, F., and Cattel, L. (2006). Stealth liposomes: review of the basic science, rationale, and clinical applications, existing and potential. Int. J. Nanomedicine 1, 297–315.
Jouvenet, N., Zhadina, M., Bieniasz, P. D., and Simon, S. M. (2011). Dynamics of ESCRT protein recruitment during retroviral assembly. Nat. Cell Biol. 13, 394–401.
Khatua, A. K., Taylor, H. E., Hildreth, J. E., and Popik, W. (2009). Exosomes packaging APOBEC3G confer human immunodeficiency virus resistance to recipient cells. J. Virol. 83, 512–521.
Kim, C. W., Lee, H. M., Lee, T. H., Kang, C., Kleinman, H. K., and Gho, Y. S. (2002). Extracellular membrane vesicles from tumor cells promote angiogenesis via sphingomyelin. Cancer Res. 62, 6312–6317.
Klibi, J., Niki, T., Riedel, A., Pioche-Durieu, C., Souquere, S., Rubinstein, E., Le Moulec, S., Guigay, J., Hirashima, M., Guemira, F., Adhikary, D., Mautner, J., and Busson, P. (2009). Blood diffusion and Th1-suppressive effects of galectin-9-containing exosomes released by Epstein-Barr virus-infected nasopharyngeal carcinoma cells. Blood 113, 1957–1966.
Kobayashi, T., Okamoto, H., Yamada, J., Setaka, M., and Kwan, T. (1984). Vesiculation of platelet plasma membranes. Dilauroylglycerophosphocholine-induced shedding of a platelet plasma membrane fraction enriched in acetylcholinesterase activity. Biochim. Biophys. Acta 778, 210–218.
Korkut, C., Ataman, B., Ramachandran, P., Ashley, J., Barria, R., Gherbesi, N., and Budnik, V. (2009). Trans-synaptic transmission of vesicular Wnt signals through Evi/Wntless. Cell 139, 393–404.
Kosaka, N., Iguchi, H., Yoshioka, Y., Takeshita, F., Matsuki, Y., and Ochiya, T. (2010). Secretory mechanisms and intercellular transfer of microRNAs in living cells. J. Biol. Chem. 285, 17442–17452.
Krämer-Albers, E. M., Bretz, N., Tenzer, S., Winterstein, C., Möbius, W., Berger, H., Nave, K. A., Schild, H., and Trotter, J. (2007). Oligodendrocytes secrete exosomes containing major myelin and stress-protective proteins: trophic support for axons? Proteomics Clin. Appl. 1, 1446–1461.
Krishnan, L., and Sprott, G. D. (2008). Archaeosome adjuvants: immunological capabilities and mechanism(s) of action. Vaccine 26, 2043–2055.
Lachenal, G., Pernet-Gallay, K., Chivet, M., Hemming, F. J., Belly, A., Bodon, G., Blot, B., Haase, G., Goldberg, Y., and Sadoul, R. (2011). Release of exosomes from differentiated neurons and its regulation by synaptic glutamatergic activity. Mol. Cell. Neurosci. 46, 409–418.
Lakkaraju, A., and Rodriguez-Boulan, E. (2008). Itinerant exosomes: emerging roles in cell and tissue polarity. Trends Cell Biol. 18, 199–209.
Lavie, L., Kitova, M., Maldener, E., Meese, E., and Mayer, J. (2005). CpG methylation directly regulates transcriptional activity of the human endogenous retrovirus family HERV-K(HML-2). J. Virol. 79, 876–883.
Lee, T. H., D’Asti, E., Magnus, N., Al-Nedawi, K., Meehan, B., and Rak, J. (2011). Microvesicles as mediators of intercellular communication in cancer–the emerging science of cellular “debris.” Semin. Immunopathol. 33, 455–467.
Lehmann, B., Paine, M., Brooks, A., and McCubrey, J. (2008). Senescence-associated exosome release from human prostate cancer cells. Cancer Res. 68, 7864–7871.
Lenassi, M., Cagney, G., Liao, M., Vaupotic, T., Bartholomeeusen, K., Cheng, Y., Krogan, N. J., Plemenitas, A., and Peterlin, B. M. (2010). HIV Nef is secreted in exosomes and triggers apoptosis in bystander CD4+ T cells. Traffic 11, 110–122.
