
High-Level γ-Glutamyl-Hydrolase (GGH) Expression is Linked to Poor Prognosis in ERG Negative Prostate Cancer

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Technical Issues
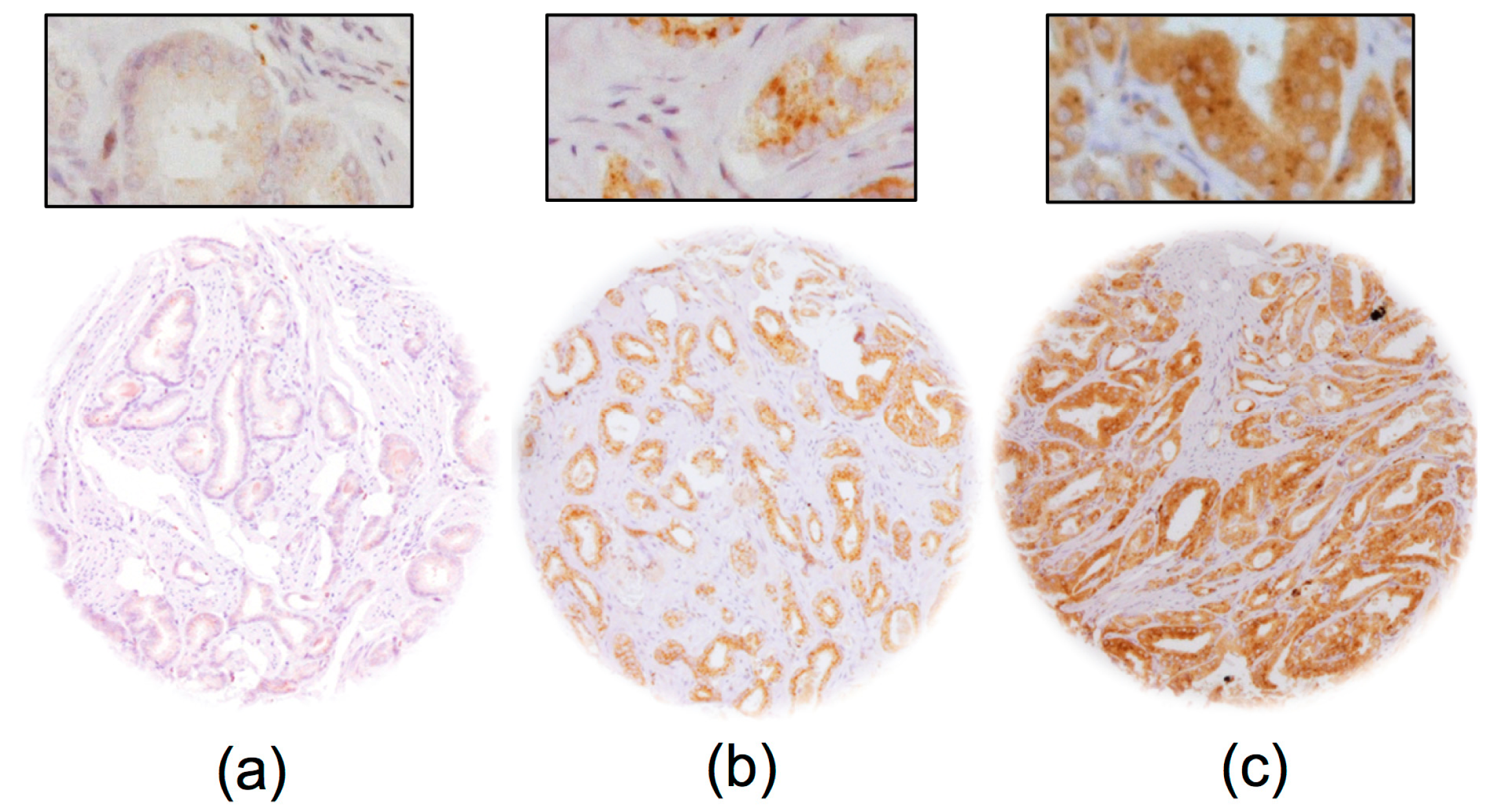
2.2. GGH Expression in Normal Prostate Tissue and Prostate Cancer
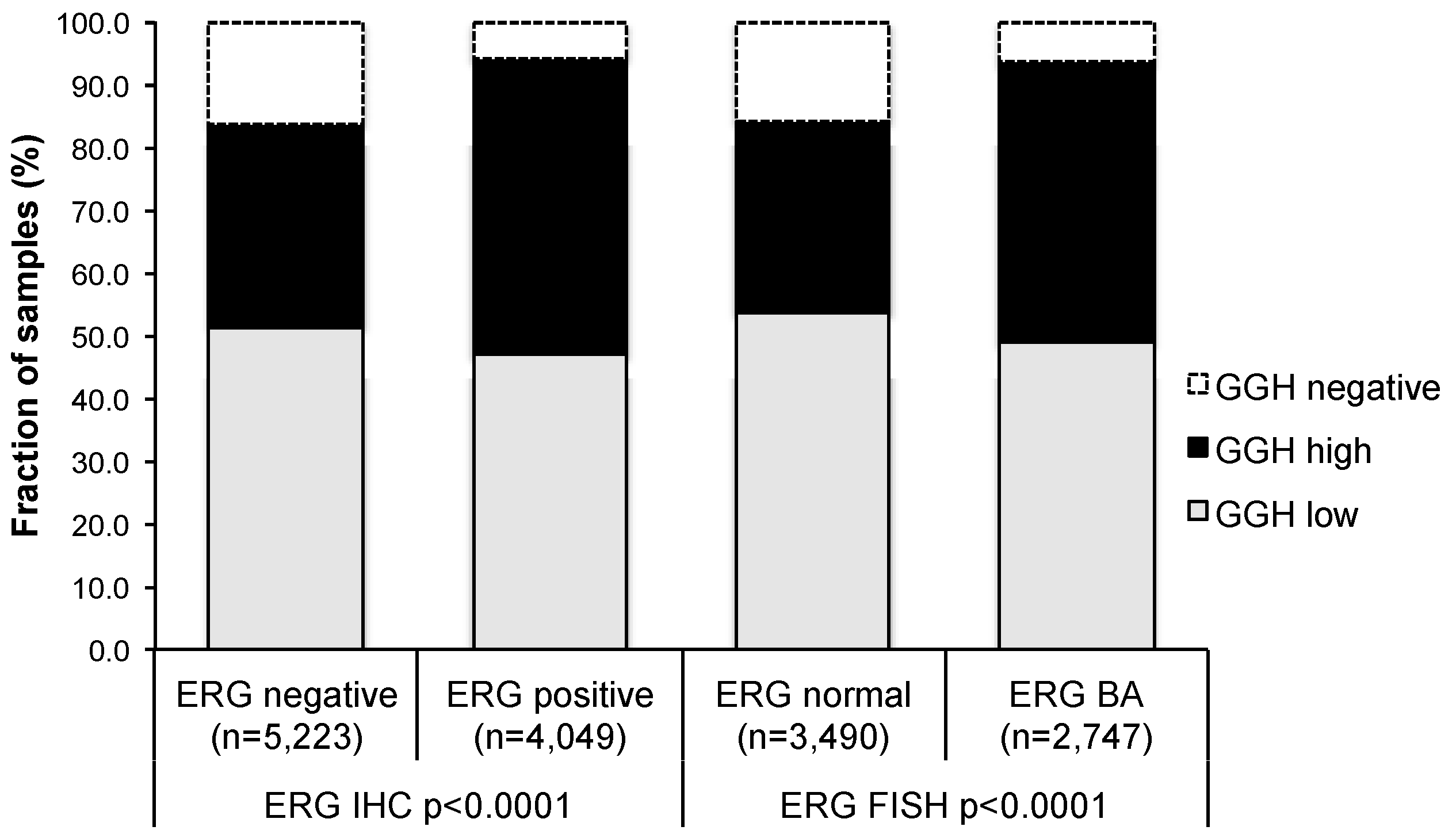
2.3. Association with TMPRSS2:ERG Fusion Status and ERG Protein Expression
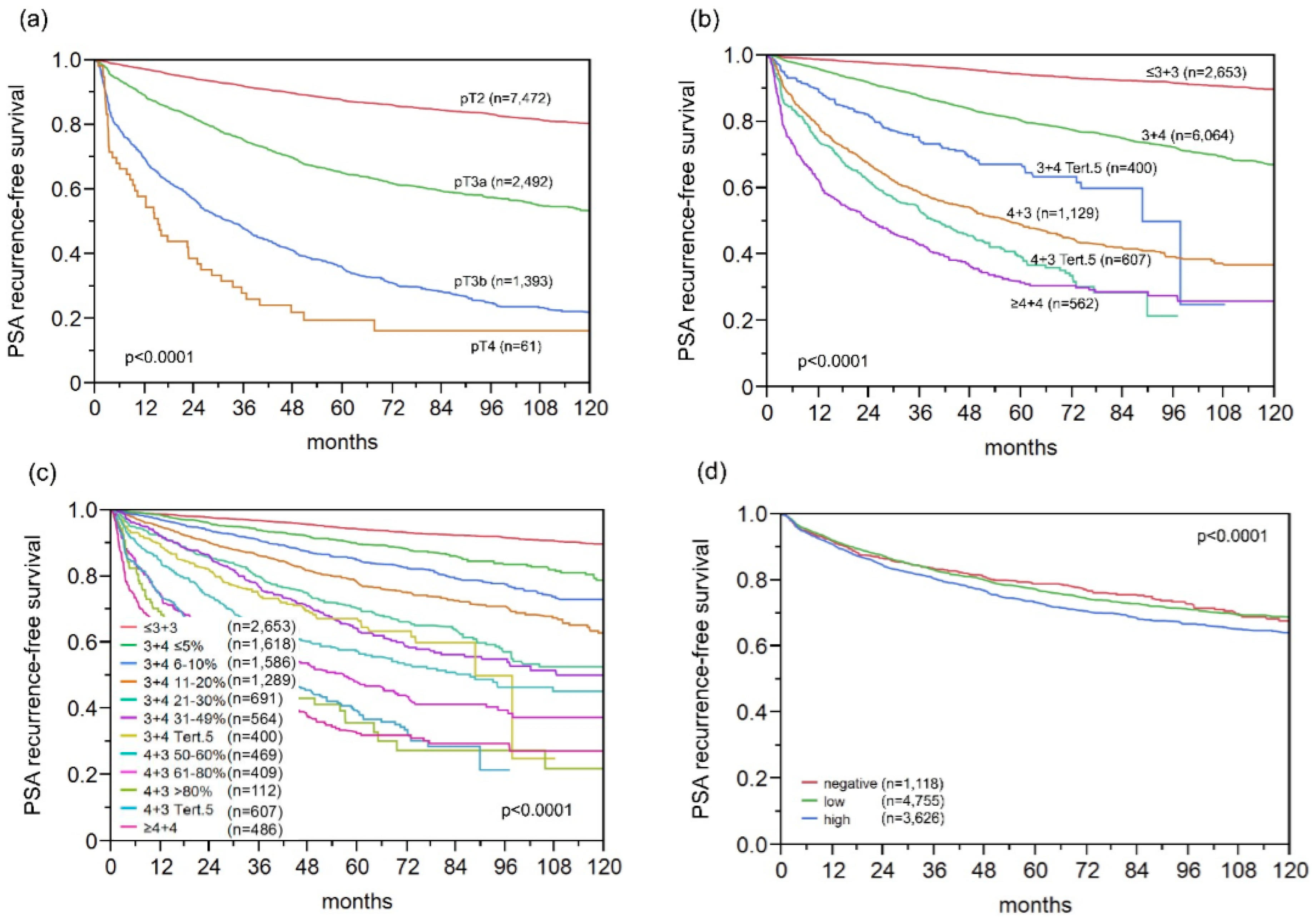
2.4. Association with Tumour Clinical Characteristics
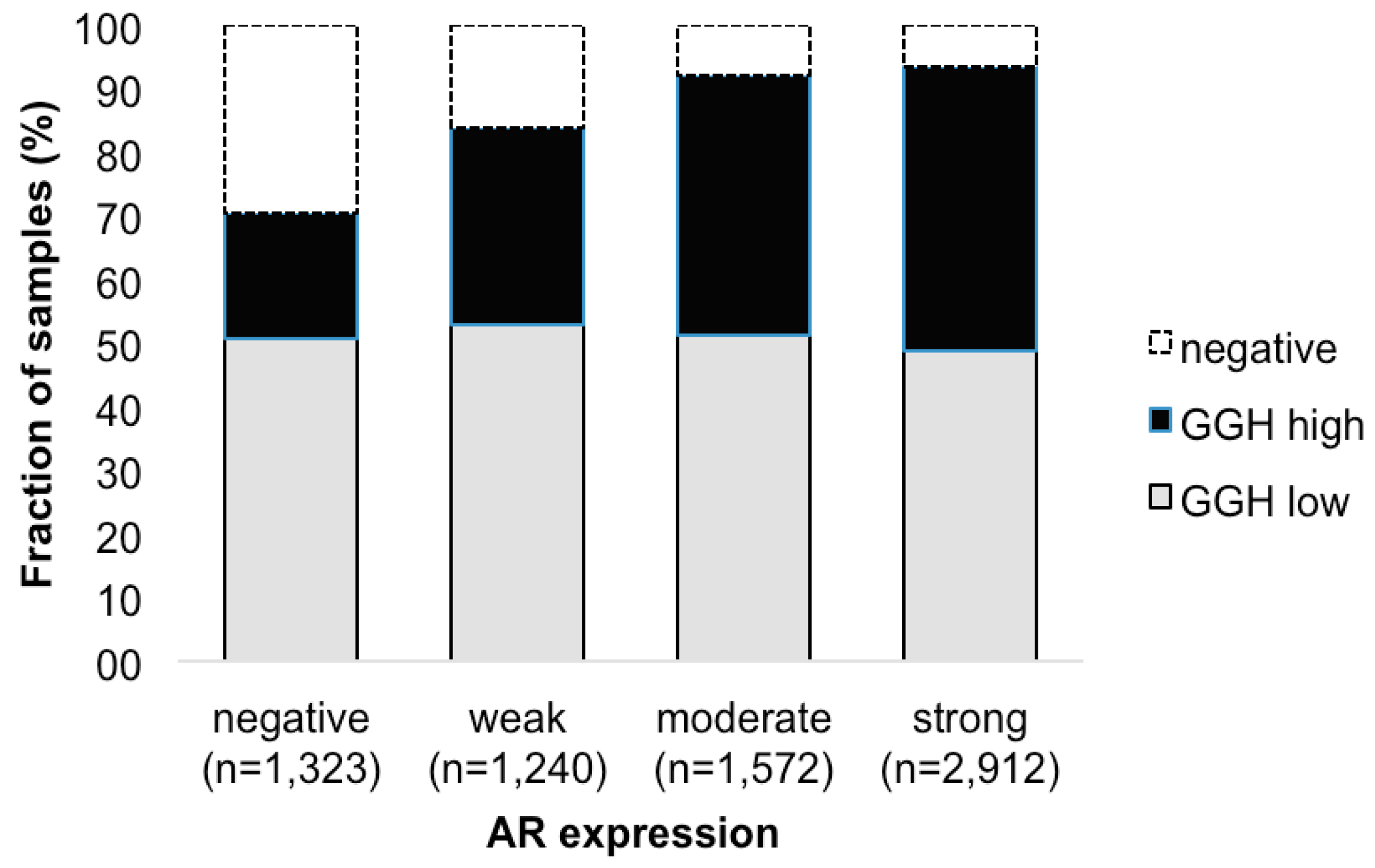
2.5. Association with Androgen Receptor
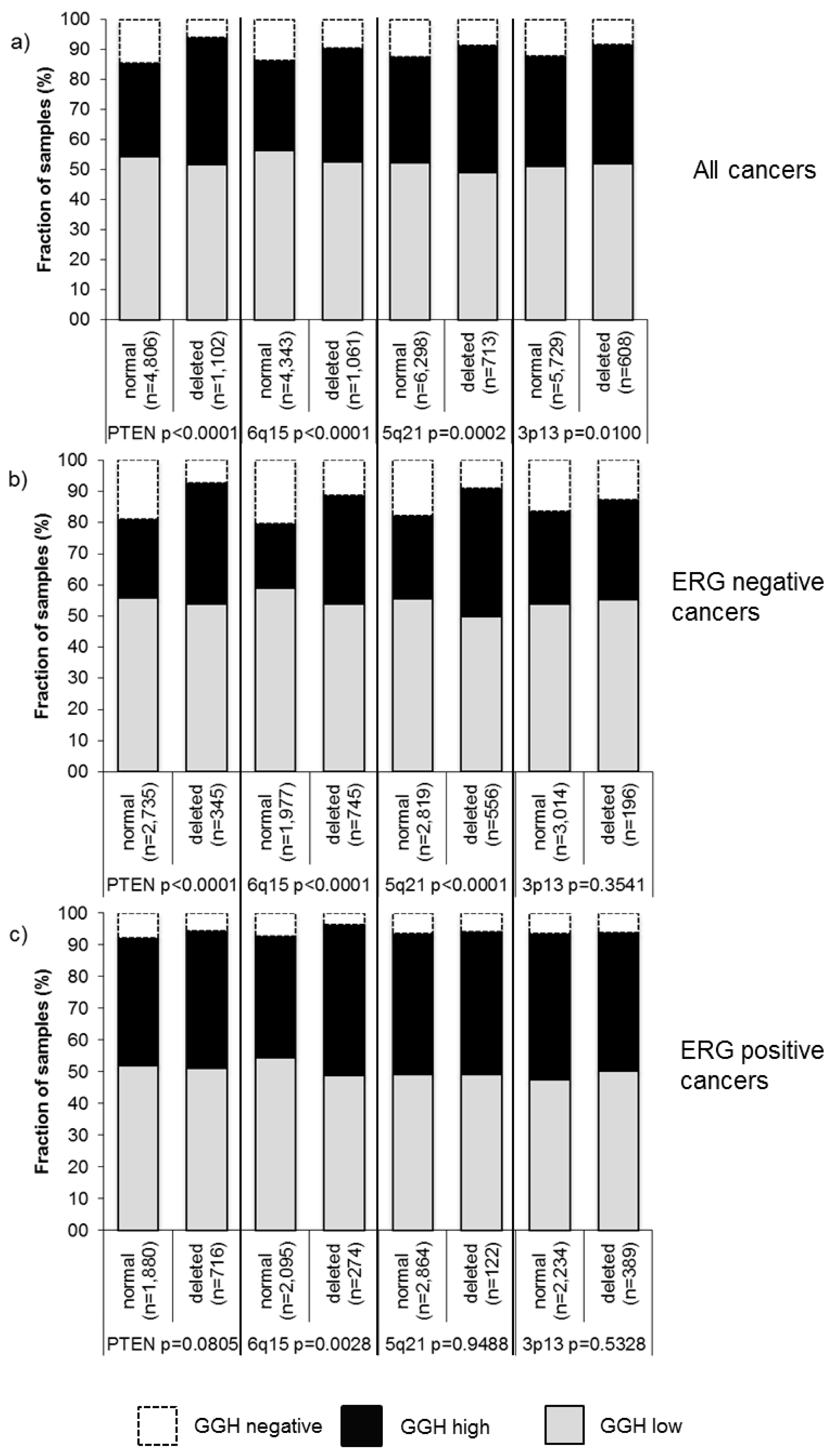
2.6. Association with Other Key Genomic Deletions
2.7. Association with Tumor Cell Proliferation
2.8. DNA Ploidy Status
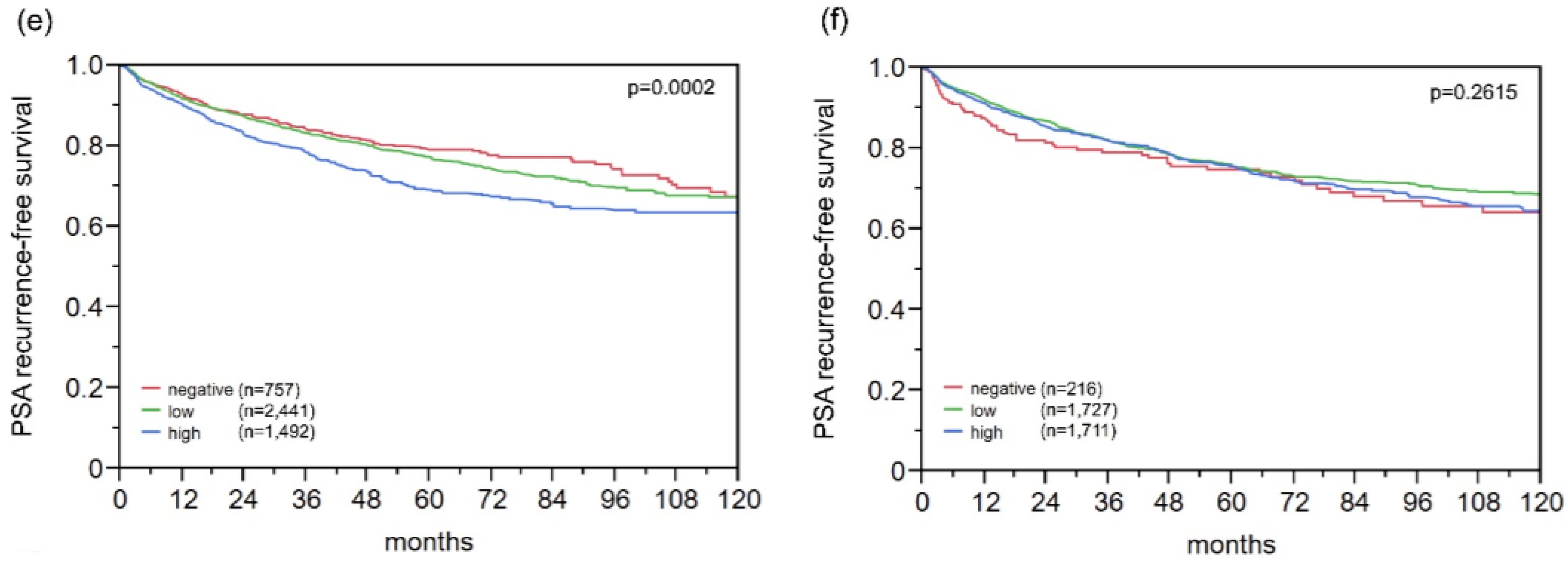
2.9. Association with PSA Recurrence
2.10. Multivariate Analysis
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Immunohistochemistry
4.3. Statistics
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AR | Androgen receptor |
| CHD1 | Chromodomain-Helicase-DNA-Binding Protein 1 |
| FISH | Fluorescence in-situ hybridization |
| FOXP1 | Forkhead box protein P1 |
| GGH | γ-Glutamyl-Hydrolase |
| IHC | Immunohistochemistry |
| Ki67LI | Ki67 labeling index |
| MAP3K | Mitogen-Activated Protein Kinase Kinase Kinase 7 |
| PSA | Prostate specific antigen |
| PTEN | Phosphatase and tensin homolog |
| RPE | Radical prostatectomy |
| TMPRSS2:ERG | Transmembrane protease, serine 2: ETS-related gene fusion |
| TMA | Tissue microarray |
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.T.; Chandler, C.J.; Halsted, C.H. Intracellular pteroylpolyglutamate hydrolase from human jejunal mucosa. Isolation and characterization. J. Biol. Chem. 1986, 261, 13551–13555. [Google Scholar] [PubMed]
- O’Connor, B.M.; Rotundo, R.F.; Nimec, Z.; McGuire, J.J.; Galivan, J. Secretion of γ-glutamyl hydrolase in vitro. Cancer Res. 1991, 51, 3874–3881. [Google Scholar] [PubMed]
- Schneider, E.; Ryan, T.J. γ-glutamyl hydrolase and drug resistance. Clin. Chim. Acta 2006, 374, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Galivan, J.; Ryan, T.; Rhee, M.; Yao, R.; Chave, K. Glutamyl hydrolase: Properties and pharmacologic impact. Semin. Oncol. 1999, 26 (Suppl. 6), 33–37. [Google Scholar] [PubMed]
- Galivan, J.; Ryan, T.J.; Chave, K.; Rhee, M.; Yao, R.; Yin, D. Glutamyl hydrolase. pharmacological role and enzymatic characterization. Pharmacol. Ther. 2000, 85, 207–215. [Google Scholar] [CrossRef]
