
Rapid and Sensitive Identification of the Herbal Tea Ingredient Taraxacum formosanum Using Loop-Mediated Isothermal Amplification

 ,
,
Abstract
:1. Introduction
2. Results
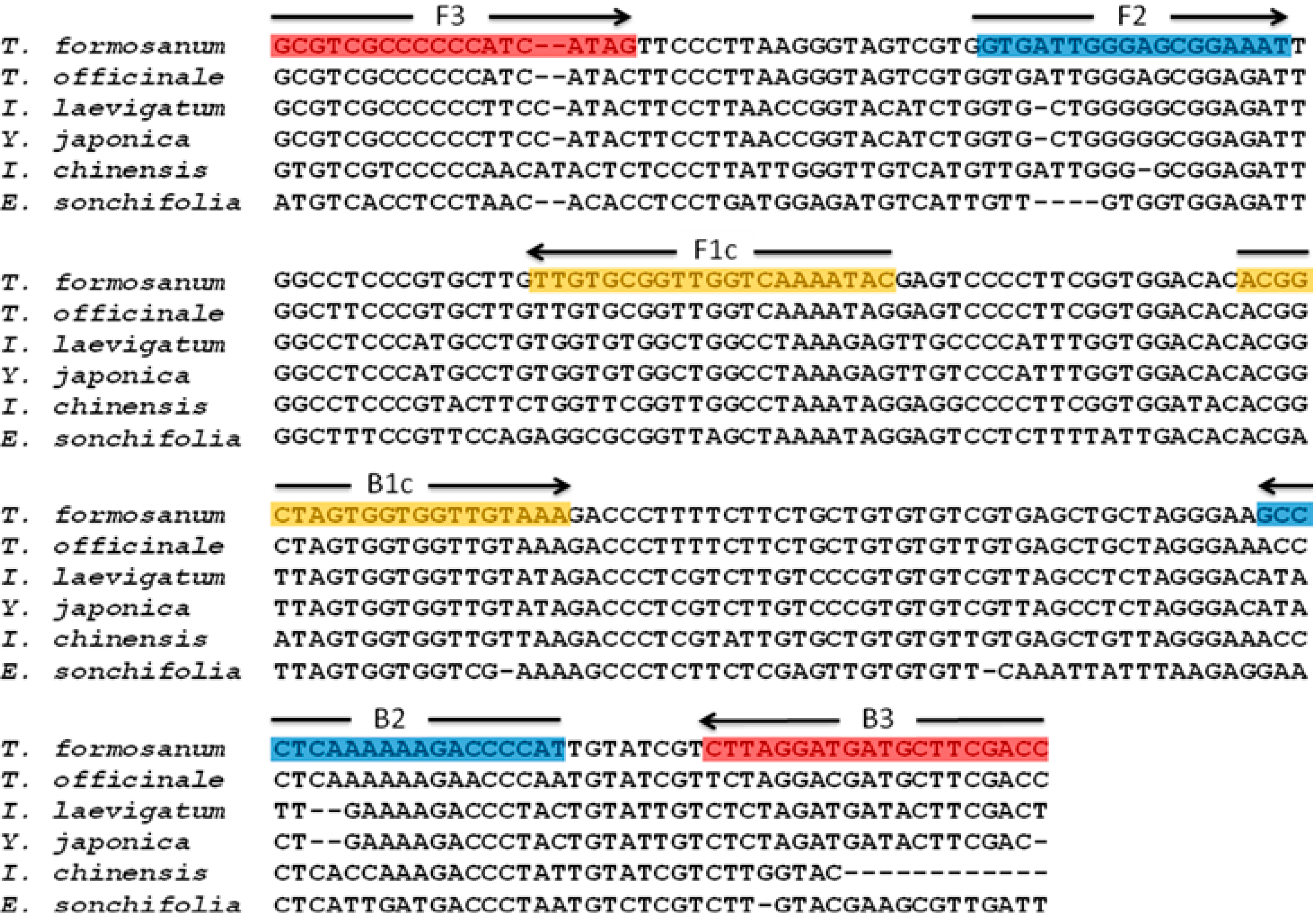
2.1. Analysis of ITS2 Sequence Alignment


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Type | Length (bp) | Sequence (5'-3') |
|---|---|---|---|
| F3 | F3 | 18 | GCGTCGCCCCCATCATAG |
| B3 | B3 | 20 | GGTCGAAGCATCATCCTAAG |
| FIP | F2 + F1c | 38 | GTATTTTGACCAACCGCAC
AAGTGATTGGGAGCGGAAT |
| BIP | B2 + B1c | 41 | ACGGCTAGTGGTGGTTGTAAAA
TGGGGTCTTTTTTGAGGGC |
2.2. Development of LAMP Primers for Authentication of T. formosanum

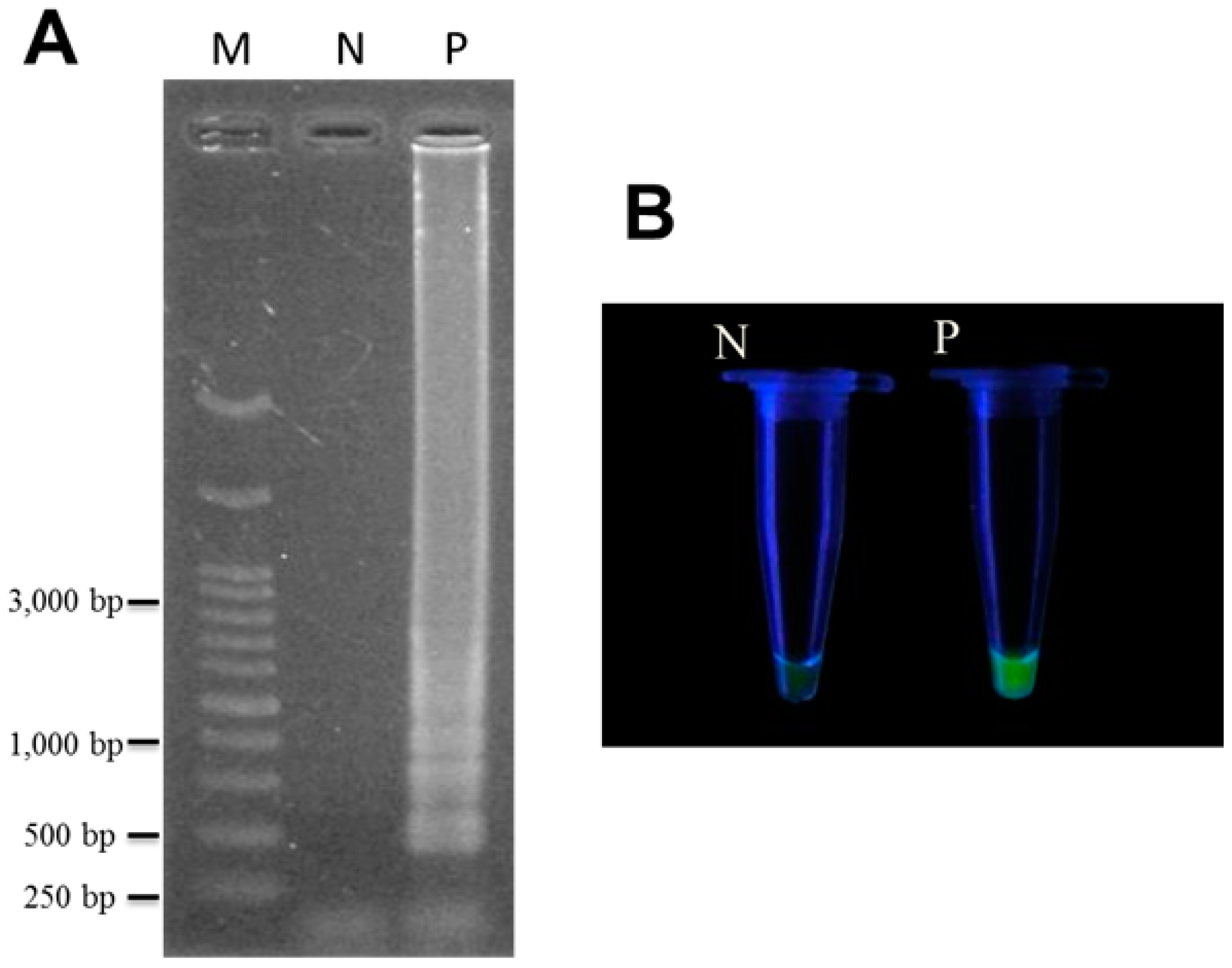
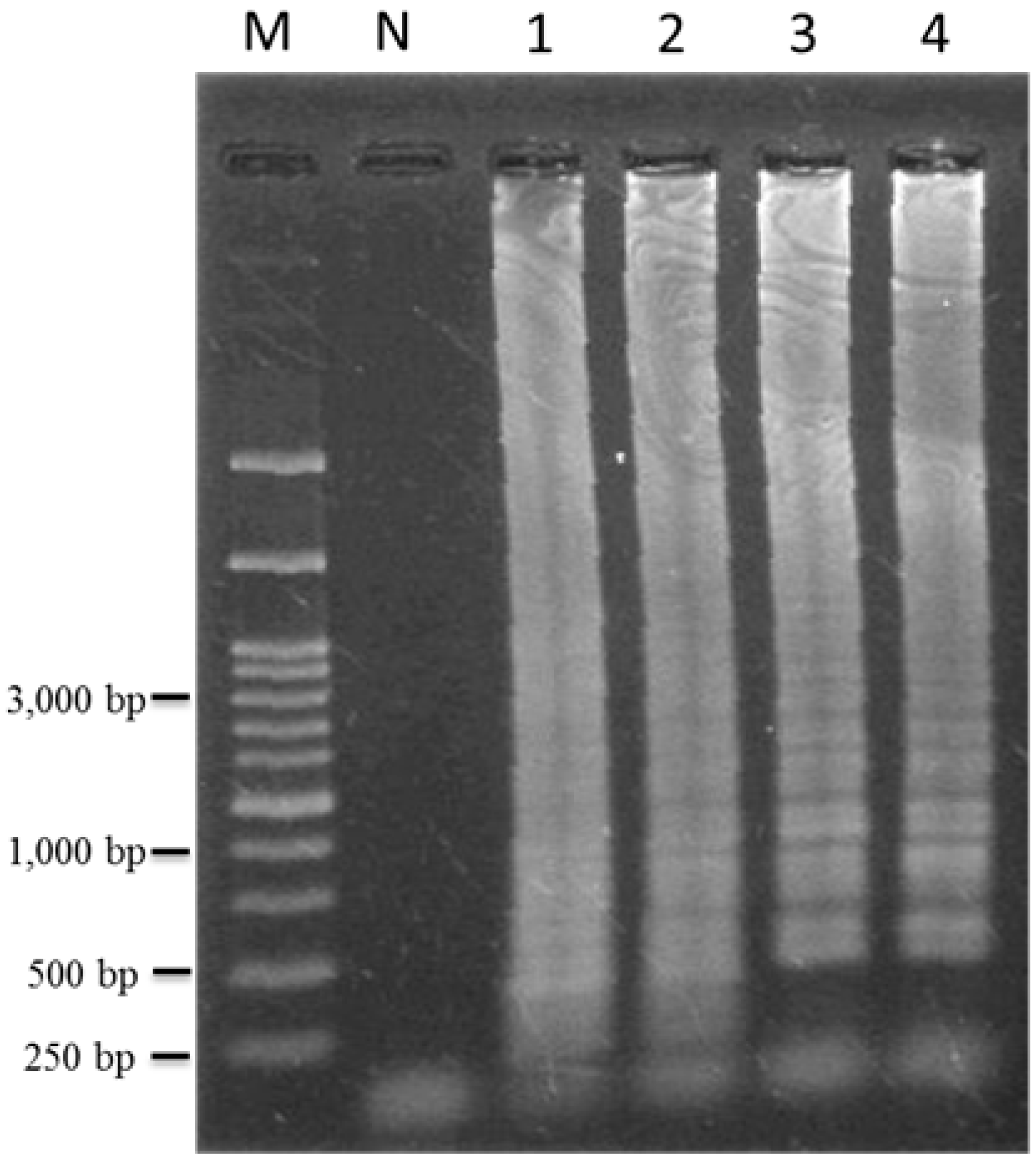
2.3. Specificity and Sensitivity of LAMP for Authentication of T. formosanum
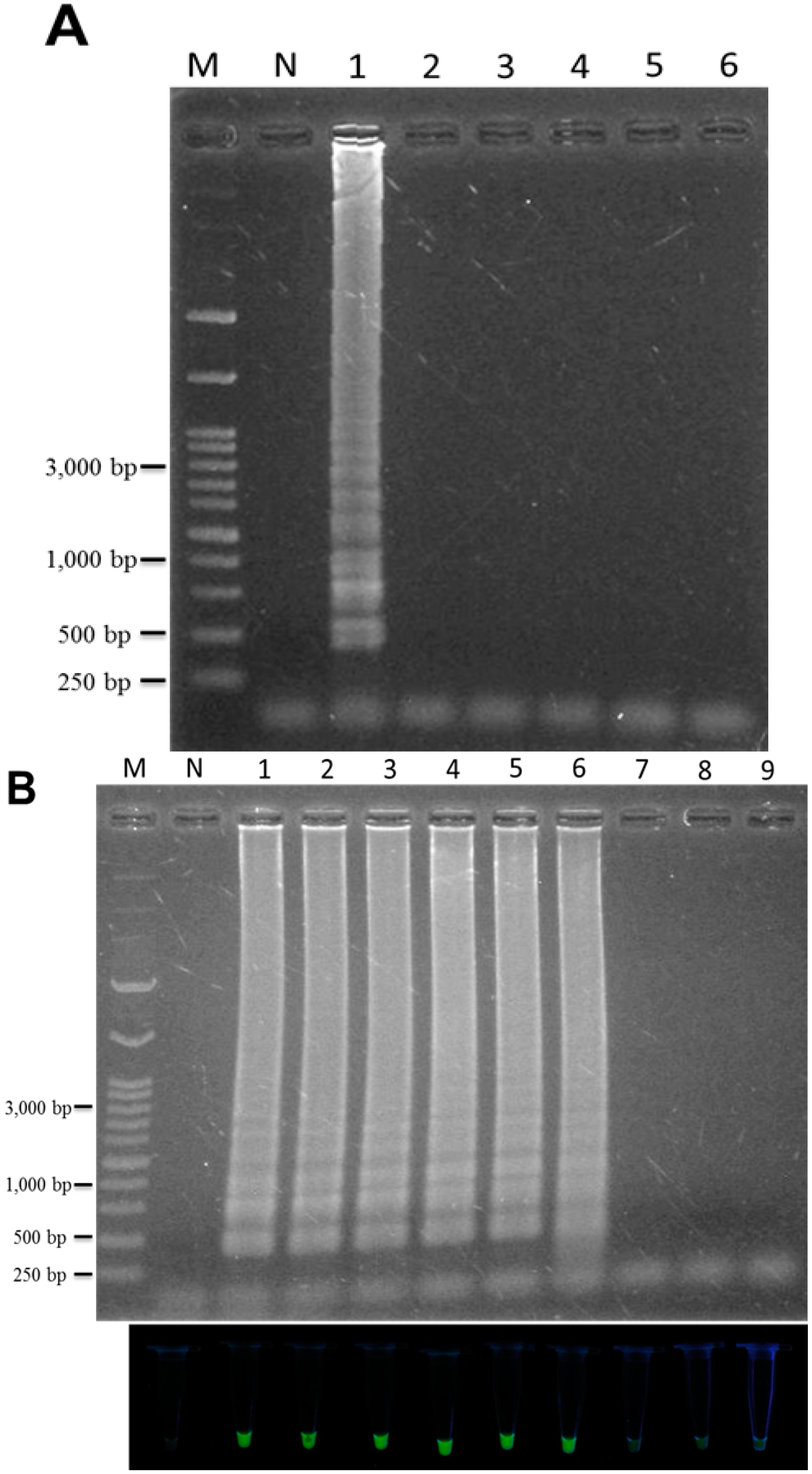
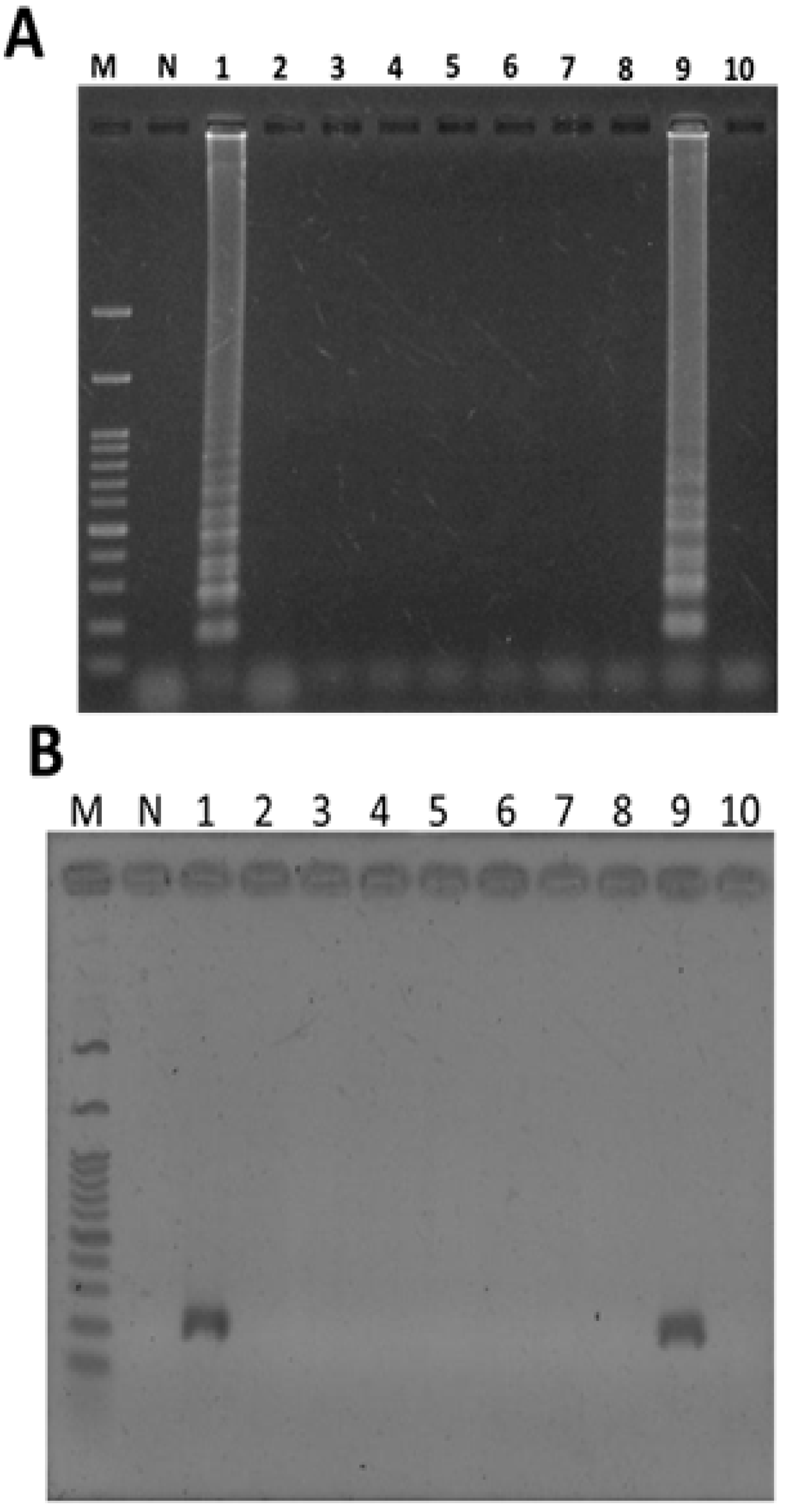
2.4. Identification of T. formosanum within a Mixed Sample



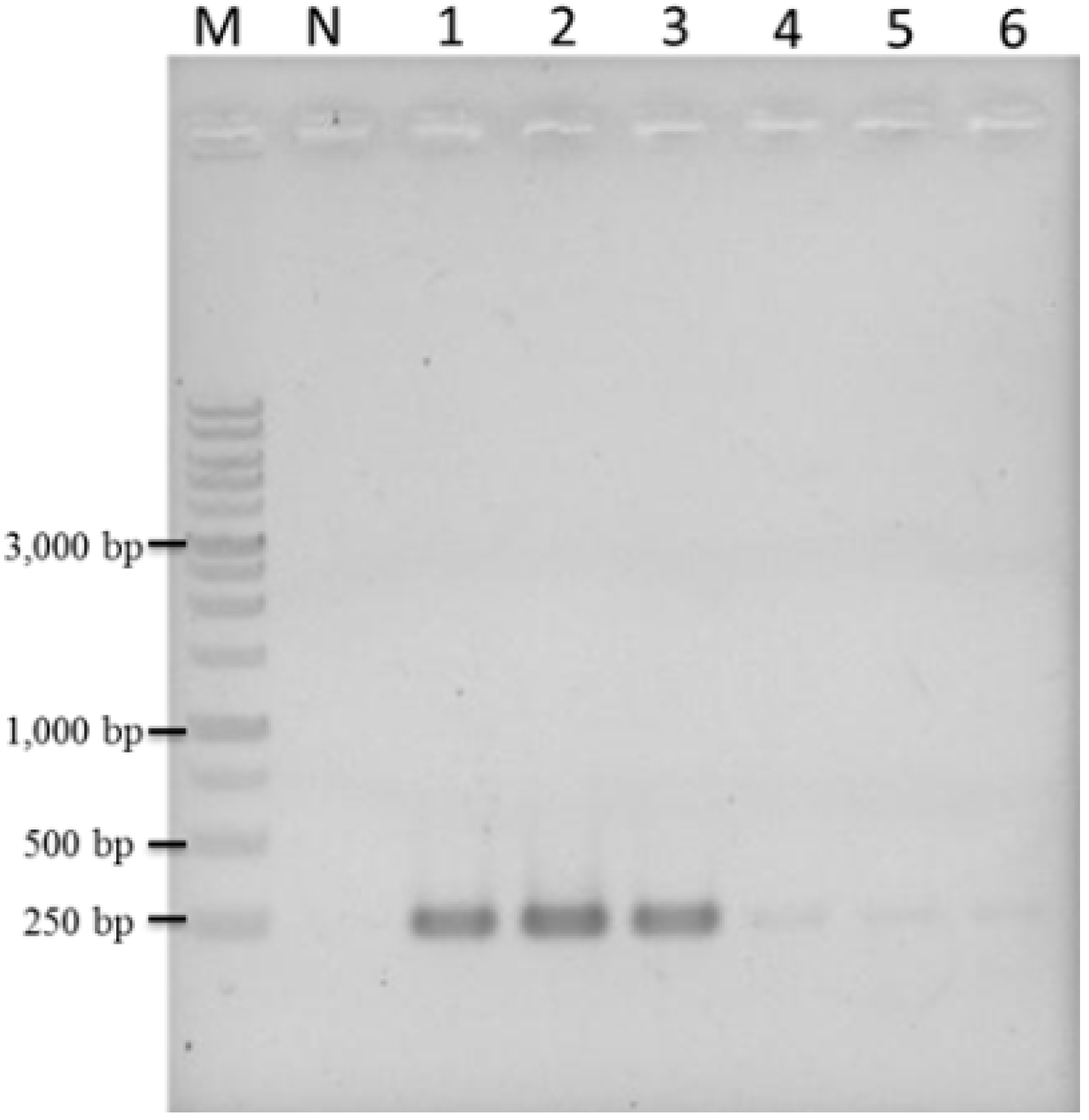
2.5. Application of LAMP to the Authentication of T. formosanum from an Herbal Market

3. Discussion
4. Experimental Section
4.1. Plant Samples
4.2. DNA Extraction
4.3. ITS Sequence Alignment
4.4. Design of LAMP Primers
4.5. LAMP Reaction
4.6. Detection of LAMP Product
4.7. PCR
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28. [Google Scholar] [CrossRef] [PubMed]