Llorente, A., van Deurs, B., and Sandvig, K. (2007). Cholesterol regulates prostasome release from secretory lysosomes in PC-3 human prostate cancer cells. Eur. J. Cell Biol. 86, 405–415.
Lo Cicero, A., Schiera, G., Proia, P., Saladino, P., Savettieri, G., Di Liegro, C. M., and Di Liegro, I. (2011). Oligodendroglioma cells shed microvesicles which contain TRAIL as well as molecular chaperones and induce cell death in astrocytes. Int. J. Oncol. 39, 1353–1357.
Lublin, A. L., and Gandy, S. (2010). Amyloid-beta oligomers: possible roles as key neurotoxins in Alzheimer’s disease. Mt. Sinai J. Med. 77, 43–49.
Mack, M., Kleinschmidt, A., Bruhl, H., Klier, C., Nelson, P. J., Cihak, J., Plachy, J., Stangassinger, M., Erfle, V., and Schlondorff, D. (2000). Transfer of the chemokine receptor CCR5 between cells by membrane-derived microparticles: a mechanism for cellular human immunodeficiency virus 1 infection. Nat. Med. 6, 769–775.
Malam, Y., Loizidou, M., and Seifalian, A. M. (2009). Liposomes and nanoparticles: nanosized vehicles for drug delivery in cancer. Trends Pharmacol. Sci. 30, 592–599.
Martinez, J., Patkaniowska, A., Urlaub, H., Lührmann, R., and Tuschl, T. (2002). Single-stranded antisense siRNAs guide target RNA cleavage in RNAi. Cell 110, 563–574.
Marzesco, A. M., Janich, P., Wilsch-Bräuninger, M., Dubreuil, V., Langenfeld, K., Corbeil, D., and Huttner, W. B. (2005). Release of extracellular membrane particles carrying the stem cell marker prominin-1 (CD133) from neural progenitors and other epithelial cells. J. Cell Sci. 118, 2849–2858.
Mathivanan, S., Fahner, C. J., Reid, G. E., and Simpson, R. J. (2011). ExoCarta 2012: database of exosomal proteins, RNA and lipids. Nucleic Acids Res. 40, D1241–D1244.
Mathivanan, S., Fahner, C. J., Reid, G. E., and Simpson, R. J. (2012). ExoCarta 2012: database of exosomal proteins, RNA and lipids. Nucleic Acids Res. 40, D1241–D1244.
Mathivanan, S., Ji, H., and Simpson, R. J. (2010). Exosomes: extracellular organelles important in intercellular communication. J. Proteomics 73, 1907–1920.
Matranga, C., Tomari, Y., Shin, C., and Bartel, D. (2005). Passenger-strand cleavage facilitates assembly of siRNA into Ago2-containing RNAi enzyme complexes. Cell 123, 607–620.
McLauchlan, J., Addison, C., Craigie, M. C., and Rixon, F. J. (1992). Noninfectious L-particles supply functions which can facilitate infection by HSV-1. Virology 190, 682–688.
Meckes, D. G. J., and Raab-Traub, N. (2011). Microvesicles and viral infection. J. Virol. 85, 12844–12854.
Meckes, D. G. J., Shair, K. H., Marquitz, A. R., Kung, C. P., Edwards, R. H., and Raab-Traub, N. (2010). Human tumor virus utilizes exosomes for intercellular communication. Proc. Natl. Acad. Sci. U.S.A. 107, 20370–20375.
Miller, S. D., and Karpus, W. J. (2007). Experimental autoimmune encephalomyelitis in the mouse. Curr. Protoc. Immunol. Chap. 15, Unit 15.1.
Millimaggi, D., Mari, M., D’Ascenzo, S., Carosa, E., Jannini, E. A., Zucker, S., Carta, G., Pavan, A., and Dolo, V. (2007). Tumor vesicle-associated CD147 modulates the angiogenic capability of endothelial cells. Neoplasia 9, 349–357.
Misra, A., Ganesh, S., and Shahiwala, A. (2003). Drug delivery to the central nervous system: a review. J. Pharm. Pharm. Sci. 6, 252–273.
