
Multidrug Efflux Pumps from Enterobacteriaceae, Vibrio cholerae and Staphylococcus aureus Bacterial Food Pathogens


Abstract
:1. Introduction
1.1. Food Microbiology: Production and Spoilage
1.1.1. Food Production
1.1.2. Food Spoilage
1.2. Bacterial Food Pathogens
Key Bacterial Pathogens, Virulence and Available Chemotherapy
Salmonella enterica
Staphylococcus aureus
Escherichia coli
Vibrio cholerae
Enterobacter spp.
1.3. Problem of Bacterial Resistance to Antimicrobial Agents
2. Mechanisms of Bacterial Drug Resistance
2.1. Biofilms

2.2. Drug Target Modification
2.3. Enzymatic Degradation of Antimicrobial Agents
2.4. Reduced Drug Permeability across the Bacterial Membrane
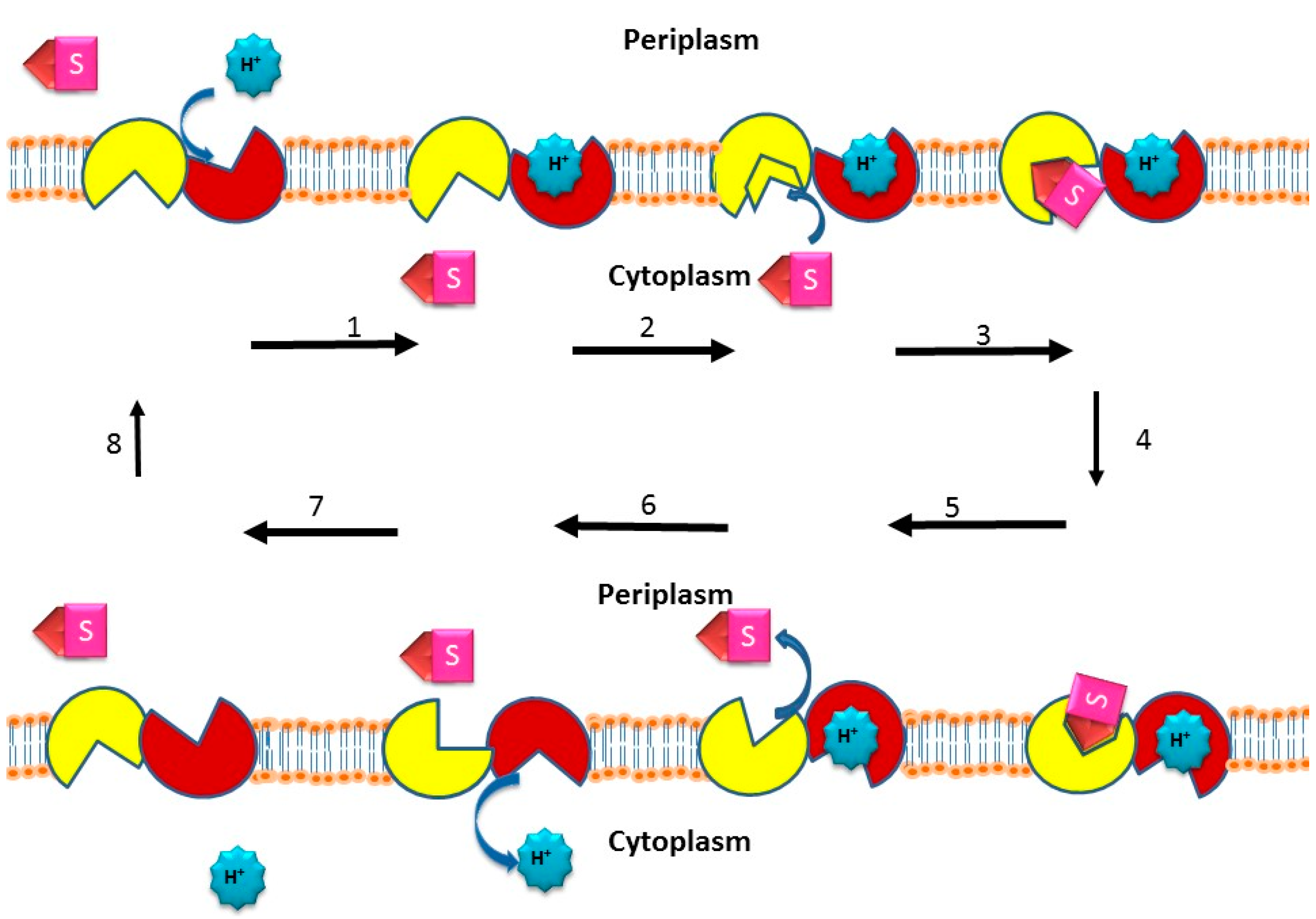
2.5. Active Efflux of Drugs from Bacterial Pathogens

2.5.1. Discovery of Single and Multidrug Efflux Pumps
2.5.2. Efflux Pumps of Staphylococcus aureus
QacA and QacB Multidrug Efflux Pumps
LmrS Multidrug Efflux Pump
MdeA Multidrug Efflux Pump
QacG, QacH, and QacJ Multidrug Efflux Pumps
NorA Multidrug Efflux Pump
NorB Multidrug Efflux Pump
NorC Multidrug Efflux Pump
NorD Multidrug Efflux Pump
TetA(K) Tetracycline Efflux Pump
Tet38 Tetracycline Efflux Pump
SdrM Multidrug Efflux Pump
Mef(A) Multidrug Efflux Pump
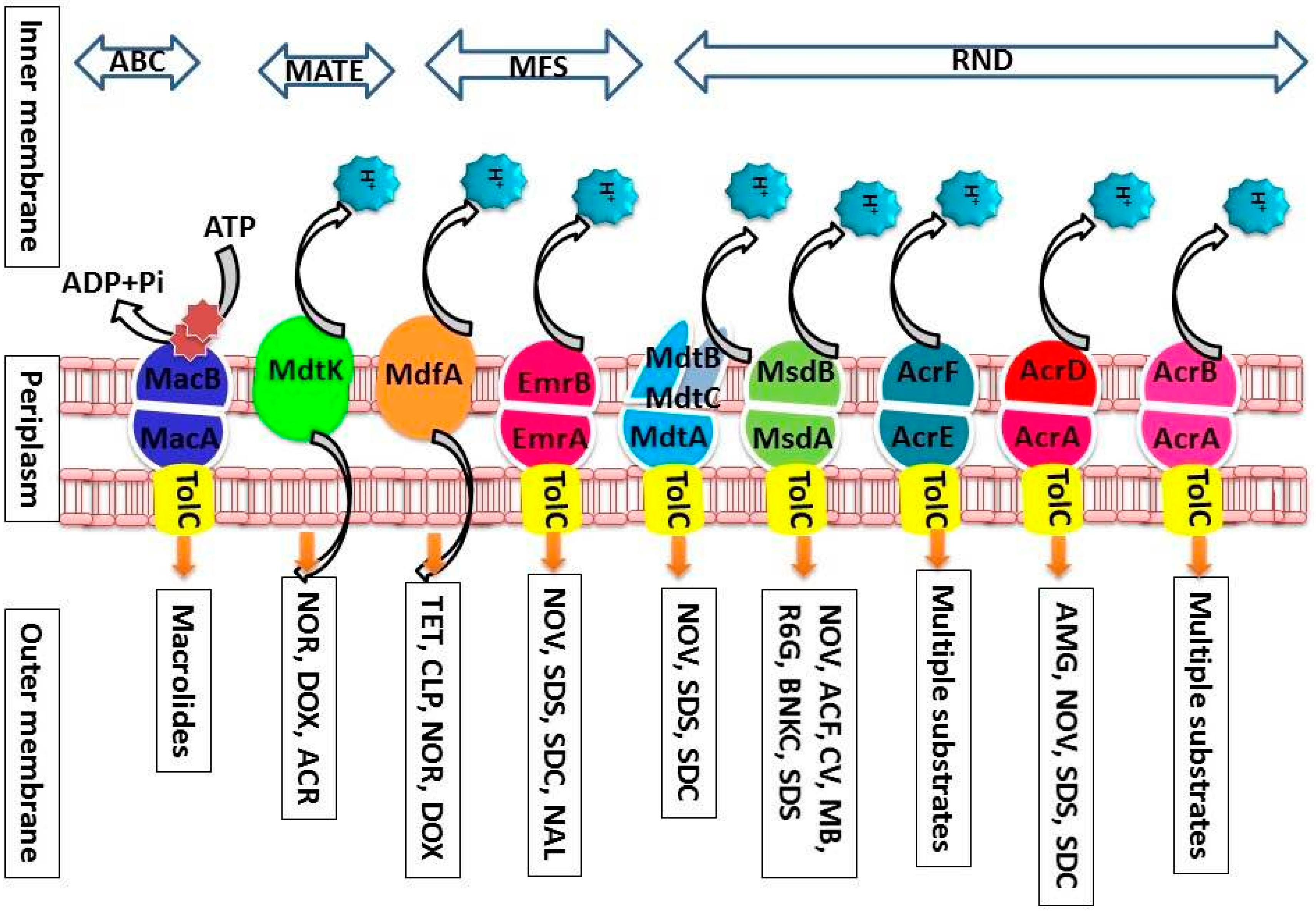
2.5.3. Efflux Pumps from Escherichia coli
The AcrA, AcrB, and TolC Multidrug Efflux Pump
EmrE and SugE Multidrug Efflux Pumps
MdfA Multidrug Efflux Pump
MacA and MacB Multidrug Efflux Pumps
2.5.4. Efflux Pumps from Vibrio cholerae
EmrD-3 Multidrug Efflux Pump
VceB Multidrug Efflux Pump
VcrM Multidrug Efflux Pump
2.5.5. Efflux Pumps from Salmonella enterica


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Pumps | Description | Database |
|---|---|---|---|
| tr|Q8Z810 | YCAD | Uncharacterized MFS-type transporter YcaD | UP |
| tr|P58530 | SOTB | Probable sugar efflux transporter | UP |
| tr|Q8XFG0 | MDTM | Multidrug resistance protein MdtM | UP |
| tr|P37594 | SMVA | Methyl viologen resistance protein SmvA | UP |
| tr|Q8Z257 | YHHS | UPF0226 protein YhhS | UP |
| tr|Q8Z4Z9 | YFCJ | UPF0226 protein YfcJ | UP |
| tr|Q8XGS2 | NEPI | Purine ribonucleoside efflux pump NepI | UP |
| tr|Q8Z7L0 | MDTH | Multidrug resistance protein MdtH | UP |
| tr|P27669 | UHPC | Regulatory protein UhpC | UP |
| tr|P33027 | SETB | Sugar efflux transporter B | UP |
| tr|Q8Z6Q5 | DTPA | Dipeptide and tripeptide permease A | UP |
| tr|P37593 | NARU | Nitrate/nitrite transporter NarU | UP |
| tr|P0A2G3 | CITA | Citrate-proton symporter | UP |
| tr|P40862 | PROP | Proline/betaine transporter | UP |
| tr|C0Q135 | Y3650 | UPF0226 membrane protein SPC_3650 | UP |
| tr|P33733 | TCR4 | Tetracycline resistance protein, class D | UP |
| tr|C0PZN3 | NANT | Putative sialic acid transporter | UP |
| tr|B5R3Y2 | Y3404 | UPF0226 membrane protein SEN3404 | UP |
| tr|Q8ZLE4 | YHHS | UPF0226 protein YhhS | UP |
| tr|B4SZP4 | Y2561 | UPF0226 membrane protein SNSL254_A2561 | UP |
| tr|Q5PLW2 | TSGA | Protein TsgA | UP |
| tr|B4T502 | LPLT | Lysophospholipid transporter LplT | UP |
| tr|Q7CQY0 | DTPD | Dipeptide permease D | UP |
| tr|Q8ZLD6 | DTPB | Dipeptide and tripeptide permease B | UP |
| tr|Q57LY2 | Y2374 | UPF0226 membrane protein SCH_2374 | UP |
| tr|Q57RY1 | ENTS | Enterobactin exporter EntS | UP |
| tr|Q9L7R5 | YIHP | Putative 2,3-dihydroxypropane-1-sulfonate exporter | UP |
| tr|Q9L7R4 | YIHO | Putative sulfoquinovose importer | UP |
| tr|A9N7L0 | MDTD | Putative multidrug resistance protein MdtD | UP |
| tr|B5F0Y5 | MdtM | Multidrug resistance protein MdtM | UP |
| tr|D0ZS93 | Multidrug translocase | Multidrug translocase | UP |
| tr|G4BZY2 | Transporter | Major facilitator family protein | UP |
| tr|E8ZZM6 | MdtM | Multidrug resistance protein MdtM | UP |
| tr|I0A7E0 | MdfA | Multidrug translocase MdfA | UP |
| tr|S5HWQ9 | Transporter | Multidrug transporter | UP |
| B7ZJI2 | TetA | SGI encoded Tetracycline efflux pump | UP |
| Q9K2Y4 | TetB | SGI encoded Tetracycline efflux pump | UP |
| B0JYJ4 | TetG | SGI encoded Tetracycline efflux pumps | UP |
| gi|575533051 | CmR | Chloramphenicol resistance pump Cmr | NB |
| WP_0029541 | KDG permease | 2-keto-3-deoxygluconate permease | NB |
| gb|ESG911| | Putative pump | Inner membrane protein | NB |
| YP_0021121 | SeSA_B0056 | Hypothetical protein | NB |
| YP2091 | Macrolide-efflux determinant | Macrolide-efflux determinant | NB |
| WP_0204371 | Putative permease | Putative permease | NB |
| WP_0232341 | MdtH | Multidrug resistance protein | NB |
| YP_0052381 | EmrAB | emrA/ emrB/ tolC efflux system | NB |
MFS Efflux Pumps of S. enterica
EmrAB of S. enterica
MdfA of S. enterica
The Efflux Pumps of the Salmonella Genomic Island I (SGI-1)