- Fenech, M. Folate (vitamin B9) and vitamin B12 and their function in the maintenance of nuclear and mitochondrial genome integrity. Mutat. Res. 2012, 733, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Shubbar, E.; Helou, K.; Kovacs, A.; Nemes, S.; Hajizadeh, S.; Enerback, C.; Einbeigi, Z. High levels of γ-glutamyl hydrolase (GGH) are associated with poor prognosis and unfavorable clinical outcomes in invasive breast cancer. BMC Cancer 2013, 13, 47. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, K.; Ooyama, A.; Ruszkiewicz, A.; Jin, M.; Watanabe, G.; Moore, J.; Oka, T.; Iacopetta, B.; Minamoto, T. Low expression of γ-glutamyl hydrolase mRNA in primary colorectal cancer with the CpG island methylator phenotype. Br. J. Cancer 2008, 98, 1555–1561. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Q.; Cheng, C.; Crews, K.R.; Ribeiro, R.C.; Pui, C.H.; Relling, M.V.; Evans, W.E. Epigenetic regulation of human γ-glutamyl hydrolase activity in acute lymphoblastic leukemia cells. Am. J. Hum. Genet. 2006, 79, 264–274. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Varticovski, L.; Bowman, E.D.; Fukuoka, J.; Welsh, J.A.; Miura, K.; Jen, J.; Gabrielson, E.; Brambilla, E.; Travis, W.D.; et al. Identification of carboxypeptidase E and γ-glutamyl hydrolase as biomarkers for pulmonary neuroendocrine tumors by cDNA microarray. Hum. Pathol. 2004, 35, 1196–1209. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.E.; Cole, P.D.; Cho, R.C.; Ly, A.; Ishiguro, L.; Sohn, K.J.; Croxford, R.; Kamen, B.A.; Kim, Y.I. γ-glutamyl hydrolase modulation and folate influence chemosensitivity of cancer cells to 5-fluorouracil and methotrexate. Br. J. Cancer 2013, 109, 2175–2188. [Google Scholar] [CrossRef] [PubMed]
- Muhale, F.A.; Wetmore, B.A.; Thomas, R.S.; McLeod, H.L. Systems pharmacology assessment of the 5-fluorouracil pathway. Pharmacogenomics 2011, 12, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Kidd, E.A.; Yu, J.; Li, X.; Shannon, W.D.; Watson, M.A.; McLeod, H.L. Variance in the expression of 5-fluorouracil pathway genes in colorectal cancer. Clin. Cancer Res. 2005, 11, 2612–2619. [Google Scholar] [CrossRef] [PubMed]
- Weischenfeldt, J.; Simon, R.; Feuerbach, L.; Schlangen, K.; Weichenhan, D.; Minner, S.; Wuttig, D.; Warnatz, H.J.; Stehr, H.; Rausch, T.; et al. Integrative genomic analyses reveal an androgen-driven somatic alteration landscape in early-onset prostate cancer. Cancer Cell 2013, 23, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Minner, S.; Enodien, M.; Sirma, H.; Luebke, A.M.; Krohn, A.; Mayer, P.S.; Simon, R.; Tennstedt, P.; Muller, J.; Scholz, L.; et al. ERG status is unrelated to PSA recurrence in radically operated prostate cancer in the absence of antihormonal therapy. Clin. Cancer Res. 2011, 17, 5878–5888. [Google Scholar] [CrossRef] [PubMed]