Mittelbrunn, M., Gutiérrez-Vázquez, C., Villarroya-Beltri, C., González, S., Sánchez-Cabo, F., González, M. Á., Bernad, A., and Sánchez-Madrid, F. (2011). Unidirectional transfer of microRNA-loaded exosomes from T cells to antigen-presenting cells. Nat. Commun. 2, 282.
Muralidharan-Chari, V., Clancy, J. W., Sedgwick, A., and D’Souza-Schorey, C. (2010). Microvesicles: mediators of extracellular communication during cancer progression. J. Cell Sci. 123, 1603–1611.
Ostrowski, M., Carmo, N. B., Krumeich, S., Fanget, I., Raposo, G., Savina, A., Moita, C. F., Schauer, K., Hume, A. N., Freitas, R. P., Goud, B., Benaroch, P., Hacohen, N., Fukuda, M., Desnos, C., Seabra, M. C., Darchen, F., Amigorena, S., Moita, L. F., and Thery, C. (2010). Rab27a and Rab27b control different steps of the exosome secretion pathway. Nat. Cell Biol. 12, 19–30.
Parolini, I., Federici, C., Raggi, C., Lugini, L., Palleschi, S., De Milito, A., Coscia, C., Iessi, E., Logozzi, M., Molinari, A., Colone, M., Tatti, M., Sargiacomo, M., and Fais, S. (2009). Microenvironmental pH is a key factor for exosome traffic in tumor cells. J. Biol. Chem. 284, 34211–34222.
Pegtel, D. M., Cosmopoulos, K., Thorley-Lawson, D. A., van Eijndhoven, M. A., Hopmans, E., Lindengey, J. L., de Gruijl, T. D., Wurdinger, T., and Middeldorp, J. M. (2010). Functional delivery of viral miRNAs via exosomes. Proc. Natl. Acad. Sci. U.S.A. 107, 6328–6333.
Piccin, A., Murphy, W. G., and Smith, O. P. (2007). Circulating microparticles: pathophysiology and clinical implications. Blood Rev. 21, 157–171.
Pilzer, D., Gasser, O., Moskovich, O., Schifferli, J. A., and Fishelson, Z. (2005). Emission of membrane vesicles: roles in complement resistance, immunity and cancer. Springer Semin. Immunopathol. 27, 375–387.
Potolicchio, I., Carven, G. J., Xu, X., Stipp, C., Riese, R. J., Stern, L. J., and Santambrogio, L. (2005). Proteomic analysis of microglia-derived exosomes: metabolic role of the aminopeptidase CD13 in neuropeptide catabolism. J. Immunol. 175, 2237–2243.
Putz, U., Howitt, J., Lackovic, J., Foot, N., Kumar, S., Silke, J., and Tan, S. S. (2008). Nedd4 family-interacting protein 1 (Ndfip1) is required for the exosomal secretion of Nedd4 family proteins. J. Biol. Chem. 283, 32621–32627.
Quaite-Randall, E., Trent, J. D., Josephs, R., and Joachimiak, A. (1995). Conformational cycle of the archaeosome, a TCP1-like chaperonin from Sulfolobus shibatae. J. Biol. Chem. 270, 28818–28823.
Rajendran, L., Honsho, M., Zahn, T. R., Keller, P., Geiger, K. D., Verkade, P., and Simons, K. (2006). Alzheimer’s disease beta-amyloid peptides are released in association with exosomes. Proc. Natl. Acad. Sci. U.S.A. 103, 11172–11177.
Ramachandran, S., and Palanisamy, V. (2011). Horizontal transfer of RNAs: exosomes as mediators of intercellular communication. Wiley Interdiscip. Rev. RNA 3, 286–293.
Rand, T. A., Petersen, S., Du, F., and Wang, X. (2005). Argonaute2 cleaves the anti-guide strand of siRNA during RISC activation. Cell 123, 621–629.
Raposo, G., Nijman, H. W., Stoorvogel, W., Liejendekker, R., Harding, C. V., Melief, C. J., and Geuze, H. J. (1996). B lymphocytes secrete antigen-presenting vesicles. J. Exp. Med. 183, 1161–1172.
Ratajczak, J., Miekus, K., Kucia, M., Zhang, J., Reca, R., Dvorak, P., and Ratajczak, M. Z. (2006a). Embryonic stem cell-derived microvesicles reprogram hematopoietic progenitors: evidence for horizontal transfer of mRNA and protein delivery. Leukemia 20, 847–856.