TetA/B/G of SGI-1
2.5.6. Efflux Pumps from Enterobacter spp.
EmmdR Multidrug Efflux Pump
The AcrAB-TolC Efflux Pump System
Outer Membrane Proteins OmpD and OmpF
SugE Multidrug Efflux Pump
3. Evolution of Drug and Multidrug Efflux Pumps
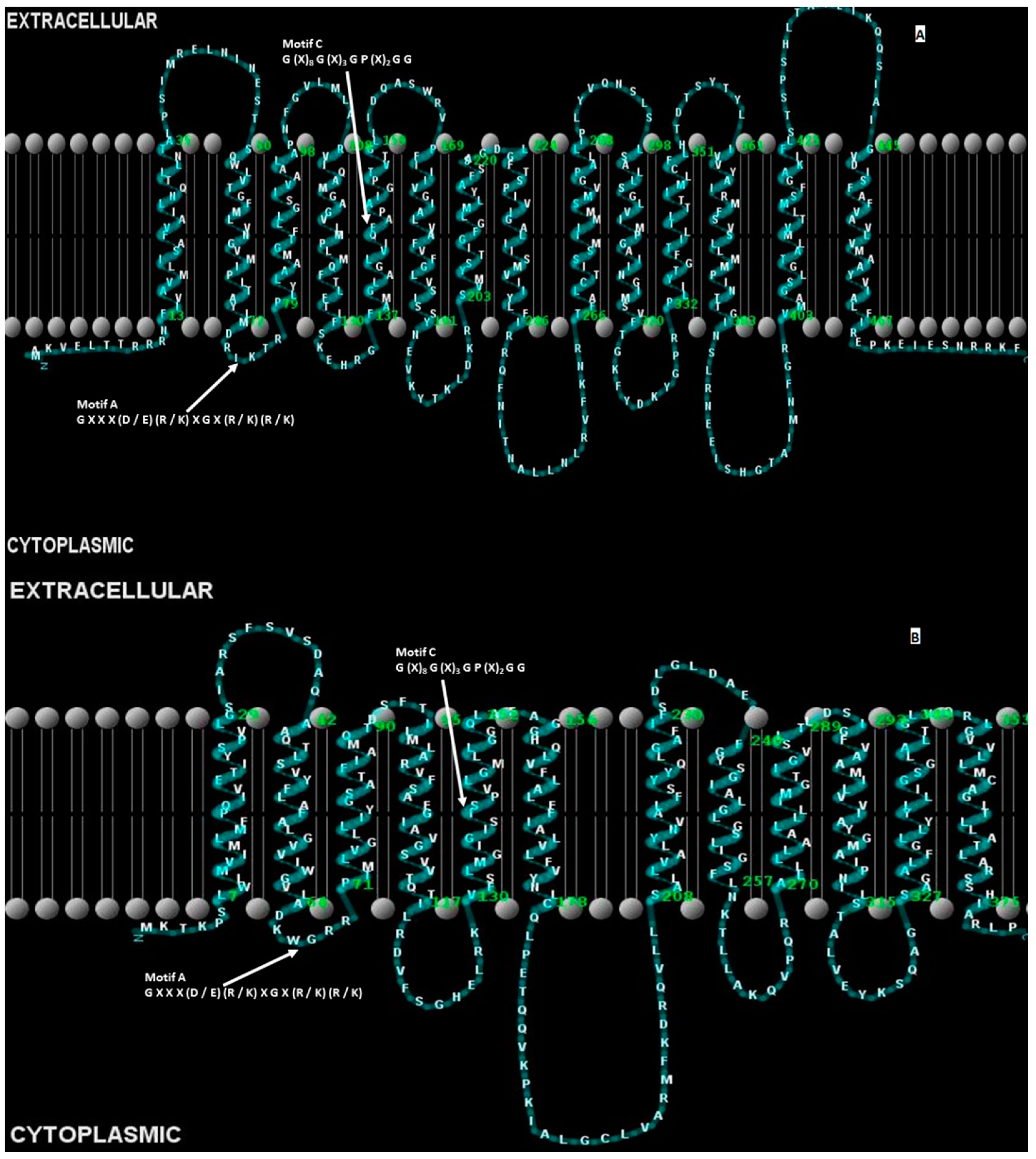
3.1. Conservation of Amino Acid Sequence Motifs

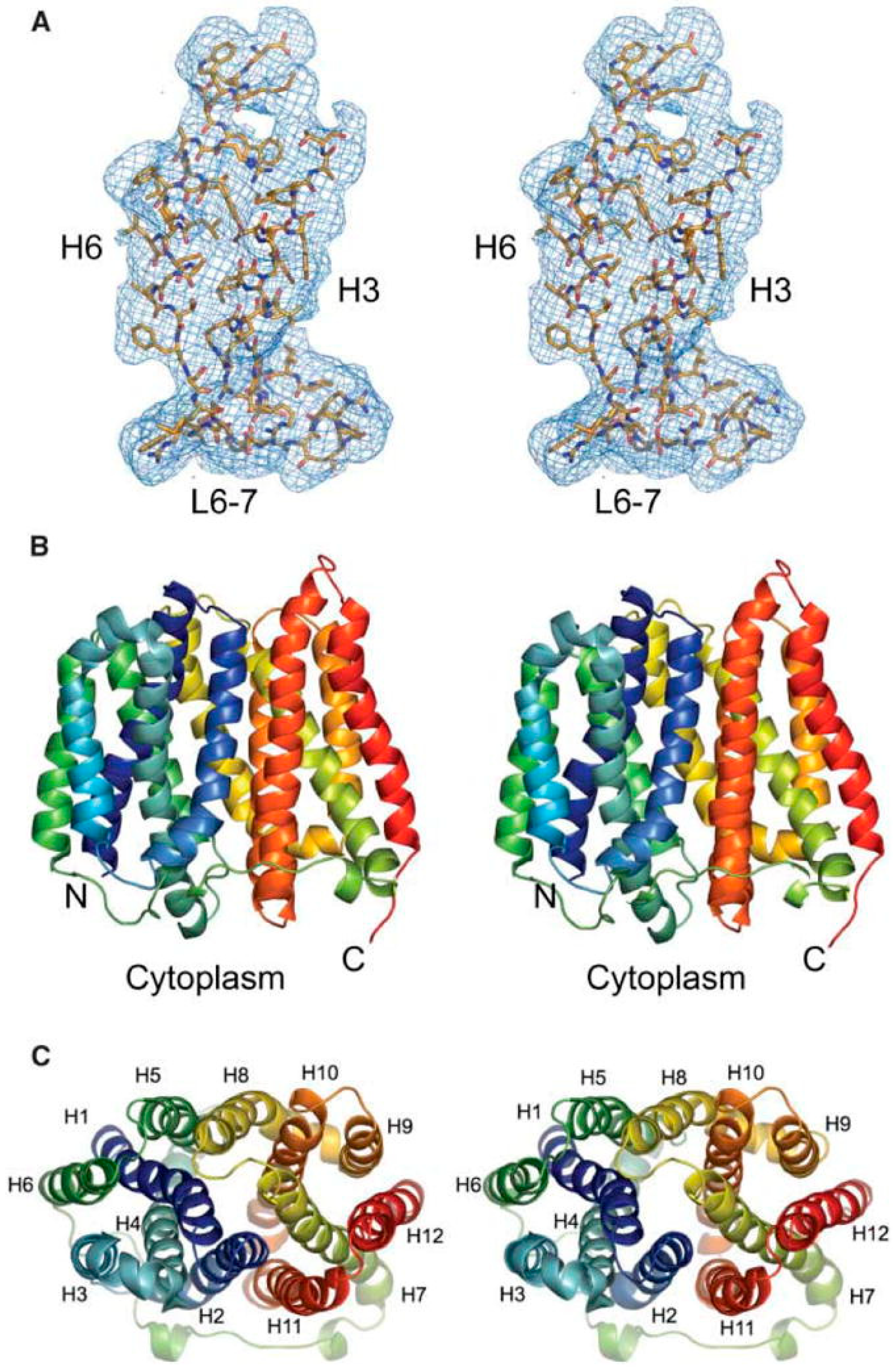
3.2. Conservation of 3-Dimensional Structures

4. Control of Drug Efflux
4.1. Control at the Level of Gene Expression
4.2. Effect on Virulence
4.3. Control at the Level of Efflux Pump Activity
5. Future Directions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Centers for Disease Control and Prevention. Incidence and trends of infection with pathogens transmitted commonly through food—Foodborne diseases active surveillance network, 10 U.S. sites, 1996–2012. MMWR 2013, 62, 283–287. [Google Scholar]
- Kuchenmuller, T.; Hird, S.; Stein, C.; Kramarz, P.; Nanda, A.; Havelaar, A.H. Estimating the global burden of foodborne diseases—A collaborative effort. Eur. Commun. Dis. Bull. 2009, 14. [Google Scholar] [CrossRef] [Green Version]
- Nyenje, M.E.; Odjadjare, C.E.; Tanih, N.F.; Green, E.; Ndip, R.N. Foodborne pathogens recovered from ready-to-eat foods from roadside cafeterias and retail outlets in Alice, Eastern Cape Province, South Africa: Public health implications. Int. J. Environ. Res. Public Health 2012, 9, 2608–2619. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, M.; Wahi, S.; Kumar, M.; Ganguli, A. Prevalence of enterotoxigenic Staphylococcus aureus and Shigella spp. in some raw street vended Indian foods. Int. J. Environ. Res. Public Health 2007, 17, 151–156. [Google Scholar] [CrossRef]
- Teplitski, M.; Wright, A.C.; Lorca, G. Biological approaches for controlling shellfish-associated pathogens. Curr. Opin. Biotechnol. 2009, 20, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Newell, D.G.; Koopmans, M.; Verhoef, L.; Duizer, E.; Aidara-Kane, A.; Sprong, H.; Opsteegh, M.; Langelaar, M.; Threfall, J.; Scheutz, F.; et al. Food-borne diseases—The challenges of 20 years ago still persist while new ones continue to emerge. Int. J. Food Microbiol. 2010, 139, S3–S15. [Google Scholar] [CrossRef] [PubMed]
- Gram, L.; Dalgaard, P. Fish spoilage bacteria—Problems and solutions. Curr. Opin. Biotechnol. 2002, 13, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Gram, L.; Melchiorsen, J. Interaction between fish spoilage bacteria Pseudomonas sp. and Shewanella putrefaciens in fish extracts and on fish tissue. J. Appl. Bacteriol. 1996, 80, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States—Major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, J.A.; Steele-Mortimer, O. Salmonella—The ultimate insider. Salmonella virulence factors that modulate intracellular survival. Cell. Microbiol. 2009, 11, 1579–1586. [Google Scholar]
- Rensch, U.; Klein, G.; Kehrenberg, C. Analysis of triclosan-selected Salmonella enterica mutants of eight serovars revealed increased aminoglycoside susceptibility and reduced growth rates. PLoS One 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Quinn, T.; O’Mahony, R.; Baird, A.W.; Drudy, D.; Whyte, P.; Fanning, S. Multi-drug resistance in Salmonella enterica: Efflux mechanisms and their relationships with the development of chromosomal resistance gene clusters. Curr. Drug Targets 2006, 7, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Nishino, K.; Nikaido, E.; Yamaguchi, A. Regulation and physiological function of multidrug efflux pumps in Escherichia coli and Salmonella. Biochimica et Biophysica Acta 2009, 1794, 834–843. [Google Scholar] [CrossRef] [PubMed]
- Fookes, M.; Schroeder, G.N.; Langridge, G.C.; Blondel, C.J.; Mammina, C.; Seth-Smith, H.; Connor, T.R.; Vernikos, G.S.; Robinson, K.S.; Sanders, M.; et al. Salmonella bongori provides insights into the evolution of the Salmonellae. PLoS Pathog. 2011, 7. [Google Scholar] [CrossRef] [Green Version]
- Wistrom, J.; Norrby, S.R. Fluoroquinolones and bacterial enteritis, when and for whom? J. Antimicrob. Chemother. 1995, 36, 23–39. [Google Scholar] [CrossRef] [PubMed]
- Waddington, C.S.; Darton, T.C.; Pollard, A.J. The challenge of enteric fever. J. Infect. 2014, 68, S38–S50. [Google Scholar] [CrossRef] [PubMed]
- Fabrega, A.; Vila, J. Salmonella enterica serovar Typhimurium skills to succeed in the host: Virulence and regulation. Clin. Microbiol. Rev. 2013, 26, 308–341. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Basis of virulence in community-associated methicillin-resistant Staphylococcus aureus. Ann. Rev. Microbiol. 2010, 64, 143–162. [Google Scholar] [CrossRef]
- Murray, P.R.; Rosenthal, K.S.; Pfaller, P.F. Medical Microbiology; Mosby Elsevier: Philadelphia, PA, USA, 2013. [Google Scholar]
- Foster, T.J. Colonization and infection of the human host by staphylococci: Adhesion, survival and immune evasion. Vet. Dermatol. 2009, 20, 456–470. [Google Scholar] [CrossRef] [PubMed]
- Manges, A.R.; Johnson, J.R. Food-borne origins of Escherichia coli causing extraintestinal infections. Clin. Infect. Dis. 2012, 55, 712–719. [Google Scholar] [CrossRef] [PubMed]
- Arthur, J.C.; Perez-Chanona, E.; Muhlbauer, M.; Tomkovich, S.; Uronis, J.M.; Fan, T.J.; Campbell, B.J.; Abujamel, T.; Dogan, B.; Rogers, A.B.; et al. Intestinal inflammation targets cancer-inducing activity of the microbiota. Science 2012, 338, 120–123. [Google Scholar] [CrossRef] [PubMed]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Porter, C.K.; Riddle, M.S.; Tribble, D.R.; Louis Bougeois, A.; McKenzie, R.; Isidean, S.D.; Sebeny, P.; Savarino, S.J. A systematic review of experimental infections with enterotoxigenic Escherichia coli (ETEC). Vaccine 2011, 29, 5869–5885. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.D.; Frankel, G. Enteropathogenic Escherichia coli: Unravelling pathogenesis. FEMS Microbiol. Rev. 2005, 29, 83–98. [Google Scholar] [CrossRef] [PubMed]
- Clarke, S.C.; Haigh, R.D.; Freestone, P.P.; Williams, P.H. Enteropathogenic Escherichia coli infection: history and clinical aspects. Br. J. Biomed. Sci. 2002, 59, 123–127. [Google Scholar] [PubMed]
- Ochoa, T.J.; Contreras, C.A. Enteropathogenic Escherichia coli infection in children. Curr. Opin. Infect. Dis. 2011, 24, 478–483. [Google Scholar] [CrossRef] [PubMed]
- Campos, L.C.; Franzolin, M.R.; Trabulsi, L.R. Diarrheagenic Escherichia coli categories among the traditional enteropathogenic E. coli O serogroups—A review. Memorias do Instituto Oswaldo Cruz 2004, 99, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Pickering, L.K. Gastroenteritis due to enteropathogenic, enterotoxigenic, and invasive Escherichia coli: A review. Am. J. Med. Technol. 1979, 45, 787–792. [Google Scholar] [PubMed]
- Welinder-Olsson, C.; Kaijser, B. Enterohemorrhagic Escherichia coli (EHEC). Scand. J. Infect. Dis. 2005, 37, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Fratamico, P.M.; Gunther, N.W.T. Shiga toxin-producing Escherichia coli. Adv. Appl. Microbiol. 2014, 86, 145–197. [Google Scholar] [PubMed]
- Erickson, M.C.; Doyle, M.P. Food as a vehicle for transmission of Shiga toxin-producing Escherichia coli. J. Food Prot. 2007, 70, 2426–2449. [Google Scholar] [PubMed]
- Bettelheim, K.A. The non-O157 shiga-toxigenic (verocytotoxigenic) Escherichia coli; under-rated pathogens. Crit. Rev. Microbiol. 2007, 33, 67–87. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Yang, Q.; Kase, J.A.; Meng, J.; Clotilde, L.M.; Lin, A.; Ge, B. Current trends in detecting non-O157 Shiga toxin-producing Escherichia coli in food. Foodborne Pathog. Dis. 2013, 10, 665–677. [Google Scholar] [CrossRef] [PubMed]
- Van den Beld, M.J.; Reubsaet, F.A. Differentiation between Shigella, enteroinvasive Escherichia coli (EIEC) and noninvasive Escherichia coli. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 899–904. [Google Scholar]
- Martinez-Medina, M.; Garcia-Gil, L.J. Escherichia coli in chronic inflammatory bowel diseases: An update on adherent invasive Escherichia coli pathogenicity. World J. Gastrointest. Pathophysiol. 2014, 5, 213–227. [Google Scholar] [PubMed]
- Parsot, C. Shigella spp. and enteroinvasive Escherichia coli pathogenicity factors. FEMS Microbiol. Lett. 2005, 252, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Faruque, S.M.; Albert, M.J.; Mekalanos, J.J. Epidemiology, genetics, and ecology of toxigenic Vibrio cholerae. Microbiol. Mol. Biol. Rev. 1998, 62, 1301–1314. [Google Scholar] [PubMed]
- Charles, R.C.; Ryan, E.T. Cholera in the 21st century. Curr. Opin. Infect. Dis. 2011, 24, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Ghose, A.C. Lessons from cholera & Vibrio cholerae. Indian J. Med. Res. 2011, 133, 164–170. [Google Scholar] [PubMed]
- Neilson, A.A.; Mayer, C.A. Cholera—Recommendations for prevention in travellers. Aust. Fam. Phys. 2010, 39, 220–226. [Google Scholar]
- Ghosh, A.; Ramamurthy, T. Antimicrobials & cholera: Are we stranded? Indian J. Med. Res. 2011, 133, 225–231. [Google Scholar] [PubMed]
- Sanders, W.E., Jr.; Sanders, C.C. Enterobacter spp.: Pathogens poised to flourish at the turn of the century. Clin. Microbiol. Rev. 1997, 10, 220–241. [Google Scholar] [PubMed]
- Bouza, E.; Cercenado, E. Klebsiella and Enterobacter: Antibiotic resistance and treatment implications. Semin. Respir. Infect. 2002, 17, 215–230. [Google Scholar] [CrossRef] [PubMed]
- Diene, S.M.; Merhej, V.; Henry, M.; el Filali, A.; Roux, V.; Robert, C.; Azza, S.; Gavory, F.; Barbe, V.; la Scola, B.; et al. The rhizome of the multidrug-resistant Enterobacter aerogenes genome reveals how new “killer bugs” are created because of a sympatric lifestyle. Mol. Biol. Evolut. 2013, 30, 369–383. [Google Scholar] [CrossRef]
- Cabral, J.P. Water microbiology. Bacterial pathogens and water. Int. J. Environ. Res. Public Health 2010, 7, 3657–3703. [Google Scholar] [CrossRef] [PubMed]
- He, G.X.; Thorpe, C.; Walsh, D.; Crow, R.; Chen, H.; Kumar, S.; Varela, M.F. EmmdR, a new member of the MATE family of multidrug transporters, extrudes quinolones from Enterobacter cloacae. Arch. Microbiol. 2011, 193, 759–765. [Google Scholar] [CrossRef] [PubMed]
- He, G.X.; Zhang, C.; Crow, R.R.; Thorpe, C.; Chen, H.; Kumar, S.; Tsuchiya, T.; Varela, M.F. SugE, a new member of the SMR family of transporters, contributes to antimicrobial resistance in Enterobacter cloacae. Antimicrob. Agents Chemother. 2011, 55, 3954–3957. [Google Scholar] [CrossRef] [PubMed]
- Levy, S.B. The future of antibiotics: Facing antibiotic resistance. Clin. Microbiol. Infect. 2000, 6, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Higgins, C.F. Multiple molecular mechanisms for multidrug resistance transporters. Nature 2007, 446, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Varela, M.F. Biochemistry of bacterial multidrug efflux pumps. Int. J. Mol. Sci. 2012, 13, 4484–4495. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Varela, M.F. Molecular mechanisms of bacterial resistance to antimicrobial agents. In Microbial Pathogens and Strategies for Combating Them: Science, Technology and Education; Méndez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2013; pp. 522–534. [Google Scholar]
- Alekshun, M.N.; Levy, S.B. Molecular mechanisms of antibacterial multidrug resistance. Cell 2007, 128, 1037–1050. [Google Scholar] [CrossRef] [PubMed]
- Pechere, J.C.; Hughes, D.; Kardas, P.; Cornaglia, G. Non-compliance with antibiotic therapy for acute community infections: A global survey. Int. J. Antimicrob. Agents 2007, 29, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C. Molecular mechanisms that confer antibacterial drug resistance. Nature 2000, 406, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Hoiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents 2010, 35, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Anderl, J.N.; Franklin, M.J.; Stewart, P.S. Role of antibiotic penetration limitation in Klebsiella pneumoniae biofilm resistance to ampicillin and ciprofloxacin. Antimicrob. Agents Chemother. 2000, 44, 1818–1824. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.R.; Allison, D.G.; Gilbert, P. Resistance of bacterial biofilms to antibiotics: A growth-rate related effect? J. Antimicrob. Chemother. 1988, 22, 777–780. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed]
- Soto, S.M. Role of efflux pumps in the antibiotic resistance of bacteria embedded in a biofilm. Virulence 2013, 4, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K. Persister cells and the riddle of biofilm survival. Biochem. Biokhimiia 2005, 70, 267–274. [Google Scholar] [CrossRef]
- Ammons, M.C. Anti-biofilm strategies and the need for innovations in wound care. Recent Pat. Anti-Infect. Drug Discov. 2010, 5, 10–17. [Google Scholar] [CrossRef]
- Fabrega, A.; Madurga, S.; Giralt, E.; Vila, J. Mechanism of action of and resistance to quinolones. Microb. Biotechnol. 2009, 2, 40–61. [Google Scholar] [CrossRef] [PubMed]
- Lambert, P.A. Bacterial resistance to antibiotics: modified target sites. Adv. Drug Deliv. Rev. 2005, 57, 1471–1485. [Google Scholar] [CrossRef] [PubMed]