- Nagamine, K.; Hase, T.; Notomi, T. Accelerated reaction by loop-mediated isothermal amplification using loop primers. Mol. Cell. Probes 2002, 16, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Salih, D.A.; Liu, Z.; Bakheit, M.A.; Ali, A.M.; Hussein, A.M.E.I.; Unger, H.; Viljoen, G.; Seitzer, U.; Ahmed, J.S. Development and evaluation of a loop-mediated isothermal amplification method for diagnosis of tropical theileriosis. Transbound. Emerg. Dis. 2008, 55, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.M.; Guo, Y.C.; Zhang, X.E.; Zhou, Y.F.; Zhang, Z.P.; Wei, H.P.; Yang, R.F.; Wang, D.B. Loop-mediated isothermal amplification for rapid detection of Bacillus anthracis spores. Biotechnol. Lett. 2007, 29, 1939–1946. [Google Scholar] [CrossRef] [PubMed]
- Aoi, Y.; Hosogai, M.; Tsuneda, S. Real-time quantitative LAMP (loop-mediated isothermal amplification of DNA) as a simple method for monitoring ammonia-oxidizing bacteria. J. Biotechnol. 2006, 125, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, W.; Taguchi, M.; Ishibashi, M.; Nukina, M.; Misawa, N.; Inoue, K. Development of a loop-mediated isothermal amplification assay for sensitive and rapid detection Camylobacter fetus. Vet. Microbiol. 2009, 136, 393–396. [Google Scholar] [CrossRef] [PubMed]
