Recent Advances in Allergy Research Using Humanized Mice

 , , , and
, , , and
Abstract
:1. Introduction
2. Development of First-Generation Humanized Mice
3. Development of Second-Generation Humanized Mice
4. Humanized Mouse Model for Allergy Research
4.1. PBMC-Transferred Allergy Models
4.2. Limitations of PBMC-Transferred Allergy Models
4.3. HSC-Transferred Allergy Model
4.4. Bone Marrow, Liver, Thymus (BLT) Allergy Model
4.5. Humanized Mouse Models of Eosinophilic Airway Inflammation
5. Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AD | Atopic dermatitis |
| AHR | Airway hyperresponsiveness |
| BLT | Bone marrow, liver, and thymus |
| GM-CSF | Granulocyte-macrophage colony stimulating factor |
| HSC | Hematopoietic stem cell |
| NOG or NSG | NOD/Shi-scid IL2rγnull |
| PBMC | Peripheral blood mononuclear cell |
| PCA | Passive cutaneous anaphylaxis |
| SCF | Stem cell factor |
References
- Gordon, J.W.; Scangos, G.A.; Plotkin, D.J.; Barbosa, J.A.; Ruddle, F.H. Genetic transformation of mouse embryos by microinjection of purified DNA. Proc. Natl. Acad. Sci. USA 1980, 77, 7380–7384. [Google Scholar] [CrossRef] [PubMed]
- Jaenisch, R. Germ line integration and Mendelian transmission of the exogenous Moloney leukemia virus. Proc. Natl. Acad. Sci. USA 1976, 73, 1260–1264. [Google Scholar] [CrossRef] [PubMed]
- Jaenisch, R.; Mintz, B. Simian virus 40 DNA sequences in DNA of healthy adult mice derived from preimplantation blastocysts injected with viral DNA. Proc. Natl. Acad. Sci. USA 1974, 71, 1250–1254. [Google Scholar] [CrossRef] [PubMed]
- Thomas, K.R.; Capecchi, M.R. Site-directed mutagenesis by gene targeting in mouse embryo-derived stem cells. Cell 1987, 51, 503–512. [Google Scholar] [CrossRef]
- Gauvreau, G.M.; Denburg, J.A. Human mast cell and basophil/eosinophil progenitors. Methods Mol. Biol. 2015, 1220, 59–68. [Google Scholar]
- Lee, J.J.; Jacobsen, E.A.; Ochkur, S.I.; McGarry, M.P.; Condjella, R.M.; Doyle, A.D.; Luo, H.; Zellner, K.R.; Protheroe, C.A.; Willetts, L.; et al. Human versus mouse eosinophils: “that which we call an eosinophil, by any other name would stain as red”. J. Allergy Clin. Immunol. 2012, 130, 572–584. [Google Scholar] [CrossRef]
- Schneider, E.; Thieblemont, N.; De Moraes, M.L.; Dy, M. Basophils: New players in the cytokine network. Eur. Cytokine Netw 2010, 21, 142–153. [Google Scholar]
- Ra, C.; Nunomura, S.; Okayama, Y. Fine-Tuning of Mast Cell Activation by FcepsilonRIbeta Chain. Front. Immunol. 2012, 3, 112. [Google Scholar]
- Jimenez-Saiz, R.; Ellenbogen, Y.; Koenig, J.F.E.; Gordon, M.E.; Walker, T.D.; Rosace, D.; Spill, P.; Bruton, K.; Kong, J.; Monteiro, K.; et al. IgG1(+) B-cell immunity predates IgE responses in epicutaneous sensitization to foods. Allergy 2019, 74, 165–175. [Google Scholar] [CrossRef]
- Bruhns, P.; Iannascoli, B.; England, P.; Mancardi, D.A.; Fernandez, N.; Jorieux, S.; Daeron, M. Specificity and affinity of human Fcgamma receptors and their polymorphic variants for human IgG subclasses. Blood 2009, 113, 3716–3725. [Google Scholar] [CrossRef]
- Maliszewski, C.R.; March, C.J.; Schoenborn, M.A.; Gimpel, S.; Shen, L. Expression cloning of a human Fc receptor for IgA. J. Exp. Med. 1990, 172, 1665–1672. [Google Scholar] [CrossRef] [PubMed]
- Decot, V.; Woerly, G.; Loyens, M.; Loiseau, S.; Quatannens, B.; Capron, M.; Dombrowicz, D. Heterogeneity of expression of IgA receptors by human, mouse, and rat eosinophils. J. Immunol. 2005, 174, 628–635. [Google Scholar] [CrossRef]
- Stelts, D.; Egan, R.W.; Falcone, A.; Garlisi, C.G.; Gleich, G.J.; Kreutner, W.; Kung, T.T.; Nahrebne, D.K.; Chapman, R.W.; Minnicozzi, M. Eosinophils retain their granule major basic protein in a murine model of allergic pulmonary inflammation. Am. J. Respir Cell Mol. Biol. 1998, 18, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Estes, J.D.; Wong, S.W.; Brenchley, J.M. Nonhuman primate models of human viral infections. Nat. Rev. Immunol. 2018, 18, 390–404. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.A.; Royer, C.M.; Pinkerton, K.E.; Schelegle, E.S. Nonhuman Primate Models of Respiratory Disease: Past, Present, and Future. Ilar J. 2017, 58, 269–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stimmer, L.; Fovet, C.M.; Serguera, C. Experimental Models of Autoimmune Demyelinating Diseases in Nonhuman Primates. Vet. Pathol. 2018, 55, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Vallender, E.J.; Miller, G.M. Nonhuman primate models in the genomic era: A paradigm shift. Ilar. J. 2013, 54, 154–165. [Google Scholar] [CrossRef]
- Sato, K.; Oiwa, R.; Kumita, W.; Henry, R.; Sakuma, T.; Ito, R.; Nozu, R.; Inoue, T.; Katano, I.; Sato, K.; et al. Generation of a Nonhuman Primate Model of Severe Combined Immunodeficiency Using Highly Efficient Genome Editing. Cell Stem Cell 2016, 19, 127–138. [Google Scholar] [CrossRef] [Green Version]