- Poehlsgaard, J.; Douthwaite, S. The bacterial ribosome as a target for antibiotics. Nat. Rev. Microbiol. 2005, 3, 870–881. [Google Scholar] [CrossRef] [PubMed]
- Schnappinger, D.; Hillen, W. Tetracyclines: Antibiotic action, uptake, and resistance mechanisms. Arch. Microbiol. 1996, 165, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, G.A. Mechanisms of resistance to quinolones. Clin. Infect. Dis. 2005, 41, S120–S126. [Google Scholar] [CrossRef] [PubMed]
- Gutkind, G.O.; di Conza, J.; Power, P.; Radice, M. β-lactamase-mediated resistance: A biochemical, epidemiological and genetic overview. Curr. Pharm. Design 2013, 19, 164–208. [Google Scholar] [CrossRef]
- De Pascale, G.; Wright, G.D. Antibiotic resistance by enzyme inactivation: From mechanisms to solutions. Chembiochem 2010, 11, 1325–1334. [Google Scholar]
- Llano-Sotelo, B.; Azucena, E.F., Jr.; Kotra, L.P.; Mobashery, S.; Chow, C.S. Aminoglycosides modified by resistance enzymes display diminished binding to the bacterial ribosomal aminoacyl-tRNA site. Chem. Biol. 2002, 9, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Kirst, H.A. New macrolide, lincosaminide and streptogramin B antibiotics. Expert Opin. Ther. Pat. 2010, 20, 1343–1357. [Google Scholar] [CrossRef] [PubMed]
- Baysarowich, J.; Koteva, K.; Hughes, D.W.; Ejim, L.; Griffiths, E.; Zhang, K.; Wright, G.D.; Junop, M. Rifamycin antibiotic resistance by ADP-ribosylation: Structure and diversity of Arr. Proc. Natl. Acad. Sci. USA 2008, 105, 4886–4891. [Google Scholar] [CrossRef] [PubMed]
- Kren, V.; Martinkova, L. Glycosides in medicine: The role of glycosidic residue in biological activity. Curr. Med. Chem. 2001, 8, 1303–1328. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Schweizer, H.P. Bacterial resistance to antibiotics: Active efflux and reduced uptake. Adv. Drug Deliv. Rev. 2005, 57, 1486–1513. [Google Scholar] [CrossRef] [PubMed]
- Gunn, J.S.; Ernst, R.K. The structure and function of Francisella lipopolysaccharide. Ann. N. Y. Acad. Sci. 2007, 1105, 202–218. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef] [PubMed]
- Pages, J.M.; James, C.E.; Winterhalter, M. The porin and the permeating antibiotic: A selective diffusion barrier in Gram-negative bacteria. Nat. Rev. Microbiol. 2008, 6, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Achouak, W.; Heulin, T.; Pages, J.M. Multiple facets of bacterial porins. FEMS Microbiol. Lett. 2001, 199, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Page, M.G. The role of the outer membrane of Gram-negative bacteria in antibiotic resistance: Ajax’ shield or Achilles’ heel? Handb. Exp. Pharmacol. 2012, 211, 67–86. [Google Scholar] [PubMed]
- Kumar, S.; Floyd, J.T.; He, G.; Varela, M.F. Bacterial antimicrobial efflux pumps of the MFS and MATE transporter families: A review. In Recent Research Developments in Antimicrobial Agents & Chemotherapy; Research Signpost, Inc.: Kerala, India, 2013; pp. 1–21. [Google Scholar]
- Yaffe, D.; Radestock, S.; Shuster, Y.; Forrest, L.R.; Schuldiner, S. Identification of molecular hinge points mediating alternating access in the vesicular monoamine transporter VMAT2. Proc. Natl. Acad. Sci. USA 2013, 110, E1332–E1341. [Google Scholar] [CrossRef] [PubMed]
- Cain, S.M.; Matzke, E.A.; Brooker, R.J. The conserved motif in hydrophilic loop 2/3 and loop 8/9 of the lactose permease of Escherichia coli. Analysis of suppressor mutations. J. Membr. Biol. 2000, 176, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Krämer, R. Functional principles of solute transport systems: concepts and perspectives. Biochimica et Biophysica Acta 1994, 1185, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Poolman, B.; Konings, W.N. Secondary solute transport in bacteria. Biochimica et Biophysica Acta 1993, 1183, 5–39. [Google Scholar] [CrossRef] [PubMed]
- Floyd, J.T.; Kumar, S.; Mukherjee, M.M.; He, G.; Varela, M.F. A review of the molecular mechanisms of drug efflux in pathogenic bacteria: A structure-function perspective. In Recent Research Developments in Membrane Biology; Shankar, P., Ed.; Research Signpost, Inc.: Kerala, India, 2013; Volume 3, pp. 15–66. [Google Scholar]
- Levy, S.B.; McMurry, L. Detection of an inducible membrane protein associated with R-factor-mediated tetracycline resistance. Biochem. Biophys. Res. Commun. 1974, 56, 1060–1068. [Google Scholar] [CrossRef] [PubMed]
- Levy, S.B.; McMurry, L. Plasmid-determined tetracycline resistance involves new transport systems for tetracycline. Nature 1978, 276, 90–92. [Google Scholar] [CrossRef] [PubMed]
- McMurry, L.; Levy, S.B. Two transport systems for tetracycline in sensitive Escherichia coli: Critical role for an initial rapid uptake system insensitive to energy inhibitors. Antimicrob. Agents Chemother. 1978, 14, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Levy, S.B. Active efflux mechanisms for antimicrobial resistance. Antimicrob. Agents Chemother. 1992, 36, 695–703. [Google Scholar] [CrossRef] [PubMed]
- West, I.; Mitchell, P. Proton-coupled β-galactoside translocation in non-metabolizing Escherichia coli. J. bioenerg. 1972, 3, 445–462. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P. Vectorial chemiosmotic processes. Ann. Rev. Biochem. 1977, 46, 996–1005. [Google Scholar] [CrossRef] [PubMed]
- Neyfakh, A.A.; Bidnenko, V.E.; Chen, L.B. Efflux-mediated multidrug resistance in Bacillus subtilis: Similarities and dissimilarities with the mammalian system. Proc. Natl. Acad. Sci. USA 1991, 88, 4781–4785. [Google Scholar] [CrossRef] [PubMed]
- Neyfakh, A.A. The multidrug efflux transporter of Bacillus subtilis is a structural and functional homolog of the Staphylococcus NorA protein. Antimicrob. Agents Chemother. 1992, 36, 484–485. [Google Scholar] [CrossRef] [PubMed]
- Neyfakh, A.A.; Borsch, C.M.; Kaatz, G.W. Fluoroquinolone resistance protein NorA of Staphylococcus aureus is a multidrug efflux transporter. Antimicrob. Agents Chemother. 1993, 37, 128–129. [Google Scholar] [CrossRef] [PubMed]
- Saier, M.H., Jr.; Beatty, J.T.; Goffeau, A.; Harley, K.T.; Heijne, W.H.; Huang, S.C.; Jack, D.L.; Jahn, P.S.; Lew, K.; Liu, J.; et al. The major facilitator superfamily. J. Mol. Microbiol. Biotechnol. 1999, 1, 257–279. [Google Scholar] [PubMed]
- Littlejohn, T.G.; Paulsen, I.T.; Gillespie, M.T.; Tennent, J.M.; Midgley, M.; Jones, I.G.; Purewal, A.S.; Skurray, R.A. Substrate specificity and energetics of antiseptic and disinfectant resistance in Staphylococcus aureus. FEMS Microbiol. Lett. 1992, 74, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Rouch, D.A.; Cram, D.S.; DiBerardino, D.; Littlejohn, T.G.; Skurray, R.A. Efflux-mediated antiseptic resistance gene qacA from Staphylococcus aureus: Common ancestry with tetracycline- and sugar-transport proteins. Mol. Microbiol. 1990, 4, 2051–2062. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.H.; Skurray, R.A. Staphylococcal multidrug efflux protein QacA. J. Mol. Microbiol. Biotechnol. 2001, 3, 163–170. [Google Scholar] [PubMed]
- Tennent, J.M.; Lyon, B.R.; Midgley, M.; Jones, I.G.; Purewal, A.S.; Skurray, R.A. Physical and biochemical characterization of the qacA gene encoding antiseptic and disinfectant resistance in Staphylococcus aureus. J. Gen. Microbiol. 1989, 135, 1–10. [Google Scholar] [PubMed]
- Paulsen, I.T.; Brown, M.H.; Littlejohn, T.G.; Mitchell, B.A.; Skurray, R.A. Multidrug resistance proteins QacA and QacB from Staphylococcus aureus: Membrane topology and identification of residues involved in substrate specificity. Proc. Natl. Acad. Sci. USA 1996, 93, 3630–3635. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, I.T.; Brown, M.H.; Skurray, R.A. Proton-dependent multidrug efflux systems. Microbiol. Rev. 1996, 60, 575–608. [Google Scholar] [PubMed]
- Ho, C.M.; Li, C.Y.; Ho, M.W.; Lin, C.Y.; Liu, S.H.; Lu, J.J. High rate of qacA- and qacB-positive methicillin-resistant Staphylococcus aureus isolates from chlorhexidine-impregnated catheter-related bloodstream infections. Antimicrob. Agents Chemother. 2012, 56, 5693–5697. [Google Scholar] [CrossRef] [PubMed]
- Otter, J.A.; Patel, A.; Cliff, P.R.; Halligan, E.P.; Tosas, O.; Edgeworth, J.D. Selection for qacA carriage in CC22, but not CC30, methicillin-resistant Staphylococcus aureus bloodstream infection isolates during a successful institutional infection control programme. J. Antimicrob. Chemother. 2013, 68, 992–999. [Google Scholar] [CrossRef] [PubMed]
- Shamsudin, M.N.; Alreshidi, M.A.; Hamat, R.A.; Alshrari, A.S.; Atshan, S.S.; Neela, V. High prevalence of qacA/B carriage among clinical isolates of meticillin-resistant Staphylococcus aureus in Malaysia. J. Hosp. Infect. 2012, 81, 206–208. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.; Branley, J. Prevalence of antiseptic resistance genes qacA/B and specific sequence types of methicillin-resistant Staphylococcus aureus in the era of hand hygiene. J. Antimicrob. Chemother. 2012, 67, 1549–1550. [Google Scholar] [CrossRef] [PubMed]
- Grkovic, S.; Brown, M.H.; Roberts, N.J.; Paulsen, I.T.; Skurray, R.A. QacR is a repressor protein that regulates expression of the Staphylococcus aureus multidrug efflux pump QacA. J. Biol. Chem. 1998, 273, 18665–18673. [Google Scholar] [CrossRef] [PubMed]
- Grkovic, S.; Brown, M.H.; Schumacher, M.A.; Brennan, R.G.; Skurray, R.A. The staphylococcal QacR multidrug regulator binds a correctly spaced operator as a pair of dimers. J. Bacteriol. 2001, 183, 7102–7109. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, M.A.; Miller, M.C.; Grkovic, S.; Brown, M.H.; Skurray, R.A.; Brennan, R.G. Structural basis for cooperative DNA binding by two dimers of the multidrug-binding protein QacR. EMBO J. 2002, 21, 1210–1218. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, M.A.; Miller, M.C.; Grkovic, S.; Brown, M.H.; Skurray, R.A.; Brennan, R.G. Structural mechanisms of QacR induction and multidrug recognition. Science 2001, 294, 2158–2163. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.S.; Viveiros, M.; Amaral, L.; Couto, I. Multidrug efflux pumps in Staphylococcus aureus: An update. Open Microbiol. J. 2013, 7, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Floyd, J.L.; Smith, K.P.; Kumar, S.H.; Floyd, J.T.; Varela, M.F. LmrS is a multidrug efflux pump of the major facilitator superfamily from Staphylococcus aureus. Antimicrob. Agents Chemother. 2010, 54, 5406–5412. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; O’Toole, P.W.; Shen, W.; Amrine-Madsen, H.; Jiang, X.; Lobo, N.; Palmer, L.M.; Voelker, L.; Fan, F.; Gwynn, M.N.; et al. Novel chromosomally encoded multidrug efflux transporter MdeA in Staphylococcus aureus. Antimicrob. Agents Chemother. 2004, 48, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Shiota, S.; Mizushima, T.; Kuroda, T.; Tsuchiya, T. Functional gene cloning and characterization of MdeA, a multidrug efflux pump from Staphylococcus aureus. Biol. Pharm. Bull. 2006, 29, 801–804. [Google Scholar] [CrossRef] [PubMed]
- Mirza, Z.M.; Kumar, A.; Kalia, N.P.; Zargar, A.; Khan, I.A. Piperine as an inhibitor of the MdeA efflux pump of Staphylococcus aureus. J. Med. Microbiol. 2011, 60, 1472–1478. [Google Scholar] [CrossRef] [PubMed]
- DeMarco, C.E.; Cushing, L.A.; Frempong-Manso, E.; Seo, S.M.; Jaravaza, T.A.; Kaatz, G.W. Efflux-related resistance to norfloxacin, dyes, and biocides in bloodstream isolates of Staphylococcus aureus. Antimicrob. Agents Chemother. 2007, 51, 3235–3239. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.S.; Mourato, C.; Viveiros, M.; Melo-Cristino, J.; Amaral, L.; Couto, I. Description of plasmid pSM52, harbouring the gene for the Smr efflux pump, and its involvement in resistance to biocides in a meticillin-resistant Staphylococcus aureus strain. Int. J. Antimicrob. Agents 2013, 41, 490–492. [Google Scholar] [CrossRef] [PubMed]
- Heir, E.; Sundheim, G.; Holck, A.L. The qacG gene on plasmid pST94 confers resistance to quaternary ammonium compounds in staphylococci isolated from the food industry. J. Appl. Microbiol. 1999, 86, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Heir, E.; Sundheim, G.; Holck, A.L. The Staphylococcus qacH gene product: A new member of the SMR family encoding multidrug resistance. FEMS Microbiol. Lett. 1998, 163, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Bjorland, J.; Steinum, T.; Sunde, M.; Waage, S.; Heir, E. Novel plasmid-borne gene qacJ mediates resistance to quaternary ammonium compounds in equine Staphylococcus aureus, Staphylococcus simulans, and Staphylococcus intermedius. Antimicrob. Agents Chemother. 2003, 47, 3046–3052. [Google Scholar] [CrossRef] [PubMed]
- Correa, J.E.; de Paulis, A.; Predari, S.; Sordelli, D.O.; Jeric, P.E. First report of qacG, qacH and qacJ genes in Staphylococcus haemolyticus human clinical isolates. J. Antimicrob. Chemother. 2008, 62, 956–960. [Google Scholar] [CrossRef] [PubMed]