- Shivappa, R.B.; Savan, R.; Kono, T.; Sakai, M.; Emmenegger, E.; Kurath, G.; Levine, J.F. Detection of spring viraemia of carp virus (SVCV) by loop-mediated isothermal amplification (LAMP) in koi carp, Cyprinus carpio L. J. Fish Dis. 2008, 31, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Kurosaki, Y.; Takada, A.; Ebihara, H.; Grolla, A.; Kamo, N.; Feldmann, H.; Kawaoka, Y.; Yasuda, J. Rapid and simple detection of Ebola virus by reverse transcription-loop-mediated isothermal amplification. J. Virol. Methods 2007, 141, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Woźniakowski, G.; Samorek-Salamonowicz, E.; Kozdruń, W. Rapid detection of Marek’s disease virus in feather follicles by loop-mediated amplification. Avian Dis. 2011, 55, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Ihira, M.; Ohta, A.; Sugata, K.; Suga, S.; Asano, Y.; Yoshikawa, T. Loop-mediated isothermal amplification for discriminating between human herpesvirus 6 A and B. J. Virol. Methods 2008, 154, 223–225. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; la Mura, M.; Allnutt, T.R.; Powell, W. Detection of genetically modified organisms (GMOs) using isothermal amplification of target DNA sequences. BMC Biotechnol. 2009, 9. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; la Mura, M.; Allnutt, T.; Powell, W.; Greenland, A. Isothermal Amplification of genetically modified DNA sequences directly from plant tissues lowers the barriers to high-throughput and field-based genotyping. J. Agric. Food Chem. 2009, 57, 9400–9402. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Luo, Y.; Tao, R.; He, R.; Jiang, K.; Wang, B.; Wang, L. Sensitive and rapid detection of genetic modified soybean (Roundup Ready) by loop-mediated isothermal amplification. Biosci. Biotechnol. Biochem. 2009, 73, 2365–2369. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Nagumo, S. Rapid identification of Curcuma longa and C. aromatica by LAMP. Biol. Pharm. Bull. 2007, 30, 2229–2230. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Omatsu, K.; Nagumo, S. Rapid detection of Panax ginseng by loop-mediated isothermal amplification and its application to authentication of ginseng. Biol. Pharm. Bull. 2008, 31, 1806–1808. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Fujimoto, T.; Aragane, M.; Yasuda, I.; Nagumo, S. Rapid and sensitive detection of Lophophora williamsii by loop-mediated isothermal amplification. Biol. Pharm. Bull. 2009, 32, 887–891. [Google Scholar] [CrossRef] [PubMed]
- Li, G.X. Pharmacology, Toxicity and Clinic of Traditional Chinese Medicine; Tianjin Science and Technique Translation Publishing House: Tianjin, China, 1992; pp. 207–208. [Google Scholar]