- Issacson, J.; Cattanach, B. Report. Mouse News Lett. 1962, 27, 31. [Google Scholar]
- Bosma, G.C.; Custer, R.P.; Bosma, M.J. A severe combined immunodeficiency mutation in the mouse. Nature 1983, 301, 527–530. [Google Scholar] [CrossRef]
- Dick, J.E. Immune-deficient mice as models for human hematopoietic disease. Mol. Genet. Med. 1991, 1, 77–115. [Google Scholar] [PubMed]
- Mosier, D.E. Adoptive transfer of human lymphoid cells to severely immunodeficient mice: Models for normal human immune function, autoimmunity, lymphomagenesis, and AIDS. Adv. Immunol. 1991, 50, 303–325. [Google Scholar] [PubMed]
- Mueller, B.M.; Reisfeld, R.A. Potential of the scid mouse as a host for human tumors. Cancer Metastasis Rev. 1991, 10, 193–200. [Google Scholar] [CrossRef] [PubMed]
- McCune, J.M.; Namikawa, R.; Kaneshima, H.; Shultz, L.D.; Lieberman, M.; Weissman, I.L. The SCID-hu mouse: Murine model for the analysis of human hematolymphoid differentiation and function. Science 1988, 241, 1632–1639. [Google Scholar] [CrossRef] [PubMed]
- Mosier, D.E.; Gulizia, R.J.; Baird, S.M.; Wilson, D.B. Transfer of a functional human immune system to mice with severe combined immunodeficiency. Nature 1988, 335, 256–259. [Google Scholar] [CrossRef] [PubMed]
- Shultz, L.D.; Schweitzer, P.A.; Christianson, S.W.; Gott, B.; Schweitzer, I.B.; Tennent, B.; McKenna, S.; Mobraaten, L.; Rajan, T.V.; Greiner, D.L. Multiple defects in innate and adaptive immunologic function in NOD/LtSz-scid mice. J. Immunol. 1995, 154, 180–191. [Google Scholar] [PubMed]
- Koyanagi, Y.; Tanaka, Y.; Tanaka, R.; Misawa, N.; Kawano, Y.; Tanaka, T.; Miyasaka, M.; Ito, M.; Ueyama, Y.; Yamamoto, N. High levels of viremia in hu-PBL-NOD-scid mice with HIV-1 infection. Leukemia 1997, 11 (Suppl. 3), 109–112. [Google Scholar]
- Ji, H.; Gauguier, D.; Ohmura, K.; Gonzalez, A.; Duchatelle, V.; Danoy, P.; Garchon, H.J.; Degott, C.; Lathrop, M.; Benoist, C.; et al. Genetic influences on the end-stage effector phase of arthritis. J. Exp. Med. 2001, 194, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Wetsel, R.A.; Fleischer, D.T.; Haviland, D.L. Deficiency of the murine fifth complement component (C5). A 2-base pair gene deletion in a 5′-exon. J. Biol. Chem. 1990, 265, 2435–2440. [Google Scholar] [PubMed]
- Legrand, N.; Huntington, N.D.; Nagasawa, M.; Bakker, A.Q.; Schotte, R.; Strick-Marchand, H.; de Geus, S.J.; Pouw, S.M.; Bohne, M.; Voordouw, A.; et al. Functional CD47/signal regulatory protein alpha (SIRP(alpha)) interaction is required for optimal human T- and natural killer- (NK) cell homeostasis in vivo. Proc. Natl. Acad. Sci. USA 2011, 108, 13224–13229. [Google Scholar] [CrossRef]
- Takenaka, K.; Prasolava, T.K.; Wang, J.C.; Mortin-Toth, S.M.; Khalouei, S.; Gan, O.I.; Dick, J.E.; Danska, J.S. Polymorphism in Sirpa modulates engraftment of human hematopoietic stem cells. Nat. Immunol. 2007, 8, 1313–1323. [Google Scholar] [CrossRef] [PubMed]
- Di Santo, J.P.; Kuhn, R.; Muller, W. Common cytokine receptor gamma chain (gamma c)-dependent cytokines: Understanding in vivo functions by gene targeting. Immunol. Rev. 1995, 148, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Leonard, W.J.; Shores, E.W.; Love, P.E. Role of the common cytokine receptor gamma chain in cytokine signaling and lymphoid development. Immunol. Rev. 1995, 148, 97–114. [Google Scholar] [CrossRef] [PubMed]
- Leonard, W.J.; Noguchi, M.; Russell, S.M.; McBride, O.W. The molecular basis of X-linked severe combined immunodeficiency: The role of the interleukin-2 receptor gamma chain as a common gamma chain, gamma c. Immunol. Rev. 1994, 138, 61–86. [Google Scholar] [CrossRef] [PubMed]
- Sugamura, K.; Asao, H.; Kondo, M.; Tanaka, N.; Ishii, N.; Ohbo, K.; Nakamura, M.; Takeshita, T. The interleukin-2 receptor gamma chain: Its role in the multiple cytokine receptor complexes and T cell development in XSCID. Annu. Rev. Immunol. 1996, 14, 179–205. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Hiramatsu, H.; Kobayashi, K.; Suzue, K.; Kawahata, M.; Hioki, K.; Ueyama, Y.; Koyanagi, Y.; Sugamura, K.; Tsuji, K.; et al. NOD/SCID/gamma(c)(null) mouse: An excellent recipient mouse model for engraftment of human cells. Blood 2002, 100, 3175–3182. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, F.; Yasukawa, M.; Lyons, B.; Yoshida, S.; Miyamoto, T.; Yoshimoto, G.; Watanabe, T.; Akashi, K.; Shultz, L.D.; Harada, M. Development of functional human blood and immune systems in NOD/SCID/IL2 receptor {gamma} chain(null) mice. Blood 2005, 106, 1565–1573. [Google Scholar] [CrossRef] [PubMed]