- Ubukata, K.; Itoh-Yamashita, N.; Konno, M. Cloning and expression of the norA gene for fluoroquinolone resistance in Staphylococcus aureus. Antimicrob. Agents Chemother. 1989, 33. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Bogaki, M.; Nakamura, S.; Ubukata, K.; Konno, M. Nucleotide sequence and characterization of the Staphylococcus aureus norA gene, which confers resistance to quinolones. J. Bacteriol. 1990, 172, 6942–6949. [Google Scholar] [PubMed]
- Kaatz, G.W.; Seo, S.M.; Ruble, C.A. Efflux-mediated fluoroquinolone resistance in Staphylococcus aureus. Antimicrob. Agents Chemother. 1993, 37, 1086–1094. [Google Scholar] [CrossRef] [PubMed]
- Ng, E.; Trucksis, M.; Hooper, D.C. Quinolone resistance mediated by norA: Physiologic characterization and relationship to flqB, a quinolone resistance locus on the Staphylococcus aureus chromosome. Antimicrob. Agents Chemother. 1994, 38, 1345–1355. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Bellido, J.L.; Manzanares, M.A.; Andrés, J.M.; Zufiaurre, M.G.; Ortiz, G.; Hernández, M.S.; García-Rodríguez, J. Efflux Pump-Mediated Quinolone Resistance in Staphylococcus aureus Strains Wild Type for gyrA, gyrB, grlA, and norA. Antimicrob. Agents Chemother. 1999, 43, 354–356. [Google Scholar] [PubMed]
- Kaatz, G.W.; Thyagarajan, R.V.; Seo, S.M. Effect of promoter region mutations and mgrA overexpression on transcription of norA, which encodes a Staphylococcus aureus multidrug efflux transporter. Antimicrob. Agents Chemother. 2005, 49, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Holler, J.G.; Slotved, H.-C.; Mølgaard, P.; Olsen, C.E.; Christensen, S.B. Chalcone inhibitors of the NorA efflux pump in Staphylococcus aureus whole cells and enriched everted membrane vesicles. Bioorganic Med. Chem. 2012, 20, 4514–4521. [Google Scholar] [CrossRef]
- Kalia, N.P.; Mahajan, P.; Mehra, R.; Nargotra, A.; Sharma, J.P.; Koul, S.; Khan, I.A. Capsaicin, a novel inhibitor of the NorA efflux pump, reduces the intracellular invasion of Staphylococcus aureus. J. Antimicrob. Chemother. 2012, 67, 2401–2408. [Google Scholar] [CrossRef] [PubMed]
- Holler, J.G.; Christensen, S.B.; Slotved, H.-C.; Rasmussen, H.B.; Gúzman, A.; Olsen, C.-E.; Petersen, B.; Mølgaard, P. Novel inhibitory activity of the Staphylococcus aureus NorA efflux pump by a kaempferol rhamnoside isolated from Persea lingue Nees. J. Antimicrob. Chemother. 2012, 67, 1138–1144. [Google Scholar] [CrossRef] [PubMed]
- German, N.; Wei, P.; Kaatz, G.W.; Kerns, R.J. Synthesis and evaluation of fluoroquinolone derivatives as substrate-based inhibitors of bacterial efflux pumps. Eur. J. Med. Chem. 2008, 43, 2453–2463. [Google Scholar] [CrossRef] [PubMed]
- Pieroni, M.; Dimovska, M.; Brincat, J.P.; Sabatini, S.; Carosati, E.; Massari, S.; Kaatz, G.W.; Fravolini, A. From 6-aminoquinolone antibacterials to 6-amino-7-thiopyranopyridinylquinolone ethyl esters as inhibitors of Staphylococcus aureus multidrug efflux pumps. J. Med. Chem. 2010, 53, 4466–4480. [Google Scholar] [CrossRef] [PubMed]
- Vidaillac, C.; Guillon, J.; Arpin, C.; Forfar-Bares, I.; Ba, B.B.; Grellet, J.; Moreau, S.; Caignard, D.-H.; Jarry, C.; Quentin, C. Synthesis of omeprazole analogues and evaluation of these as potential inhibitors of the multidrug efflux pump NorA of Staphylococcus aureus. Antimicrob. Agents Chemother. 2007, 51, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Stavri, M.; Piddock, L.J.; Gibbons, S. Bacterial efflux pump inhibitors from natural sources. J. Antimicrob. Chemother. 2007, 59, 1247–1260. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Sun, F.; Ji, Q.; Liang, H.; Missiakas, D.; Lan, L.; He, C. Expression of multidrug resistance efflux pump gene norA is iron responsive in Staphylococcus aureus. J. Bacteriol. 2012, 194, 1753–1762. [Google Scholar] [CrossRef] [PubMed]
- Truong-Bolduc, Q.C.; Hooper, D.C. Phosphorylation of MgrA and its effect on expression of the NorA and NorB efflux pumps of Staphylococcus aureus. J. Bacteriol. 2010, 192, 2525–2534. [Google Scholar] [CrossRef] [PubMed]
- Truong-Bolduc, Q.C.; Ding, Y.; Hooper, D.C. Posttranslational modification influences the effects of MgrA on norA expression in Staphylococcus aureus. J. Bacteriol. 2008, 190, 7375–7381. [Google Scholar] [CrossRef] [PubMed]
- Truong-Bolduc, Q.C.; Hooper, D.C. The transcriptional regulators NorG and MgrA modulate resistance to both quinolones and β-lactams in Staphylococcus aureus. J. Bacteriol. 2007, 189, 2996–3005. [Google Scholar] [CrossRef] [PubMed]
- Luong, T.T.; Newell, S.W.; Lee, C.Y. Mgr, a novel global regulator in Staphylococcus aureus. J. Bacteriol. 2003, 185, 3703–3710. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zheng, L.; Landwehr, C.; Lunsford, D.; Holmes, D.; Ji, Y. Global regulation of gene expression by ArlRS, a two-component signal transduction regulatory system of Staphylococcus aureus. J. Bacteriol. 2005, 187, 5486–5492. [Google Scholar] [CrossRef] [PubMed]
- Truong-Bolduc, Q.; Dunman, P.; Strahilevitz, J.; Projan, S.; Hooper, D. MgrA is a multiple regulator of two new efflux pumps in Staphylococcus aureus. J. Bacteriol. 2005, 187, 2395–2405. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Onodera, Y.; Lee, J.C.; Hooper, D.C. NorB, an efflux pump in Staphylococcus aureus strain MW2, contributes to bacterial fitness in abscesses. J. Bacteriol. 2008, 190, 7123–7129. [Google Scholar] [CrossRef] [PubMed]
- Truong-Bolduc, Q.C.; Bolduc, G.R.; Okumura, R.; Celino, B.; Bevis, J.; Liao, C.-H.; Hooper, D.C. Implication of the NorB efflux pump in the adaptation of Staphylococcus aureus to growth at acid pH and in resistance to moxifloxacin. Antimicrob. Agents Chemother. 2011, 55, 3214–3219. [Google Scholar] [CrossRef] [PubMed]
- Truong-Bolduc, Q.C.; Hsing, L.C.; Villet, R.; Bolduc, G.R.; Estabrooks, Z.; Taguezem, G.F.; Hooper, D.C. Reduced aeration affects the expression of the NorB efflux pump of Staphylococcus aureus by posttranslational modification of MgrA. J. Bacteriol. 2012, 194, 1823–1834. [Google Scholar] [CrossRef] [PubMed]
- Truong-Bolduc, Q.C.; Strahilevitz, J.; Hooper, D.C. NorC, a new efflux pump regulated by MgrA of Staphylococcus aureus. Antimicrob. Agents Chemother. 2006, 50, 1104–1107. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Fu, Y.; Lee, J.C.; Hooper, D.C. Staphylococcus aureus NorD, a putative efflux pump coregulated with the Opp1 oligopeptide permease, contributes selectively to fitness in vivo. J. Bacteriol. 2012, 194, 6586–6593. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Feßler, A.T.; Hauschild, T.; Kehrenberg, C.; Kadlec, K. Plasmid‐mediated resistance to protein biosynthesis inhibitors in staphylococci. Ann. N. Y. Acad. Sci. 2011, 1241, 82–103. [Google Scholar] [CrossRef] [PubMed]
- Krulwich, T.A.; Jin, J.; Guffanti, A.A.; Bechhofer, D.H. Functions of tetracycline efflux proteins that do not involve tetracycline. J. Mol. Microbiol. Biotechnol. 2001, 3, 237–246. [Google Scholar] [PubMed]
- Ginn, S.L.; Brown, M.H.; Skurray, R.A. The TetA (K) tetracycline/H+ antiporter from Staphylococcus aureus: Mutagenesis and functional analysis of motif C. J. Bacteriol. 2000, 182, 1492–1498. [Google Scholar] [CrossRef] [PubMed]
- Guay, G.G.; Rothstein, D.M. Expression of the tetK gene from Staphylococcus aureus in Escherichia coli: Comparison of substrate specificities of TetA (B), TetA (C), and TetK efflux proteins. Antimicrob. Agents Chemother. 1993, 37, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Fujihira, E.; Kimura, T.; Shiina, Y.; Yamaguchi, A. Transmembrane glutamic acid residues play essential roles in the metal-tetracycline/H+ antiporter of Staphylococcus aureus. FEBS Lett. 1996, 391, 243–246. [Google Scholar] [CrossRef] [PubMed]
- Sudano Roccaro, A.; Blanco, A.R.; Giuliano, F.; Rusciano, D.; Enea, V. Epigallocatechin-gallate enhances the activity of tetracycline in staphylococci by inhibiting its efflux from bacterial cells. Antimicrob. Agents Chemother. 2004, 48, 1968–1973. [Google Scholar] [CrossRef] [PubMed]
- Oluwatuyi, M.; Kaatz, G.W.; Gibbons, S. Antibacterial and resistance modifying activity of Rosmarinus officinalis. Phytochemistry 2004, 65, 3249–3254. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, S.; Oluwatuyi, M.; Veitch, N.C.; Gray, A.I. Bacterial resistance modifying agents from Lycopus europaeus. Phytochemistry 2003, 62, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Handzlik, J.; Matys, A.; Kieć-Kononowicz, K. Recent advances in multi-drug resistance (MDR) efflux pump inhibitors of Gram-positive bacteria S. aureus. Antibiotics 2013, 2, 28–45. [Google Scholar] [CrossRef]
- Yamada, Y.; Hideka, K.; Shiota, S.; Kuroda, T.; Tsuchiya, T. Gene cloning and characterization of SdrM, a chromosomally-encoded multidrug efflux pump, from Staphylococcus aureus. Biol. Pharm. Bull. 2006, 29, 554–556. [Google Scholar] [CrossRef] [PubMed]
- Sutcliffe, J.; Tait-Kamradt, A.; Wondrack, L. Streptococcus pneumoniae and Streptococcus pyogenes resistant to macrolides but sensitive to clindamycin: A common resistance pattern mediated by an efflux system. Antimicrob. Agents Chemother. 1996, 40, 1817–1824. [Google Scholar] [PubMed]
- Poole, K. Efflux-mediated antimicrobial resistance. J. Antimicrob. Chemother. 2005, 56, 20–51. [Google Scholar] [CrossRef] [PubMed]
- Clancy, J.; Petitpas, J.; Dib‐Hajj, F.; Yuan, W.; Cronan, M.; Kamath, A.V.; Bergeron, J.; Retsema, J.A. Molecular cloning and functional analysis of a novel macrolide‐resistance determinant, mefA, from Streptococcus pyogenes. Mol. Microbiol. 1996, 22, 867–879. [Google Scholar] [CrossRef] [PubMed]
- Luna, V.A.; Heiken, M.; Judge, K.; Ulep, C.; van Kirk, N.; Luis, H.; Bernardo, M.; Leitao, J.; Roberts, M.C. Distribution of mef (A) in gram-positive bacteria from healthy Portuguese children. Antimicrob. Agents Chemother. 2002, 46, 2513–2517. [Google Scholar] [CrossRef] [PubMed]
- Luna, V.A.; Coates, P.; Eady, E.A.; Cove, J.H.; Nguyen, T.T.; Roberts, M.C. A variety of gram-positive bacteria carry mobile mef genes. J. Antimicrob. Chemother. 1999, 44, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Ojo, K.; Ulep, C.; van Kirk, N.; Luis, H.; Bernardo, M.; Leitao, J.; Roberts, M. The mef (A) gene predominates among seven macrolide resistance genes identified in Gram-negative strains representing 13 genera, isolated from healthy Portuguese children. Antimicrob. Agents Chemother. 2004, 48, 3451–3456. [Google Scholar] [CrossRef] [PubMed]
- Tait-Kamradt, A.; Clancy, J.; Cronan, M.; Dib-Hajj, F.; Wondrack, L.; Yuan, W.; Sutcliffe, J. mefE is necessary for the erythromycin-resistant M phenotype in Streptococcus pneumoniae. Antimicrob. Agents Chemother. 1997, 41, 2251–2255. [Google Scholar] [PubMed]
- Du, D.; Wang, Z.; James, N.R.; Voss, J.E.; Klimont, E.; Ohene-Agyei, T.; Venter, H.; Chiu, W.; Luisi, B.F. Structure of the AcrAB-TolC multidrug efflux pump. Nature 2014, 509, 512–515. [Google Scholar] [CrossRef] [PubMed]
- Weeks, J.W.; Bavro, V.N.; Misra, R. Genetic assessment of the role of AcrB β-hairpins in the assembly of the TolC-AcrAB multidrug efflux pump of Escherichia coli. Mol. Microbiol. 2014, 91, 965–975. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Yokota, S.; Okubo, T.; Ishihara, K.; Ueno, H.; Muramatsu, Y.; Fujii, N.; Tamura, Y. Contribution of the AcrAB-TolC efflux pump to high-level fluoroquinolone resistance in Escherichia coli isolated from dogs and humans. J. Vet. Med. Sci. 2013, 75, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Sikora, C.W.; Turner, R.J. SMR proteins SugE and EmrE bind ligand with similar affinity and stoichiometry. Biochem. Biophys. Res. Commun. 2005, 335, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Son, M.S.; Del Castilho, C.; Duncalf, K.A.; Carney, D.; Weiner, J.H.; Turner, R.J. Mutagenesis of SugE, a small multidrug resistance protein. Biochem. Biophys. Res. Commun. 2003, 312, 914–921. [Google Scholar] [CrossRef] [PubMed]
- Cortez-Cordova, J.; Kumar, A. Activity of the efflux pump inhibitor phenylalanine-arginine β-naphthylamide against the AdeFGH pump of Acinetobacter baumannii. Int. J. Antimicrob. Agents 2011, 37, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Reddy, V.S.; Shlykov, M.A.; Castillo, R.; Sun, E.I.; Saier, M.H., Jr. The major facilitator superfamily (MFS) revisited. FEBS J. 2012, 279, 2022–2035. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Z.; Nikaido, H. Efflux-mediated drug resistance in bacteria: An update. Drugs 2009, 69, 1555–1623. [Google Scholar] [CrossRef] [PubMed]
- Sigal, N.; Cohen-Karni, D.; Siemion, S.; Bibi, E. MdfA from Escherichia coli, a model protein for studying secondary multidrug transport. J. Mol. Microbiol. Biotechnol. 2006, 11, 308–317. [Google Scholar] [CrossRef] [PubMed]
- White, D.G.; Hudson, C.; Maurer, J.J.; Ayers, S.; Zhao, S.; Lee, M.D.; Bolton, L.; Foley, T.; Sherwood, J. Characterization of chloramphenicol and florfenicol resistance in Escherichia coli associated with bovine diarrhea. J. Clin. Microbiol. 2000, 38, 4593–4598. [Google Scholar] [PubMed]