- Kluth, M.; Hesse, J.; Heinl, A.; Krohn, A.; Steurer, S.; Sirma, H.; Simon, R.; Mayer, P.S.; Schumacher, U.; Grupp, K.; et al. Genomic deletion of MAP3K7 at 6q12–22 is associated with early PSA recurrence in prostate cancer and absence of TMPRSS2:ERG fusions. Mod. Pathol. 2013, 26, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Burkhardt, L.; Fuchs, S.; Krohn, A.; Masser, S.; Mader, M.; Kluth, M.; Bachmann, F.; Huland, H.; Steuber, T.; Graefen, M.; et al. CHD1 is a 5q21 tumor suppressor required for ERG rearrangement in prostate cancer. Cancer Res. 2013, 73, 2795–2805. [Google Scholar] [CrossRef] [PubMed]
- Krohn, A.; Diedler, T.; Burkhardt, L.; Mayer, P.S.; De Silva, C.; Meyer-Kornblum, M.; Kotschau, D.; Tennstedt, P.; Huang, J.; Gerhauser, C.; et al. Genomic deletion of PTEN is associated with tumor progression and early PSA recurrence in ERG fusion-positive and fusion-negative prostate cancer. Am. J. Pathol. 2012, 181, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Krohn, A.; Seidel, A.; Burkhardt, L.; Bachmann, F.; Mader, M.; Grupp, K.; Eichenauer, T.; Becker, A.; Adam, M.; Graefen, M.; et al. Recurrent deletion of 3p13 targets multiple tumour suppressor genes and defines a distinct subgroup of aggressive ERG fusion-positive prostate cancers. J. Pathol. 2013, 231, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Epstein, J.I.; Feng, Z.; Trock, B.J.; Pierorazio, P.M. Upgrading and downgrading of prostate cancer from biopsy to radical prostatectomy: Incidence and predictive factors using the modified Gleason grading system and factoring in tertiary grades. Eur. Urol. 2012, 61, 1019–1024. [Google Scholar] [CrossRef] [PubMed]
- Yin, D.; Chave, K.J.; Macaluso, C.R.; Galivan, J.; Yao, R. Structural organization of the human γ-glutamyl hydrolase gene. Gene 1999, 238, 463–470. [Google Scholar] [CrossRef]
- Saini, P.K.; Rosenberg, I.H. Isolation of pteroly-γ-oligoglutamyl endopeptidase from chicken intestine with the aid of affinity chromatography. J. Biol. Chem. 1974, 249, 5131–5134. [Google Scholar] [PubMed]
- Yao, R.; Schneider, E.; Ryan, T.J.; Galivan, J. Human γ-glutamyl hydrolase: Cloning and characterization of the enzyme expressed in vitro. Proc. Natl. Acad. Sci. USA 1996, 93, 10134–10138. [Google Scholar] [CrossRef] [PubMed]