Ratajczak, J., Wysoczynski, M., Hayek, F., Janowska-Wieczorek, A., and Ratajczak, M. Z. (2006b). Membrane-derived microvesicles: important and underappreciated mediators of cell-to-cell communication. Leukemia 20, 1487–1495.
Ristorcelli, E., Beraud, E., Mathieu, S., Lombardo, D., and Verine, A. (2009). Essential role of Notch signaling in apoptosis of human pancreatic tumoral cells mediated by exosomal nanoparticles. Int. J. Cancer 125, 1016–1026.
Ristorcelli, E., Beraud, E., Verrando, P., Villard, C., Lafitte, D., Sbarra, V., Lombardo, D., and Verine, A. (2008). Human tumor nanoparticles induce apoptosis of pancreatic cancer cells. FASEB J. 22, 3358–3369.
Rountree, R. B., Mandl, S. J., Nachtwey, J. M., Dalpozzo, K., Do, L., Lombardo, J. R., Schoonmaker, P. L., Brinkmann, K., Dirmeier, U., Laus, R., and Delcayre, A. (2011). Exosome targeting of tumor antigens expressed by cancer vaccines can improve antigen immunogenicity and therapeutic efficacy. Cancer Res. 71, 5235–5244.
Rozmyslowicz, T., Majka, M., Kijowski, J., Murphy, S. L., Conover, D. O., Poncz, M., Ratajczak, J., Gaulton, G. N., and Ratajczak, M. Z. (2003). Platelet- and megakaryocyte-derived microparticles transfer CXCR4 receptor to CXCR4-null cells and make them susceptible to infection by X4-HIV. AIDS 17, 33–42.
Rustom, A., Saffrich, R., Markovic, I., Walther, P., and Gerdes, H.-H. (2004). Nanotubular highways for intercellular organelle transport. Science 303, 1007.
Sakar, Y., Nazaret, C., Lettéron, P., Ait Omar, A., Avenati, M., Viollet, B., Ducroc, R., and Bado, A. (2009). Positive regulatory control loop between gut leptin and intestinal GLUT2/GLUT5 transporters links to hepatic metabolic functions in rodents. PLoS ONE 4, e7935. doi:10.1371/journal.pone.0007935
Salzer, U., Hinterdorfer, P., Hunger, U., Borken, C., and Prohaska, R. (2002). Ca(++)-dependent vesicle release from erythrocytes involves stomatin-specific lipid rafts, synexin (annexin VII), and sorcin. Blood 99, 2569–2577.
Saman, S., Kim, W., Raya, M., Visnick, Y., Miro, S., Saman, S., Jackson, B., McKee, A. C., Alvarez, V. E., Lee, N. C., and Hall, G. F. (2012). Exosome-associated Tau is secreted in tauopathy models and is selectively phosphorylated in cerebrospinal fluid in early Alzheimer disease. J. Biol. Chem. 287, 3842–3849.
Sang, Q., Kim, M. H., Kumar, S., Bye, N., Morganti-Kossman, M. C., Gunnersen, J., Fuller, S., Howitt, J., Hyde, L., Beissbarth, T., Scott, H. S., Silke, J., and Tan, S. S. (2006). Nedd4-WW domain-binding protein 5 (Ndfip1) is associated with neuronal survival after acute cortical brain injury. J. Neurosci. 26, 7234–7244.
Scott, S., Pendlebury, S. A., and Green, C. (1984). Lipid organization in erythrocyte membrane microvesicles. Biochem. J. 224, 285–290.
Shedden, K., Xie, X. T., Chandaroy, P., Chang, Y. T., and Rosania, G. R. (2003). Expulsion of small molecules in vesicles shed by cancer cells: association with gene expression and chemosensitivity profiles. Cancer Res. 63, 4331–4337.
Sheldon, H., Heikamp, E., Turley, H., Dragovic, R., Thomas, P., Oon, C. E., Leek, R., Edelmann, M., Kessler, B., Sainson, R. C., Sargent, I., Li, J. L., and Harris, A. L. (2010). New mechanism for Notch signaling to endothelium at a distance by Delta-like 4 incorporation into exosomes. Blood 116, 2385–2394.