- Ou, M.; Li, Y.W. The Traditional Chinese Drug and Its Usage; Hai Feng Press: Hong Kong, China, 1994; pp. 49–51. [Google Scholar]
- Zhao, L.; Yang, Y.; Lin, J.D. Economic values of Dandelion. Liaoning Agric. Sci. 2006, 6, 33–35. [Google Scholar]
- Cao, H.; But, P.P.; Shaw, P. Identification of herbal Taraxaci and its adulterants in Hong Kong market by DNA fingerprinting with random primed PCR. Chin. J. Chin. Mater. Med. 1997, 22, 197–200. [Google Scholar]
- Tong, C.F.; He, Y.L.; Zhang, X.Z.; Cai, H.Y.; Zhang, Y.X. Investigation of adulteration and substitution species of Chinese crude drugs in Taiwan. Chin. Med. Coll. J. 1999, 8, 35–46. [Google Scholar]
- Yuan, C.C. Textual research of material medica Taraxacum mongolicum and varietal identification. Chin. Wild Plant Res. 2001, 20, 6–8. [Google Scholar]
- Chiang, Y.C.; Chen, M.D.; Lai, G.H.; Chen, H.J.; Chao, J.; Chang, W.T.; Lin, M.K.; Chang, Y.S.; Chou, Y.M.; Lee, M.S.; et al. Rapid identification of the medicinal plant Taraxacum formosanum and distinguishing of this plant from its adulterants by ribosomal DNA internal transcribed spacer (ITS) based DNA barcode. Afr. J. Biotechnol. 2011, 10, 4838–4843. [Google Scholar]