- Xu, Y.; Sim, S.H.; Nam, K.H.; Jin, X.L.; Kim, H.M.; Hwang, K.Y.; Lee, K.; Ha, N.C. Crystal structure of the periplasmic region of MacB, a noncanonic ABC transporter. Biochemistry 2009, 48, 5218–5225. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Sim, S.H.; Song, S.; Piao, S.; Kim, H.M.; Jin, X.L.; Lee, K.; Ha, N.C. The tip region of the MacA α-hairpin is important for the binding to TolC to the Escherichia coli MacAB-TolC pump. Biochem. Biophys. Res. Commun. 2010, 394, 962–965. [Google Scholar] [CrossRef] [PubMed]
- Guyer, D.M.; Kao, J.S.; Mobley, H.L. Genomic analysis of a pathogenicity island in uropathogenic Escherichia coli CFT073: Distribution of homologous sequences among isolates from patients with pyelonephritis, cystitis, and Catheter-associated bacteriuria and from fecal samples. Infect. Immun. 1998, 66, 4411–4417. [Google Scholar] [PubMed]
- Hayashi, T.; Makino, K.; Ohnishi, M.; Kurokawa, K.; Ishii, K.; Yokoyama, K.; Han, C.G.; Ohtsubo, E.; Nakayama, K.; Murata, T.; et al. Complete genome sequence of enterohemorrhagic Escherichia coli O157:H7 and genomic comparison with a laboratory strain K-12. DNA Res. 2001, 8, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.P.; Kumar, S.; Varela, M.F. Identification, cloning, and functional characterization of EmrD-3, a putative multidrug efflux pump of the major facilitator superfamily from Vibrio cholerae O395. Arch. Microbiol. 2009, 191, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, M.; Kakarla, P.; Kumar, S.; Gonzalez, E.; Floyd, J.T.; Inupakutika, M.; Devireddy, A.R.; Tirrell, S.R.; Bruns, M.; He, G.; et al. Comparative genome analysis of non-toxigenic non-O1 versus toxigenic O1 Vibrio cholerae. Genomics Discov. 2014, 2. [Google Scholar] [CrossRef]
- Woolley, R.C.; Vediyappan, G.; Anderson, M.; Lackey, M.; Ramasubramanian, B.; Jiangping, B.; Borisova, T.; Colmer, J.A.; Hamood, A.N.; McVay, C.S.; et al. Characterization of the Vibrio cholerae vceCAB multiple-drug resistance efflux operon in Escherichia coli. J. Bacteriol. 2005, 187, 5500–5503. [Google Scholar] [CrossRef] [PubMed]
- Colmer, J.A.; Fralick, J.A.; Hamood, A.N. Isolation and characterization of a putative multidrug resistance pump from Vibrio cholerae. Mol. Microbiol. 1998, 27, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Huda, M.N.; Chen, J.; Morita, Y.; Kuroda, T.; Mizushima, T.; Tsuchiya, T. Gene cloning and characterization of VcrM, a Na+-coupled multidrug efflux pump, from Vibrio cholerae non-O1. Microbiol. Immunol. 2003, 47, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Mukherjee, M.M.; Varela, M.F. Modulation of bacterial multidrug resistance efflux pumps of the major facilitator superfamily. Int. J. Bacteriol. 2013, 10. [Google Scholar] [CrossRef]
- Sun, J.; Deng, Z.; Yan, A. Bacterial multidrug efflux pumps: Mechanisms, physiology and pharmacological exploitations. Biochem. Biophys. Res. Commun. 2014, 453, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Horiyama, T.; Yamaguchi, A.; Nishino, K. TolC dependency of multidrug efflux systems in Salmonella enterica serovar Typhimurium. J. Antimicrob. Chemother. 2010, 65, 1372–1376. [Google Scholar] [CrossRef] [PubMed]
- Nishino, K.; Latifi, T.; Groisman, E.A. Virulence and drug resistance roles of multidrug efflux systems of Salmonella enterica serovar Typhimurium. Mol. Microbiol. 2006, 59, 126–141. [Google Scholar] [CrossRef] [PubMed]
- Fluman, N.; Bibi, E. Bacterial multidrug transport through the lens of the major facilitator superfamily. Biochimica et Biophysica Acta 2009, 1794, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Rensch, U.; Nishino, K.; Klein, G.; Kehrenberg, C. Salmonella enterica serovar Typhimurium multidrug efflux pumps EmrAB and AcrEF support the major efflux system AcrAB in decreased susceptibility to triclosan. Int. J. Antimicrob. Agents 2014, 44, 179–180. [Google Scholar] [CrossRef] [PubMed]
- Lomovskaya, O.; Lewis, K.; Matin, A. EmrR is a negative regulator of the Escherichia coli multidrug resistance pump EmrAB. J. Bacteriol. 1995, 177, 2328–2334. [Google Scholar] [PubMed]
- Lewinson, O.; Padan, E.; Bibi, E. Alkali tolerance: A biological function for a multidrug transporter in pH homeostasis. Proc. Natl. Acad. Sci. USA 2004, 101, 14073–14078. [Google Scholar] [CrossRef] [PubMed]
- Braibant, M.; Chevalier, J.; Chaslus-Dancla, E.; Pages, J.M.; Cloeckaert, A. Structural and functional study of the phenicol-specific efflux pump FloR belonging to the major facilitator superfamily. Antimicrob. Agents Chemother. 2005, 49, 2965–2971. [Google Scholar] [CrossRef] [PubMed]
- Cheung, T.K.; Chu, Y.W.; Chu, M.Y.; Ma, C.H.; Yung, R.W.; Kam, K.M. Plasmid-mediated resistance to ciprofloxacin and cefotaxime in clinical isolates of Salmonella enterica serotype Enteritidis in Hong Kong. J. Antimicrob. Chemother. 2005, 56, 586–589. [Google Scholar] [CrossRef] [PubMed]
- Randall, L.P.; Cooles, S.W.; Osborn, M.K.; Piddock, L.J.; Woodward, M.J. Antibiotic resistance genes, integrons and multiple antibiotic resistance in thirty-five serotypes of Salmonella enterica isolated from humans and animals in the UK. J. Antimicrob. Chemother. 2004, 53, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Hur, J.; Jawale, C.; Lee, J.H. Antimicrobial resistance of Salmonella isolated from food animals: A review. Food Res. Int. 2012, 45, 819–830. [Google Scholar] [CrossRef]
- Randall, L.P.; Woodward, M.J. The multiple antibiotic resistance (mar) locus and its significance. Res. Vet. Sci. 2002, 72, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Arcangioli, M.A.; Leroy-Setrin, S.; Martel, J.L.; Chaslus-Dancla, E. A new chloramphenicol and florfenicol resistance gene flanked by two integron structures in Salmonella typhimurium DT104. FEMS Microbiol. Lett. 1999, 174, 327–332. [Google Scholar]
- Akiyama, T.; Presedo, J.; Khan, A.A. The tetA gene decreases tigecycline sensitivity of Salmonella enterica isolates. Int. J. Antimicrob. Agents 2013, 42, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Shahada, F.; Amamoto, A.; Chuma, T.; Shirai, A.; Okamoto, K. Antimicrobial susceptibility phenotypes, resistance determinants and DNA fingerprints of Salmonella enterica serotype Typhimurium isolated from bovine in Southern Japan. Int. J. Antimicrob. Agents 2007, 30, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Shahada, F.; Chuma, T.; Tobata, T.; Okamoto, K.; Sueyoshi, M.; Takase, K. Molecular epidemiology of antimicrobial resistance among Salmonella enterica serovar infantis from poultry in Kagoshima, Japan. Int. J. Antimicrob. Agents 2006, 28, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, L.; Nodwell, J.R. The TetR family of regulators. Microbiol. Mol. Biol. Rev. 2013, 77, 440–475. [Google Scholar] [CrossRef] [PubMed]
- He, G.X.; Kuroda, T.; Mima, T.; Morita, Y.; Mizushima, T.; Tsuchiya, T. An H+-coupled multidrug efflux pump, PmpM, a member of the MATE family of transporters, from Pseudomonas aeruginosa. J. Bacteriol. 2004, 186, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.H.; Paulsen, I.T.; Skurray, R.A. The multidrug efflux protein NorM is a prototype of a new family of transporters. Mol. Microbiol. 1999, 31, 394–395. [Google Scholar] [CrossRef] [PubMed]
- Dridi, L.; Tankovic, J.; Petit, J.C. CdeA of Clostridium difficile, a new multidrug efflux transporter of the MATE family. Microb. Drug Resist. 2004, 10, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Perez, A.; Poza, M.; Fernandez, A.; Fernandez Mdel, C.; Mallo, S.; Merino, M.; Rumbo-Feal, S.; Cabral, M.P.; Bou, G. Involvement of the AcrAB-TolC efflux pump in the resistance, fitness, and virulence of Enterobacter cloacae. Antimicrob. Agents Chemother. 2012, 56, 2084–2090. [Google Scholar] [CrossRef] [PubMed]
- Perez, A.; Poza, M.; Aranda, J.; Latasa, C.; Medrano, F.J.; Tomas, M.; Romero, A.; Lasa, I.; Bou, G. Effect of transcriptional activators SoxS, RobA, and RamA on expression of multidrug efflux pump AcrAB-TolC in Enterobacter cloacae. Antimicrob. Agents Chemother. 2012, 56, 6256–6266. [Google Scholar] [CrossRef] [PubMed]
- Szabo, D.; Silveira, F.; Hujer, A.M.; Bonomo, R.A.; Hujer, K.M.; Marsh, J.W.; Bethel, C.R.; Doi, Y.; Deeley, K.; Paterson, D.L. Outer membrane protein changes and efflux pump expression together may confer resistance to ertapenem in Enterobacter cloacae. Antimicrob. Agents Chemother. 2006, 50, 2833–2835. [Google Scholar] [CrossRef] [PubMed]
- Sacha, P.; Wieczorek, P.; Ojdana, D.; Hauschild, T.; Milewski, R.; Czaban, S.; Poniatowski, B.; Tryniszewska, E. Expression of MexAB-OprM efflux pump system and susceptibility to antibiotics of different Pseudomonas aeruginosa clones isolated from patients hospitalized in two intensive care units at University Hospital in Bialystok (northeastern Poland) between January 2002 and December 2009. APMIS 2014, 122, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Bay, D.C.; Rommens, K.L.; Turner, R.J. Small multidrug resistance proteins: A multidrug transporter family that continues to grow. Biochimica et Biophysica Acta 2008, 1778, 1814–1838. [Google Scholar] [CrossRef] [PubMed]
- Griffith, J.K.; Baker, M.E.; Rouch, D.A.; Page, M.G.; Skurray, R.A.; Paulsen, I.T.; Chater, K.F.; Baldwin, S.A.; Henderson, P.J. Membrane transport proteins: Implications of sequence comparisons. Curr. Opin. Cell Biol. 1992, 4, 684–695. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, S.A.; Henderson, P.J. Homologies between sugar transporters from eukaryotes and prokaryotes. Ann. Rev. Physiol. 1989, 51, 459–471. [Google Scholar] [CrossRef]
- Henderson, P.J.; Roberts, P.E.; Martin, G.E.; Seamon, K.B.; Walmsley, A.R.; Rutherford, N.G.; Varela, M.F.; Griffith, J.K. Homologous sugar-transport proteins in microbes and man. Biochem. Soc. Trans. 1993, 21, 1002–1006. [Google Scholar] [PubMed]
- Henderson, P.J.; Maiden, M.C. Homologous sugar transport proteins in Escherichia coli and their relatives in both prokaryotes and eukaryotes. Philos. Trans. R. Soc. Lond. Ser. B. 1990, 326, 391–410. [Google Scholar] [CrossRef]
- Pao, S.S.; Paulsen, I.T.; Saier, M.H., Jr. Major facilitator superfamily. Microbiol. Mol. Biol. Rev. 1998, 62, 1–34. [Google Scholar] [PubMed]
- Saier, M.H., Jr.; Reddy, V.S.; Tamang, D.G.; Vastermark, A. The transporter classification database. Nucleic Acids Res. 2014, 42, D251–D258. [Google Scholar] [CrossRef] [PubMed]
- Hirai, T.; Heymann, J.A.; Maloney, P.C.; Subramaniam, S. Structural model for 12-helix transporters belonging to the major facilitator superfamily. J. Bacteriol. 2003, 185, 1712–1718. [Google Scholar] [CrossRef] [PubMed]
- Saier, M.H., Jr. A functional-phylogenetic classification system for transmembrane solute transporters. Microbiol. Mol. Biol. Rev. 2000, 64, 354–411. [Google Scholar] [CrossRef] [PubMed]
- Bolhuis, H.; Poelarends, G.; van Veen, H.W.; Poolman, B.; Driessen, A.J.; Konings, W.N. The Lactococcal lmrP gene encodes a proton motive force-dependent drug transporter. J. Biol. Chem. 1995, 270, 26092–26098. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; He, X.; Szewczyk, P.; Nguyen, T.; Chang, G. Structure of the multidrug transporter EmrD from Escherichia coli. Science 2006, 312, 741–744. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.; Bibi, E. MdfA, an Escherichia coli multidrug resistance protein with an extraordinarily broad spectrum of drug recognition. J. Bacteriol. 1997, 179, 2274–2280. [Google Scholar] [PubMed]
- Yelin, R.; Schuldiner, S. The pharmacological profile of the vesicular monoamine transporter resembles that of multidrug transporters. FEBS Lett. 1995, 377, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Sutcliffe, J.G. Complete nucleotide sequence of the Escherichia coli plasmid pBR322. Cold Spring Harb. Symp. Quant. Biol. 1979, 43, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Hillen, W.; Schollmeier, K. Nucleotide sequence of the Tn10 encoded tetracycline resistance gene. Nucleic acids research 1983, 11, 525–539. [Google Scholar] [CrossRef] [PubMed]
- Goffeau, A.; Park, J.; Paulsen, I.T.; Jonniaux, J.L.; Dinh, T.; Mordant, P.; Saier, M.H., Jr. Multidrug-resistant transport proteins in yeast: Complete inventory and phylogenetic characterization of yeast open reading frames with the major facilitator superfamily. Yeast 1997, 13, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Lomovskaya, O.; Lewis, K. Emr, an Escherichia coli locus for multidrug resistance. Proc. Natl. Acad. Sci. USA 1992, 89, 8938–8942. [Google Scholar] [CrossRef] [PubMed]
- Hongo, E.; Morimyo, M.; Mita, K.; Machida, I.; Hama-Inaba, H.; Tsuji, H.; Ichimura, S.; Noda, Y. The methyl viologen-resistance-encoding gene smvA of Salmonella typhimurium. Gene 1994, 148, 173–174. [Google Scholar] [CrossRef] [PubMed]
- Guffanti, A.A.; Krulwich, T.A. Tetracycline/H+ antiport and Na+/H+ antiport catalyzed by the Bacillus subtilis TetA(L) transporter expressed in Escherichia coli. J. Bacteriol. 1995, 177, 4557–4561. [Google Scholar] [PubMed]
