Population-Specific Associations of Deleterious Rare Variants in Coding Region of P2RY1–P2RY12 Purinergic Receptor Genes in Large-Vessel Ischemic Stroke Patients

 , ,
, ,
Abstract
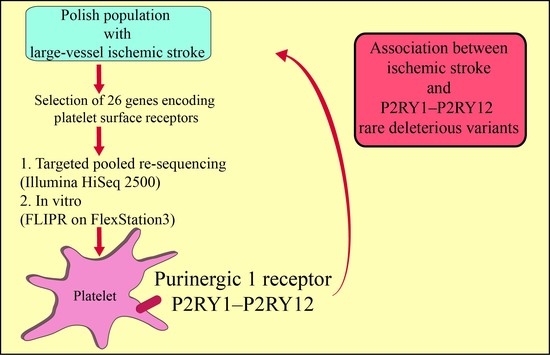
:
1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Genotyping
4.2.1. Statistical Tests and Calculations
4.2.2. Sample Size and Power Considerations
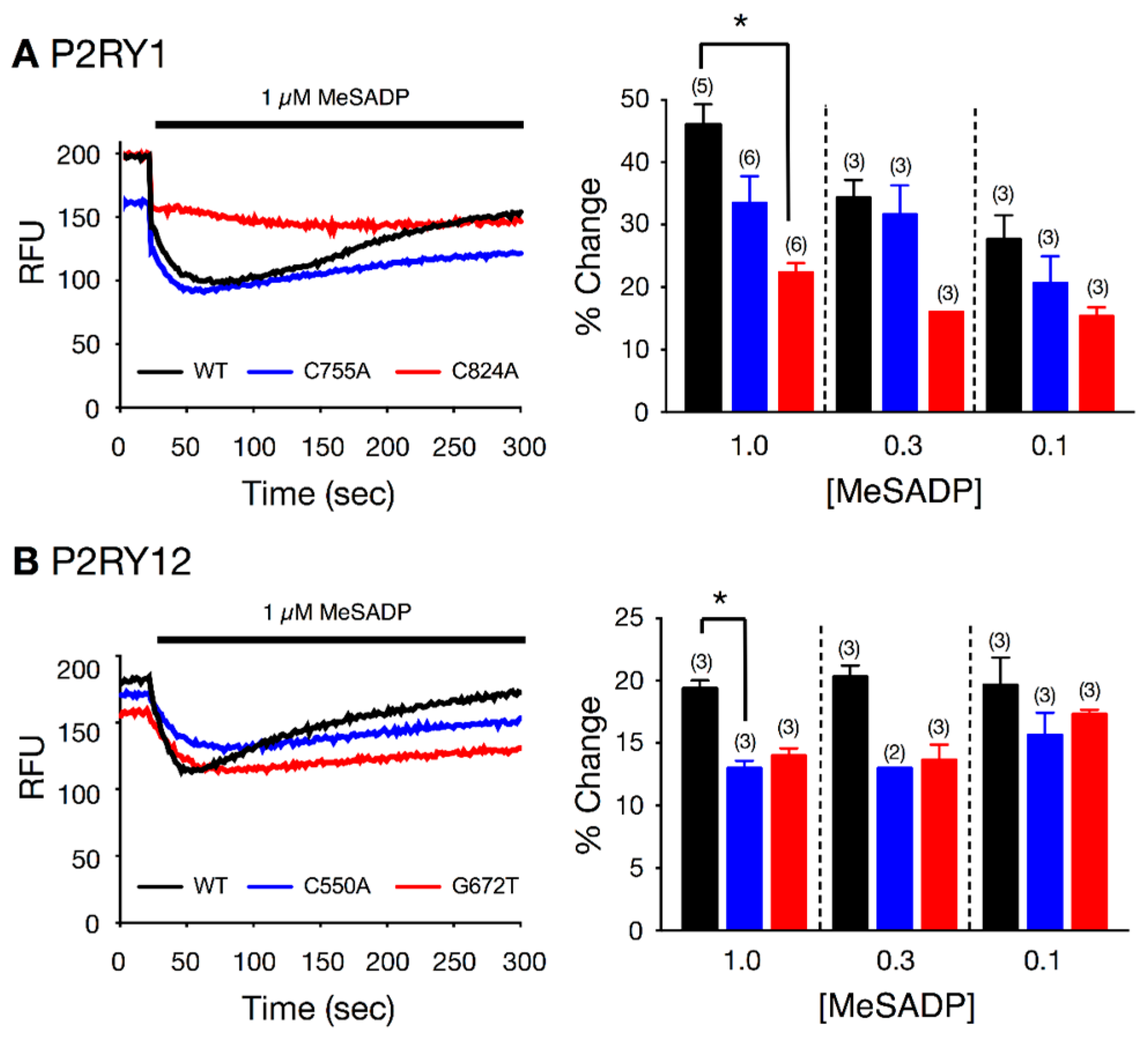
4.3. Fluorescence-Based Functional Assay for P2RY1 and P2RY12 Receptor Activation in L Cells
5. Limitations
6. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| IS | Ischemic stroke |
| GWAS | Genome wide association studies |
| MAF | Minor allele frequency |
| SNVs | Single nucleotide variants |
| SNPs | Single nucleotide polymorphism |
| CADD | Combined Annotation Dependent Depletion |
| TOAST | Trial of Org 10172 in Acute Stroke Treatment |
| CAD | Coronary artery disease |
| CHF | Congestive heart failure |
| DM | Diabetes mellitus |
| cMAF | Combined MAF |
| CMAT | Cumulative minor-allele test |
| SNP | Single nucleotide polymorphism |
| GIRK | G protein inwardly-rectifying K+ |
| MeSAD | 2-Methylthioadenosine diphosphate |
| TIA | Transient ischemic attack |
References
- Welter, D.; MacArthur, J.; Morales, J.; Burdett, T.; Hall, P.; Junkins, H.; Klemm, A.; Flicek, P.; Manolio, T.; Hindorff, L.; et al. The NHGRI GWAS Catalog, a curated resource of SNP-trait associations. Nucleic Acids Res. 2014, 42, D1001–D1006. [Google Scholar] [CrossRef] [PubMed]
- Holliday, E.G.; Maguire, J.M.; Evans, T.J.; Koblar, S.A.; Jannes, J.; Sturm, J.W.; Hankey, G.J.; Baker, R.; Golledge, J.; Parsons, M.W.; et al. Common variants at 6p21.1 are associated with large artery atherosclerotic stroke. Nat. Genet. 2012, 44, 1147–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bevan, S.; Traylor, M.; Adib-Samii, P.; Malik, R.; Paul, N.L.; Jackson, C.; Farrall, M.; Rothwell, P.M.; Sudlow, C.; Dichgans, M.; et al. Genetic heritability of ischemic stroke and the contribution of previously reported candidate gene and genomewide associations. Stroke 2012, 43, 3161–3167. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G. Rare and common variants: Twenty arguments. Nat. Rev. Genet. 2011, 13, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Francioli, L.C.; Menelaou, A.; Pulit, S.L.; van Dijk, F.; Palamara, P.F.; Elbers, C.C.; Neerincx, P.B.; Ye, K.; Guryev, V.; Kloosterman, W.P.; et al. Whole-genome sequence variation, population structure and demographic history of the Dutch population. Nat. Genet. 2014, 46, 818–825. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.C.; Cole, J.W.; Kittner, S.J.; Mitchell, B.D. Genetics of ischemic stroke in young adults. Circ. Cardiovasc. Genet. 2014, 7, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Auer, P.L.; Nalls, M.; Meschia, J.F.; Worrall, B.B.; Longstreth, W.T., Jr.; Seshadri, S.; Kooperberg, C.; Burger, K.M.; Carlson, C.S.; Carty, C.L.; et al. Rare and Coding Region Genetic Variants Associated With Risk of Ischemic Stroke: The NHLBI Exome Sequence Project. JAMA Neurol. 2015, 72, 781–788. [Google Scholar] [CrossRef] [PubMed]
- Anyanwu, C.; Hahn, M.; Nath, M.; Li, J.; Barone, F.C.; Rosenbaum, D.M.; Zhou, J. Platelets Pleiotropic Roles in Ischemic Stroke. Austin J. Cerebrovasc. Dis. Stroke 2016, 3, 1048. [Google Scholar]
- Postula, M.; Janicki, P.K.; Milanowski, L.; Pordzik, J.; Eyileten, C.; Karlinski, M.; Wylezol, P.; Solarska, M.; Czlonkowka, A.; Kurkowska-Jastrzebka, I.; et al. Association of frequent genetic variants in platelet activation pathway genes with large-vessel ischemic stroke in Polish population. Platelets 2017, 28, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Milanowski, L.; Pordzik, J.; Janicki, P.K.; Postula, M. Common genetic variants in platelet surface receptors and its association with ischemic stroke. Pharmacogenomics 2016, 17, 953–971. [Google Scholar] [CrossRef] [PubMed]
- Kircher, M.; Witten, D.M.; Jain, P.; O’Roak, B.J.; Cooper, G.M.; Shendure, J. A general framework for estimating the relative pathogenicity of human genetic variants. Nat. Genet. 2014, 46, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Visschedijk, M.C.; Alberts, R.; Mucha, S.; Deelen, P.; de Jong, D.J.; Pierik, M.; Spekhorst, L.M.; Imhann, F.; van der Meulen-de, A.E.; van der Woude, C.J.; et al. Pooled Resequencing of 122 Ulcerative Colitis Genes in a Large Dutch Cohort Suggests Population-Specific Associations of Rare Variants in MUC2. PLoS ONE 2016, 11, e0159609. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Z.; van Sommeren, S.; Huang, H.; Ng, S.C.; Alberts, R.; Takahashi, A.; Ripke, S.; Lee, J.C.; Jostins, L.; Shah, T.; et al. Association analyses identify 38 susceptibility loci for inflammatory bowel disease and highlight shared genetic risk across populations. Nat. Genet. 2015, 47, 979–986. [Google Scholar] [CrossRef] [PubMed]
- Hong, K.W.; Shin, M.S.; Ahn, Y.B.; Lee, H.J.; Kim, H.D. Genomewide association study on chronic periodontitis in Korean population: Results from the Yangpyeong health cohort. J. Clin. Periodontol. 2015, 42, 703–710. [Google Scholar] [CrossRef] [PubMed]
