
Zoledronate Causes a Systemic Shift of Macrophage Polarization towards M1 In Vivo

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. General Considerations
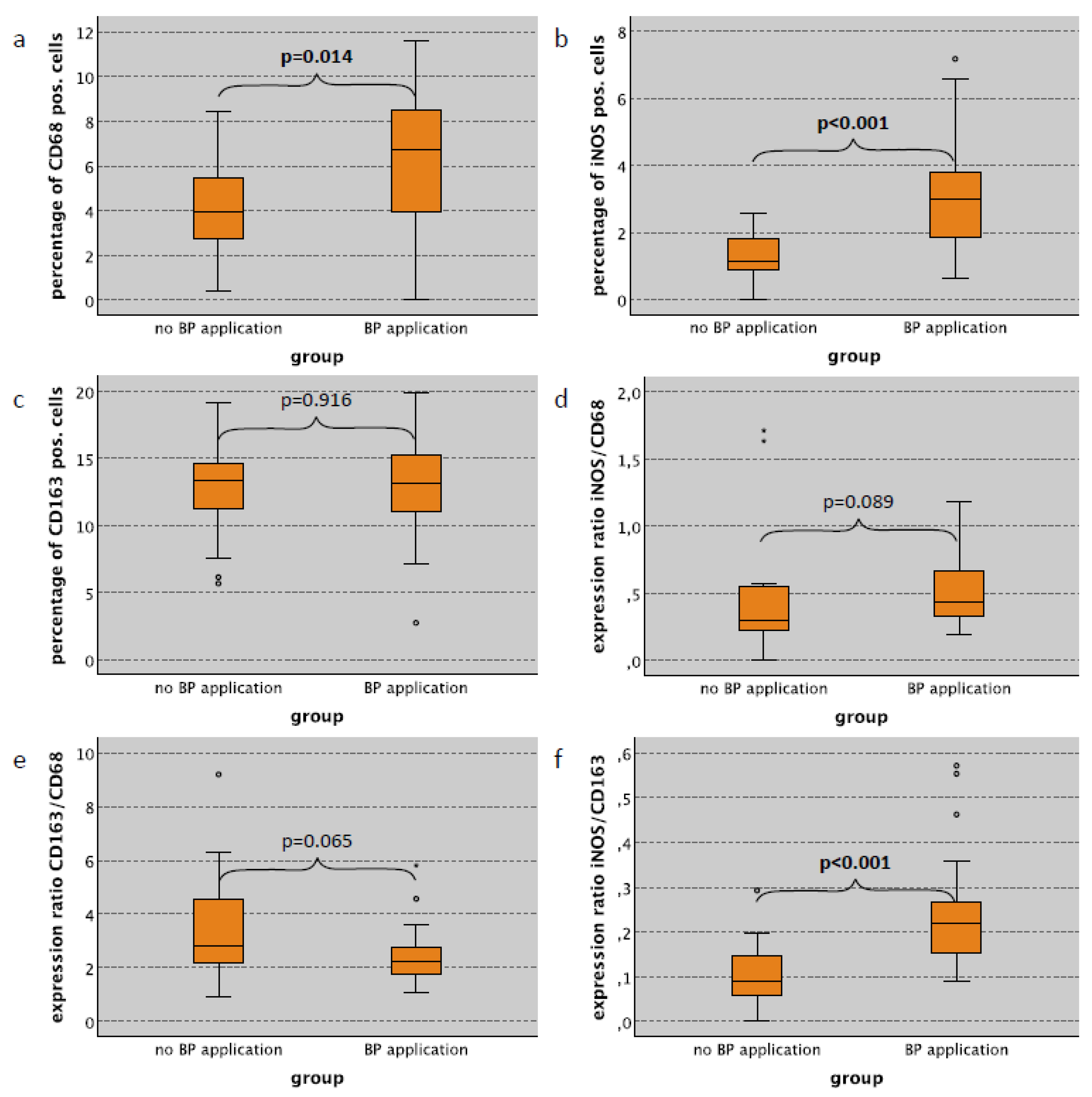
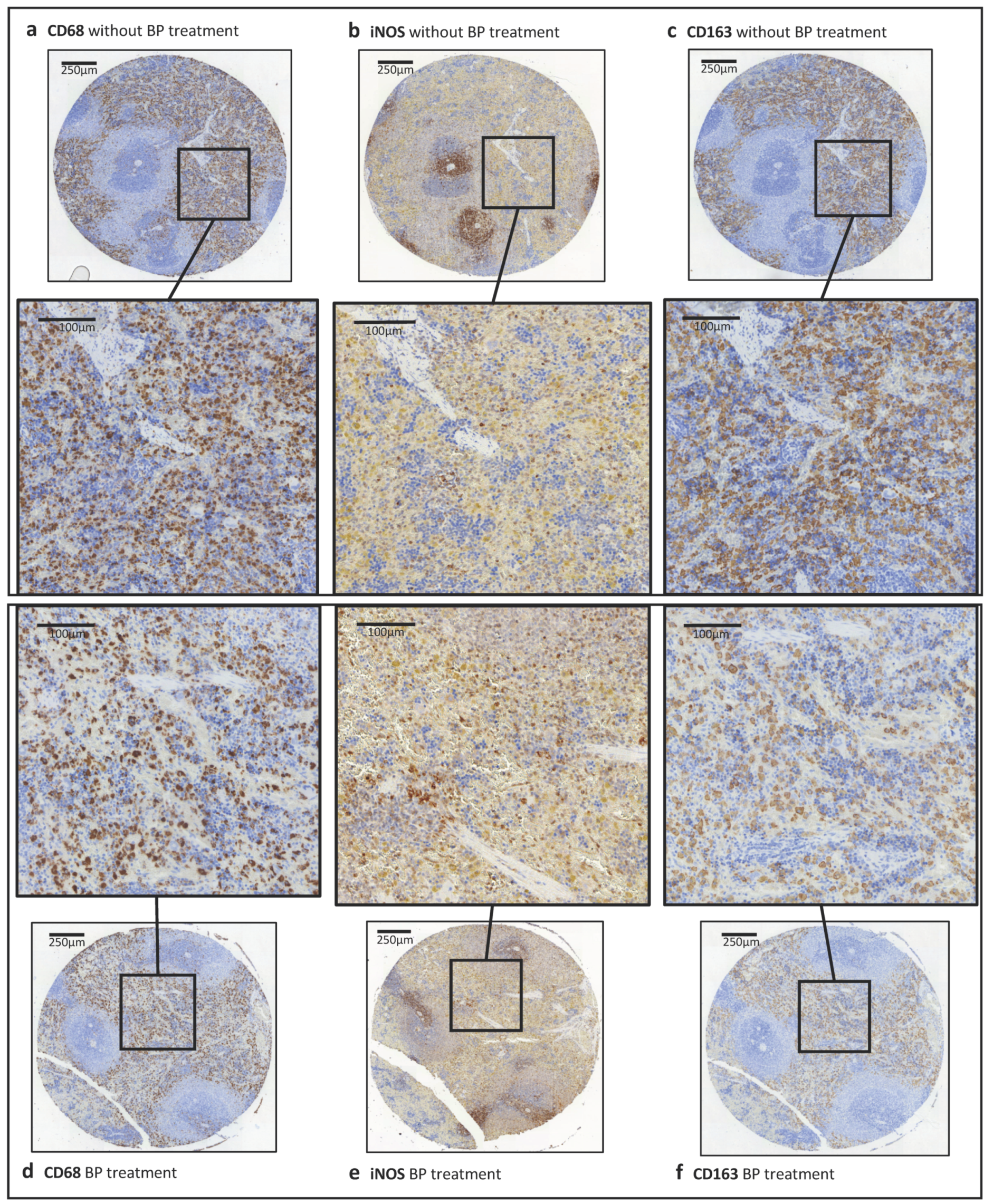
2.2. Macrophage Infiltration and Polarization in the Skin in Relation to Bisphosphonate Application
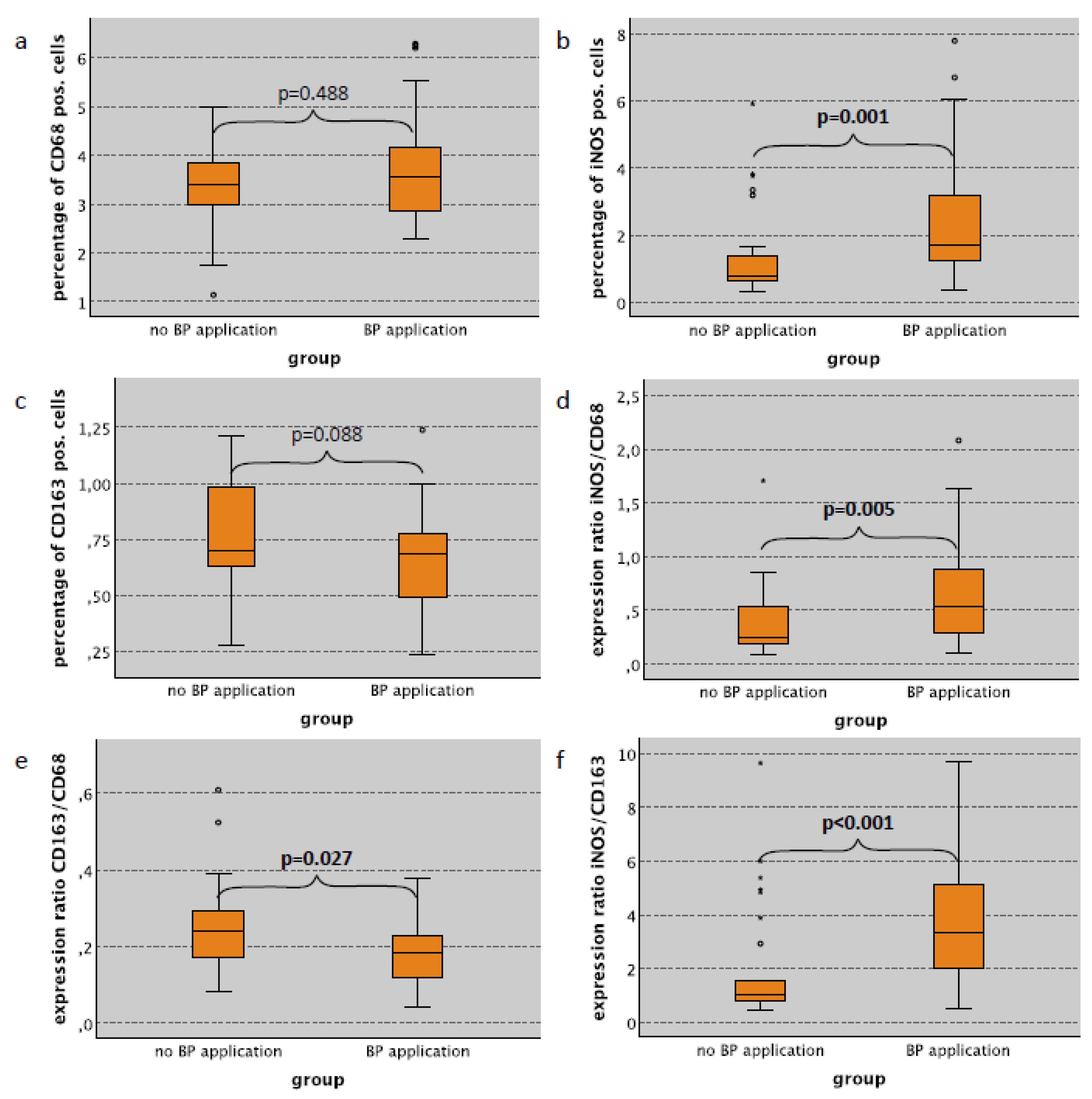
2.3. Macrophage Infiltration and Polarization in the Lung in Relation to Bisphosphonate Application
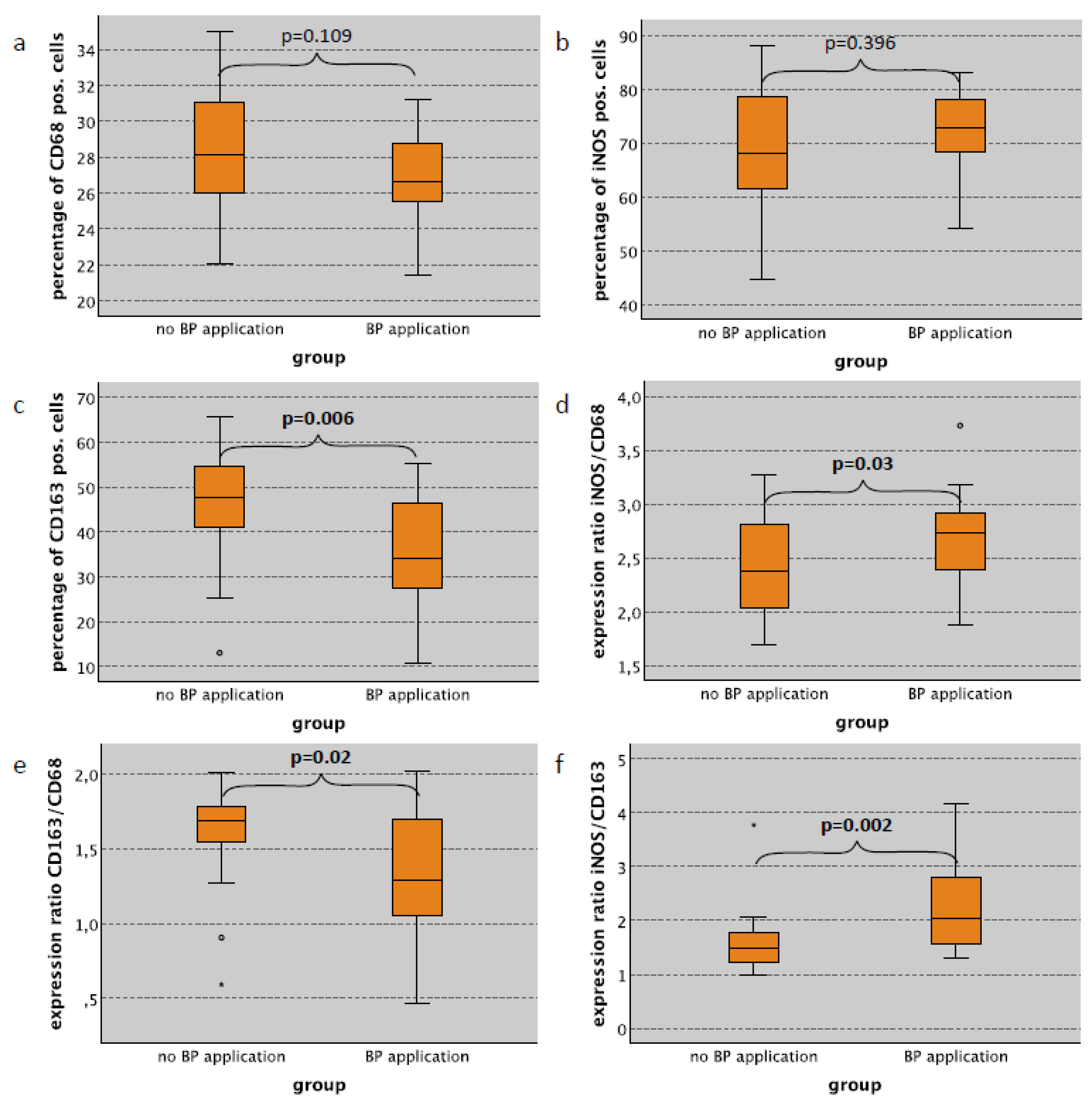
2.4. Macrophage Infiltration and Polarization in the Spleen in Relation to Bisphosphonate Application
2.5. Macrophage Infiltration and Polarization in the Skin Subepithelium in Relation to Surgical Trauma
2.6. Macrophage Infiltration and Polarization in the Lung in Relation to Surgical Trauma
2.7. Macrophage Infiltration and Polarization in the Spleen Red Pulp in Relation to Surgical Trauma
3. Discussion
3.1. Bisphosphonate-Derived Alteration of Systemic Macrophage Polarization and the Possible Influence on MRONJ
3.2. Bisphosphonate-Derived Alteration of Systemic Macrophage Polarization and the Possible Influence on Cancer
3.3. Influence of Surgical Trauma on Systemic Macrophage Polarization
3.4. Macrophages Polarization and Its Potential Influence on Dental Implant and Bone Grafting Success
3.5. Limitations of the Study
4. Materials and Methods
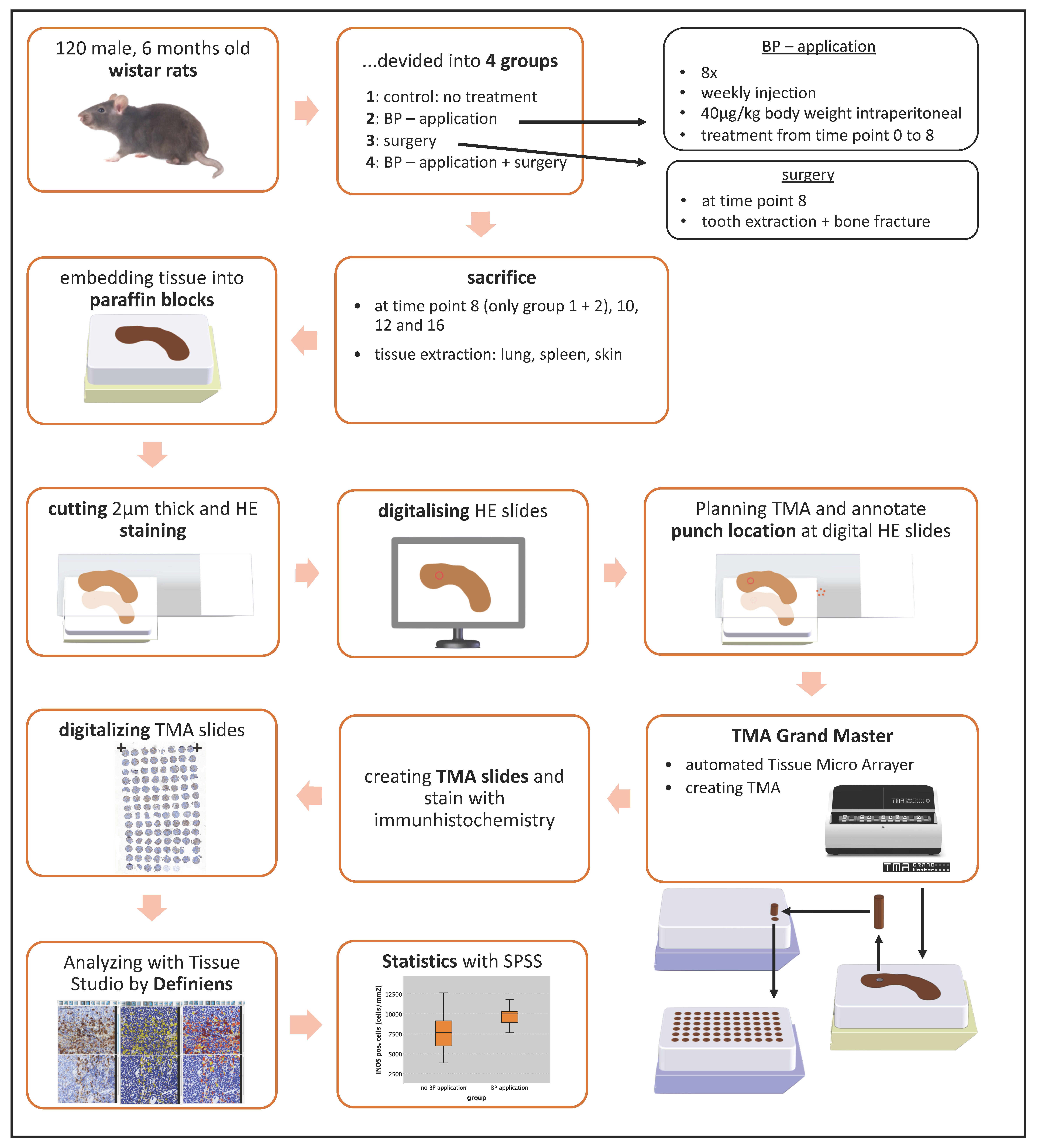
4.1. Animal Experiment
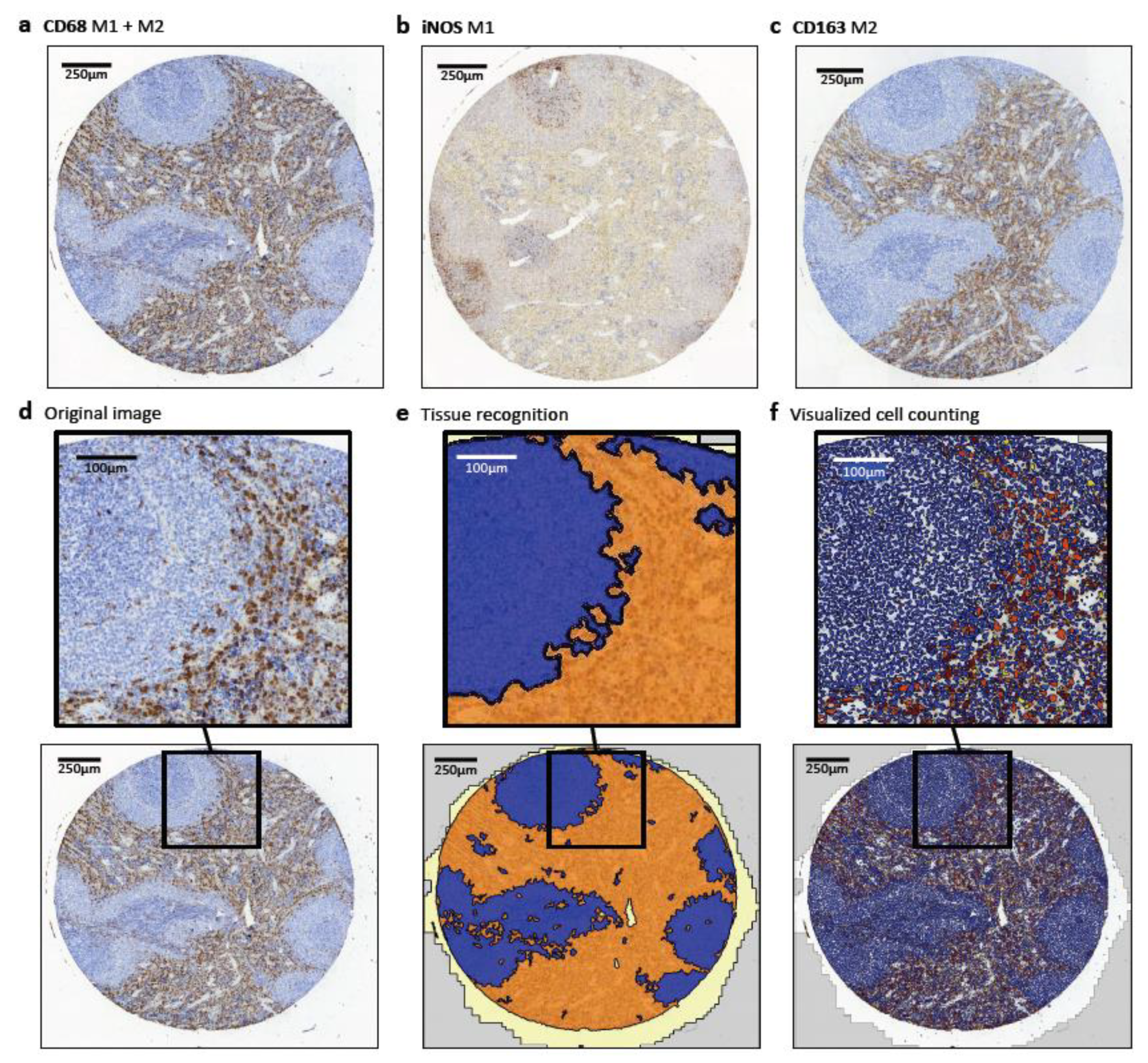
4.2. Design of Tissue Microarrays (TMA)
4.3. Immunohistochemical Staining and Quantitative Analysis
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stockmann, P.; Vairaktaris, E.; Wehrhan, F.; Seiss, M.; Schwarz, S.; Spriewald, B.; Neukam, F.W.; Nkenke, E. Osteotomy and primary wound closure in bisphosphonate-associated osteonecrosis of the jaw: A prospective clinical study with 12 months follow-up. Support Care Cancer 2010, 18, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Wehrhan, F.; Gross, C.; Creutzburg, K.; Amann, K.; Ries, J.; Kesting, M.; Geppert, C.I.; Weber, M. Osteoclastic expression of higher-level regulators NFATc1 and BCL6 in medication-related osteonecrosis of the jaw secondary to bisphosphonate therapy: A comparison with osteoradionecrosis and osteomyelitis. J. Transl. Med. 2019, 17, 69. [Google Scholar] [CrossRef] [PubMed]
- Gross, C.; Weber, M.; Creutzburg, K.; Möbius, P.; Preidl, R.; Amann, K.; Wehrhan, F. Osteoclast profile of medication-related osteonecrosis of the jaw secondary to bisphosphonate therapy: A comparison with osteoradionecrosis and osteomyelitis. J. Transl. Med. 2017, 15, 128. [Google Scholar] [CrossRef] [PubMed]
- Hadji, P.; Coleman, R.E.; Wilson, C.; Powles, T.J.; Clezardin, P.; Aapro, M.; Costa, L.; Body, J.J.; Markopoulos, C.; Santini, D.; et al. Adjuvant bisphosphonates in early breast cancer: Consensus guidance for clinical practice from a European Panel. Ann. Oncol. 2016, 27, 379–390. [Google Scholar] [CrossRef] [PubMed]