- Pollard, C.; Nitz, M.; Baras, A.; Williams, P.; Moskaluk, C.; Theodorescu, D. Genoproteomic mining of urothelial cancer suggests γ-glutamyl hydrolase and diazepam-binding inhibitor as putative urinary markers of outcome after chemotherapy. Am. J. Pathol. 2009, 175, 1824–1830. [Google Scholar] [CrossRef] [PubMed]
- Odin, E.; Wettergren, Y.; Nilsson, S.; Willen, R.; Carlsson, G.; Spears, C.P.; Larsson, L.; Gustavsson, B. Altered gene expression of folate enzymes in adjacent mucosa is associated with outcome of colorectal cancer patients. Clin. Cancer Res. 2003, 9, 6012–6019. [Google Scholar] [PubMed]
- Eto, I.; Krumdieck, C.L. Role of vitamin B12 and folate deficiencies in carcinogenesis. Adv. Exp. Med. Biol. 1986, 206, 313–330. [Google Scholar] [PubMed]
- Blount, B.C.; Ames, B.N. DNA damage in folate deficiency. Baillieres Clin. Haematol. 1995, 8, 461–478. [Google Scholar] [CrossRef]
- Blount, B.C.; Mack, M.M.; Wehr, C.M.; MacGregor, J.T.; Hiatt, R.A.; Wang, G.; Wickramasinghe, S.N.; Everson, R.B.; Ames, B.N. Folate deficiency causes uracil misincorporation into human DNA and chromosome breakage: Implications for cancer and neuronal damage. Proc. Natl. Acad. Sci. USA 1997, 94, 3290–3295. [Google Scholar] [CrossRef] [PubMed]
- Okun, J.G.; Horster, F.; Farkas, L.M.; Feyh, P.; Hinz, A.; Sauer, S.; Hoffmann, G.F.; Unsicker, K.; Mayatepek, E.; Kolker, S. Neurodegeneration in methylmalonic aciduria involves inhibition of complex II and the tricarboxylic acid cycle, and synergistically acting excitotoxicity. J. Biol. Chem. 2002, 277, 14674–14680. [Google Scholar] [PubMed]
- Fowler, B.; Leonard, J.V.; Baumgartner, M.R. Causes of and diagnostic approach to methylmalonic acidurias. J. Inherit. Metab. Dis. 2008, 31, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Ribas, G.S.; Manfredini, V.; de Marco, M.G.; Vieira, R.B.; Wayhs, C.Y.; Vanzin, C.S.; Biancini, G.B.; Wajner, M.; Vargas, C.R. Prevention by l-carnitine of DNA damage induced by propionic and l-methylmalonic acids in human peripheral leukocytes in vitro. Mutat. Res. 2010, 702, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.E.; Hinoue, T.; Kim, M.S.; Sohn, K.J.; Cho, R.C.; Cole, P.D.; Weisenberger, D.J.; Laird, P.W.; Kim, Y.I. γ-Glutamyl hydrolase modulation significantly influences global and gene-specific DNA methylation and gene expression in human colon and breast cancer cells. Genes Nutr. 2015, 10, 444. [Google Scholar] [CrossRef] [PubMed]