- Prescott, N.J.; Lehne, B.; Stone, K.; Lee, J.C.; Taylor, K.; Knight, J.; Papouli, E.; Mirza, M.M.; Simpson, M.A.; Spain, S.L.; et al. Pooled Sequencing of 531 Genes in Inflammatory Bowel Disease Identifies an Associated Rare Variant in BTNL2 and Implicates Other Immune Related Genes. PLoS Genet. 2015, 11, e1004955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziegler, S.; Schillinger, M.; Funk, M.; Felber, K.; Exner, M.; Mlekusch, W.; Sabeti, S.; Amighi, J.; Minar, E.; Brunner, M.; et al. Association of a functional polymorphism in the clopidogrel target receptor gene, P2Y12, and the risk for ischemic cerebrovascular events in patients with peripheral artery disease. Stroke 2005, 36, 1394–1399. [Google Scholar] [CrossRef] [PubMed]
- Zee, R.Y.; Michaud, S.E.; Diehl, K.A.; Chasman, D.I.; Emmerich, J.; Gaussem, P.; Aiach, M.; Ridker, P.M. Purinergic receptor P2Y, G-protein coupled, 12 gene variants and risk of incident ischemic stroke, myocardial infarction, and venous thromboembolism. Atherosclerosis 2008, 197, 694–699. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Nakayama, T.; Sato, N.; Yamaguchi, M.; Izumi, Y.; Kasamaki, Y.; Ohta, M.; Soma, M.; Aoi, N.; Ozawa, Y.; et al. Purinergic receptor P2Y, G-protein coupled, 2 (P2RY2) gene is associated with cerebral infarction in Japanese subjects. Hypertens. Res. 2009, 32, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Zhou, Q.; Wang, C.; Lin, J.; Liu, P.; Fu, C. Platelet receptor Gene (P2Y12, P2Y1) and platelet glycoprotein Gene (GPIIIa) polymorphisms are associated with antiplatelet drug responsiveness and clinical outcomes after acute minor ischemic stroke. Eur. J. Clin. Pharmacol. 2017, 73, 437–443. [Google Scholar] [CrossRef] [PubMed]
- You, J.; Johnson, T.D.; Marrelli, S.P.; Mombouli, J.V.; Bryan, R.M., Jr. P2u receptor-mediated release of endothelium-derived relaxing factor/nitric oxide and endothelium-derived hyperpolarizing factor from cerebrovascular endothelium in rats. Stroke 1999, 30, 1125–1133. [Google Scholar] [CrossRef] [PubMed]
- Marrelli, S.P. Altered endothelial Ca2+ regulation after ischemia/reperfusion produces potentiated endothelium-derived hyperpolarizing factor-mediated dilations. Stroke 2002, 33, 2285–2291. [Google Scholar] [CrossRef] [PubMed]
- Ignatovica, V.; Latkovskis, G.; Peculis, R.; Megnis, K.; Schioth, H.B.; Vaivade, I.; Fridmanis, D.; Pirags, V.; Erglis, A.; Klovins, J. Single nucleotide polymorphisms of the purinergic 1 receptor are not associated with myocardial infarction in a Latvian population. Mol. Biol. Rep. 2012, 39, 1917–1925. [Google Scholar] [CrossRef] [PubMed]