- O’Carrigan, B.; Wong, M.H.; Willson, M.L.; Stockler, M.R.; Pavlakis, N.; Goodwin, A. Bisphosphonates and other bone agents for breast cancer. Cochrane Database Syst. Rev. 2017, 10. [Google Scholar] [CrossRef]
- Stockmann, P.; Burger, M.; von Wilmowsky, C.; Ebker, T.; Lutz, R.; Bauersachs, A.; Nkenke, E.; Neukam, F.W.; Wehrhan, F. The outcome after surgical therapy of bisphosphonate-associated osteonecrosis of the jaw–results of a clinical case series with an average follow-up of 20 months. Clin. Oral Investig. 2014, 18, 1299–1304. [Google Scholar] [CrossRef] [PubMed]
- Wehrhan, F.; Amann, K.; Mobius, P.; Weber, M.; Preidl, R.; Ries, J.; Stockmann, P. BRONJ-related jaw bone is associated with increased Dlx-5 and suppressed osteopontin-implication in the site-specific alteration of angiogenesis and bone turnover by bisphosphonates. Clin. Oral Investig. 2015, 19, 1289–1298. [Google Scholar] [CrossRef]
- Wehrhan, F.; Hyckel, P.; Amann, K.; Ries, J.; Stockmann, P.; Schlegel, K.; Neukam, F.; Nkenke, E. Msx-1 is suppressed in bisphosphonate-exposed jaw bone analysis of bone turnover-related cell signalling after bisphosphonate treatment. Oral Dis. 2011, 17, 433–442. [Google Scholar] [CrossRef] [Green Version]
- Wehrhan, F.; Moebius, P.; Amann, K.; Ries, J.; Preidl, R.; Neukam, F.W.; Weber, M. Macrophage and osteoclast polarization in bisphosphonate associated necrosis and osteoradionecrosis. J. Cranio-Maxillofac. Surg. 2017, 45, 944–953. [Google Scholar] [CrossRef]
- Mantovani, A.; Biswas, S.K.; Galdiero, M.R.; Sica, A.; Locati, M. Macrophage plasticity and polarization in tissue repair and remodelling. J. Pathol. 2013, 229, 176–185. [Google Scholar] [CrossRef]
- Sica, A.; Mantovani, A. Macrophage plasticity and polarization: In vivo veritas. J. Clin. Investig. 2012, 122, 787–795. [Google Scholar] [CrossRef]
- Mantovani, A.; Sica, A.; Locati, M. New vistas on macrophage differentiation and activation. Eur. J. Immunol. 2007, 37, 14–16. [Google Scholar] [CrossRef]
- Hao, N.B.; Lu, M.H.; Fan, Y.H.; Cao, Y.L.; Zhang, Z.R.; Yang, S.M. Macrophages in tumor microenvironments and the progression of tumors. Clin. Dev. Immunol. 2012, 2012, 948098. [Google Scholar] [CrossRef] [Green Version]
- Werneck-Gomes, H.; Campolina-Silva, G.H.; Maria, B.T.; Barata, M.C.; Mahecha, G.A.B.; Hess, R.A.; Oliveira, C.A. Tumor-Associated Macrophages (TAM) are recruited to the aging prostate epithelial lesions and become intermingled with basal cells. Andrology 2020. [Google Scholar] [CrossRef]
- Bloomer, S.A.; Moyer, E.D.; Brown, K.E.; Kregel, K.C. Aging results in accumulation of M1 and M2 hepatic macrophages and a differential response to gadolinium chloride. Histochem. Cell Biol. 2020, 153, 37–48. [Google Scholar] [CrossRef]
- Ashouri, F.; Beyranvand, F.; Boroujeni, N.B.; Tavafi, M.; Sheikhian, A.; Varzi, A.M.; Shahrokhi, S. Macrophage polarization in wound healing: Role of aloe vera/chitosan nanohydrogel. Drug Deliv. Transl. Res. 2019, 9, 1027–1042. [Google Scholar] [CrossRef]