- Hiramatsu, H.; Nishikomori, R.; Heike, T.; Ito, M.; Kobayashi, K.; Katamura, K.; Nakahata, T. Complete reconstitution of human lymphocytes from cord blood CD34+ cells using the NOD/SCID/gammacnull mice model. Blood 2003, 102, 873–880. [Google Scholar] [CrossRef] [PubMed]
- Ito, R.; Katano, I.; Ida-Tanaka, M.; Kamisako, T.; Kawai, K.; Suemizu, H.; Aiso, S.; Ito, M. Efficient xenoengraftment in severe immunodeficient NOD/Shi-scid IL2rgammanull mice is attributed to a lack of CD11c+B220+CD122+ cells. J. Immunol. 2012, 189, 4313–4320. [Google Scholar] [CrossRef] [PubMed]
- Chijiwa, T.; Kawai, K.; Noguchi, A.; Sato, H.; Hayashi, A.; Cho, H.; Shiozawa, M.; Kishida, T.; Morinaga, S.; Yokose, T.; et al. Establishment of patient-derived cancer xenografts in immunodeficient NOG mice. Int. J. Oncol. 2015, 47, 61–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyakawa, Y.; Ohnishi, Y.; Tomisawa, M.; Monnai, M.; Kohmura, K.; Ueyama, Y.; Ito, M.; Ikeda, Y.; Kizaki, M.; Nakamura, M. Establishment of a new model of human multiple myeloma using NOD/SCID/gammac(null) (NOG) mice. Biochem. Biophys. Res. Commun. 2004, 313, 258–262. [Google Scholar] [CrossRef] [PubMed]
- Shultz, L.D.; Goodwin, N.; Ishikawa, F.; Hosur, V.; Lyons, B.L.; Greiner, D.L. Human cancer growth and therapy in immunodeficient mouse models. Cold Spring Harb Protoc 2014, 2014, 694–708. [Google Scholar] [CrossRef] [PubMed]
- Morton, C.L.; Houghton, P.J. Establishment of human tumor xenografts in immunodeficient mice. Nat Protoc. 2007, 2, 247–250. [Google Scholar] [CrossRef] [PubMed]
- Machida, K.; Suemizu, H.; Kawai, K.; Ishikawa, T.; Sawada, R.; Ohnishi, Y.; Tsuchiya, T. Higher susceptibility of NOG mice to xenotransplanted tumors. J. Toxicol. Sci. 2009, 34, 123–127. [Google Scholar] [CrossRef] [Green Version]
- Ito, R.; Katano, I.; Kawai, K.; Hirata, H.; Ogura, T.; Kamisako, T.; Eto, T.; Ito, M. Highly sensitive model for xenogenic GVHD using severe immunodeficient NOG mice. Transplantation 2009, 87, 1654–1658. [Google Scholar] [CrossRef] [PubMed]
- King, M.A.; Covassin, L.; Brehm, M.A.; Racki, W.; Pearson, T.; Leif, J.; Laning, J.; Fodor, W.; Foreman, O.; Burzenski, L.; et al. Human peripheral blood leucocyte non-obese diabetic-severe combined immunodeficiency interleukin-2 receptor gamma chain gene mouse model of xenogeneic graft-versus-host-like disease and the role of host major histocompatibility complex. Clin. Exp. Immunol. 2009, 157, 104–118. [Google Scholar] [CrossRef] [PubMed]
- Brehm, M.A.; Jouvet, N.; Greiner, D.L.; Shultz, L.D. Humanized mice for the study of infectious diseases. Curr. Opin. Immunol. 2013, 25, 428–435. [Google Scholar] [CrossRef] [Green Version]
- Carrillo, M.A.; Zhen, A.; Kitchen, S.G. The Use of the Humanized Mouse Model in Gene Therapy and Immunotherapy for HIV and Cancer. Front. Immunol. 2018, 9, 746. [Google Scholar] [CrossRef]
- Fujiwara, S.; Matsuda, G.; Imadome, K. Humanized mouse models of epstein-barr virus infection and associated diseases. Pathogens 2013, 2, 153–176. [Google Scholar] [CrossRef]
- Munz, C. Humanized mouse models for Epstein Barr virus infection. Curr. Opin. Virol. 2017, 25, 113–118. [Google Scholar] [CrossRef] [Green Version]
- Morton, J.J.; Bird, G.; Refaeli, Y.; Jimeno, A. Humanized Mouse Xenograft Models: Narrowing the Tumor-Microenvironment Gap. Cancer Res. 2016, 76, 6153–6158. [Google Scholar] [CrossRef] [PubMed]
- Satheesan, S.; Li, H.; Burnett, J.C.; Takahashi, M.; Li, S.; Wu, S.X.; Synold, T.W.; Rossi, J.J.; Zhou, J. HIV Replication and Latency in a Humanized NSG Mouse Model during Suppressive Oral Combinational Antiretroviral Therapy. J. Virol. 2018, 92, e02118-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De La Rochere, P.; Guil-Luna, S.; Decaudin, D.; Azar, G.; Sidhu, S.S.; Piaggio, E. Humanized Mice for the Study of Immuno-Oncology. Trends Immunol. 2018, 39, 748–763. [Google Scholar] [CrossRef] [PubMed]
- Takagi, S.; Saito, Y.; Hijikata, A.; Tanaka, S.; Watanabe, T.; Hasegawa, T.; Mochizuki, S.; Kunisawa, J.; Kiyono, H.; Koseki, H.; et al. Membrane-bound human SCF/KL promotes in vivo human hematopoietic engraftment and myeloid differentiation. Blood 2012, 119, 2768–2777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Billerbeck, E.; Barry, W.T.; Mu, K.; Dorner, M.; Rice, C.M.; Ploss, A. Development of human CD4+FoxP3+ regulatory T cells in human stem cell factor-, granulocyte-macrophage colony-stimulating factor-, and interleukin-3-expressing NOD-SCID IL2Rgamma(null) humanized mice. Blood 2011, 117, 3076–3086. [Google Scholar] [CrossRef] [PubMed]