- Abbracchio, M.P.; Burnstock, G.; Boeynaems, J.-M.; Barnard, E.A.; Boyer, J.L.; Kennedy, C.; Knight, G.E.; Fumagalli, M.; Gachet, C.; Jacobson, K.A.; et al. International Union of Pharmacology LVIII: Update on the P2Y G Protein-Coupled Nucleotide Receptors: From Molecular Mechanisms and Pathophysiology to Therapy. Pharmacol. Rev. 2006, 58, 281–341. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.; Zhang, S.; Zou, J.; Chen, Y.; Chen, H. P2Y12 receptor gene polymorphism and the risk of resistance to clopidogrel: A meta-analysis and review of the literature. Adv. Clin. Exp. Med. 2017, 26, 343–349. [Google Scholar] [PubMed]
- Liu, R.; Zhou, Z.; Chen, Y.; Li, J.L.; Jin, J.; Huang, M.; Zhao, M.; Yu, W.B.; Chen, X.M.; Cai, Y.F.; et al. Associations of CYP3A4, NR1I2, CYP2C19 and P2RY12 polymorphisms with clopidogrel resistance in Chinese patients with ischemic stroke. Acta Pharmacol. Sin. 2016, 37, 882–888. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Lin, J.; Wang, Y.; Zhou, J.; Zhou, Q. Interaction among CYP2C8, GPIIIa and P2Y12 variants increase susceptibility to ischemic stroke in Chinese population. Oncotarget 2017, 8, 70811–70820. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Nakayama, T.; Sato, N.; Izumi, Y.; Kasamaki, Y.; Ohta, M.; Soma, M.; Aoi, N.; Ozawa, Y.; Ma, Y. The purinergic receptor P2Y, G-protein coupled, 2 (P2RY2) gene associated with essential hypertension in Japanese men. J. Hum. Hypertens. 2010, 24, 327–335. [Google Scholar] [CrossRef] [PubMed]
- WHO Task Force on Stroke and Other Cerebrovascular Diseases. Recommendations on stroke prevention, diagnosis, and therapy. Report of the WHO task force on stroke and other cerebrovascular disorders. Stroke 1989, 20, 1407–1431. [Google Scholar]
- Foulkes, M.; Wolf, P.; Price, T.; Mohr, J.; Hier, D. The Stroke Data Bank: Design, methods, and baseline characteristics. Stroke 1988, 19, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Grabska, K.; Gromadzka, G.; Członkowska, A. Infections and ischemic stroke outcome. Neurol. Res. Int. 2011, 2011, 691348. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Abecasis, G.R.; Boehnke, M.; Lin, X. Rare-variant association analysis: Study designs and statistical tests. Am. J. Hum. Genet. 2014, 95, 5–23. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Bost, D.; Wilson, S.; Maruki, T.; Harrison, S. Population-genetic inference from pooled-sequencing data. Genome Biol. Evol. 2014, 6, 1210–1218. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Chr | Gene | Position | Ref | Alt | dbSNP149 | cDNA | Protein AA | CADD | MAF Ctrl | MAF Stroke |
|---|---|---|---|---|---|---|---|---|---|---|