- Kaneko, J.; Okinaga, T.; Hikiji, H.; Ariyoshi, W.; Yoshiga, D.; Habu, M.; Tominaga, K.; Nishihara, T. Zoledronic acid exacerbates inflammation through M1 macrophage polarization. Inflamm. Regen. 2018, 38, 16. [Google Scholar] [CrossRef]
- Zhu, W.; Xu, R.; Du, J.; Fu, Y.; Li, S.; Zhang, P.; Liu, L.; Jiang, H. Zoledronic acid promotes TLR-4-mediated M1 macrophage polarization in bisphosphonate-related osteonecrosis of the jaw. FASEB J. 2019, 33, 5208–5219. [Google Scholar] [CrossRef]
- Zhou, J.; Tang, Z.; Gao, S.; Li, C.; Feng, Y.; Zhou, X. Tumor-Associated Macrophages: Recent Insights and Therapies. Front. Oncol. 2020, 10, 188. [Google Scholar] [CrossRef]
- Ramos, R.N.; Rodriguez, C.; Hubert, M.; Ardin, M.; Treilleux, I.; Ries, C.H.; Lavergne, E.; Chabaud, S.; Colombe, A.; Tredan, O.; et al. CD163(+) tumor-associated macrophage accumulation in breast cancer patients reflects both local differentiation signals and systemic skewing of monocytes. Clin. Transl. Immunol. 2020, 9, e1108. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Che, X.; Qiu, X.; Li, Z.; Yang, B.; Wang, S.; Hou, K.; Fan, Y.; Qu, X.; Liu, Y. M2 macrophage infiltration into tumor islets leads to poor prognosis in non-small-cell lung cancer. Cancer Manag. Res. 2019, 11, 6125–6138. [Google Scholar] [CrossRef] [Green Version]
- Weber, M.; Iliopoulos, C.; Moebius, P.; Buttner-Herold, M.; Amann, K.; Ries, J.; Preidl, R.; Neukam, F.W.; Wehrhan, F. Prognostic significance of macrophage polarization in early stage oral squamous cell carcinomas. Oral Oncol. 2016, 52, 75–84. [Google Scholar] [CrossRef]
- Weber, M.; Büttner-Herold, M.; Hyckel, P.; Moebius, P.; Distel, L.; Ries, J.; Amann, K.; Neukam, F.W.; Wehrhan, F. Small oral squamous cell carcinomas with nodal lymphogenic metastasis show increased infiltration of M2 polarized macrophages—An immunohistochemical analysis. J. Cranio-Maxillofac. Surg. 2014, 42, 1087–1094. [Google Scholar] [CrossRef]
- Weber, M.; Wehrhan, F.; Baran, C.; Agaimy, A.; Buttner-Herold, M.; Ozturk, H.; Neubauer, K.; Wickenhauser, C.; Kesting, M.; Ries, J. Malignant transformation of oral leukoplakia is associated with macrophage polarization. J. Transl. Med. 2020, 18, 11. [Google Scholar] [CrossRef] [Green Version]
- Hesketh, M.; Sahin, K.B.; West, Z.E.; Murray, R.Z. Macrophage Phenotypes Regulate Scar Formation and Chronic Wound Healing. Int. J. Mol. Sci. 2017, 18, 1545. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Nair, M.G. Macrophages in wound healing: Activation and plasticity. Immunol. Cell Biol. 2019, 97, 258–267. [Google Scholar] [CrossRef]
- Weber, M.; Moebius, P.; Buttner-Herold, M.; Amann, K.; Preidl, R.; Neukam, F.W.; Wehrhan, F. Macrophage polarisation changes within the time between diagnostic biopsy and tumour resection in oral squamous cell carcinomas—An immunohistochemical study. Br. J. Cancer 2015, 113, 510–519. [Google Scholar] [CrossRef] [Green Version]
- Das, A.; Sinha, M.; Datta, S.; Abas, M.; Chaffee, S.; Sen, C.K.; Roy, S. Monocyte and macrophage plasticity in tissue repair and regeneration. Am. J. Pathol. 2015, 185, 2596–2606. [Google Scholar] [CrossRef] [Green Version]