- Khan, S.A.; Novick, R.P. Complete nucleotide sequence of pT181, a tetracycline-resistance plasmid from Staphylococcus aureus. Plasmid 1983, 10, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Marger, M.D.; Saier, M.H., Jr. A major superfamily of transmembrane facilitators that catalyse uniport, symport and antiport. Trends Biochem. Sci. 1993, 18, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Kimura-Someya, T.; Iwaki, S.; Konishi, S.; Tamura, N.; Kubo, Y.; Yamaguchi, A. Cysteine-scanning mutagenesis around transmembrane segments 1 and 11 and their flanking loop regions of Tn10-encoded metal-Tetracycline/H+ antiporter. J. Biol. Chem. 2000, 275, 18692–18697. [Google Scholar] [CrossRef] [PubMed]
- Varela, M.F.; Wilson, T.H.; Rodon-Rivera, V.; Shepherd, S.; Dehne, T.A.; Rector, A.C. Mutants of the lactose carrier of Escherichia coli which show altered sugar recognition plus a severe defect in sugar accumulation. J. Membr. Biol. 2000, 174, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Varela, M.F.; Wilson, T.H. Molecular biology of the lactose carrier of Escherichia coli. Biochimica et Biophysica Acta 1996, 1276, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Van Camp, B.M.; Crow, R.R.; Peng, Y.; Varela, M.F. Amino acids that confer transport of raffinose and maltose sugars in the raffinose permease (RafB) of Escherichia coli as implicated by spontaneous mutations at Val-35, Ser-138, Ser-139, Gly-389 and Ile-. J. Membr. Biol. 2007, 220, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, A.; Nakatani, M.; Sawai, T. Aspartic acid-66 is the only essential negatively charged residue in the putative hydrophilic loop region of the metal-tetracycline/H+ antiporter encoded by transposon Tn10 of Escherichia coli. Biochemistry 1992, 31, 8344–8348. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, A.; Ono, N.; Akasaka, T.; Noumi, T.; Sawai, T. Metal-tetracycline/H+ antiporter of Escherichia coli encoded by transposon Tn10: The role of the conserved dipeptide, Ser65-Asp66, in tetracycline transport. J. Biol. Chem. 1990, 265, 15525–15530. [Google Scholar] [PubMed]
- Tamura, N.; Konishi, S.; Yamaguchi, A. Mechanisms of drug/H+ antiport: Complete cysteine-scanning mutagenesis and the protein engineering approach. Curr. Opin. Chem. Biol. 2003, 7, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Someya, Y.; Kimura-Someya, T.; Yamaguchi, A. Role of the charge interaction between Arg(70) and Asp(120) in the Tn10-encoded metal-tetracycline/H+ antiporter of Escherichia coli. J. Biol. Chem. 2000, 275, 210–214. [Google Scholar] [CrossRef] [PubMed]
- Someya, Y.; Yamaguchi, A. Mercaptide formed between the residue Cys70 and Hg2+ or Co2+ behaves as a functional positively charged side chain operative in the Arg70→Cys mutant of the metal-tetracycline/H+ antiporter of Escherichia coli. Biochemistry 1996, 35, 9385–9391. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Nakatani, M.; Kawabe, T.; Yamaguchi, A. Roles of conserved arginine residues in the metal-tetracycline/H+ antiporter of Escherichia coli. Biochemistry 1998, 37, 5475–5480. [Google Scholar] [CrossRef] [PubMed]
- Pazdernik, N.J.; Matzke, E.A.; Jessen-Marshall, A.E.; Brooker, R.J. Roles of charged residues in the conserved motif, G-X-X-X-D/E-R/K-X-G-[X]-R/K-R/K, of the lactose permease of Escherichia coli. J. Membr. Biol. 2000, 174, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Jessen-Marshall, A.E.; Parker, N.J.; Brooker, R.J. Suppressor analysis of mutations in the loop 2–3 motif of lactose permease: Evidence that glycine-64 is an important residue for conformational changes. J. Bacteriol. 1997, 179, 2616–2622. [Google Scholar] [PubMed]
- Jessen-Marshall, A.E.; Paul, N.J.; Brooker, R.J. The conserved motif, GXXX(D/E)(R/K)XG[X](R/K)(R/K), in hydrophilic loop 2/3 of the lactose permease. J. Biol. Chem. 1995, 270, 16251–16257. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Zhao, Y.; Wang, X.; Fan, J.; Heng, J.; Liu, X.; Feng, W.; Kang, X.; Huang, B.; Liu, J.; Zhang, X.C. Structure of the YajR transporter suggests a transport mechanism based on the conserved motif A. Proc. Natl. Acad. Sci. USA 2013, 110, 14664–14669. [Google Scholar] [CrossRef] [PubMed]
- Varela, M.F.; Griffith, J.K. Nucleotide and deduced protein sequences of the class D tetracycline resistance determinant: relationship to other antimicrobial transport proteins. Antimicrob. Agents Chemother. 1993, 37, 1253–1258. [Google Scholar] [CrossRef] [PubMed]
- Varela, M.F.; Sansom, C.E.; Griffith, J.K. Mutational analysis and molecular modelling of an amino acid sequence motif conserved in antiporters but not symporters in a transporter superfamily. Mol. Membr. Biol. 1995, 12, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Saraceni-Richards, C.A.; Levy, S.B. Second-site suppressor mutations of inactivating substitutions at Gly247 of the tetracycline efflux protein, Tet(B). J. Bacteriol. 2000, 182, 6514–6516. [Google Scholar] [CrossRef]
- De Jesus, M.; Jin, J.; Guffanti, A.A.; Krulwich, T.A. Importance of the GP dipeptide of the antiporter motif and other membrane-embedded proline and glycine residues in tetracycline efflux protein Tet(L). Biochemistry 2005, 44, 12896–12904. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Krulwich, T.A. Site-directed mutagenesis studies of selected motif and charged residues and of cysteines of the multifunctional tetracycline efflux protein Tet(L). J. Bacteriol. 2002, 184, 1796–1800. [Google Scholar] [CrossRef] [PubMed]
- Perreten, V.; Schwarz, F.V.; Teuber, M.; Levy, S.B. Mdt(A), a new efflux protein conferring multiple antibiotic resistance in Lactococcus lactis and Escherichia coli. Antimicrob. Agents Chemother. 2001, 45, 1109–1114. [Google Scholar] [CrossRef] [PubMed]
- Hassan, K.A.; Galea, M.; Wu, J.; Mitchell, B.A.; Skurray, R.A.; Brown, M.H. Functional effects of intramembranous proline substitutions in the staphylococcal multidrug transporter QacA. FEMS Microbiol. Lett. 2006, 263, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Pasrija, R.; Banerjee, D.; Prasad, R. Structure and function analysis of CaMdr1p, a major facilitator superfamily antifungal efflux transporter protein of Candida albicans: identification of amino acid residues critical for drug/H+ transport. Eukaryotic Cell 2007, 6, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Walther, C.; Rossano, A.; Thomann, A.; Perreten, V. Antibiotic resistance in Lactococcus species from bovine milk: Presence of a mutated multidrug transporter mdt(A) gene in susceptible Lactococcus garvieae strains. Vet. Microbiol. 2008, 131, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, A.; Ojeda, A.M.; Kolmakova, N.G.; Parsons, S.M. Mutational and bioinformatics analysis of proline- and glycine-rich motifs in vesicular acetylcholine transporter. J. Neurochem. 2006, 98, 1551–1559. [Google Scholar] [CrossRef] [PubMed]
- Ugolev, Y.; Segal, T.; Yaffe, D.; Gros, Y.; Schuldiner, S. Identification of conformationally sensitive residues essential for inhibition of vesicular monoamine transport by the noncompetitive inhibitor tetrabenazine. J. Biol. Chem. 2013, 288, 32160–32171. [Google Scholar] [CrossRef] [PubMed]
- Iancu, C.V.; Zamoon, J.; Woo, S.B.; Aleshin, A.; Choe, J.Y. Crystal structure of a glucose/H+ symporter and its mechanism of action. Proc. Natl. Acad. Sci. USA 2013, 110, 17862–17867. [Google Scholar] [CrossRef] [PubMed]
- Maiden, M.C.; Davis, E.O.; Baldwin, S.A.; Moore, D.C.; Henderson, P.J. Mammalian and bacterial sugar transport proteins are homologous. Nature 1987, 325, 641–643. [Google Scholar] [CrossRef] [PubMed]
- Newstead, S.; Drew, D.; Cameron, A.D.; Postis, V.L.; Xia, X.; Fowler, P.W.; Ingram, J.C.; Carpenter, E.P.; Sansom, M.S.; McPherson, M.J.; Baldwin, S.A.; Iwata, S. Crystal structure of a prokaryotic homologue of the mammalian oligopeptide-proton symporters, PepT1 and PepT2. EMBO J. 2011, 30, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Lemieux, M.J.; Song, J.; Kim, M.J.; Huang, Y.; Villa, A.; Auer, M.; Li, X.D.; Wang, D.N. Three-dimensional crystallization of the Escherichia coli glycerol-3-phosphate transporter: A member of the major facilitator superfamily. Protein Sci. 2003, 12, 2748–2756. [Google Scholar] [CrossRef] [PubMed]
- Abramson, J.; Smirnova, I.; Kasho, V.; Verner, G.; Kaback, H.R.; Iwata, S. Structure and mechanism of the lactose permease of Escherichia coli. Science 2003, 301, 610–615. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Wisedchaisri, G.; Gonen, T. Crystal structure of a nitrate/nitrite exchanger. Nature 2013, 497, 647–651. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Huang, W.; Yan, C.; Gong, X.; Jiang, S.; Zhao, Y.; Wang, J.; Shi, Y. Structure and mechanism of a nitrate transporter. Cell Rep. 2013, 3, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Dang, S.; Sun, L.; Huang, Y.; Lu, F.; Liu, Y.; Gong, H.; Wang, J.; Yan, N. Structure of a fucose transporter in an outward-open conformation. Nature 2010, 467, 734–738. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zeng, X.; Yan, C.; Sun, X.; Gong, X.; Rao, Y.; Yan, N. Crystal structure of a bacterial homologue of glucose transporters GLUT1–4. Nature 2012, 490, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.P.; Kumar, H.; Waight, A.B.; Risenmay, A.J.; Roe-Zurz, Z.; Chau, B.H.; Schlessinger, A.; Bonomi, M.; Harries, W.; Sali, A.; et al. Crystal structure of a eukaryotic phosphate transporter. Nature 2013, 496, 533–536. [Google Scholar] [CrossRef] [PubMed]
- Deng, D.; Xu, C.; Sun, P.; Wu, J.; Yan, C.; Hu, M.; Yan, N. Crystal structure of the human glucose transporter GLUT1. Nature 2014, 510, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Yan, N. Structural advances for the major facilitator superfamily (MFS) transporters. Trends Biochem. Sci. 2013, 38, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H.; Pages, J.M. Broad-specificity efflux pumps and their role in multidrug resistance of Gram-negative bacteria. FEMS Microbiol. Rev. 2012, 36, 340–363. [Google Scholar] [CrossRef] [PubMed]
- Swick, M.C.; Morgan-Linnell, S.K.; Carlson, K.M.; Zechiedrich, L. Expression of multidrug efflux pump genes acrAB-tolC, mdfA, and norE in Escherichia coli clinical isolates as a function of fluoroquinolone and multidrug resistance. Antimicrob. Agents Chemother. 2011, 55, 921–924. [Google Scholar] [CrossRef] [PubMed]
- Yasufuku, T.; Shigemura, K.; Shirakawa, T.; Matsumoto, M.; Nakano, Y.; Tanaka, K.; Arakawa, S.; Kinoshita, S.; Kawabata, M.; Fujisawa, M. Correlation of overexpression of efflux pump genes with antibiotic resistance in Escherichia coli strains clinically isolated from urinary tract infection patients. J. Clin. Microbiol. 2011, 49, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Davin-Regli, A.; Bolla, J.M.; James, C.E.; Lavigne, J.P.; Chevalier, J.; Garnotel, E.; Molitor, A.; Pages, J.M. Membrane permeability and regulation of drug “influx and efflux” in enterobacterial pathogens. Curr. Drug Targets 2008, 9, 750–759. [Google Scholar] [CrossRef] [PubMed]
- Piddock, L.J. Clinically relevant chromosomally encoded multidrug resistance efflux pumps in bacteria. Clin. Microbiol. Rev. 2006, 19, 382–402. [Google Scholar] [CrossRef] [PubMed]
- Bornet, C.; Chollet, R.; Mallea, M.; Chevalier, J.; Davin-Regli, A.; Pages, J.M.; Bollet, C. Imipenem and expression of multidrug efflux pump in Enterobacter aerogenes. Biochem. Biophys. Res. Commun. 2003, 301, 985–990. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, P.; Ferreira, B.S.; Cabral, J.M. Solvent tolerance in bacteria: Role of efflux pumps and cross-resistance with antibiotics. Int. J. Antimicrob. Agents 2003, 22, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Mazzariol, A.; Zuliani, J.; Cornaglia, G.; Rossolini, G.M.; Fontana, R. AcrAB Efflux System: Expression and Contribution to Fluoroquinolone Resistance in Klebsiella spp. Antimicrob. Agents Chemother. 2002, 46, 3984–3986. [Google Scholar] [CrossRef] [PubMed]
- Giamarellou, H.; Poulakou, G. Multidrug-resistant Gram-negative infections: What are the treatment options? Drugs 2009, 69, 1879–1901. [Google Scholar] [CrossRef] [PubMed]
- Rice, L.B. The clinical consequences of antimicrobial resistance. Curr. Opin. Microbiol. 2009, 12, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Gwynn, M.N.; Portnoy, A.; Rittenhouse, S.F.; Payne, D.J. Challenges of antibacterial discovery revisited. Ann. N. Y. Acad. Sci. 2010, 1213, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Z.; Nikaido, H. Efflux-mediated drug resistance in bacteria. Drugs 2004, 64, 159–204. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H. Multidrug resistance in bacteria. Ann. Rev. Biochem. 2009, 78, 119–146. [Google Scholar] [CrossRef] [PubMed]
- Warner, D.M.; Shafer, W.M.; Jerse, A.E. Clinically relevant mutations that cause derepression of the Neisseria gonorrhoeae MtrC-MtrD-MtrE Efflux pump system confer different levels of antimicrobial resistance and in vivo fitness. Mol. Microbiol. 2008, 70, 462–478. [Google Scholar] [CrossRef] [PubMed]