- Rongvaux, A.; Willinger, T.; Martinek, J.; Strowig, T.; Gearty, S.V.; Teichmann, L.L.; Saito, Y.; Marches, F.; Halene, S.; Palucka, A.K.; et al. Development and function of human innate immune cells in a humanized mouse model. Nat. Biotechnol. 2014, 32, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Ito, R.; Takahashi, T.; Katano, I.; Kawai, K.; Kamisako, T.; Ogura, T.; Ida-Tanaka, M.; Suemizu, H.; Nunomura, S.; Ra, C.; et al. Establishment of a human allergy model using human IL-3/GM-CSF-transgenic NOG mice. J. Immunol. 2013, 191, 2890–2899. [Google Scholar] [CrossRef] [PubMed]
- Hanazawa, A.; Ito, R.; Katano, I.; Kawai, K.; Goto, M.; Suemizu, H.; Kawakami, Y.; Ito, M.; Takahashi, T. Generation of Human Immunosuppressive Myeloid Cell Populations in Human Interleukin-6 Transgenic NOG Mice. Front. Immunol. 2018, 9, 152. [Google Scholar] [CrossRef] [Green Version]
- Brehm, M.A.; Racki, W.J.; Leif, J.; Burzenski, L.; Hosur, V.; Wetmore, A.; Gott, B.; Herlihy, M.; Ignotz, R.; Dunn, R.; et al. Engraftment of human HSCs in nonirradiated newborn NOD-scid IL2rgamma null mice is enhanced by transgenic expression of membrane-bound human SCF. Blood 2012, 119, 2778–2788. [Google Scholar] [CrossRef]
- Herndler-Brandstetter, D.; Shan, L.; Yao, Y.; Stecher, C.; Plajer, V.; Lietzenmayer, M.; Strowig, T.; de Zoete, M.R.; Palm, N.W.; Chen, J.; et al. Humanized mouse model supports development, function, and tissue residency of human natural killer cells. Proc. Natl. Acad. Sci. USA 2017, 114, E9626–E9634. [Google Scholar] [CrossRef] [Green Version]
- Katano, I.; Takahashi, T.; Ito, R.; Kamisako, T.; Mizusawa, T.; Ka, Y.; Ogura, T.; Suemizu, H.; Kawakami, Y.; Ito, M. Predominant development of mature and functional human NK cells in a novel human IL-2-producing transgenic NOG mouse. J. Immunol. 2015, 194, 3513–3525. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Di Santo, J.P. Probing Human NK Cell Biology Using Human Immune System (HIS) Mice. Curr. Top Microbiol. Immunol. 2016, 395, 191–208. [Google Scholar] [PubMed]
- Shultz, L.D.; Saito, Y.; Najima, Y.; Tanaka, S.; Ochi, T.; Tomizawa, M.; Doi, T.; Sone, A.; Suzuki, N.; Fujiwara, H.; et al. Generation of functional human T-cell subsets with HLA-restricted immune responses in HLA class I expressing NOD/SCID/IL2r gamma(null) humanized mice. Proc. Natl. Acad. Sci. USA 2010, 107, 13022–13027. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Takahashi, T.; Katano, I.; Ito, R.; Ito, M.; Harigae, H.; Ishii, N.; Sugamura, K. Induction of human humoral immune responses in a novel HLA-DR-expressing transgenic NOD/Shi-scid/gammacnull mouse. Int. Immunol. 2012, 24, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Ito, R.; Takahashi, T.; Ito, M. Humanized mouse models: Application to human diseases. J. Cell. Physiol. 2018, 233, 3723–3728. [Google Scholar] [CrossRef]
- Theocharides, A.P.; Rongvaux, A.; Fritsch, K.; Flavell, R.A.; Manz, M.G. Humanized hemato-lymphoid system mice. Haematologica 2016, 101, 5–19. [Google Scholar] [CrossRef]
- Walsh, N.C.; Kenney, L.L.; Jangalwe, S.; Aryee, K.E.; Greiner, D.L.; Brehm, M.A.; Shultz, L.D. Humanized Mouse Models of Clinical Disease. Annu. Rev. Pathol. 2017, 12, 187–215. [Google Scholar] [CrossRef] [Green Version]
- Akkina, R. New generation humanized mice for virus research: Comparative aspects and future prospects. Virology 2013, 435, 14–28. [Google Scholar] [CrossRef]
- Lemmermann, N.A.; Reddehase, M.J. Refining human T-cell immunotherapy of cytomegalovirus disease: A mouse model with ‘humanized’ antigen presentation as a new preclinical study tool. Med. Microbiol. Immunol. 2016, 205, 549–561. [Google Scholar] [CrossRef]
- Rongvaux, A.; Takizawa, H.; Strowig, T.; Willinger, T.; Eynon, E.E.; Flavell, R.A.; Manz, M.G. Human hemato-lymphoid system mice: Current use and future potential for medicine. Annu. Rev. Immunol. 2013, 31, 635–674. [Google Scholar] [CrossRef]
- Wunderlich, M.; Chou, F.S.; Link, K.A.; Mizukawa, B.; Perry, R.L.; Carroll, M.; Mulloy, J.C. AML xenograft efficiency is significantly improved in NOD/SCID-IL2RG mice constitutively expressing human SCF, GM-CSF and IL-3. Leukemia 2010, 24, 1785–1788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimi, A.; Balasis, M.E.; Vedder, A.; Feldman, K.; Ma, Y.; Zhang, H.; Lee, S.C.; Letson, C.; Niyongere, S.; Lu, S.X.; et al. Robust patient-derived xenografts of MDS/MPN overlap syndromes capture the unique characteristics of CMML and JMML. Blood 2017, 130, 397–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spengler, J.R.; Lavender, K.J.; Martellaro, C.; Carmody, A.; Kurth, A.; Keck, J.G.; Saturday, G.; Scott, D.P.; Nichol, S.T.; Hasenkrug, K.J.; et al. Ebola Virus Replication and Disease Without Immunopathology in Mice Expressing Transgenes to Support Human Myeloid and Lymphoid Cell Engraftment. J. Infect Dis. 2016, 214 (Suppl. 3), S308–S318. [Google Scholar] [CrossRef]