- Sindrilaru, A.; Peters, T.; Wieschalka, S.; Baican, C.; Baican, A.; Peter, H.; Hainzl, A.; Schatz, S.; Qi, Y.; Schlecht, A.; et al. An unrestrained proinflammatory M1 macrophage population induced by iron impairs wound healing in humans and mice. J. Clin. Investig. 2011, 121, 985–997. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Atsuta, I.; Liu, S.; Chen, C.; Shi, S.; Shi, S.; Le, A.D. IL-17-mediated M1/M2 macrophage alteration contributes to pathogenesis of bisphosphonate-related osteonecrosis of the jaws. Clin. Cancer Res. 2013, 19, 3176–3188. [Google Scholar] [CrossRef] [Green Version]
- Aft, R.; Naughton, M.; Trinkaus, K.; Watson, M.; Ylagan, L.; Chavez-MacGregor, M.; Zhai, J.; Kuo, S.; Shannon, W.; Diemer, K.; et al. Effect of zoledronic acid on disseminated tumour cells in women with locally advanced breast cancer: An open label, randomised, phase 2 trial. Lancet Oncol. 2010, 11, 421–428. [Google Scholar] [CrossRef] [Green Version]
- Heymann, P.G.; Ziebart, T.; Kammerer, P.W.; Mandic, R.; Saydali, A.; Braun, A.; Neff, A.; Draenert, G.F. The enhancing effect of a laser photochemotherapy with cisplatin or zolendronic acid in primary human osteoblasts and osteosarcoma cells in vitro. J. Oral Pathol. Med. 2016, 45, 803–809. [Google Scholar] [CrossRef]
- Obeid, E.; Nanda, R.; Fu, Y.X.; Olopade, O.I. The role of tumor-associated macrophages in breast cancer progression (review). Int. J. Oncol. 2013, 43, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Sousa, S.; Brion, R.; Lintunen, M.; Kronqvist, P.; Sandholm, J.; Monkkonen, J.; Kellokumpu-Lehtinen, P.L.; Lauttia, S.; Tynninen, O.; Joensuu, H.; et al. Human breast cancer cells educate macrophages toward the M2 activation status. Breast Cancer Res. 2015, 17, 101. [Google Scholar] [CrossRef] [Green Version]
- Tariq, M.; Zhang, J.; Liang, G.; Ding, L.; He, Q.; Yang, B. Macrophage Polarization: Anti-Cancer Strategies to Target Tumor-Associated Macrophage in Breast Cancer. J. Cell Biochem. 2017, 118, 2484–2501. [Google Scholar] [CrossRef]
- Kurotaki, D.; Uede, T.; Tamura, T. Functions and development of red pulp macrophages. Microbiol. Immunol. 2015, 59, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Najafi, M.; Farhood, B.; Mortezaee, K. Contribution of regulatory T cells to cancer: A review. J. Cell Physiol. 2019, 234, 7983–7993. [Google Scholar] [CrossRef]
- Miller, P.D.; Pannacciulli, N.; Malouf-Sierra, J.; Singer, A.; Czerwinski, E.; Bone, H.G.; Wang, C.; Huang, S.; Chines, A.; Lems, W.; et al. Efficacy and safety of denosumab vs. bisphosphonates in postmenopausal women previously treated with oral bisphosphonates. Osteoporos. Int. 2020, 31, 181–191. [Google Scholar] [CrossRef]
- Raje, N.; Terpos, E.; Willenbacher, W.; Shimizu, K.; Garcia-Sanz, R.; Durie, B.; Legiec, W.; Krejci, M.; Laribi, K.; Zhu, L.; et al. Denosumab versus zoledronic acid in bone disease treatment of newly diagnosed multiple myeloma: An international, double-blind, double-dummy, randomised, controlled, phase 3 study. Lancet Oncol. 2018, 19, 370–381. [Google Scholar] [CrossRef]