Shen, B., Wu, N., Yang, J. M., and Gould, S. J. (2011). Protein targeting to exosomes/microvesicles by plasma membrane anchors. J. Biol. Chem. 286, 14383–14395.
Shim, M. S., and Kwon, Y. J. (2010). Efficient and targeted delivery of siRNA in vivo. FEBS J. 277, 4814–4827.
Simak, J., and Gelderman, M. P. (2006). Cell membrane microparticles in blood and blood products: potentially pathogenic agents and diagnostic markers. Transfus. Med. Rev. 20, 1–26.
Simpson, R. J., Jensen, S. S., and Lim, J. W. E. (2008). Proteomic profiling of exosomes: current perspectives. Proteomics 8, 4083–4099.
Skog, J., Würdinger, T., van Rijn, S., Meijer, D. H., Gainche, L., Curry, W. T., Carter, B. S., Krichevsky, A. M., and Breakefield, X. O. (2008). Glioblastoma microvesicles transport RNA and proteins that promote tumor growth and provide diagnostic biomarkers. Nat. Cell Biol. 10, 1470–1476.
Smalheiser, N. R. (2007). Exosomal transfer of proteins and RNAs at synapses in the nervous system. Biol. Direct 2, 35.
Steiner, I. (2011). Herpes simplex virus encephalitis: new infection or reactivation? Curr. Opin. Neurol. 24, 268–274.
Stuart, M. C., Damoiseaux, J. G., Frederik, P. M., Arends, J. W., and Reutelingsperger, C. P. (1998). Surface exposure of phosphatidylserine during apoptosis of rat thymocytes precedes nuclear changes. Eur. J. Cell Biol. 76, 77–83.
Subra, C., Laulagnier, K., Perret, B., and Record, M. (2007). Exosome lipidomics unravels lipid sorting at the level of multivesicular bodies. Biochimie 89, 205–212.
Sun, D., Zhuang, X., Xiang, X., Liu, Y., Zhang, S., Liu, C., Barnes, S., Grizzle, W., Miller, D., and Zhang, H.-G. (2009). A novel nanoparticle drug delivery system: the anti-inflammatory activity of curcumin is enhanced when encapsulated in exosomes. Mol. Ther. 18, 1606–1614.
Taïeb, J., Chaput, N., Schartz, N., Roux, S., Novault, S., Ménard, C., Ghiringhelli, F., Terme, M., Carpentier, A. F., Darrasse-Jèze, G., Lemonnier, F., and Zitvogel, L. (2006). Chemoimmunotherapy of tumors: cyclophosphamide synergizes with exosome based vaccines. J. Immunol. 176, 2722–2729.
Tan, A., De La Peña,., and Seifalian, A. M. (2010). The application of exosomes as a nanoscale cancer vaccine. Int. J. Nanomedicine 5, 889–900.
Tani, E., Nakano, M., Itagaki, T., and Fukumori, T. (1978). Cell membrane structure of human giant-celled glioblastoma. Acta Neuropathol. 41, 61–65.
Taraboletti, G., D’Ascenzo, S., Giusti, I., Marchetti, D., Borsotti, P., Millimaggi, D., Giavazzi, R., Pavan, A., and Dolo, V. (2006). Bioavailability of VEGF in tumor-shed vesicles depends on vesicle burst induced by acidic pH. Neoplasia 8, 96–103.
Taylor, A. R., Robinson, M. B., Gifondorwa, D. J., Tytell, M., and Milligan, C. E. (2007). Regulation of heat shock protein 70 release in astrocytes: role of signaling kinases. Dev. Neurobiol. 67, 1815–1829.
Taylor, D. D., and Gerçel-Taylor, C. (2005). Tumor-derived exosomes and their role in cancer-associated T-cell signalling defects. Br. J. Cancer 92, 305–311.
Temchura, V. V., Tenbusch, M., Nchinda, G., Nabi, G., Tippler, B., Zelenyuk, M., Wildner, O., Uberla, K., and Kuate, S. (2008). Enhancement of immunostimulatory properties of exosomal vaccines by incorporation of fusion-competent G protein of vesicular stomatitis virus. Vaccine 26, 3662–3672.