- Bryce, P.J.; Falahati, R.; Kenney, L.L.; Leung, J.; Bebbington, C.; Tomasevic, N.; Krier, R.A.; Hsu, C.L.; Shultz, L.D.; Greiner, D.L.; et al. Humanized mouse model of mast cell-mediated passive cutaneous anaphylaxis and passive systemic anaphylaxis. J. Allergy. Clin. Immunol. 2016, 138, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Wunderlich, M.; Mulloy, J.C. MISTRG extends PDX modeling to favorable AMLs. Blood 2016, 128, 2111–2112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Rongvaux, A.; Taylor, A.; Jiang, T.; Tebaldi, T.; Balasubramanian, K.; Bagale, A.; Terzi, Y.K.; Gbyli, R.; Wang, X.; et al. A highly efficient and faithful MDS patient-derived xenotransplantation model for pre-clinical studies. Nat. Commun. 2019, 10, 366. [Google Scholar] [CrossRef]
- Saito, Y.; Ellegast, J.M.; Rafiei, A.; Song, Y.; Kull, D.; Heikenwalder, M.; Rongvaux, A.; Halene, S.; Flavell, R.A.; Manz, M.G. Peripheral blood CD34(+) cells efficiently engraft human cytokine knock-in mice. Blood 2016, 128, 1829–1833. [Google Scholar] [CrossRef] [PubMed]
- Perdomo-Celis, F.; Medina-Moreno, S.; Davis, H.; Bryant, J.; Zapata, J.C. HIV Replication in Humanized IL-3/GM-CSF-Transgenic NOG Mice. Pathogens 2019, 8, 33. [Google Scholar] [CrossRef]
- Burton, O.T.; Stranks, A.J.; Tamayo, J.M.; Koleoglou, K.J.; Schwartz, L.B.; Oettgen, H.C. A humanized mouse model of anaphylactic peanut allergy. J. Allergy Clin. Immunol. 2017, 139, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wilson, S.J.; Kuei, C.; Lovenberg, T.W. Comparison of human, mouse, rat, and guinea pig histamine H4 receptors reveals substantial pharmacological species variation. J. Pharm. Exp. 2001, 299, 121–130. [Google Scholar]
- Maekawa, A.; Kanaoka, Y.; Lam, B.K.; Austen, K.F. Identification in mice of two isoforms of the cysteinyl leukotriene 1 receptor that result from alternative splicing. Proc. Natl. Acad. Sci. USA 2001, 98, 2256–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duez, C.; Tsicopoulos, A.; Janin, A.; Tillie-Leblond, I.; Thyphronitis, G.; Marquillies, P.; Hamid, Q.; Wallaert, B.; Tonnel, A.B.; Pestel, J. An in vivo model of allergic inflammation: Pulmonary human cell infiltrate in allergen-challenged allergic Hu-SCID mice. Eur. J. Immunol. 1996, 26, 1088–1093. [Google Scholar] [CrossRef] [PubMed]
- Herz, U.; Botchkarev, V.A.; Paus, R.; Renz, H. Increased airway responsiveness, allergy-type-I skin responses and systemic anaphylaxis in a humanized-severe combined immuno-deficiency mouse model. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2004, 34, 478–487. [Google Scholar] [CrossRef]
- Weigmann, B.; Schughart, N.; Wiebe, C.; Sudowe, S.; Lehr, H.A.; Jonuleit, H.; Vogel, L.; Becker, C.; Neurath, M.F.; Grabbe, S.; et al. Allergen-induced IgE-dependent gut inflammation in a human PBMC-engrafted murine model of allergy. J. Allergy Clin. Immunol. 2012, 129, 1126–1135. [Google Scholar] [CrossRef] [PubMed]
- Nolte, T.; Zadeh-Khorasani, M.; Safarov, O.; Rueff, F.; Varga, R.; Herbach, N.; Wanke, R.; Wollenberg, A.; Mueller, T.; Gropp, R.; et al. Induction of oxazolone-mediated features of atopic dermatitis in NOD-scid IL2Rgamma(null) mice engrafted with human peripheral blood mononuclear cells. Dis. Model. Mech. 2013, 6, 125–134. [Google Scholar] [CrossRef]
- Pagovich, O.E.; Wang, B.; Chiuchiolo, M.J.; Kaminsky, S.M.; Sondhi, D.; Jose, C.L.; Price, C.C.; Brooks, S.F.; Mezey, J.G.; Crystal, R.G. Anti-hIgE gene therapy of peanut-induced anaphylaxis in a humanized murine model of peanut allergy. J. Allergy Clin. Immunol. 2016, 138, 1652–1662. [Google Scholar] [CrossRef]
- Martin, H.; Reuter, S.; Dehzad, N.; Heinz, A.; Bellinghausen, I.; Saloga, J.; Haasler, I.; Korn, S.; Jonuleit, H.; Buhl, R.; et al. CD4-mediated regulatory T-cell activation inhibits the development of disease in a humanized mouse model of allergic airway disease. J. Allergy Clin. Immunol. 2012, 129, 521–528. [Google Scholar] [CrossRef]
- Bellinghausen, I.; Saloga, J. Analysis of allergic immune responses in humanized mice. Cell Immunol. 2016, 308, 7–12. [Google Scholar] [CrossRef]
- Eschborn, M.; Weigmann, B.; Reissig, S.; Waisman, A.; Saloga, J.; Bellinghausen, I. Activated glycoprotein A repetitions predominant (GARP)-expressing regulatory T cells inhibit allergen-induced intestinal inflammation in humanized mice. J. Allergy Clin. Immunol. 2015, 136, 159–168. [Google Scholar] [CrossRef]
- Cabanillas, B.; Brehler, A.C.; Novak, N. Atopic dermatitis phenotypes and the need for personalized medicine. Curr. Opin. Allergy Clin. Immunol. 2017, 17, 309–315. [Google Scholar] [CrossRef] [Green Version]
- David Boothe, W.; Tarbox, J.A.; Tarbox, M.B. Atopic Dermatitis: Pathophysiology. Adv. Exp. Med. Biol. 2017, 1027, 21–37. [Google Scholar] [PubMed]