- Jeronimo, C.; Bastian, P.J.; Bjartell, A.; Carbone, G.M.; Catto, J.W.; Clark, S.J.; Henrique, R.; Nelson, W.G.; Shariat, S.F. Epigenetics in prostate cancer: Biologic and clinical relevance. Eur. Urol. 2011, 60, 753–766. [Google Scholar] [CrossRef] [PubMed]
- Feinberg, A.P.; Vogelstein, B. Hypomethylation distinguishes genes of some human cancers from their normal counterparts. Nature 1983, 301, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Baylin, S.B.; Ohm, J.E. Epigenetic gene silencing in cancer—A mechanism for early oncogenic pathway addiction? Nat. Rev. Cancer 2006, 6, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Herman, J.G.; Baylin, S.B. Gene silencing in cancer in association with promoter hypermethylation. N. Engl. J. Med. 2003, 349, 2042–2054. [Google Scholar] [CrossRef] [PubMed]
- Stumm, L.; Burkhardt, L.; Steurer, S.; Simon, R.; Adam, M.; Becker, A.; Sauter, G.; Minner, S.; Schlomm, T.; Sirma, H.; et al. Strong expression of the neuronal transcription factor FOXP2 is linked to an increased risk of early PSA recurrence in ERG fusion-negative cancers. J. Clin. Pathol. 2013, 66, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Grupp, K.; Ospina-Klinck, D.; Tsourlakis, M.C.; Koop, C.; Wilczak, W.; Adam, M.; Simon, R.; Sauter, G.; Izbicki, J.R.; Graefen, M.; et al. NY-ESO-1 expression is tightly linked to TMPRSS2-ERG fusion in prostate cancer. Prostate 2014, 74, 1012–1022. [Google Scholar] [CrossRef] [PubMed]
- Grupp, K.; Jedrzejewska, K.; Tsourlakis, M.C.; Koop, C.; Wilczak, W.; Adam, M.; Quaas, A.; Sauter, G.; Simon, R.; Izbicki, J.R.; et al. High mitochondria content is associated with prostate cancer disease progression. Mol. Cancer 2013, 12, 145. [Google Scholar] [CrossRef] [PubMed]
- Burdelski, C.; Bujupi, E.; Tsourlakis, M.C.; Hube-Magg, C.; Kluth, M.; Melling, N.; Lebok, P.; Minner, S.; Koop, C.; Graefen, M.; et al. Loss of SOX9 expression is associated with PSA recurrence in ERG-positive and PTEN deleted prostate cancers. PLoS ONE 2015, 10, e0128525. [Google Scholar] [CrossRef] [PubMed]
- Burdelski, C.; Menan, D.; Tsourlakis, M.C.; Kluth, M.; Hube-Magg, C.; Melling, N.; Minner, S.; Koop, C.; Graefen, M.; Heinzer, H.; et al. The prognostic value of SUMO1/Sentrin specific peptidase 1 (SENP1) in prostate cancer is limited to ERG-fusion positive tumors lacking PTEN deletion. BMC Cancer 2015, 15, 538. [Google Scholar] [CrossRef] [PubMed]
- Grupp, K.; Boumesli, R.; Tsourlakis, M.C.; Koop, C.; Wilczak, W.; Adam, M.; Sauter, G.; Simon, R.; Izbicki, J.R.; Graefen, M.; et al. The prognostic impact of high Nijmegen breakage syndrome (NBS1) gene expression in ERG-negative prostate cancers lacking PTEN deletion is driven by KPNA2 expression. Int. J. Cancer 2014, 135, 1399–1407. [Google Scholar] [CrossRef] [PubMed]