Temme, S., Eis-Hübinger, A. M., McLellan, A. D., and Koch, N. (2010). The herpes simplex virus-1 encoded glycoprotein B diverts HLA-DR into the exosome pathway. J. Immunol. 184, 236–243.
Teo, B. H., and Wong, S. H. (2010). MHC class II-associated invariant chain (Ii) modulates dendritic cells-derived microvesicles (DCMV)-mediated activation of microglia. Biochem. Biophys. Res. Commun. 400, 673–678.
Théry, C., Boussac, M., Véron, P., Ricciardi-Castagnoli, P., Raposo, G., Garin, J., and Amigorena, S. (2001). Proteomic analysis of dendritic cell-derived exosomes: a secreted subcellular compartment distinct from apoptotic vesicles. J. Immunol. 166, 7309–7318.
Théry, C., Ostrowski, M., and Segura, E. (2009). Membrane vesicles as conveyors of immune responses. Nat. Rev. Immunol. 9, 581–593.
Thomas, A. A., Ernstoff, M. S., and Fadul, C. E. (2012). Immunotherapy for the treatment of glioblastoma. Cancer J. 18, 59–68.
Trajkovic, K., Hsu, C., Chiantia, S., Rajendran, L., Wenzel, D., Wieland, F., Schwille, P., Brügger, B., and Simons, M. (2008). Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science 319, 1244–1247.
Trumpfheller, C., Longhi, M. P., Caskey, M., Idoyaga, J., Bozzacco, L., Keler, T., Schlesinger, S. J., and Steinman, R. M. (2012). Dendritic cell-targeted protein vaccines: a novel approach to induce T-cell immunity. J. Intern. Med. 271, 183–192.
Turchinovich, A., Weiz, L., Langheinz, A., and Burwinkel, B. (2011). Characterization of extracellular circulating microRNA. Nucleic Acids Res. 39, 7223–7233.
Twiss, J. L., and Fainzilber, M. (2009). Ribosomes in axons–scrounging from the neighbors? Trends Cell Biol. 19, 236–243.
Valadi, H., Ekström, K., Bossios, A., Sjöstrand, M., Lee, J. J., and Lötvall, J. O. (2007). Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 9, 654–659.
Vallhov, H., Gutzeit, C., Johansson, S. M., Nagy, N., Paul, M., Li, Q., Friend, S., George, T. C., Klein, E., Scheynius, A., and Gabrielsson, S. (2011). Exosomes containing glycoprotein 350 released by EBV-transformed B cells selectively target B cells through CD21 and block EBV infection in vitro. J. Immunol. 186, 73–82.
van der Vos, K. E., Balaj, L., Skog, J., and Breakefield, X. O. (2011). Brain tumor microvesicles: insights into intercellular communication in the nervous system. Cell. Mol. Neurobiol. 31, 949–959.
van Dommelen, S. M., Vader, P., Lakhal, S., Kooijmans, S. A. A., van Solinge, W. W., Wood, M. J. A., and Schiffelers, R. M. (2011). Microvesicles and exosomes: opportunities for cell-derived membrane vesicles in drug delivery. J. Control Release 1–10.
Vella, L. J., Sharples, R. A., Lawson, V. A., Masters, C. L., Cappai, R., and Hill, A. F. (2007). Packaging of prions into exosomes is associated with a novel pathway of PrP processing. J. Pathol. 211, 582–590.
Vella, L. J., Sharples, R. A., Nisbet, R. M., Cappai, R., and Hill, A. F. (2008). The role of exosomes in the processing of proteins associated with neurodegenerative diseases. Eur. Biophys. J. 37, 323–332.
Viaud, S., Ploix, S., Lapierre, V., Théry, C., Commere, P.-H., Tramalloni, D., Gorrichon, K., Virault-Rocroy, P., Tursz, T., Lantz, O., Zitvogel, L., and Chaput, N. (2011). Updated technology to produce highly immunogenic dendritic cell-derived exosomes of clinical grade: a critical role of interferon-γ. J. Immunother. 34, 65–75.
