
MiR-21 Protected Cardiomyocytes against Doxorubicin-Induced Apoptosis by Targeting BTG2

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Effects of DOX on Cardiac Functions of Mice
2.2. The Effect of DOX Treatment on the Expression of miR-21 in Mice Myocardium and H9C2 Cells


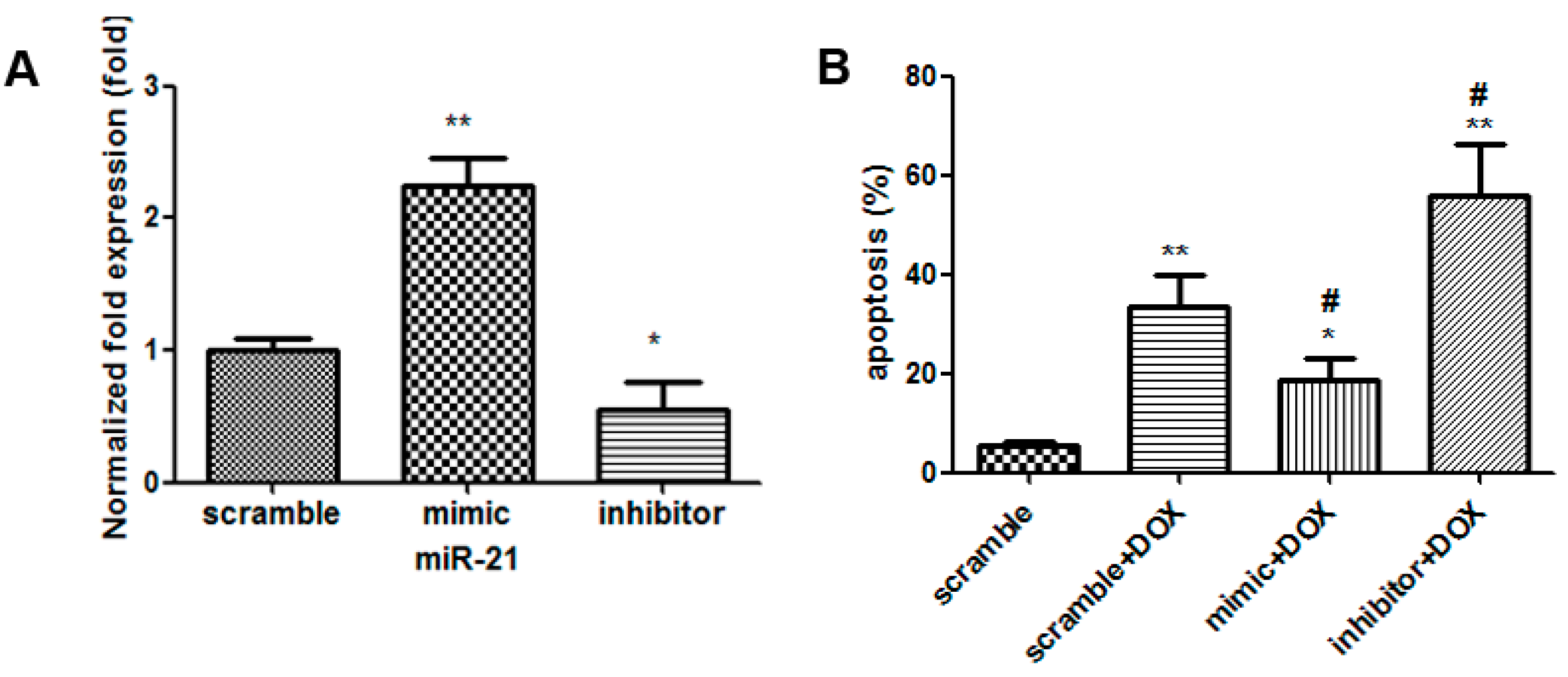
2.3. The Effects of miR-21 Mimics and Inhibitors on the Injury Mediated by DOX in H9C2 Cells


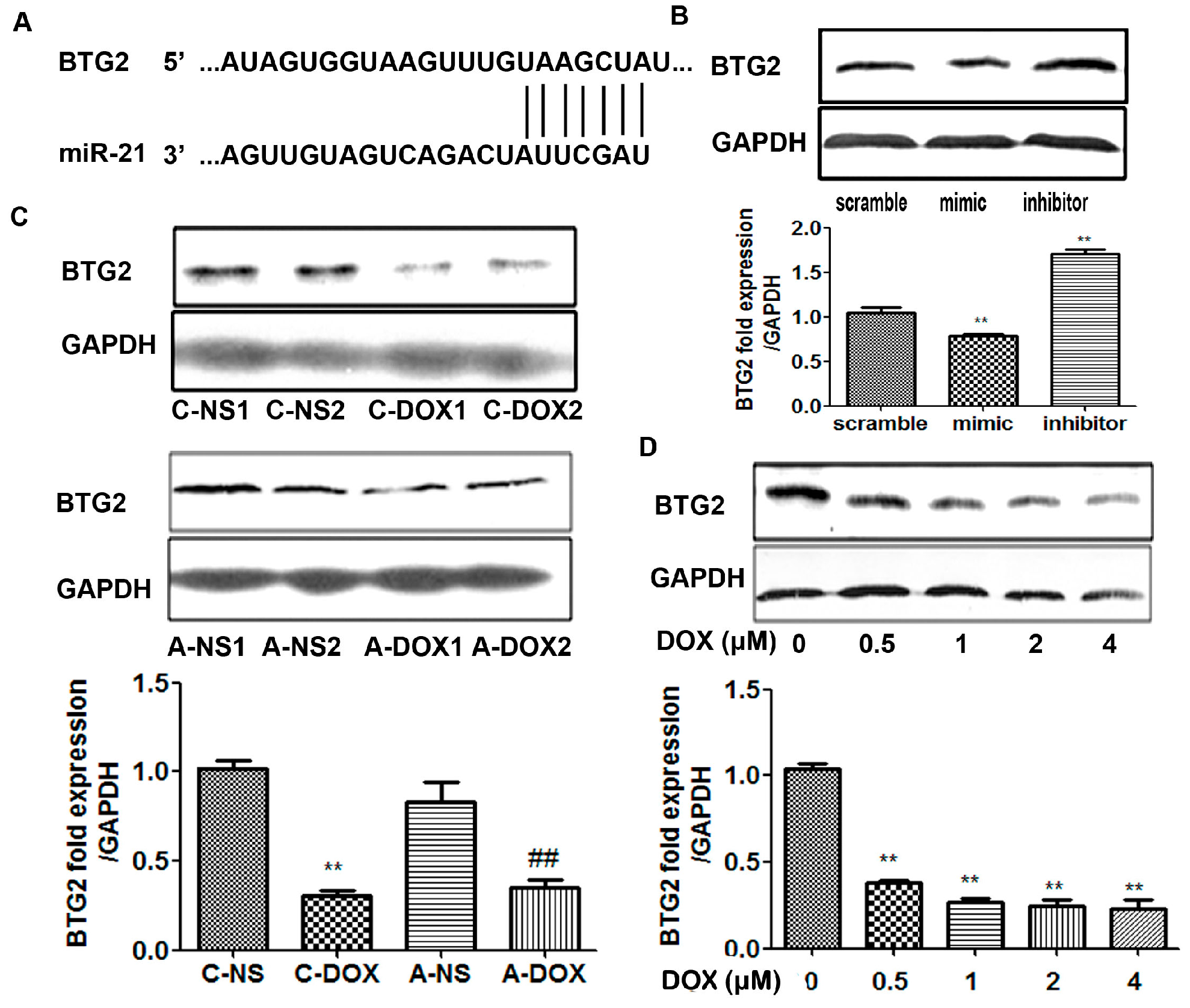
2.4. miR-21Regulatedthe Expression of BTG-2 in H9C2 Cells


3. Discussion
4. Materials and Methods
4.1. Animals and Treatment
4.2. Cardiac Haemodynamic Measurements
4.3. Cell Culture
4.4. Transfection Experiments
4.5. RNA Isolation and Quantitative Real-Time PCR Analysis
4.6. Protein Preparation and Western Blotting
4.7. Assays of Cell Viability and Activities of Lactate Dehydrogenase
4.8. Apoptosis Assessment by TUNEL Assay
4.9. Flow Cytometry
4.10. Computational Prediction of miRNA Target Genes
4.11. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Minotti, G.; Menna, P.; Salvatorelli, E.; Cairo, G.; Gianni, L. Anthracyclines: Molecular advances and pharmacologic developments in antitumor activity and cardiotoxicity. Pharmacol. Rev. 2004, 56, 185–229. [Google Scholar] [CrossRef] [PubMed]
- Lefrak, E.A.; Pitha, J.; Rosenheim, S.; Gottlieb, J.A. A clinicopathologic analysis of adriamycin cardiotoxicity. Cancer 1973, 32, 302–314. [Google Scholar] [CrossRef]
- Buja, L.M.; Ferrans, V.J.; Mayer, R.J.; Roberts, W.C.; Henderson, E.S. Cardiac ultrastructural changes induced by daunorubicin therapy. Cancer 1973, 32, 771–788. [Google Scholar] [CrossRef]
- Roca-Alonso, L.; Pellegrino, L.; Castellano, L.; Stebbing, J. Breast cancer treatment and adverse cardiac events: What are the molecular mechanisms? Cardiology 2012, 122, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Ueda, Y.; Juan, Y.; Katsuda, S.; Takahashi, H.; Koh, E. Fas-mediated apoptosis in adriamycin-induced cardiomyopathy in rats: In vivo study. Circulation 2000, 102, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.W.; Shi, J.; Li, Y.J.; Wei, L. Cardiomyocyte death in doxorubicin-induced cardiotoxicity. Arch. Immunol. Ther. Exp. 2009, 57, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Chen, X.; Cao, Y. New role of microRNA: Carcinogenesis and clinical application in cancer. Acta Biochim. Biophys. Sin. 2011, 43, 