- Zhu, S.; Fushimi, H.; Cai, S.; Komatsu, K. Species identification from Ginseng drugs by multiplex amplification refractory mutation system (MARMS). Planta Med. 2004, 70, 189–192. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, K.; Minami, M.; Kawahara, K.; Nakamura, I.; Shibata, T. Discrimination among three species of medicinal Scutellaria plants using RAPD markers. Planta Med. 2000, 66, 270–272. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, V.; Sharma, K.; Gupta, S.; Kumar, R.; Prasad, M. Identification of sex in Simmondsia Chinensis (Jojoba) using PADP markers. Plant Biotechnol. Rep. 2007, 1, 207–210. [Google Scholar] [CrossRef]
- Chaseb, M.W.; Salamin, N.; Wilkison, M.; Dunwell, J.M.; Keasanakurthi, R.P.; Haidar, N.; Savolainen, V. Land plants and DNA barcodes: Short-term and long-tern goals. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 1889–1895. [Google Scholar] [CrossRef] [PubMed]
- Hon, C.C.; Chow, Y.C.; Zeng, F.Y.; Leung, F.C. Genetic authentication of ginseng and other traditional Chinese medicine. Acta Pharmacol. Sin. 2003, 24, 841–846. [Google Scholar] [PubMed]
- Pang, X.; Song, J.; Zhu, Y.; Xie, C.; Chen, S. Using DNA barcoding to identify species with Euphorbiaceae. Planta Med. 2010, 76, 1784–1786. [Google Scholar] [CrossRef] [PubMed]