- Diel, I.; Ansorge, S.; Hohmann, D.; Giannopoulou, C.; Niepel, D.; Intorcia, M. Real-world use of denosumab and bisphosphonates in patients with solid tumours and bone metastases in Germany. Support Care Cancer 2020, 28, 5223–5233. [Google Scholar] [CrossRef] [Green Version]
- Patntirapong, S.; Poolgesorn, M. Alteration of macrophage viability, differentiation, and function by bisphosphonates. Oral Dis. 2018, 24, 1294–1302. [Google Scholar] [CrossRef]
- Patntirapong, S.; Phupunporn, P.; Vanichtantiphong, D.; Thanetchaloempong, W. Inhibition of macrophage viability by bound and free bisphosphonates. Acta Histochem. 2019, 121, 400–406. [Google Scholar] [CrossRef]
- Moest, T.; Frabschka, J.; Kesting, M.R.; Schmitt, C.M.; Frohwitter, G.; Lutz, R.; Schlegel, K.A. Osseous ingrowth in allogeneic bone blocks applied for vertical bone augmentation: A preclinical randomised controlled study. Clin. Oral Investig. 2020, 24, 2867–2879. [Google Scholar] [CrossRef]
- Moest, T.; Wehrhan, F.; Lutz, R.; Schmitt, C.M.; Neukam, F.W.; Schlegel, K.A. Extra-oral defect augmentation using autologous, bovine and equine bone blocks: A preclinical histomorphometrical comparative study. J. Craniomaxillofac. Surg. 2015, 43, 559–566. [Google Scholar] [CrossRef]
- von Wilmowsky, C.; Stockmann, P.; Harsch, I.; Amann, K.; Metzler, P.; Lutz, R.; Moest, T.; Neukam, F.W.; Schlegel, K.A. Diabetes mellitus negatively affects peri-implant bone formation in the diabetic domestic pig. J. Clin. Periodontol. 2011, 38, 771–779. [Google Scholar] [CrossRef]
- Schlegel, K.A.; Prechtl, C.; Möst, T.; Seidl, C.; Lutz, R.; von Wilmowsky, C. Osseointegration of SLActive implants in diabetic pigs. Clin. Oral Implants Res. 2013, 24, 128–134. [Google Scholar] [CrossRef]
- Silveira, F.M.; Etges, A.; Correa, M.B.; Vasconcelos, A.C. Microscopic Evaluation of the Effect of Oral Microbiota on the Development of Bisphosphonate-Related Osteonecrosis of the Jaws in Rats. J. Oral Maxillofac. Res. 2016, 7, e3. [Google Scholar] [CrossRef] [Green Version]
- Koneski, F.; Popovic-Monevska, D.; Gjorgoski, I.; Krajoska, J.; Popovska, M.; Muratovska, I.; Velickovski, B.; Petrushevska, G.; Popovski, V. In vivo effects of geranylgeraniol on the development of bisphosphonate-related osteonecrosis of the jaws. J. Cranio-Maxillofac. Surg. 2018, 46, 230–236. [Google Scholar] [CrossRef]
- Zandi, M.; Dehghan, A.; Malekzadeh, H.; Janbaz, P.; Ghadermazi, K.; Amini, P. Introducing a protocol to create bisphosphonate-related osteonecrosis of the jaw in rat animal model. J. Cranio-Maxillofac. Surg. 2016, 44, 271–278. [Google Scholar] [CrossRef]
- Jayaraman, P.; Alfarano, M.G.; Svider, P.F.; Parikh, F.; Lu, G.; Kidwai, S.; Xiong, H.; Sikora, A.G. iNOS expression in CD4+ T cells limits Treg induction by repressing TGFbeta1: Combined iNOS inhibition and Treg depletion unmask endogenous antitumor immunity. Clin. Cancer Res. 2014, 20, 6439–6451. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ((pos. Cells/all Cells in %)/(pos. Cells/all Cells in %)) in Skin Subepithelium Tissue of Wistar Rats | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Marker | n | CD68 | iNOS | CD163 | Ratio iNOS/CD68 | Ratio CD163/CD68 | Ratio iNOS/CD163 | ||||||