- Brase, J.C.; Johannes, M.; Mannsperger, H.; Falth, M.; Metzger, J.; Kacprzyk, L.A.; Andrasiuk, T.; Gade, S.; Meister, M.; Sirma, H.; et al. TMPRSS2-ERG-specific transcriptional modulation is associated with prostate cancer biomarkers and TGF-β signaling. BMC Cancer 2011, 11, 507. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Iljin, K.; Sara, H.; Mpindi, J.P.; Mirtti, T.; Vainio, P.; Rantala, J.; Alanen, K.; Nees, M.; Kallioniemi, O. FZD4 as a mediator of ERG oncogene-induced WNT signaling and epithelial-to-mesenchymal transition in human prostate cancer cells. Cancer Res. 2010, 70, 6735–6745. [Google Scholar] [CrossRef] [PubMed]
- Jhavar, S.; Brewer, D.; Edwards, S.; Kote-Jarai, Z.; Attard, G.; Clark, J.; Flohr, P.; Christmas, T.; Thompson, A.; Parker, M.; et al. Integration of ERG gene mapping and gene-expression profiling identifies distinct categories of human prostate cancer. BJU Int. 2009, 103, 1256–1269. [Google Scholar] [CrossRef] [PubMed]
- Cuzick, J.; Swanson, G.P.; Fisher, G.; Brothman, A.R.; Berney, D.M.; Reid, J.E.; Mesher, D.; Speights, V.O.; Stankiewicz, E.; Foster, C.S.; et al. Prognostic value of an RNA expression signature derived from cell cycle proliferation genes in patients with prostate cancer: A retrospective study. Lancet Oncol. 2011, 12, 245–255. [Google Scholar] [CrossRef]
- Knezevic, D.; Goddard, A.D.; Natraj, N.; Cherbavaz, D.B.; Clark-Langone, K.M.; Snable, J.; Watson, D.; Falzarano, S.M.; Magi-Galluzzi, C.; Klein, E.A.; et al. Analytical validation of the oncotype DX prostate cancer assay—A clinical RT-PCR assay optimized for prostate needle biopsies. BMC Genom. 2013, 14, 690. [Google Scholar] [CrossRef] [PubMed]
- Sauter, G.; Steurer, S.; Clauditz, T.S.; Krech, T.; Wittmer, C.; Lutz, F.; Lennartz, M.; Janssen, T.; Hakimi, N.; Simon, R.; et al. Clinical Utility of quantitative gleason grading in prostate biopsies and prostatectomy specimens. Eur. Urol. 2016, 69, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Rhee, M.S.; Wang, Y.; Nair, M.G.; Galivan, J. Acquisition of resistance to antifolates caused by enhanced γ-glutamyl hydrolase activity. Cancer Res. 1993, 53 (Suppl. 10), 2227–2230. [Google Scholar] [PubMed]
- McCloskey, D.E.; McGuire, J.J.; Russell, C.A.; Rowan, B.G.; Bertino, J.R.; Pizzorno, G.; Mini, E. Decreased folylpolyglutamate synthetase activity as a mechanism of methotrexate resistance in CCRF-CEM human leukemia sublines. J. Biol. Chem. 1991, 266, 6181–6187. [Google Scholar] [PubMed]