| chr3 | ITPR1 | 4714920 | A | G | rs35789999 | c.A2260G | p.M754V | 15.92 | 0.0014 | |
| chr3 | ITPR1 | 4716885 | C | T | rs201519806 | c.C2687T | p.A896V | 12.80 | 0.0011 | |
| chr3 | ITPR1 | 4774887 | G | T | c.G5147T | p.G1716V | 15.77 | 0.0014 | ||
| chr3 | ITPR1 | 4821291 | G | T | rs373973399 | c.G6160T | p.A2054S | 20.2 | 0.0013 | |
| chr3 | ITPR1 | 4842276 | G | A | rs201144431 | c.G6910A | p.A2304T | 16.91 | 0.0015 | 0.001 |
| chr17 | GP1BA | 4837662 | T | C | rs201408072 | c.T1763C | p.V588A | 15.12 | 0.0013 | |
| chr3 | RAF1 | 12641707 | C | T | rs555034652 | c.G934A | p.V312M | 12.39 | 0.0017 | |
| chr1 | PTAFR | 28477192 | T | C | rs138629813 | c.A341G | p.N114S | 20.8 | 0.0018 | |
| chr1 | PTAFR | 28477408 | C | A | c.G125T | p.R42L | 14.45 | 0.0015 | ||
| chr17 | ITGA2B | 42453084 | C | T | rs74988902 | c.G2602A | p.V868M | 13.50 | 0.0014 | |
| chr17 | ITGA2B | 42455791 | G | A | rs200481952 | c.C2033T | p.A678V | 20.4 | 0.0015 | |
| chr17 | ITGA2B | 42457474 | G | A | rs548977341 | c.C1648T | p.R550W | 22.3 | 0.0018 | |
| chr17 | ITGA2B | 42463054 | G | C | rs76066357 | c.C439G | p.L147V | 11.09 | 0.0025 | |
| chr17 | ITGB3 | 45363765 | A | G | rs56173532 | c.A754G | p.I252V | 13.67 | 0.0012 | |
| chr17 | ITGB3 | 45376796 | G | A | rs144884023 | c.G1813A | p.G605S | 35 | 0.0013 | |
| chr19 | PTGIR | 47126849 | G | A | rs4987262 | c.C634T | p.R212C | 22.3 | 0.0034 | |
| chr5 | ITGA2 | 52344487 | A | G | rs55973669 | c.A517G | p.I173V | 12.10 | 0.0022 | |
| chr19 | GP6 | 55543660 | G | A | rs199588110 | c.C172T | p.R58C | 18.28 | 0.0025 | 0.0016 |
| chr19 | GP6 | 55543660 | G | A | rs199588110 | c.C172T | p.R58C | 18.28 | 0.0023 | |
| chr19 | GP6 | 55543692 | C | T | rs750889036 | c.G140A | p.R47Q | 10.75 | 0.0019 | |
| chr11 | FERMT3 | 63974970 | T | G | rs759179590 | c.T134G | p.V45G | 33 | 0.0035 | 0.0042 |
| chr11 | FERMT3 | 63974995 | C | G | rs142815441 | c.C159G | p.I53M | 12.17 | 0.0013 | |
| chr11 | FERMT3 | 63978538 | G | A | rs762181713 | c.G409A | p.E137K | 29.0 | 0.0014 | |
| chr11 | P2RY2 | 72945434 | T | C | rs148391446 | c.T230C | p.V77A | 14.97 | 0.0017 | |
| chr11 | P2RY2 | 72945799 | A | G | rs141776297 | c.A595G | p.S199G | 15.02 | 0.0021 | |
| chr11 | P2RY2 | 72945799 | A | G | rs141776297 | c.A595G | p.S199G | 15.02 | 0.0019 | 0.0026 |
| chr11 | P2RY2 | 72946279 | T | C | rs74472890 | c.T1075C | p.S359P | 12.54 | 0.0024 | |
| chr3 | GP9 | 128781048 | G | A | rs3796130 | c.G466A | p.A156T | 12.21 | 0.0015 | |
| chr3 | P2RY12 * | 151055962 | C | A | c.G672T | p.R224S | 15 | 0.0021 | ||
| chr3 | P2RY12 * | 151056084 | G | T | c.C550A | p.L184I | 15.13 | 0.0013 | ||