Viaud, S., Terme, M., Flament, C., Taïeb, J., André, F., Novault, S., Escudier, B., Robert, C., Caillat-Zucman, S., Tursz, T., Zitvogel, L., and Chaput, N. (2009). Dendritic cell-derived exosomes promote natural killer cell activation and proliferation: a role for NKG2D ligands and IL-15Ralpha. PLoS ONE 4, e4942. doi:10.1371/journal.pone.0004942
Vidal, M., Sainte-Marie, J., Philippot, J. R., and Bienvenue, A. (1989). Asymmetric distribution of phospholipids in the membrane of vesicles released during in vitro maturation of guinea pig reticulocytes: evidence precluding a role for “aminophospholipid translocase.” J. Cell. Physiol. 140, 455–462.
Vingtdeux, C., Buée, L., and Sergeant, N. (2007). “Contribution of multivesicular bodies of the prion-like propagation of lesions in Alzheimer’s disease,” in Alzheimer’s Disease Pathogenesis – Core Concepts, Shifting Paradigms and Therapeutic Targets, Chap. 6, ed. S. De La Monte (Rijeka: InTech), 107–130.
Waldenström, A., Gennebäck, N., Hellman, U., and Ronquist, G. (2012). Cardiomyocyte microvesicles contain DNA/RNA and convey biological messages to target cells. PLoS ONE 7, e34653. doi:10.1371/journal.pone.0034653
Wang, S., Cesca, F., Loers, G., Schweizer, M., Buck, F., Benfenati, F., Schachner, M., and Kleene, R. (2011). Synapsin I is an oligomannose-carrying glycoprotein, acts as an oligomannose-binding lectin, and promotes neurite outgrowth and neuronal survival when released via glia-derived exosomes. J. Neurosci. 31, 7275–7290.
Whitehead, K. A., Dahlman, J., Langer, R. S., and Anderson, D. G. (2011). Silencing or stimulation? siRNA delivery and the immune system. Ann. Rev. Chem. Biomol. Eng. 2, 77–96.
Wolfers, J., Lozier, A., Raposo, G., Regnault, A., Théry, C., Masurier, C., Flament, C., Pouzieux, S., Faure, F., Tursz, T., Angevin, E., Amigorena, S., and Zitvogel, L. (2001). Tumor-derived exosomes are a source of shared tumor rejection antigens for CTL cross-priming. Nat. Med. 7, 297–303.
Wurdinger, T., Gatson, N. A., Balaj, L., Kaur, B., Breakefield, X. O., and Pegtel, D. M. (2012). Extracellular vesicles and their convergence with viral pathways. Adv. Virol. (in press).
Würdinger, T., Tannous, B. A., Saydam, O., Skog, J., Grau, S., Soutschek, J., Weissleder, R., Breakefield, X. O., and Krichevsky, A. M. (2008). miR-296 regulates growth factor receptor overexpression in angiogenic endothelial cells. Cancer Cell 14, 382–393.
Xie, Y., Bai, O., Zhang, H., Yuan, J., Zong, S., Chibbar, R., Slattery, K., Qureshi, M., Wei, Y., Deng, Y., and Xiang, J. (2010). Membrane-bound HSP70-engineered myeloma cell-derived exosomes stimulate more efficient CD8+ CTL- and NK-mediated antitumor immunity than exosomes released from heat-shocked tumor cells expressing cytoplasmic HSP70. J. Cell. Mol. Med. 14, 2655–2666.
Xiu, F., Cai, Z., Yang, Y., Wang, X., Wang, J., and Cao, X. (2007). Surface anchorage of superantigen SEA promotes induction of specific antitumor immune response by tumor-derived exosomes. J. Mol. Med. 85, 511–521.
Yang, M., Chen, J., Su, F., Yu, B., Su, F., Lin, L., Liu, Y., Huang, J. D., and Song, E. (2011). Microvesicles secreted by macrophages shuttle invasion-potentiating microRNAs into breast cancer cells. Mol. Cancer 10, 117.
Yang, Y., Xiu, F., Cai, Z., Wang, J., Wang, Q., Fu, Y., and Cao, X. (2007). Increased induction of antitumor response by exosomes derived from interleukin-2 gene-modified tumor cells. J. Cancer Res. Clin. Oncol. 133, 389–399.
Yu, X., Harris, S. L., and Levine, A. J. (2006). The regulation of exosome secretion: a novel function of the p53 protein. Cancer Res. 66, 4795–4801.