| Median | SD | Median | SD | Median | SD | Median | SD | Median | SD | Median | SD | ||
| BP application—all cases | |||||||||||||
| no | 43 | 3.47 | 2.46 | 1.18 | 1.99 | 13.55 | 4.93 | 0.38 | 3.71 | 3.38 | 5.38 | 0.11 | 0.11 |
| yes | 33 | 6.66 | 3.17 | 2.11 | 1.75 | 13.20 | 5.10 | 0.36 | 1.31 | 2.24 | 2.58 | 0.18 | 0.14 |
| p-value | 0.004 | 0.017 | 0.892 | 0.906 | 0.007 | 0.012 | |||||||
| BP application—only no surgical trauma cases | |||||||||||||
| no | 21 | 3.96 | 2.18 | 1.12 | 0.72 | 13.37 | 3.94 | 0.29 | 0.70 | 2.79 | 6.99 | 0.09 | 0.07 |
| yes | 23 | 6.72 | 3.05 | 2.99 | 1.74 | 13.20 | 3.75 | 0.44 | 0.67 | 2.22 | 2.11 | 0.22 | 0.13 |
| p-value | 0.014 | <0.001 | 0.916 | 0.089 | 0.065 | <0.001 | |||||||
| BP application—only surgical trauma cases | |||||||||||||
| no | 22 | 3.42 | 2.74 | 1.31 | 2.63 | 13.97 | 5.81 | 0.42 | 5.14 | 3.61 | 3.22 | 0.13 | 0.14 |
| yes | 10 | 5.72 | 3.52 | 1.04 | 0.96 | 14.03 | 7.62 | 0.23 | 2.20 | 2.47 | 3.51 | 0.08 | 0.06 |
| p-value | 0.251 | 0.434 | 0.826 | 0.047 | 0.129 | 0.164 | |||||||
| surgical trauma—all cases | |||||||||||||
| no | 44 | 5.21 | 2.84 | 1.87 | 1.61 | 13.29 | 3.80 | 0.40 | 0.68 | 2.45 | 5.23 | 0.15 | 0.13 |
| yes | 32 | 3.68 | 3.02 | 1.24 | 2.28 | 13.97 | 6.31 | 0.36 | 4.41 | 3.31 | 3.29 | 0.08 | 0.12 |
| p-value | 0.105 | 0.119 | 0.556 | 0.622 | 0.252 | 0.03 | |||||||
| surgical trauma—only no BP application cases | |||||||||||||
| no | 21 | 3.96 | 2.18 | 1.12 | 0.72 | 13.37 | 3.94 | 0.29 | 0.70 | 2.79 | 6.99 | 0.09 | 0.07 |
| yes | 22 | 3.42 | 2.74 | 1.31 | 2.63 | 13.97 | 5.81 | 0.42 | 5.14 | 3.61 | 3.22 | 0.13 | 0.14 |
| p-value | 0.423 | 0.369 | 0.752 | 0.224 | 0.593 | 0.399 | |||||||
| surgical trauma—only BP application cases | |||||||||||||
| no | 23 | 6.72 | 3.05 | 2.99 | 1.74 | 13.20 | 3.75 | 0.44 | 0.67 | 2.22 | 2.11 | 0.22 | 0.13 |
| yes | 10 | 5.72 | 3.52 | 1.04 | 0.96 | 14.03 | 7.62 | 0.23 | 2.20 | 2.47 | 3.51 | 0.08 | 0.06 |
| p-value | 0.428 | 0.003 | 0.63 | 0.014 | 0.857 | <0.001 | |||||||
| ((pos. Cells/all Cells)/(pos. Cells/all Cells)) in Lung Tissue of Wistar Rats | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Marker | n | CD68 | iNOS | CD163 | Ratio iNOS/CD68 | Ratio CD163/CD68 | Ratio iNOS/CD163 | ||||||
| Median | SD | Median | SD | Median | SD | Median | SD | Median | SD | Median | SD | ||
| BP application—all cases | |||||||||||||
| no | 53 | 3.33 | 1.01 | 1.22 | 1.20 | 0.68 | 0.27 | 0.41 | 0.34 | 0.22 | 0.11 | 1.76 | 1.87 |
| yes | 52 | 3.63 | 2.09 | 2.31 | 2.31 | 0.69 | 0.27 | 0.71 | 0.66 | 0.18 | 0.09 | 3.81 | 3.11 |
| p-value | 0.209 | <0.001 | 0.509 | 0.002 | 0.109 | <0.001 | |||||||
| BP application—only no surgical trauma cases | |||||||||||||
| no | 29 | 3.40 | 0.90 | 0.80 | 1.33 | 0.70 | 0.24 | 0.24 | 0.35 | 0.24 | 0.12 | 1.02 | 2.18 |