831–839. [Google Scholar] [CrossRef] [PubMed]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Thum, T.; Catalucci, D.; Bauersachs, J. MicroRNAs: Novel regulators in cardiac development and disease. Cardiovasc. Res. 2008, 79, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Jazbutyte, V.; Thum, T. MicroRNA-21: From cancer to cardiovascular disease. Curr. Drug Targets 2010, 11, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Thum, T.; Gross, C.; Fiedler, J.; Fischer, T.; Kissler, S.; Bussen, M.; Galuppo, P.; Just, S.; Rottbauer, W.; Frantz, S.; et al. MicroRNA-21 contributes to myocardial disease by stimulating map kinase signalling in fibroblasts. Nature 2008, 456, 980–984. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Khanna, S.; Hussain, S.R.; Biswas, S.; Azad, A.; Rink, C.; Gnyawali, S.; Shilo, S.; Nuovo, G.J.; Sen, C.K. MicroRNA expression in response to murine myocardial infarction: miR-21 regulates fibroblast metalloprotease-2 via phosphatase and tensin homologue. Cardiovasc. Res. 2009, 82, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Cheng, Y.; Yang, J.; Li, J.; Liu, X.; Wang, X.; Wang, D.; Krall, T.J.; Delphin, E.S.; Zhang, C. MicroRNA expression signature and the role of microRNA-21 in the early phase of acute myocardial infarction. J. Biol. Chem. 2009, 284, 29514–29525. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Yang, K.; Li, A. MicroRNA-21 protects against ischemia-reperfusion and hypoxia-reperfusion-induced cardiocyte apoptosis via the phosphatase and tensin homolog/akt-dependent mechanism. Mol. Med. Rep. 2014, 9, 2213–2220. [Google Scholar] [CrossRef] [PubMed]
- Mao, B.; Zhang, Z.; Wang, G. BTG2: A rising star of tumor suppressors (review). Int. J. Oncol. 2015, 46, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Tang, Z.L.; Li, K.; Liu, B.; Yu, M.; Zhao, S.H. Molecular characterization of the BTG2 and BTG3 genes in fetal muscle development of pigs. Gene 2007, 403, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.W.; Park, T.J.; Kim, H.S.; Lim, I.K. Signals regulating necrosis of cardiomyoblast by BTG2/TIS21/PC3 via activation of GSK3β and opening of mitochondrial permeability transition pore in response to H2O2. Biochem. Biophys. Res. Commun. 