| chr3 | P2RY1 * | 152554155 | G | A | c.G584A | p.R195H | 18.35 | 0.0012 | 0.0034 | |
| chr3 | P2RY1 * | 152554326 | C | A | c.C755A | p.S252Y | 22.4 | 0.0015 | ||
| chr3 | P2RY1 * | 152554395 | C | A | c.C824A | p.P275H | 22.8 | 0.0013 | ||
| chr3 | P2RY1 * | 152554482 | C | T | rs868057570 | c.C911T | p.A304V | 12.84 | 0.0026 | |
| chr1 | PEAR1 | 156878116 | C | T | c.C1099T | p.R367W | 18.01 | 0.0012 | ||
| chr1 | SELP | 169576246 | G | A | c.C1460T | p.A487V | 12.76 | 0.0017 | ||
| chr1 | SELP | 169581608 | G | A | rs139249907 | c.C808T | p.R270X | 13.86 | 0.0016 | |
| chr3 | GP5 | 194117640 | C | A | c.G1372T | p.A458S | 10.81 | 0.0012 |
| Gene | All Individuals (n = 1000). Number of Variant Carriers for Each Locus and Cohort in Brackets | Subjects without Cardiac Disease (n = 605). Number of Variants Carriers for Each Locus and Cohort in Brackets | ||||
|---|---|---|---|---|---|---|
| Region | cMAF ctrl | cMAF stroke | CMAT P/Fisher | cMAF ctrl | cMAF stroke | CMAT P/Fisher |
| GP6 | 0.006 (3) | 0.002 (1) | 0.3700 | 0.000 (0) | 0.000 (0) | NA |
| ITGA2 | 0.000 (0) | 0.004 (2) | 0.4900 | 0.0000 (0) | 0.002 (2) | 0.51 |
| ITGA2B/ITGB3 | 0.004 (2) | 0.014 (7) | 0.1200 | 0.005 (2) | 0.006 (5) | 0.62 |
| ITPR1 | 0.004 (2) | 0.006 (3) | 0.4900 | 0.0025 (1) | 0.002 (2) | 0.90 |
| P2RY1/P2RY12 | 0.002 (1) | 0.02 (10) | 0.0007 ** | 0.0025 (1) | 0.123 (10) | 0.002 ** |
| P2RY2 | 0.004 (2) | 0.006 (3) | 0.2100 | 0.0006 (1) | 0.004 (3) | 0.62 |
| PEAR1 | 0.0000 (0) | 0.002 (1) | 1.0000 | 0.000 (0) | 0.000 (0) | NA |
| PTAFR | 0.0000 (0) | 0.002 (1) | 1.0000 | 0.0000 (0) | 0.001 (1) | 0.99 |
| SELP | 0.0000 (0) | 0.004 (2) | 0.1200 | 0.0000 (0) | 0.002 (2) | 0.14 |
| PTGIR | 0.0000 (0) | 0.004 (2) | 0.2500 | 0.0000 (0) | 0.002 (2) | 0.26 |
| Total # | 0.02 (10) | 0.064 (32) | 0.0005 * | 0.0125 (5) | 0.033 (27) | 0.03 * |
| OR | 3.4 (7.6–6.9) | 2.8 (1.1–7.3) | ||||
| Gene | Protein Product | Chr Location |
|---|---|---|
| P2RY2 | purinergic receptor P2Y, G-protein coupled, 2 | 11q13.5-q14.1 |
| P2RY12 | purinergic receptor P2Y, G-protein coupled, 12 | 3q25.1 |
| P2RY1 | Purinergic receptor P2Y, G-protein coupled, 1 | 3q25.2 |
| ITGB3 | integrin, beta 3 (platelet glycoprotein IIIa, antigen CD61) | 17q21.32 |
| ITGA2B | integrin, alpha 2b (platelet glycoprotein IIb of IIb/IIIa complex, antigen CD41) | 17q21.32 |
| GP5 | glycoprotein V | 3q29 |
| GP9 | glycoprotein IX | 3q21.3 |
| GP6 | glycoprotein VI | 19q13.42 |
| GP1Bα | glycoprotein 1bα | 17p13.2 |
| GP1BB | glycoprotein Ib (platelet), beta polypeptide, antigen CD42c | 22q11.21 |