- Barakat, R.R.; Li, W.W.; Lovelace, C.; Bertino, J.R. Intrinsic resistance of cervical squamous cell carcinoma cell lines to methotrexate (MTX) as a result of decreased accumulation of intracellular MTX polyglutamates. Gynecol. Oncol. 1993, 51, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Cole, P.D.; Kamen, B.A.; Gorlick, R.; Banerjee, D.; Smith, A.K.; Magill, E.; Bertino, J.R. Effects of overexpression of γ-glutamyl hydrolase on methotrexate metabolism and resistance. Cancer Res. 2001, 61, 4599–4604. [Google Scholar] [PubMed]
- Shelley, M.; Harrison, C.; Coles, B.; Staffurth, J.; Wilt, T.J.; Mason, M.D. Chemotherapy for hormone-refractory prostate cancer. Cochrane Database Syst. Rev. 2006, 4, CD005247. [Google Scholar]
- Erbersdobler, A.; Hammerer, P.; Huland, H.; Henke, R.P. Numerical chromosomal aberrations in transition-zone carcinomas of the prostate. J. Urol. 1997, 158, 1594–1598. [Google Scholar] [CrossRef]
- Mirlacher, M.; Simon, R. Recipient block TMA technique. Methods Mol. Biol. 2010, 664, 37–44. [Google Scholar] [PubMed]
- Kononen, J.; Bubendorf, L.; Kallioniemi, A.; Barlund, M.; Schraml, P.; Leighton, S.; Torhorst, J.; Mihatsch, M.J.; Sauter, G.; Kallioniemi, O.P. Tissue microarrays for high-throughput molecular profiling of tumor specimens. Nat. Med. 1998, 4, 844–847. [Google Scholar] [CrossRef] [PubMed]
- Lennartz, M.; Minner, S.; Brasch, S.; Wittmann, H.; Paterna, L.; Angermeier, K.; Ozturk, E.; Shihada, R.; Ruge, M.; Kluth, M.; et al. The Combination of DNA ploidy status and PTEN/6q15 deletions provides strong and independent prognostic information in prostate cancer. Clin. Cancer Res. 2016, 22, 2802–2811. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | GGH IHC Result (%) | p Value | |||
|---|---|---|---|---|---|
| N Evaluable | Negative | Low | High | ||
| All cancers | 10,562 | 11.7 | 49.6 | 38.6 | |
| Tumor stage | |||||
| pT2 | 6851 | 12.3 | 50.8 | 36.9 | <0.0001 |
| pT3a | 2333 | 11.0 | 46.5 | 42.5 | |
| pT3b-pT4 | 1338 | 9.9 | 49.1 | 41.0 | |
| Gleason grade | |||||
| ≤3 + 3 | 2329 | 16.7 | 52.3 | 31.0 | <0.0001 |
| 3 + 4 | 5430 | 10.8 | 49.1 | 40.1 | |
| 3 + 4 Tertiary 5 | 397 | 10.1 | 51.9 | 38.0 | |
| 4 + 3 | 914 | 10.2 | 48.1 | 41.7 | |
| 4 + 3 Tertiary 5 | 568 | 8.1 | 48.2 | 43.7 | |
| ≥4 + 4 | 450 | 12.0 | 48.9 | 39.1 | |
| Lymph node metastasis | |||||
| N0 | 5996 | 10.3 | 48.0 | 41.7 | 0.0928 |
| N+ | 608 | 12.2 | 50.3 | 37.5 | |
| Preoperative PSA level (ng/mL) | |||||