2013, 434, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Safra, T.; Muggia, F.; Jeffers, S.; Tsao-Wei, D.D.; Groshen, S.; Lyass, O.; Henderson, R.; Berry, G.; Gabizon, A. Pegylated liposomal doxorubicin (doxil): Reduced clinical cardiotoxicity in patients reaching or exceeding cumulative doses of 500 mg/m2. Ann. Oncol. 2000, 11, 1029–1033. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.C.; Zhou, X.; Wang, X.; Song, G.; Qian, J.; Nicolaou, P.; Chen, G.; Ren, X.; Kranias, E.G. Heat shock protein 20 interacting with phosphorylated akt reduces doxorubicin-triggered oxidative stress and cardiotoxicity. Circ. Res. 2008, 103, 1270–1279. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Maity, B.; Huang, J.; Gao, Z.; Stewart, A.; Weiss, R.M.; Anderson, M.E.; Fisher, R.A. G-protein inactivator RGS6 mediates myocardial cell apoptosis and cardiomyopathy caused by doxorubicin. Cancer Res. 2013, 73, 1662–1667. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, K.; Kanagasabai, R.; Druhan, L.J.; Ilangovan, G. Heat shock protein 25-enriched plasma transfusion preconditions the heart against doxorubicin-induced dilated cardiomyopathy in mice. J. Pharmacol. Exp. Ther. 2012, 341, 829–839. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Yang, Y.B.; Zhang, X.H.; Yu, X.L.; Wang, Z.B.; Cheng, X.C. MicroRNA-21 gene and cancer. Med. Oncol. 2013, 30, 376. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Han, L.; Wei, J.; Shi, Z.; Pu, P.; Zhang, J.; Yuan, X.; Kang, C. Combination treatment with doxorubicin and microRNA-21 inhibitor synergistically augments anticancer activity through upregulation of tumor suppressing genes. Int. J. Oncol. 2015, 46, 1589–1600. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Kriegel, A.J.; Jiao, X.; Liu, H.; Bai, X.; Olson, J.; Liang, M.; Ding, X. Mir-21 in ischemia/reperfusion injury: A double-edged sword? Physiol. Genomics 2014, 46, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Ma, W.; Hao, B.; Hu, F.; Yan, L.; Yan, X.; Wang, Y.; Chen, Z.; Wang, Z. MicroRNA-21 promotes cardiac fibrosis and development of heart failure with preserved left ventricular ejection fraction by up-regulating BCL-2. Int. J. Clin. Exp. Pathol. 2014, 7, 565–574. [Google Scholar] [PubMed]