| ITGA2 (GPIa) | integrin, alpha 2 (alpha 2 subunit of VLA-2 receptor, antigen CD49B) | 5q11.2 |
| ADRA2A | adrenoceptor alpha 2A | 10q25.2 |
| TBXA2R | thromboxane A2 receptor | 19p13.3 |
| HTR2A | 5-hydroxytryptamine (serotonin) receptor 2A, G protein-coupled | 13q14-q21 |
| F2R (PAR-1) | proteinase-activated receptor 1 (PAR1), coagulation factor II (thrombin) receptor | 5q13 |
| F2RL3 (PAR-4) | protease activated receptor 4 (PAR-4), coagulation factor II (thrombin) receptor-like 3 | 19p12 |
| PEAR1 | platelet endothelial aggregation receptor-1 | 1q23.1 |
| GNB3 | guanine nucleotide binding protein (G protein), beta polypeptide 3 | 12p13 |
| CD148 | receptor-type protein tyrosine phosphatase | 11p11.1 |
| ITPR1 | inositol 1,4,5-trisphosphate receptor, type 1 | 3p26.1 |
| CD36 | thrombospondin receptor, antigen CD36 | 7q11.2 |
| CD40 | TNF receptor superfamily member 5, antigen CD40 | 20q12-q13.2 |
| EPR1 | effector cell peptidase receptor 1 | 17q25 |
| PECAM-1 | platelet/endothelial cell adhesion molecule 1 | 17q23.3 |
| FERMT3 | ferritin family member 3 | 11q13.1 |
| PTAFR | platelet-activating factor receptor | 1p35-p34.3 |
| PTGIR | prostaglandin I2 (prostacyclin) receptor (IP) | 19q13. |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janicki, P.K.; Eyileten, C.; Ruiz-Velasco, V.; Sedeek, K.A.; Pordzik, J.; Czlonkowska, A.; Kurkowska-Jastrzebska, I.; Sugino, S.; Imamura-Kawasawa, Y.; Mirowska-Guzel, D.; et al. Population-Specific Associations of Deleterious Rare Variants in Coding Region of P2RY1–P2RY12 Purinergic Receptor Genes in Large-Vessel Ischemic Stroke Patients. Int. J. Mol. Sci. 2017, 18, 2678. https://doi.org/10.3390/ijms18122678
Janicki PK, Eyileten C, Ruiz-Velasco V, Sedeek KA, Pordzik J, Czlonkowska A, Kurkowska-Jastrzebska I, Sugino S, Imamura-Kawasawa Y, Mirowska-Guzel D, et al. Population-Specific Associations of Deleterious Rare Variants in Coding Region of P2RY1–P2RY12 Purinergic Receptor Genes in Large-Vessel Ischemic Stroke Patients. International Journal of Molecular Sciences. 2017; 18(12):2678. https://doi.org/10.3390/ijms18122678
Chicago/Turabian StyleJanicki, Piotr K., Ceren Eyileten, Victor Ruiz-Velasco, Khaled Anwar Sedeek, Justyna Pordzik, Anna Czlonkowska, Iwona Kurkowska-Jastrzebska, Shigekazu Sugino, Yuka Imamura-Kawasawa, Dagmara Mirowska-Guzel, and et al. 2017. "Population-Specific Associations of Deleterious Rare Variants in Coding Region of P2RY1–P2RY12 Purinergic Receptor Genes in Large-Vessel Ischemic Stroke Patients" International Journal of Molecular Sciences 18, no. 12: 2678. https://doi.org/10.3390/ijms18122678