| <4 | 1290 | 9.5 | 50.5 | 39.9 | <0.0001 |
| 4–10 | 6362 | 11.3 | 48.9 | 39.9 | |
| 10–20 | 2076 | 12.6 | 51.4 | 36.0 | |
| >20 | 726 | 17.1 | 49.9 | 33.1 | |
| Surgical margin | |||||
| Negative | 8413 | 11.7 | 49.9 | 38.4 | 0.9259 |
| Positive | 1949 | 11.9 | 49.4 | 38.7 | |
| Subset | Scenario | N Analyzable | p Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Preoperative PSA-Level | pT Stage | cT Stage | Gleason Grade RPE 1 | Gleason Grade Biopsy | N-Stage | R-Status | GGH-Expression | |||
| Total | 1 | 5773 | <0.0001 | <0.0001 | - | <0.0001 | - | <0.0001 | 0.0010 | 0.0290 |
| 2 | 9281 | <0.0001 | <0.0001 | - | <0.0001 | - | - | <0.0001 | 0.0047 | |
| 3 | 9163 | <0.0001 | - | <0.0001 | <0.0001 | - | - | - | 0.0246 | |
| 4 | 9038 | <0.0001 | - | <0.0001 | - | <0.0001 | - | - | 0.0032 | |
| ERG neg. subset | 1 | 2928 | 0.0002 | <0.0001 | - | <0.0001 | - | 0.0006 | 0.0894 | 0.0094 |
| 2 | 4583 | <0.0001 | <0.0001 | - | <0.0001 | - | - | 0.0006 | 0.0111 | |
| 3 | 4549 | <0.0001 | - | <0.0001 | <0.0001 | - | - | - | 0.1042 | |
| 4 | 4492 | <0.0001 | - | <0.0001 | - | <0.0001 | - | - | 0.0197 | |
| ERG pos. subset | 1 | 2255 | 0.0060 | <0.0001 | - | <0.0001 | - | 0.0038 | 0.0143 | 0.4035 |
| 2 | 3573 | <0.0001 | <0.0001 | - | <0.0001 | - | - | <0.0001 | 0.2573 | |
| 3 | 3500 | <0.0001 | - | <0.0001 | <0.0001 | - | - | - | 0.1868 | |
| 4 | 3447 | <0.0001 | - | <0.0001 | - | <0.0001 | - | - | 0.2427 | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melling, N.; Rashed, M.; Schroeder, C.; Hube-Magg, C.; Kluth, M.; Lang, D.; Simon, R.; Möller-Koop, C.; Steurer, S.; Sauter, G.; et al. High-Level γ-Glutamyl-Hydrolase (GGH) Expression is Linked to Poor Prognosis in ERG Negative Prostate Cancer. Int. J. Mol. Sci. 2017, 18, 286. https://doi.org/10.3390/ijms18020286
Melling N, Rashed M, Schroeder C, Hube-Magg C, Kluth M, Lang D, Simon R, Möller-Koop C, Steurer S, Sauter G, et al. High-Level γ-Glutamyl-Hydrolase (GGH) Expression is Linked to Poor Prognosis in ERG Negative Prostate Cancer. International Journal of Molecular Sciences. 2017; 18(2):286. https://doi.org/10.3390/ijms18020286
Chicago/Turabian StyleMelling, Nathaniel, Masoud Rashed, Cornelia Schroeder, Claudia Hube-Magg, Martina Kluth, Dagmar Lang, Ronald Simon, Christina Möller-Koop, Stefan Steurer, Guido Sauter, and et al. 2017. "High-Level γ-Glutamyl-Hydrolase (GGH) Expression is Linked to Poor Prognosis in ERG Negative Prostate Cancer" International Journal of Molecular Sciences 18, no. 2: 286. https://doi.org/10.3390/ijms18020286