- Tirone, F. The gene PC3 (TIS21/BTG2), prototype member of the PC3/BTG/TOB family: Regulator in control of cell growth, differentiation, and DNA repair? J. Cell. Physiol. 2001, 187, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Wei, L.; Liu, M.; Li, J.; Zheng, Y.Q.; Gao, Y.; Li, X.R. BTG2 inhibits the proliferation, invasion, and apoptosis of MDA-MB-231 triple-negative breast cancer cells. Tumour Biol. 2013, 34, 1605–1613. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Hao, C.; Li, X.; Zhao, H.; Chen, J.; Zhou, Q. Effects of BTG2 on proliferation inhibition and anti-invasion in human lung cancer cells. Tumour Biol. 2012, 33, 1223–1230. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wu, H.; Liu, T.; Li, Y.; Wang, F.; Wan, H.; Li, X.; Tang, H. Regulation of the cell cycle gene, BTG2, by MIR-21 in human laryngeal carcinoma. Cell Res. 2009, 19, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Zhang, B.; Liang, P.; Chen, G.; Zhou, B.; Lv, C.; Tu, Z.; Xiao, X. Nucleolin protects the heart from ischaemia-reperfusion injury by up-regulating heat shock protein 32. Cardiovas. Res. 2013, 99, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Monte, E.; Mouillesseaux, K.; Chen, H.; Kimball, T.; Ren, S.; Wang, Y.; Chen, J.N.; Vondriska, T.M.; Franklin, S. Systems proteomics of cardiac chromatin identifies nucleolin as a regulator of growth and cellular plasticity in cardiomyocytes. Am. J. Physiol. Heart Circ. Physiol. 2013, 305, H1624–H1638. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Feng, Y.; Qu, S.; Wei, X.; Zhu, H.; Luo, Q.; Liu, M.; Chen, G.; Xiao, X. Resveratrol attenuates doxorubicin-induced cardiomyocyte apoptosis in mice through SIRT1-mediated deacetylation of p53. Cardiovas. Res. 2011, 90, 538–545. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tong, Z.; Jiang, B.; Wu, Y.; Liu, Y.; Li, Y.; Gao, M.; Jiang, Y.; Lv, Q.; Xiao, X. MiR-21 Protected Cardiomyocytes against Doxorubicin-Induced Apoptosis by Targeting BTG2. Int. J. Mol. Sci. 2015, 16, 14511-14525. https://doi.org/10.3390/ijms160714511
Tong Z, Jiang B, Wu Y, Liu Y, Li Y, Gao M, Jiang Y, Lv Q, Xiao X. MiR-21 Protected Cardiomyocytes against Doxorubicin-Induced Apoptosis by Targeting BTG2. International Journal of Molecular Sciences. 2015; 16(7):14511-14525. https://doi.org/10.3390/ijms160714511
Chicago/Turabian StyleTong, Zhongyi, Bimei Jiang, Yanyang Wu, Yanjuan Liu, Yuanbin Li, Min Gao, Yu Jiang, Qinglan Lv, and Xianzhong Xiao. 2015. "MiR-21 Protected Cardiomyocytes against Doxorubicin-Induced Apoptosis by Targeting BTG2" International Journal of Molecular Sciences 16, no. 7: 14511-14525. https://doi.org/10.3390/ijms160714511