- Olliver, A.; Valle, M.; Chaslus-Dancla, E.; Cloeckaert, A. Overexpression of the multidrug efflux operon acrEF by insertional activation with IS1 or IS10 elements in Salmonella enterica serovar Typhimurium DT204 acrB mutants selected with fluoroquinolones. Antimicrob. Agents Chemother. 2005, 49, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Vettoretti, L.; Plesiat, P.; Muller, C.; el Garch, F.; Phan, G.; Attree, I.; Ducruix, A.; Llanes, C. Efflux unbalance in Pseudomonas aeruginosa isolates from cystic fibrosis patients. Antimicrob. Agents Chemother. 2009, 53, 1987–1997. [Google Scholar] [CrossRef] [PubMed]
- Chuanchuen, R.; Beinlich, K.; Hoang, T.T.; Becher, A.; Karkhoff-Schweizer, R.R.; Schweizer, H.P. Cross-resistance between triclosan and antibiotics in Pseudomonas aeruginosa is mediated by multidrug efflux pumps: exposure of a susceptible mutant strain to triclosan selects nfxB mutants overexpressing MexCD-OprJ. Antimicrob. Agents Chemother. 2001, 45, 428–432. [Google Scholar] [CrossRef] [PubMed]
- Saleh, S.; Haddadin, R.N.; Baillie, S.; Collier, P.J. Triclosan—An update. Lett. Appl. Microbiol. 2011, 52, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Dann, A.B.; Hontela, A. Triclosan: Environmental exposure, toxicity and mechanisms of action. J. Appl. Toxicol. 2011, 31, 285–311. [Google Scholar] [CrossRef] [PubMed]
- Yazdankhah, S.P.; Scheie, A.A.; Hoiby, E.A.; Lunestad, B.T.; Heir, E.; Fotland, T.O.; Naterstad, K.; Kruse, H. Triclosan and antimicrobial resistance in bacteria: An overview. Microb. Drug Resist. 2006, 12, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Fralick, J.A. Evidence that TolC is required for functioning of the Mar/AcrAB efflux pump of Escherichia coli. J. Bacteriol. 1996, 178, 5803–5805. [Google Scholar] [PubMed]
- Yamane, K.; Wachino, J.; Suzuki, S.; Kimura, K.; Shibata, N.; Kato, H.; Shibayama, K.; Konda, T.; Arakawa, Y. New plasmid-mediated fluoroquinolone efflux pump, QepA, found in an Escherichia coli clinical isolate. Antimicrob. Agents Chemother. 2007, 51, 3354–3360. [Google Scholar] [CrossRef] [PubMed]
- Blair, J.M.; Piddock, L.J. Structure, function and inhibition of RND efflux pumps in Gram-negative bacteria: An update. Curr. Opin. Microbiol. 2009, 12, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Morita, Y.; Kodama, K.; Shiota, S.; Mine, T.; Kataoka, A.; Mizushima, T.; Tsuchiya, T. NorM, a putative multidrug efflux protein, of Vibrio parahaemolyticus and its homolog in Escherichia coli. Antimicrob. Agents Chemother. 1998, 42, 1778–1782. [Google Scholar] [PubMed]
- O’Regan, E.; Quinn, T.; Pages, J.M.; McCusker, M.; Piddock, L.; Fanning, S. Multiple regulatory pathways associated with high-level ciprofloxacin and multidrug resistance in Salmonella enterica serovar enteritidis: Involvement of RamA and other global regulators. Antimicrob. Agents Chemother. 2009, 53, 1080–1087. [Google Scholar] [CrossRef] [PubMed]
- Lunn, A.D.; Fabrega, A.; Sanchez-Cespedes, J.; Vila, J. Prevalence of mechanisms decreasing quinolone-susceptibility among Salmonella spp. clinical isolates. Int. Microbiol. 2010, 13, 15–20. [Google Scholar] [PubMed]
- Giraud, E.; Cloeckaert, A.; Kerboeuf, D.; Chaslus-Dancla, E. Evidence for active efflux as the primary mechanism of resistance to ciprofloxacin in Salmonella enterica serovar Typhimurium. Antimicrob. Agents Chemother. 2000, 44, 1223–1228. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, L.; Hancock, R.E. Adaptive and mutational resistance: Role of porins and efflux pumps in drug resistance. Clin. Microbiol. Rev. 2012, 25, 661–681. [Google Scholar] [CrossRef]
- Martins, A.; Hunyadi, A.; Amaral, L. Mechanisms of resistance in bacteria: An evolutionary approach. Open Microbiol. J. 2013, 7, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Poole, K. Efflux pumps as antimicrobial resistance mechanisms. Ann. Med. 2007, 39, 162–176. [Google Scholar] [CrossRef] [PubMed]
- Alekshun, M.N.; Levy, S.B. Regulation of chromosomally mediated multiple antibiotic resistance: The mar regulon. Antimicrob. Agents Chemother. 1997, 41, 2067–2075. [Google Scholar] [PubMed]
- Keeney, D.; Ruzin, A.; McAleese, F.; Murphy, E.; Bradford, P.A. MarA-mediated overexpression of the AcrAB efflux pump results in decreased susceptibility to tigecycline in Escherichia coli. J. Antimicrob. Chemother. 2008, 61, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Baucheron, S.; Tyler, S.; Boyd, D.; Mulvey, M.R.; Chaslus-Dancla, E.; Cloeckaert, A. AcrAB-TolC directs efflux-mediated multidrug resistance in Salmonella enterica serovar Typhimurium DT. Antimicrob. Agents Chemother. 2004, 48, 3729–3735. [Google Scholar] [CrossRef] [PubMed]
- Piddock, L.J.; White, D.G.; Gensberg, K.; Pumbwe, L.; Griggs, D.J. Evidence for an efflux pump mediating multiple antibiotic resistance in Salmonella enterica serovar Typhimurium. Antimicrob. Agents Chemother. 2000, 44, 3118–3121. [Google Scholar] [CrossRef] [PubMed]
- Hocquet, D.; Vogne, C.; el Garch, F.; Vejux, A.; Gotoh, N.; Lee, A.; Lomovskaya, O.; Plesiat, P. MexXY-OprM efflux pump is necessary for a adaptive resistance of Pseudomonas aeruginosa to aminoglycosides. Antimicrob. Agents Chemother. 2003, 47, 1371–1375. [Google Scholar] [CrossRef] [PubMed]
- Masuda, N.; Sakagawa, E.; Ohya, S.; Gotoh, N.; Tsujimoto, H.; Nishino, T. Contribution of the MexX-MexY-oprM efflux system to intrinsic resistance in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2000, 44, 2242–2246. [Google Scholar] [CrossRef] [PubMed]
- Eaves, D.J.; Ricci, V.; Piddock, L.J. Expression of acrB, acrF, acrD, marA, and soxS in Salmonella enterica serovar Typhimurium: Role in multiple antibiotic resistance. Antimicrob. Agents Chemother. 2004, 48, 1145–1150. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, L.; Machado, D.; Couto, I.; Amaral, L.; Viveiros, M. Contribution of efflux activity to isoniazid resistance in the Mycobacterium tuberculosis complex. Infect. Genet. Evol. 2012, 12, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Kwak, Y.G.; Truong-Bolduc, Q.C.; Bin Kim, H.; Song, K.H.; Kim, E.S.; Hooper, D.C. Association of norB overexpression and fluoroquinolone resistance in clinical isolates of Staphylococcus aureus from Korea. J. Antimicrob. Chemother. 2013, 68, 2766–2772. [Google Scholar] [CrossRef] [PubMed]
- Llanes, C.; Hocquet, D.; Vogne, C.; Benali-Baitich, D.; Neuwirth, C.; Plesiat, P. Clinical strains of Pseudomonas aeruginosa overproducing MexAB-OprM and MexXY efflux pumps simultaneously. Antimicrob. Agents Chemother. 2004, 48, 1797–1802. [Google Scholar] [CrossRef] [PubMed]
- Ziha-Zarifi, I.; Llanes, C.; Kohler, T.; Pechere, J.C.; Plesiat, P. In vivo emergence of multidrug-resistant mutants of Pseudomonas aeruginosa overexpressing the active efflux system MexA-MexB-OprM. Antimicrob. Agents Chemother. 1999, 43, 287–291. [Google Scholar] [PubMed]
- Becnel Boyd, L.; Maynard, M.J.; Morgan-Linnell, S.K.; Horton, L.B.; Sucgang, R.; Hamill, R.J.; Jimenez, J.R.; Versalovic, J.; Steffen, D.; Zechiedrich, L. Relationships among ciprofloxacin, gatifloxacin, levofloxacin, and norfloxacin MICs for fluoroquinolone-resistant Escherichia coli clinical isolates. Antimicrob. Agents Chemother. 2009, 53, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Morgan-Linnell, S.K.; Becnel Boyd, L.; Steffen, D.; Zechiedrich, L. Mechanisms accounting for fluoroquinolone resistance in Escherichia coli clinical isolates. Antimicrob. Agents Chemother. 2009, 53, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Yoon, E.J.; Courvalin, P.; Grillot-Courvalin, C. RND-type efflux pumps in multidrug-resistant clinical isolates of Acinetobacter baumannii: Major role for AdeABC overexpression and AdeRS mutations. Antimicrob. Agents Chemother. 2013, 57, 2989–2995. [Google Scholar] [CrossRef] [PubMed]
- Coyne, S.; Courvalin, P.; Perichon, B. Efflux-mediated antibiotic resistance in Acinetobacter spp. Antimicrob. Agents Chemother. 2011, 55, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Magnet, S.; Courvalin, P.; Lambert, T. Resistance-nodulation-cell division-type efflux pump involved in aminoglycoside resistance in Acinetobacter baumannii strain BM4. Antimicrob. Agents Chemother. 2001, 45, 3375–3380. [Google Scholar] [CrossRef] [PubMed]
- Pannek, S.; Higgins, P.G.; Steinke, P.; Jonas, D.; Akova, M.; Bohnert, J.A.; Seifert, H.; Kern, W.V. Multidrug efflux inhibition in Acinetobacter baumannii: comparison between 1-(1-naphthylmethyl)-piperazine and phenyl-arginine-β-naphthylamide. J. Antimicrob. Chemother. 2006, 57, 970–974. [Google Scholar] [CrossRef] [PubMed]
- Marchand, I.; Damier-Piolle, L.; Courvalin, P.; Lambert, T. Expression of the RND-type efflux pump AdeABC in Acinetobacter baumannii is regulated by the AdeRS two-component system. Antimicrob. Agents Chemother. 2004, 48, 3298–3304. [Google Scholar] [CrossRef] [PubMed]
- Peleg, A.Y.; Potoski, B.A.; Rea, R.; Adams, J.; Sethi, J.; Capitano, B.; Husain, S.; Kwak, E.J.; Bhat, S.V.; Paterson, D.L. Acinetobacter baumannii bloodstream infection while receiving tigecycline: A cautionary report. J. Antimicrob. Chemother. 2007, 59, 128–131. [Google Scholar] [CrossRef] [PubMed]
- Ruzin, A.; Keeney, D.; Bradford, P.A. AdeABC multidrug efflux pump is associated with decreased susceptibility to tigecycline in Acinetobacter calcoaceticus-Acinetobacter baumannii complex. J. Antimicrob. Chemother. 2007, 59, 1001–1004. [Google Scholar] [CrossRef] [PubMed]
- Linares, J.F.; Lopez, J.A.; Camafeita, E.; Albar, J.P.; Rojo, F.; Martinez, J.L. Overexpression of the multidrug efflux pumps MexCD-OprJ and MexEF-OprN is associated with a reduction of type III secretion in Pseudomonas aeruginosa. J. Bacteriol. 2005, 187, 1384–1391. [Google Scholar] [CrossRef] [PubMed]
- Pumbwe, L.; Randall, L.P.; Woodward, M.J.; Piddock, L.J. Evidence for multiple-antibiotic resistance in Campylobacter jejuni not mediated by CmeB or CmeF. Antimicrob. Agents Chemother. 2005, 49, 1289–1293. [Google Scholar] [CrossRef] [PubMed]
- Poelarends, G.J.; Mazurkiewicz, P.; Putman, M.; Cool, R.H.; Veen, H.W.; Konings, W.N. An ABC-type multidrug transporter of Lactococcus lactis possesses an exceptionally broad substrate specificity. Drug Resist. Updat. 2000, 3, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, K.T.; Carleton, J.D.; Quillin, S.J.; Rollins, S.D.; Portnoy, D.A.; Leber, J.H. Hyperinduction of host β-interferon by a Listeria monocytogenes strain naturally overexpressing the multidrug efflux pump MdrT. Infect. Immun. 2012, 80, 1537–1545. [Google Scholar] [CrossRef] [PubMed]
- Crimmins, G.T.; Herskovits, A.A.; Rehder, K.; Sivick, K.E.; Lauer, P.; Dubensky, T.W., Jr.; Portnoy, D.A. Listeria monocytogenes multidrug resistance transporters activate a cytosolic surveillance pathway of innate immunity. Proc. Natl. Acad. Sci. USA 2008, 105, 10191–10196. [Google Scholar] [CrossRef] [PubMed]
- Woodward, J.J.; Iavarone, A.T.; Portnoy, D.A. c-di-AMP secreted by intracellular Listeria monocytogenes activates a host type I interferon response. Science 2010, 328, 1703–1705. [Google Scholar] [CrossRef] [PubMed]
- Bina, J.E.; Provenzano, D.; Wang, C.; Bina, X.R.; Mekalanos, J.J. Characterization of the Vibrio cholerae vexAB and vexCD efflux systems. Arch. Microbiol. 2006, 186, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Blair, J.M.; la Ragione, R.M.; Woodward, M.J.; Piddock, L.J. Periplasmic adaptor protein AcrA has a distinct role in the antibiotic resistance and virulence of Salmonella enterica serovar Typhimurium. J. Antimicrob. Chemother. 2009, 64, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.; Iversen, C.; Rodrigues, L.; Spengler, G.; Ramos, J.; Kern, W.V.; Couto, I.; Viveiros, M.; Fanning, S.; Pages, J.M.; et al. An AcrAB-mediated multidrug-resistant phenotype is maintained following restoration of wild-type activities by efflux pump genes and their regulators. Int. J. Antimicrob. Agents 2009, 34, 602–604. [Google Scholar] [CrossRef] [PubMed]
- Padilla, E.; Llobet, E.; Domenech-Sanchez, A.; Martinez-Martinez, L.; Bengoechea, J.A.; Alberti, S. Klebsiella pneumoniae AcrAB efflux pump contributes to antimicrobial resistance and virulence. Antimicrob. Agents Chemother. 2010, 54, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Webber, M.A.; Randall, L.P.; Cooles, S.; Woodward, M.J.; Piddock, L.J. Triclosan resistance in Salmonella enterica serovar Typhimurium. J. Antimicrob. Chemother. 2008, 62, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Alhede, M.; Bjarnsholt, T.; Givskov, M.; Alhede, M. Pseudomonas aeruginosa biofilms: Mechanisms of immune evasion. Adv. Appl. Microbiol. 2014, 86, 1–40. [Google Scholar] [PubMed]
- Gillis, R.J.; White, K.G.; Choi, K.H.; Wagner, V.E.; Schweizer, H.P.; Iglewski, B.H. Molecular basis of azithromycin-resistant Pseudomonas aeruginosa biofilms. Antimicrob. Agents Chemother. 2005, 49, 3858–3867. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Mah, T.F. Involvement of a novel efflux system in biofilm-specific resistance to antibiotics. J. Bacteriol. 2008, 190, 4447–4452. [Google Scholar] [CrossRef] [PubMed]
- Kvist, M.; Hancock, V.; Klemm, P. Inactivation of efflux pumps abolishes bacterial biofilm formation. Appl. Environ. Microbiol. 2008, 74, 7376–7382. [Google Scholar] [CrossRef] [PubMed]