- Nakajima, S.; Nomura, T.; Common, J.; Kabashima, K. Insights into atopic dermatitis gained from genetically defined mouse models. J. Allergy Clin. Immunol. 2019, 143, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Silverberg, J.I.; Kantor, R. The Role of Interleukins 4 and/or 13 in the Pathophysiology and Treatment of Atopic Dermatitis. Derm. Clin. 2017, 35, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Man, M.Q.; Hatano, Y.; Lee, S.H.; Man, M.; Chang, S.; Feingold, K.R.; Leung, D.Y.; Holleran, W.; Uchida, Y.; Elias, P.M. Characterization of a hapten-induced, murine model with multiple features of atopic dermatitis: Structural, immunologic, and biochemical changes following single versus multiple oxazolone challenges. J. Invest. Derm. 2008, 128, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Mizukoshi, K.; Oyobikawa, M.; Ohshima, H.; Tagami, H. Establishment of an atopic dermatitis-like skin model in a hairless mouse by repeated elicitation of contact hypersensitivity that enables to conduct functional analyses of the stratum corneum with various non-invasive biophysical instruments. Ski. Res. Technol. 2004, 10, 122–129. [Google Scholar] [CrossRef]
- Jin, H.; He, R.; Oyoshi, M.; Geha, R.S. Animal models of atopic dermatitis. J. Invest. Derm. 2009, 129, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Berin, M.C.; Sampson, H.A. Food allergy: An enigmatic epidemic. Trends Immunol 2013, 34, 390–397. [Google Scholar] [CrossRef]
- Jones, S.M.; Burks, A.W. Food Allergy. N Engl. J. Med. 2017, 377, 1168–1176. [Google Scholar] [CrossRef]
- Reber, L.L.; Hernandez, J.D.; Galli, S.J. The pathophysiology of anaphylaxis. J. Allergy Clin. Immunol. 2017, 140, 335–348. [Google Scholar] [CrossRef]
- Yue, D.; Ciccolini, A.; Avilla, E.; Waserman, S. Food allergy and anaphylaxis. J. Asthma Allergy 2018, 11, 111–120. [Google Scholar] [CrossRef]
- Finkelman, F.D. Anaphylaxis: Lessons from mouse models. J. Allergy Clin. Immunol. 2007, 120, 506–515, quiz 516-5177. [Google Scholar] [CrossRef] [PubMed]
- Finkelman, F.D.; Rothenberg, M.E.; Brandt, E.B.; Morris, S.C.; Strait, R.T. Molecular mechanisms of anaphylaxis: Lessons from studies with murine models. J. Allergy Clin. Immunol. 2005, 115, 449–457, quiz 458. [Google Scholar] [CrossRef] [PubMed]
- Strait, R.T.; Morris, S.C.; Yang, M.; Qu, X.W.; Finkelman, F.D. Pathways of anaphylaxis in the mouse. J. Allergy Clin. Immunol. 2002, 109, 658–668. [Google Scholar] [CrossRef] [PubMed]
- Ito, R.; Katano, I.; Kawai, K.; Yagoto, M.; Takahashi, T.; Ka, Y.; Ogura, T.; Takahashi, R.; Ito, M. A Novel Xenogeneic Graft-Versus-Host Disease Model for Investigating the Pathological Role of Human CD4+ or CD8+ T Cells Using Immunodeficient NOG Mice. Am. J. Transpl. 2017, 17, 1216–1228. [Google Scholar] [CrossRef] [PubMed]
- Brainard, D.M.; Seung, E.; Frahm, N.; Cariappa, A.; Bailey, C.C.; Hart, W.K.; Shin, H.S.; Brooks, S.F.; Knight, H.L.; Eichbaum, Q.; et al. Induction of robust cellular and humoral virus-specific adaptive immune responses in human immunodeficiency virus-infected humanized BLT mice. J. Virol. 2009, 83, 7305–7321. [Google Scholar] [CrossRef] [PubMed]
- Denton, P.W.; Estes, J.D.; Sun, Z.; Othieno, F.A.; Wei, B.L.; Wege, A.K.; Powell, D.A.; Payne, D.; Haase, A.T.; Garcia, J.V. Antiretroviral pre-exposure prophylaxis prevents vaginal transmission of HIV-1 in humanized BLT mice. Plos Med. 2008, 5, e16. [Google Scholar] [CrossRef] [PubMed]
- Wege, A.K.; Melkus, M.W.; Denton, P.W.; Estes, J.D.; Garcia, J.V. Functional and phenotypic characterization of the humanized BLT mouse model. Curr. Top Microbiol. Immunol. 2008, 324, 149–165. [Google Scholar]
- Lan, P.; Tonomura, N.; Shimizu, A.; Wang, S.; Yang, Y.G. Reconstitution of a functional human immune system in immunodeficient mice through combined human fetal thymus/liver and CD34+ cell transplantation. Blood 2006, 108, 487–492. [Google Scholar] [CrossRef] [Green Version]
- Fattouh, R.; Pouladi, M.A.; Alvarez, D.; Johnson, J.R.; Walker, T.D.; Goncharova, S.; Inman, M.D.; Jordana, M. House dust mite facilitates ovalbumin-specific allergic sensitization and airway inflammation. Am. J. Respir. Crit. Care Med. 2005, 172, 314–321. [Google Scholar] [CrossRef]
- Johnson, J.R.; Wiley, R.E.; Fattouh, R.; Swirski, F.K.; Gajewska, B.U.; Coyle, A.J.; Gutierrez-Ramos, J.C.; Ellis, R.; Inman, M.D.; Jordana, M. Continuous exposure to house dust mite elicits chronic airway inflammation and structural remodeling. Am. J. Respir. Crit. Care Med. 2004, 169, 378–385. [Google Scholar] [CrossRef]