- Kress, W.J.; Wurdack, K.J.; Zimmer, E.A.; Weigt, L.A.; Janzen, D.H. Use of DNA barcodes to identify flowering plants. Proc. Natl. Acad. Sci. USA 2005, 102, 8369–8374. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.I.; Lin, L.C.; Kao, C.L.; Chiang, M.Y. PCR-RFLP marker of ribosomal DNA used in detection of adulterant species of Taraxacum mongolicum. Crop Environ. Bioinform. 2007, 4, 285–296. [Google Scholar]
- Fukuta, S.; Mizukami, Y.; Ishida, A.; Kanbe, M. Development of loop-mediated isothermal amplification (LAMP)-based SNP markers for shelf-life in melon (Cucumis melo L.). J. Appl. Genet. 2006, 47, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.H.; Lai, G.H.; Lee, M.S.; Lin, W.H.; Lien, Y.Y.; Hsueh, S.C.; Kao, J.Y.; Chang, W.T.; Lu, T.C.; Lin, W.N.; et al. Development and evaluation of a loop-mediated isothermal amplification assay for rapid detection of chicken anaemia virus. J. Appl. Microbiol. 2010, 108, 917–924. [Google Scholar] [CrossRef]
- Yoshikawa, T.; Matsuo, T.; Kawamura, Y.; Ohashi, M.; Yonekawa, T.; Kanda, H.; Notomi, T.; Ihira, M. Direct detection of human herpesvirus 6B by the LAMP method using newly developed dry-reagents. J. Virol. Methods 2014, 201, 65–67. [Google Scholar] [CrossRef] [PubMed]
- Woźniakowski, G.; Samorek-Salamonowicz, E. Direct detection of Marek’s disease virus in poultry dust by loop-mediated isothermal amplification. Arch. Virol. 2014, 159, 3083–3087. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.S. Authentication on Easy-Confused Chinese Medicinal Material. Presented at the Symposium of Identification on Easy-Confused Chinese Medicinal Material for TCM Pratitioners; Committee of Chinese Medicine and Pharmacy & Department of Health, Executive Yuan: Taichung, Taiwan, 2009; pp. 85–86.
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, G.-H.; Chao, J.; Lin, M.-K.; Chang, W.-T.; Peng, W.-H.; Sun, F.-C.; Lee, M.-S.; Lee, M.-S. Rapid and Sensitive Identification of the Herbal Tea Ingredient Taraxacum formosanum Using Loop-Mediated Isothermal Amplification. Int. J. Mol. Sci. 2015, 16, 1562-1575. https://doi.org/10.3390/ijms16011562
Lai G-H, Chao J, Lin M-K, Chang W-T, Peng W-H, Sun F-C, Lee M-S, Lee M-S. Rapid and Sensitive Identification of the Herbal Tea Ingredient Taraxacum formosanum Using Loop-Mediated Isothermal Amplification. International Journal of Molecular Sciences. 2015; 16(1):1562-1575. https://doi.org/10.3390/ijms16011562
Chicago/Turabian StyleLai, Guan-Hua, Jung Chao, Ming-Kuem Lin, Wen-Te Chang, Wen-Huang Peng, Fang-Chun Sun, Meng-Shiunn Lee, and Meng-Shiou Lee. 2015. "Rapid and Sensitive Identification of the Herbal Tea Ingredient Taraxacum formosanum Using Loop-Mediated Isothermal Amplification" International Journal of Molecular Sciences 16, no. 1: 1562-1575. https://doi.org/10.3390/ijms16011562