| yes | 32 | 3.54 | 1.35 | 1.68 | 1.78 | 0.69 | 0.22 | 0.54 | 0.46 | 0.18 | 0.08 | 3.35 | 2.41 |
| p-value | 0.488 | 0.001 | 0.088 | 0.005 | 0.027 | <0.001 | |||||||
| BP application—only surgical trauma cases | |||||||||||||
| no | 24 | 3.20 | 1.14 | 1.59 | 0.99 | 0.58 | 0.30 | 0.49 | 0.33 | 0.17 | 0.11 | 2.81 | 1.29 |
| yes | 20 | 3.67 | 2.89 | 3.15 | 2.80 | 0.69 | 0.32 | 0.79 | 0.86 | 0.18 | 0.10 | 4.20 | 3.92 |
| p-value | 0.258 | 0.002 | 0.239 | 0.038 | 0.832 | 0.004 | |||||||
| surgical trauma—all cases | |||||||||||||
| no | 61 | 3.50 | 1.17 | 1.34 | 1.66 | 0.69 | 0.24 | 0.45 | 0.43 | 0.20 | 0.11 | 2.05 | 2.46 |
| yes | 44 | 3.58 | 2.16 | 2.00 | 2.23 | 0.66 | 0.31 | 0.53 | 0.66 | 0.18 | 0.10 | 3.51 | 3.00 |
| p-value | 0.59 | 0.004 | 0.608 | 0.026 | 0.413 | 0.012 | |||||||
| surgical trauma–only no BP application cases | |||||||||||||
| no | 29 | 3.40 | 0.90 | 0.80 | 1.33 | 0.70 | 0.24 | 0.24 | 0.35 | 0.24 | 0.12 | 1.02 | 2.18 |
| yes | 24 | 3.20 | 1.14 | 1.59 | 0.99 | 0.58 | 0.30 | 0.49 | 0.33 | 0.17 | 0.11 | 2.81 | 1.29 |
| p-value | 0.872 | 0.004 | 0.054 | 0.019 | 0.116 | 0.001 | |||||||
| surgical trauma—only BP application cases | |||||||||||||
| no | 32 | 3.54 | 1.35 | 1.68 | 1.78 | 0.69 | 0.22 | 0.54 | 0.46 | 0.18 | 0.08 | 3.35 | 2.41 |
| yes | 20 | 3.67 | 2.89 | 3.15 | 2.80 | 0.69 | 0.32 | 0.79 | 0.86 | 0.18 | 0.10 | 4.20 | 3.92 |
| p-value | 0.367 | 0.024 | 0.367 | 0.17 | 0.707 | 0.137 | |||||||
| ((pos. Cells/all Cells in %)/(pos. Cells/all Cells in %)) in Spleen Red Pulp of Wistar Rats | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Marker | n | CD68 | iNOS | CD163 | Ratio iNOS/CD68 | Ratio CD163/CD68 | Ratio iNOS/CD163 | ||||||
| Median | SD | Median | SD | Median | SD | Median | SD | Median | SD | Median | SD | ||
| BP application—all cases | |||||||||||||
| no | 48 | 28.52 | 2.90 | 72.59 | 11.70 | 50.46 | 9.83 | 2.49 | 0.41 | 1.75 | 0.28 | 1.43 | 0.43 |
| yes | 44 | 27.51 | 2.38 | 75.64 | 7.16 | 39.93 | 11.04 | 2.72 | 0.33 | 1.37 | 0.39 | 1.88 | 0.91 |
| p-value | 0.116 | 0.099 | <0.001 | 0.008 | <0.001 | <0.001 | |||||||
| BP application—only no surgical trauma cases | |||||||||||||
| no | 26 | 28.15 | 3.10 | 68.18 | 12.15 | 47.58 | 11.91 | 2.37 | 0.44 | 1.68 | 0.31 | 1.48 | 0.55 |
| yes | 25 | 26.61 | 2.27 | 72.76 | 7.73 | 34.17 | 12.47 | 2.73 | 0.40 | 1.29 | 0.43 | 2.03 | 1.15 |
| p-value | 0.109 | 0.396 | 0.006 | 0.03 | 0.02 | 0.002 | |||||||
| BP application–only surgical trauma cases | |||||||||||||
| no | 22 | 28.85 | 2.67 | 73.55 | 10.53 | 51.77 | 5.03 | 2.55 | 0.37 | 1.79 | 0.19 | 1.37 | 0.23 |
| yes | 19 | 28.68 | 2.09 | 76.43 | 4.59 | 42.69 | 7.59 | 2.70 | 0.23 | 1.48 | 0.31 | 1.82 | 0.33 |
| p-value | 0.676 | 0.075 | <0.001 | 0.166 | <0.001 | <0.001 | |||||||
| surgical trauma–all cases | |||||||||||||
| no | 51 | 27.19 | 2.79 | 72.53 | 10.28 | 43.36 | 13.18 | 2.64 | 0.44 | 1.63 | 0.40 | 1.63 | 0.96 |
| yes | 41 | 28.72 | 2.39 | 75.96 | 8.43 | 49.37 | 8.07 | 2.61 | 0.32 | 1.72 | 0.30 | 1.61 | 0.36 |
| p-value | 0.013 | 0.011 | 0.011 | 0.364 | 0.033 | 0.302 | |||||||