- Bartemes, K.R.; Kita, H. Innate and adaptive immune responses to fungi in the airway. J. Allergy Clin. Immunol. 2018, 142, 353–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, S.K.; Leung, D.Y.M. Dog and Cat Allergies: Current State of Diagnostic Approaches and Challenges. Allergy Asthma Immunol. Res. 2018, 10, 97–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takkouche, B.; Gonzalez-Barcala, F.J.; Etminan, M.; Fitzgerald, M. Exposure to furry pets and the risk of asthma and allergic rhinitis: A meta-analysis. Allergy 2008, 63, 857–864. [Google Scholar] [CrossRef] [PubMed]
- Wiesner, D.L.; Klein, B.S. Lung epithelium: Barrier immunity to inhaled fungi and driver of fungal-associated allergic asthma. Curr. Opin. Microbiol. 2017, 40, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Sonar, S.S.; Hsu, Y.M.; Conrad, M.L.; Majeau, G.R.; Kilic, A.; Garber, E.; Gao, Y.; Nwankwo, C.; Willer, G.; Dudda, J.C.; et al. Antagonism of TIM-1 blocks the development of disease in a humanized mouse model of allergic asthma. J. Clin. Invest. 2010, 120, 2767–2781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christianson, C.A.; Goplen, N.P.; Zafar, I.; Irvin, C.; Good, J.T., Jr.; Rollins, D.R.; Gorentla, B.; Liu, W.; Gorska, M.M.; Chu, H.; et al. Persistence of asthma requires multiple feedback circuits involving type 2 innate lymphoid cells and IL-33. J. Allergy Clin. Immunol. 2015, 136, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, N.; Nabe, T.; Yoshino, S. IL-17A promotes the exacerbation of IL-33-induced airway hyperresponsiveness by enhancing neutrophilic inflammation via CXCR2 signaling in mice. J. Immunol. 2014, 192, 1372–1384. [Google Scholar] [CrossRef] [PubMed]
- Nagarkar, D.R.; Ramirez-Carrozzi, V.; Choy, D.F.; Lee, K.; Soriano, R.; Jia, G.; Abbas, A.R.; Modrusan, Z.; Pappu, R.; Arron, J.R. IL-13 mediates IL-33-dependent mast cell and type 2 innate lymphoid cell effects on bronchial epithelial cells. J. Allergy Clin. Immunol. 2015, 136, 202–205. [Google Scholar] [CrossRef]
- Snelgrove, R.J.; Gregory, L.G.; Peiro, T.; Akthar, S.; Campbell, G.A.; Walker, S.A.; Lloyd, C.M. Alternaria-derived serine protease activity drives IL-33-mediated asthma exacerbations. J. Allergy Clin. Immunol. 2014, 134, 583–592. [Google Scholar] [CrossRef]
- Ito, R.; Maruoka, S.; Soda, K.; Katano, I.; Kawai, K.; Yagoto, M.; Hanazawa, A.; Takahashi, T.; Ogura, T.; Goto, M.; et al. A humanized mouse model to study asthmatic airway inflammation via the human IL-33/IL-13 axis. Jci. Insight 2018, 3, 121580. [Google Scholar] [CrossRef]
- Shikotra, A.; Choy, D.F.; Ohri, C.M.; Doran, E.; Butler, C.; Hargadon, B.; Shelley, M.; Abbas, A.R.; Austin, C.D.; Jackman, J.; et al. Increased expression of immunoreactive thymic stromal lymphopoietin in patients with severe asthma. J. Allergy Clin. Immunol. 2012, 129, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Ashizawa, T.; Iizuka, A.; Nonomura, C.; Kondou, R.; Maeda, C.; Miyata, H.; Sugino, T.; Mitsuya, K.; Hayashi, N.; Nakasu, Y.; et al. Antitumor Effect of Programmed Death-1 (PD-1) Blockade in Humanized the NOG-MHC Double Knockout Mouse. Clin. Cancer Res. 2017, 23, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Kametani, Y.; Katano, I.; Miyamoto, A.; Kikuchi, Y.; Ito, R.; Muguruma, Y.; Tsuda, B.; Habu, S.; Tokuda, Y.; Ando, K.; et al. NOG-hIL-4-Tg, a new humanized mouse model for producing tumor antigen-specific IgG antibody by peptide vaccination. Plos ONE 2017, 12, e0179239. [Google Scholar] [CrossRef] [PubMed]
- Yoshihara, S.; Li, Y.; Xia, J.; Danzl, N.; Sykes, M.; Yang, Y.G. Posttransplant Hemophagocytic Lymphohistiocytosis Driven by Myeloid Cytokines and Vicious Cycles of T-Cell and Macrophage Activation in Humanized Mice. Front. Immunol. 2019, 10, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oldenborg, P.A.; Gresham, H.D.; Lindberg, F.P. CD47-signal regulatory protein alpha (SIRPalpha) regulates Fcgamma and complement receptor-mediated phagocytosis. J. Exp. Med. 2001, 193, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Oldenborg, P.A.; Zheleznyak, A.; Fang, Y.F.; Lagenaur, C.F.; Gresham, H.D.; Lindberg, F.P. Role of CD47 as a marker of self on red blood cells. Science 2000, 288, 2051–2054. [Google Scholar] [CrossRef] [PubMed]
- Hammad, H.; Chieppa, M.; Perros, F.; Willart, M.A.; Germain, R.N.; Lambrecht, B.N. House dust mite allergen induces asthma via Toll-like receptor 4 triggering of airway structural cells. Nat. Med. 2009, 15, 410–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, R.; Takahashi, T.; Katano, I.; Ito, M. Current advances in humanized mouse models. Cell Mol. Immunol. 2012, 9, 208–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitoma, H.; Horiuchi, T.; Tsukamoto, H.; Tamimoto, Y.; Kimoto, Y.; Uchino, A.; To, K.; Harashima, S.; Hatta, N.; Harada, M. Mechanisms for cytotoxic effects of anti-tumor necrosis factor agents on transmembrane tumor necrosis factor alpha-expressing cells: Comparison among infliximab, etanercept, and adalimumab. Arthritis Rheum. 2008, 58, 1248–1257. [Google Scholar] [CrossRef]