Yuyama, K., Sun, H., Mitsutake, S., and Igarashi, Y. (2012). Sphingolipid-modulated exosome secretion promotes the clearance of amyloid-β by microglia. J. Biol. Chem. 287, 10977–10989.
Zeelenberg, I. S., van Maren, W. W. C., Boissonnas, A., Van Hout-Kuijer, M. A., Brok, Den, M. H. M. G. M., Wagenaars, J. A. L., van der Schaaf, A., Jansen, E. J. R., Amigorena, S., Théry, C., Figdor, C. G., and Adema, G. J. (2011). Antigen localization controls T cell-mediated tumor immunity. J. Immunol. 187, 1281–1288.
Zhang, B., Une, Y., Fu, X., Yan, J., Ge, F., Yao, J., Sawashita, J., Mori, M., Tomozawa, H., Kametani, F., and Higuchi, K. (2008). Fecal transmission of AA amyloidosis in the cheetah contributes to high incidence of disease. Proc. Natl. Acad. Sci. U.S.A. 105, 7263–7268.
Zhang, H.-G., Kim, H., Liu, C., Yu, S., Wang, J., Grizzle, W. E., Kimberly, R. P., and Barnes, S. (2007). Curcumin reverses breast tumor exosomes mediated immune suppression of NK cell tumor cytotoxicity. Biochim. Biophys. Acta 1773, 1116–1123.
Zhang, Y., Liu, D., Chen, X., Li, J., Li, L., Bian, Z., Sun, F., Lu, J., Yin, Y., Cai, X., Sun, Q., Wang, K., Ba, Y., Wang, Q., Wang, D., Yang, J., Liu, P., Xu, T., Yan, Q., Zhang, J., Zen, K., and Zhang, C. Y. (2010). Secreted monocytic miR-150 enhances targeted endothelial cell migration. Mol. Cell 39, 133–144.
Zhuang, X., Xiang, X., Grizzle, W., Sun, D., Zhang, S., Axtell, R. C., Ju, S., Mu, J., Zhang, L., Steinman, L., Miller, D., and Zhang, H. G. (2011). Treatment of brain inflammatory diseases by delivering exosome encapsulated anti-inflammatory drugs from the nasal region to the brain. Mol. Ther. 19, 1769–1779.
Zitvogel, L., Regnault, A., Lozier, A., Wolfers, J., Flament, C., Tenza, D., Ricciardi-Castagnoli, P., Raposo, G., and Amigorena, S. (1998). Eradication of established murine tumors using a novel cell-free vaccine: dendritic cell-derived exosomes. Nat. Med. 4, 594–600.
Zomer, A., Vendrig, T., Hopmans, E. S., van Eijndhoven, M., Middeldorp, J. M., and Pegtel, D. M. (2010). Exosomes: fit to deliver small RNA. Commun. Integr. Biol. 3, 447–450.
Zwaal, R. F., Comfurius, P., and Bevers, E. M. (1992). Platelet procoagulant activity and microvesicle formation. Its putative role in hemostasis and thrombosis. Biochim Biophys Acta 1180, 1–8.
Keywords: microvesicles, exosomes, neuron, neuroregeneration, neurodegeneration, development, cancer, therapy
Citation: Lai CP-K and Breakefield XO (2012) Role of exosomes/microvesicles in the nervous system and use in emerging therapies. Front. Physio. 3:228. doi: 10.3389/fphys.2012.00228
Received: 21 March 2012; Paper pending published: 11 April 2012;
Accepted: 06 June 2012; Published online: 27 June 2012.
Edited by:
Claudia Verderio, CNR Institute of Neuroscience, ItalyReviewed by:
Patrizia Rosa, Italian National Research Council, ItalyRachel Susan Kraut, Nanyang Technological University, Singapore
Stefano Pluchino, University of Cambridge, UK
Mikael Simons, Max-Planck-Institut für Experimentelle Medizin, Germany
Copyright: © 2012 Lai and Breakefield. This is an open-access article distributed under the terms of the Creative Commons Attribution Non Commercial License, which permits non-commercial use, distribution, and reproduction in other forums, provided the original authors and source are credited.
*Correspondence: Xandra Owen Breakefield, Molecular Neurogenetics Unit, Massachusetts General Hospital-East, 13th Street, Building 149, Charlestown, MA 02129, USA. e-mail: breakefield@hms.harvard.edu