| surgical trauma—only no BP application cases | |||||||||||||
| no | 26 | 28.15 | 3.10 | 68.18 | 12.15 | 47.58 | 11.91 | 2.37 | 0.44 | 1.68 | 0.31 | 1.48 | 0.55 |
| yes | 22 | 28.85 | 2.67 | 73.55 | 10.53 | 51.77 | 5.03 | 2.55 | 0.37 | 1.79 | 0.19 | 1.37 | 0.23 |
| p-value | 0.352 | 0.172 | 0.031 | 0.207 | 0.012 | 0.363 | |||||||
| surgical trauma—only BP application cases | |||||||||||||
| no | 25 | 26.61 | 2.27 | 72.76 | 7.73 | 34.17 | 12.47 | 2.73 | 0.40 | 1.29 | 0.43 | 2.03 | 1.15 |
| yes | 19 | 28.68 | 2.09 | 76.43 | 4.59 | 42.69 | 7.59 | 2.70 | 0.23 | 1.48 | 0.31 | 1.82 | 0.33 |
| p-value | 0.008 | 0.008 | 0.07 | 0.878 | 0.231 | 0.562 | |||||||
| Group | n | Total: 108 Animals, 6-Month-Old Male Wistar Rats | ||||
|---|---|---|---|---|---|---|
| 1 | 30 | no BP application | no surgery (control group) | sacrifice | sacrifice | sacrifice |
| (control group) | sacrifice + tissue extraction | + tissue extraction | + tissue extraction | + tissue extraction | ||
| 2 | 32 | 40 μg/kg body weight Zoledronate | no surgery | sacrifice | sacrifice | sacrifice |
| i.p. per week (8× at time point 0–7) | sacrifice + tissue extraction | + tissue extraction | + tissue extraction | + tissue extraction | ||
| 3 | 26 | no bisphosphonate application | bone fracture + tooth extraction | sacrifice | sacrifice | sacrifice |
| + tissue extraction | + tissue extraction | + tissue extraction | ||||
| 4 | 20 | 40 μg/kg body weight Zoledronate | bone fracture + tooth extraction | sacrifice | sacrifice | sacrifice |
| i.p. per week (8× at time point 0–7) | + tissue extraction | + tissue extraction | + tissue extraction | |||
| time point | 0 | 8 | 10 | 12 | 16 | |
| start of experiment | weeks after start of experiment | |||||
| Soft Tissue | Punch Target | Number of | Punch Diameter | Number of Punches | Number of |
|---|---|---|---|---|---|
| Tissue Samples | in mm | Per Target | TMA Blocks | ||
| lung | lung tissue | 103 | 1.5 | 3 | 3 |
| spleen | red pulp | 104 | 1.5 | 3 | 3 |
| white pulp | |||||
| skin | skin epithelium | 102 | 1 | 3 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weber, M.; Homm, A.; Müller, S.; Frey, S.; Amann, K.; Ries, J.; Geppert, C.; Preidl, R.; Möst, T.; Kämmerer, P.W.; et al. Zoledronate Causes a Systemic Shift of Macrophage Polarization towards M1 In Vivo. Int. J. Mol. Sci. 2021, 22, 1323. https://doi.org/10.3390/ijms22031323
Weber M, Homm A, Müller S, Frey S, Amann K, Ries J, Geppert C, Preidl R, Möst T, Kämmerer PW, et al. Zoledronate Causes a Systemic Shift of Macrophage Polarization towards M1 In Vivo. International Journal of Molecular Sciences. 2021; 22(3):1323. https://doi.org/10.3390/ijms22031323
Chicago/Turabian StyleWeber, Manuel, Andi Homm, Stefan Müller, Silke Frey, Kerstin Amann, Jutta Ries, Carol Geppert, Raimund Preidl, Tobias Möst, Peer W. Kämmerer, and et al. 2021. "Zoledronate Causes a Systemic Shift of Macrophage Polarization towards M1 In Vivo" International Journal of Molecular Sciences 22, no. 3: 1323. https://doi.org/10.3390/ijms22031323