- Wechsler, M.E. Inhibiting interleukin-4 and interleukin-13 in difficult-to-control asthma. N Engl. J. Med. 2013, 368, 2511–2513. [Google Scholar] [CrossRef]
- Haldar, P.; Brightling, C.E.; Hargadon, B.; Gupta, S.; Monteiro, W.; Sousa, A.; Marshall, R.P.; Bradding, P.; Green, R.H.; Wardlaw, A.J.; et al. Mepolizumab and exacerbations of refractory eosinophilic asthma. N Engl. J. Med. 2009, 360, 973–984. [Google Scholar] [CrossRef] [PubMed]
- FitzGerald, J.M.; Bleecker, E.R.; Nair, P.; Korn, S.; Ohta, K.; Lommatzsch, M.; Ferguson, G.T.; Busse, W.W.; Barker, P.; Sproule, S.; et al. Benralizumab, an anti-interleukin-5 receptor alpha monoclonal antibody, as add-on treatment for patients with severe, uncontrolled, eosinophilic asthma (CALIMA): A randomised, double-blind, placebo-controlled phase 3 trial. Lancet 2016, 388, 2128–2141. [Google Scholar] [CrossRef]
- Matlawska-Wasowska, K.; Ward, E.; Stevens, S.; Wang, Y.; Herbst, R.; Winter, S.S.; Wilson, B.S. Macrophage and NK-mediated killing of precursor-B acute lymphoblastic leukemia cells targeted with a-fucosylated anti-CD19 humanized antibodies. Leukemia 2013, 27, 1263–1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murata, Y.; Tanaka, D.; Hazama, D.; Yanagita, T.; Saito, Y.; Kotani, T.; Oldenborg, P.A.; Matozaki, T. Anti-human SIRPalpha antibody is a new tool for cancer immunotherapy. Cancer Sci. 2018, 109, 1300–1308. [Google Scholar] [CrossRef] [PubMed]
- Miyata, J.; Fukunaga, K.; Kawashima, Y.; Watanabe, T.; Saitoh, A.; Hirosaki, T.; Araki, Y.; Kikawada, T.; Betsuyaku, T.; Ohara, O.; et al. Dysregulated fatty acid metabolism in nasal polyp-derived eosinophils from patients with chronic rhinosinusitis. Allergy 2019. Jan 22. [Google Scholar] [CrossRef] [PubMed]


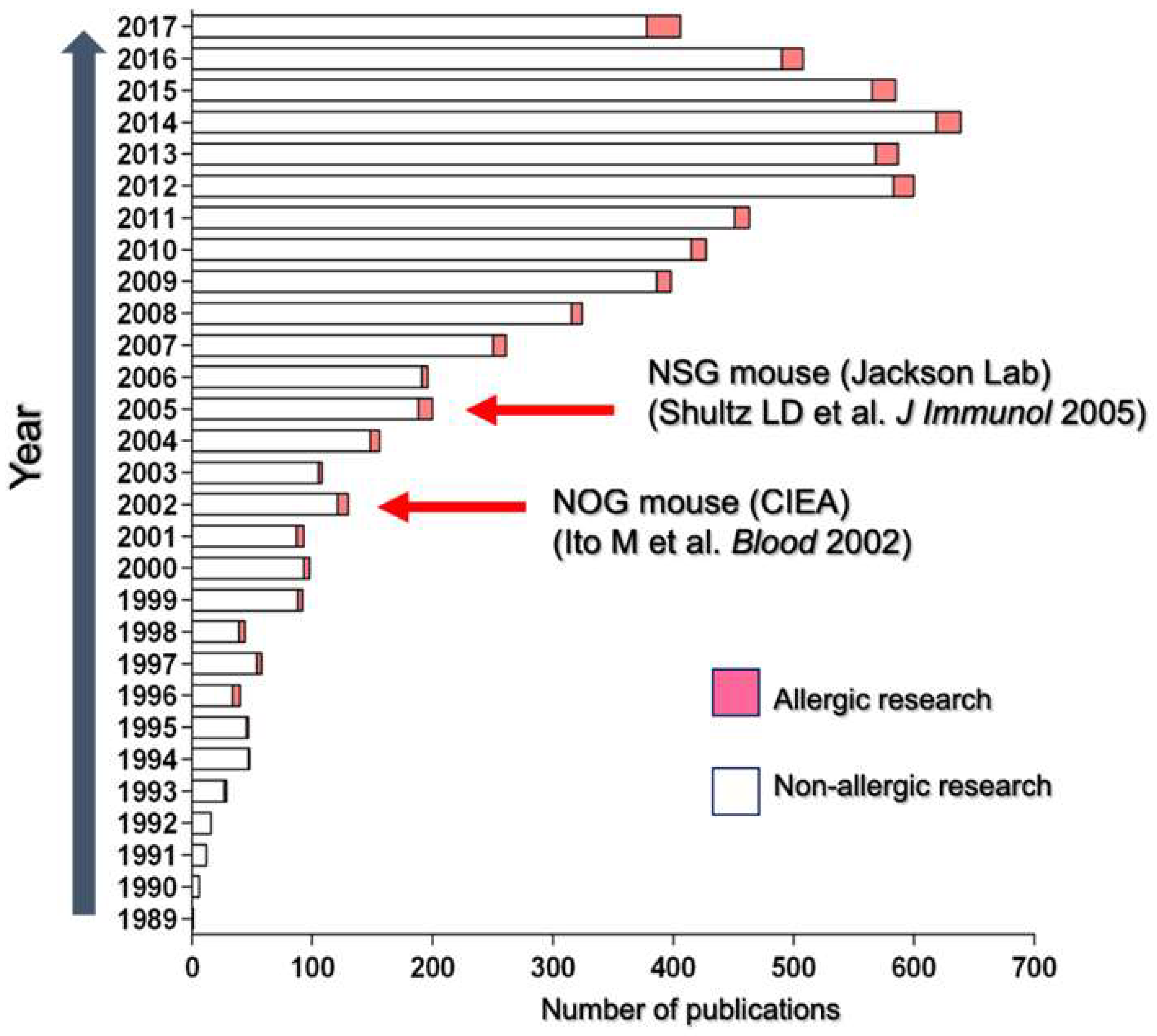{kind=link}
| Strain | NOG IL-3/GM-CSF Tg | NSG-SGM3 (Tg) | MISTRG (knockin) | NSG-hSCF Tg | |
| Transgene | human IL-3. GM-CSF | human IL-3, GM-CSF, SCF | human IL-3, GM-CSF, M-CSF, TPO, SIRPa ** | membrane-bound human SCF | |
| Expression level | 30-100 pg/ml | 2,000-4,000 pg/ml * | Not reported | Not reported | |
| Level of human myeloid cells compared to conventional humanized mice | Mono/Macrophage | Increased | Increased | Increased | No difference |
| Granulocyte | Eo/Baso increased but Neutro did not | Eo/Baso increased but Neutro did not | Eo/Baso increased but Neutro did not | No difference | |
| Mast cell | Increased | Increased | Not reported | Increased | |
| Erythrocyte | None | None | None | None | |
| Platelet | Slightly increased | Decreased | Not reported | Not reported | |
| Human disease model | Immune diseases | Allergy, Asthma, Adverse effect of immunotherapy or chemotherapy | Allergy, Macrophage activation syndrome | Not reported | Allergy |
| PDX | Not reported | AML, MDS, MML | AML, MDS, MM | Not reported | |
| Infection | HIV | Ebora | Listeria, Influenza, | Not reported | |
| Original publication | 57 | 55 | 56 | 54, 59 | |
| Strain | Graft | Induction | Treatment | Reference |
|---|---|---|---|---|
| NSG | Patient-derived PBMC | Hazelnut-induced allergic gut or airway | - | 84 |
| NSG | Patient-derived PBMC | Oxazolone-induced atopic dermatitis | - | 85 |
| NSG | Patient-derived PBMC | Allergen-induced allergic gut inflammation | Omalizumab, rGARP, Treg | 89 |
| NSG | Patient-derived PBMC | Peanut-induced PCA and PSA | Omalizumab | 86 |
| NSG | Patient-derived PBMC | Birch pollen-induced airway inflammation | gp120 | 87 |
| NOG IL-3/GM-CSF Tg | HSC | hIgE+antigen-induced PCA | C3 antagonist | 57 |
| NOG IL-3/GM-CSF Tg | HSC | IL-33-induced airway inflammation | anti-hIL-13 Ab | 104 |
| NSG-SGM3 | BLT | hIgE+antigen-induced PCA and PSA | - | 74 |
| NSG-hSCF Tg | HSC | Peanut-induced PSA | Omalizumab | 79 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ito, R.; Maruoka, S.; Gon, Y.; Katano, I.; Takahashi, T.; Ito, M.; Izuhara, K.; Nunomura, S. Recent Advances in Allergy Research Using Humanized Mice. Int. J. Mol. Sci. 2019, 20, 2740. https://doi.org/10.3390/ijms20112740
Ito R, Maruoka S, Gon Y, Katano I, Takahashi T, Ito M, Izuhara K, Nunomura S. Recent Advances in Allergy Research Using Humanized Mice. International Journal of Molecular Sciences. 2019; 20(11):2740. https://doi.org/10.3390/ijms20112740
Chicago/Turabian StyleIto, Ryoji, Shuichiro Maruoka, Yasuhiro Gon, Ikumi Katano, Takeshi Takahashi, Mamoru Ito, Kenji Izuhara, and Satoshi Nunomura. 2019. "Recent Advances in Allergy Research Using Humanized Mice" International Journal of Molecular Sciences 20, no. 11: 2740. https://doi.org/10.3390/ijms20112740