- Baugh, S.; Ekanayaka, A.S.; Piddock, L.J.; Webber, M.A. Loss of or inhibition of all multidrug resistance efflux pumps of Salmonella enterica serovar Typhimurium results in impaired ability to form a biofilm. J. Antimicrob. Chemother. 2012, 67, 2409–2417. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, A.K.; Mohanty, P. Bacterial efflux pumps involved in multidrug resistance and their inhibitors: Rejuvinating the antimicrobial chemotherapy. Recent Pat. Anti-Infect. Drug Discov. 2012, 7, 73–89. [Google Scholar] [CrossRef]
- Zechini, B.; Versace, I. Inhibitors of multidrug resistant efflux systems in bacteria. Recent Pat. Anti-Infect. Drug Discov. 2009, 4, 37–50. [Google Scholar] [CrossRef]
- Lewis, K. In search of natural substrates and inhibitors of MDR pumps. J. Mol. Microbiol. Biotechnol. 2001, 3, 247–254. [Google Scholar] [PubMed]
- Markham, P.N.; Westhaus, E.; Klyachko, K.; Johnson, M.E.; Neyfakh, A.A. Multiple novel inhibitors of the NorA multidrug transporter of Staphylococcus aureus. Antimicrob. Agents Chemother. 1999, 43, 2404–2408. [Google Scholar] [PubMed]
- Lomovskaya, O.; Warren, M.S.; Lee, A.; Galazzo, J.; Fronko, R.; Lee, M.; Blais, J.; Cho, D.; Chamberland, S.; Renau, T.; et al. Identification and characterization of inhibitors of multidrug resistance efflux pumps in Pseudomonas aeruginosa: novel agents for combination therapy. Antimicrob. Agents Chemother. 2001, 45, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [PubMed]
- Takatsuka, Y.; Chen, C.; Nikaido, H. Mechanism of recognition of compounds of diverse structures by the multidrug efflux pump AcrB of Escherichia coli. Proc. Natl. Acad. Sci. USA 2010, 107, 6559–6565. [Google Scholar] [CrossRef] [PubMed]
- Murakami, S.; Nakashima, R.; Yamashita, E.; Yamaguchi, A. Crystal structure of bacterial multidrug efflux transporter AcrB. Nature 2002, 419, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Sennhauser, G.; Amstutz, P.; Briand, C.; Storchenegger, O.; Grutter, M.G. Drug export pathway of multidrug exporter AcrB revealed by DARPin inhibitors. PLoS Biol. 2007, 5. [Google Scholar] [CrossRef] [PubMed]
- Sennhauser, G.; Bukowska, M.A.; Briand, C.; Grutter, M.G. Crystal structure of the multidrug exporter MexB from Pseudomonas aeruginosa. J. Mol. Biol. 2009, 389, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, R.; Sakurai, K.; Yamasaki, S.; Hayashi, K.; Nagata, C.; Hoshino, K.; Onodera, Y.; Nishino, K.; Yamaguchi, A. Structural basis for the inhibition of bacterial multidrug exporters. Nature 2013, 500, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Kourtesi, C.; Ball, A.R.; Huang, Y.Y.; Jachak, S.M.; Vera, D.M.; Khondkar, P.; Gibbons, S.; Hamblin, M.R.; Tegos, G.P. Microbial efflux systems and inhibitors: approaches to drug discovery and the challenge of clinical implementation. Open Microbiol. J. 2013, 7, 34–52. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.R.; Perng, C.L.; Lin, J.C.; Yang, Y.S.; Chan, M.C.; Chang, T.Y.; Lin, F.M.; Chiueh, T.S. AdeRS combination codes differentiate the response to efflux pump inhibitors in tigecycline-resistant isolates of extensively drug-resistant Acinetobacter baumannii. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 12. [Google Scholar] [CrossRef]
- Baugh, S.; Phillips, C.R.; Ekanayaka, A.S.; Piddock, L.J.; Webber, M.A. Inhibition of multidrug efflux as a strategy to prevent biofilm formation. J. Antimicrob. Chemother. 2014, 69, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Hannula, M.; Hanninen, M.L. Effect of putative efflux pump inhibitors and inducers on the antimicrobial susceptibility of Campylobacter jejuni and Campylobacter coli. J. Med. Microbiol. 2008, 57, 851–855. [Google Scholar] [CrossRef] [PubMed]
- Bohnert, J.A.; Kern, W.V. Selected arylpiperazines are capable of reversing multidrug resistance in Escherichia coli overexpressing RND efflux pumps. Antimicrob. Agents Chemother. 2005, 49, 849–852. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, A.; Steinke, P.; Bohnert, J.A.; Akova, M.; Jonas, D.; Kern, W.V. Effect of 1-(1-naphthylmethyl)-piperazine, a novel putative efflux pump inhibitor, on antimicrobial drug susceptibility in clinical isolates of Enterobacteriaceae other than Escherichia coli. J. Antimicrob. Chemother. 2006, 57, 344–348. [Google Scholar] [CrossRef] [PubMed]
- Mallea, M.; Mahamoud, A.; Chevalier, J.; Alibert-Franco, S.; Brouant, P.; Barbe, J.; Pages, J.M. Alkylaminoquinolines inhibit the bacterial antibiotic efflux pump in multidrug-resistant clinical isolates. Biochem. J. 2003, 376, 801–805. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, J.; Bredin, J.; Mahamoud, A.; Mallea, M.; Barbe, J.; Pages, J.M. Inhibitors of antibiotic efflux in resistant Enterobacter aerogenes and Klebsiella pneumoniae strains. Antimicrob. Agents Chemother. 2004, 48, 1043–1046. [Google Scholar] [CrossRef] [PubMed]
- Nagata, S. Lethal effect of carbonyl cyanide m-chlorophenylhydrazone on Escherichia coli and a halotolerant Brevibacterium species. Microbios 1995, 81, 73–83. [Google Scholar] [PubMed]
- Zhang, Z.; Liu, Z.Q.; Zheng, P.Y.; Tang, F.A.; Yang, P.C. Influence of efflux pump inhibitors on the multidrug resistance of Helicobacter pylori. World J. Gastroenterol. 2010, 16, 1279–1284. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Kumar, M.; Sharma, S.; Nargotra, A.; Koul, S.; Khan, I.A. Piperine as an inhibitor of Rv1258c, a putative multidrug efflux pump of Mycobacterium tuberculosis. J. Antimicrob. Chemother. 2010, 65, 1694–1701. [Google Scholar] [CrossRef] [PubMed]
- Sangwan, P.L.; Koul, J.L.; Koul, S.; Reddy, M.V.; Thota, N.; Khan, I.A.; Kumar, A.; Kalia, N.P.; Qazi, G.N. Piperine analogs as potent Staphylococcus aureus NorA efflux pump inhibitors. Bioorganic Med. Chem. 2008, 16, 9847–9857. [Google Scholar] [CrossRef]
- Khan, I.A.; Mirza, Z.M.; Kumar, A.; Verma, V.; Qazi, G.N. Piperine, a phytochemical potentiator of ciprofloxacin against Staphylococcus aureus. Antimicrob. Agents Chemother. 2006, 50, 810–812. [Google Scholar] [CrossRef] [PubMed]
- Handzlik, J.; Szymanska, E.; Alibert, S.; Chevalier, J.; Otrebska, E.; Pekala, E.; Kiec-Kononowicz, K.; Pages, J.M. Search for new tools to combat Gram-negative resistant bacteria among amine derivatives of 5-arylidenehydantoin. Bioorganic Med. Chem. 2013, 21, 135–145. [Google Scholar] [CrossRef]
- Aparna, V.; Dineshkumar, K.; Mohanalakshmi, N.; Velmurugan, D.; Hopper, W. Identification of natural compound inhibitors for multidrug efflux pumps of Escherichia coli and Pseudomonas aeruginosa using in silico high-throughput virtual screening and in vitro validation. PLoS One 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Kuete, V.; Alibert-Franco, S.; Eyong, K.O.; Ngameni, B.; Folefoc, G.N.; Nguemeving, J.R.; Tangmouo, J.G.; Fotso, G.W.; Komguem, J.; Ouahouo, B.M.; et al. Antibacterial activity of some natural products against bacteria expressing a multidrug-resistant phenotype. Int. J. Antimicrob. Agents 2011, 37, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Barnes, C.A.; Rasmussen, S.L.; Petrich, J.W.; Rasmussen, M.A. Determination of the concentration of potential efflux pump inhibitors, pheophorbide a and pyropheophorbide a, in the feces of animals by fluorescence spectroscopy. J. Agric. Food Chem. 2012, 60, 10456–10460. [Google Scholar] [CrossRef] [PubMed]
- Agerso, Y.; Aarestrup, F.M. Voluntary ban on cephalosporin use in Danish pig production has effectively reduced extended-spectrum cephalosporinase-producing Escherichia coli in slaughter pigs. J. Antimicrob. Chemother. 2013, 68, 569–572. [Google Scholar] [CrossRef] [PubMed]
- Aarestrup, F.M.; Jensen, V.F.; Emborg, H.D.; Jacobsen, E.; Wegener, H.C. Changes in the use of antimicrobials and the effects on productivity of swine farms in Denmark. Am. J. Vet. Res. 2010, 71, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.R.; Cho, I.H.; Jeong, B.C.; Lee, S.H. Strategies to minimize antibiotic resistance. Int. J. Environ. Res. Public Health 2013, 10, 4274–4305. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, D.H. Alternatives to conventional antimicrobial drugs: A review of future prospects. Vet. Dermatol. 2012, 23, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Hsu, L.Y.; Kwa, A.L.; Lye, D.C.; Chlebicki, M.P.; Tan, T.Y.; Ling, M.L.; Wong, S.Y.; Goh, L.G. Reducing antimicrobial resistance through appropriate antibiotic usage in Singapore. Singap. Med. J. 2008, 49, 749–755. [Google Scholar]
- Garcia-Migura, L.; Hendriksen, R.S.; Fraile, L.; Aarestrup, F.M. Antimicrobial resistance of zoonotic and commensal bacteria in Europe: The missing link between consumption and resistance in veterinary medicine. Vet. Microbiol. 2014, 170, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Aarestrup, F.M.; Wegener, H.C.; Collignon, P. Resistance in bacteria of the food chain: Epidemiology and control strategies. Expert Rev. Anti-Infect. Ther. 2008, 6, 733–750. [Google Scholar] [CrossRef] [PubMed]
- Aarestrup, F.M.; Wegener, H.C. The effects of antibiotic usage in food animals on the development of antimicrobial resistance of importance for humans in Campylobacter and Escherichia coli. Microbes Infect. 1999, 1, 639–644. [Google Scholar] [CrossRef] [PubMed]
- MacDougall, C.; Polk, R.E. Antimicrobial stewardship programs in health care systems. Clin. Microbiol. Rev. 2005, 18, 638–656. [Google Scholar] [CrossRef] [PubMed]
- Paskovaty, A.; Pflomm, J.M.; Myke, N.; Seo, S.K. A multidisciplinary approach to antimicrobial stewardship: Evolution into the 21st century. Int. J. Antimicrob. Agents 2005, 25, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Croucher, N.J.; Hanage, W.P.; Harris, S.R.; McGee, L.; van der Linden, M.; de Lencastre, H.; Sa-Leao, R.; Song, J.H.; Ko, K.S.; Beall, B.; et al. Variable recombination dynamics during the emergence, transmission and “disarming” of a multidrug-resistant pneumococcal clone. BMC Biol. 2014, 12. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Lindquist, I.E.; Sundararajan, A.; Rajanna, C.; Floyd, J.T.; Smith, K.P.; Andersen, J.L.; He, G.; Ayers, R.M.; Johnson, J.A.; et al. Genome Sequence of Non-O1 Vibrio cholerae PS. Genome Announce. 2013, 1, e00227:1–e00227:12. [Google Scholar]
- Tiwari, R.; Dhama, K.; Kumar, A.; Rahal, A.; Kapoor, S. Bacteriophage therapy for safeguarding animal and human health: A review. Pak. J. Biol. Sci. 2014, 17, 301–315. [Google Scholar] [CrossRef] [PubMed]
- Endersen, L.; O’Mahony, J.; Hill, C.; Ross, R.P.; McAuliffe, O.; Coffey, A. Phage therapy in the food industry. Ann. Rev. Food Sci. Technol. 2014, 5, 327–349. [Google Scholar] [CrossRef]
- Drawz, S.M.; Papp-Wallace, K.M.; Bonomo, R.A. New β-lactamase inhibitors: A therapeutic renaissance in an MDR world. Antimicrob. Agents Chemother. 2014, 58, 1835–1846. [Google Scholar] [CrossRef] [PubMed]
- King, D.T.; Strynadka, N.C. Targeting metallo-β-lactamase enzymes in antibiotic resistance. Future Med. Chem. 2013, 5, 1243–1263. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.; Ma, J.; Rothnie, A.; Biggin, P.C.; Kerr, I.D. Towards understanding promiscuity in multidrug efflux pumps. Trends Biochem. Sci. 2014, 39, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Schweizer, H.P. Understanding efflux in Gram-negative bacteria: opportunities for drug discovery. Expert Opin. Drug discov. 2012, 7, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Schindler, B.D.; Jacinto, P.; Kaatz, G.W. Inhibition of drug efflux pumps in Staphylococcus aureus: Current status of potentiating existing antibiotics. Future Microbiol. 2013, 8, 491–507. [Google Scholar] [CrossRef] [PubMed]
- Varela, M.F.; Kumar, S.; He, G. Potential for inhibition of bacterial efflux pumps in multidrug-resistant Vibrio cholerae. Indian J. Med. Res. 2013, 138. [Google Scholar] [CrossRef]
- Henderson, P.J. Studies of translocation catalysis. Biosci. Rep. 1991, 11, 477–453. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andersen, J.L.; He, G.-X.; Kakarla, P.; KC, R.; Kumar, S.; Lakra, W.S.; Mukherjee, M.M.; Ranaweera, I.; Shrestha, U.; Tran, T.; et al. Multidrug Efflux Pumps from Enterobacteriaceae, Vibrio cholerae and Staphylococcus aureus Bacterial Food Pathogens. Int. J. Environ. Res. Public Health 2015, 12, 1487-1547. https://doi.org/10.3390/ijerph120201487
Andersen JL, He G-X, Kakarla P, KC R, Kumar S, Lakra WS, Mukherjee MM, Ranaweera I, Shrestha U, Tran T, et al. Multidrug Efflux Pumps from Enterobacteriaceae, Vibrio cholerae and Staphylococcus aureus Bacterial Food Pathogens. International Journal of Environmental Research and Public Health. 2015; 12(2):1487-1547. https://doi.org/10.3390/ijerph120201487
Chicago/Turabian StyleAndersen, Jody L., Gui-Xin He, Prathusha Kakarla, Ranjana KC, Sanath Kumar, Wazir Singh Lakra, Mun Mun Mukherjee, Indrika Ranaweera, Ugina Shrestha, Thuy Tran, and et al. 2015. "Multidrug Efflux Pumps from Enterobacteriaceae, Vibrio cholerae and Staphylococcus aureus Bacterial Food Pathogens" International Journal of Environmental Research and Public Health 12, no. 2: 1487-1547. https://doi.org/10.3390/ijerph120201487