
Role of Palatine Tonsil and Epipharyngeal Lymphoid Tissue in the Development of Glomerular Active Lesions (Glomerular vasculitis) in Immunoglobulin A Nephropathy

Abstract
:1. Importance of Hematuria in Immunoglobulin A Nephropathy (IgAN)
2. Role of Palatine Tonsil in IgAN
2.1. Characteristics of the Palatine Tonsil
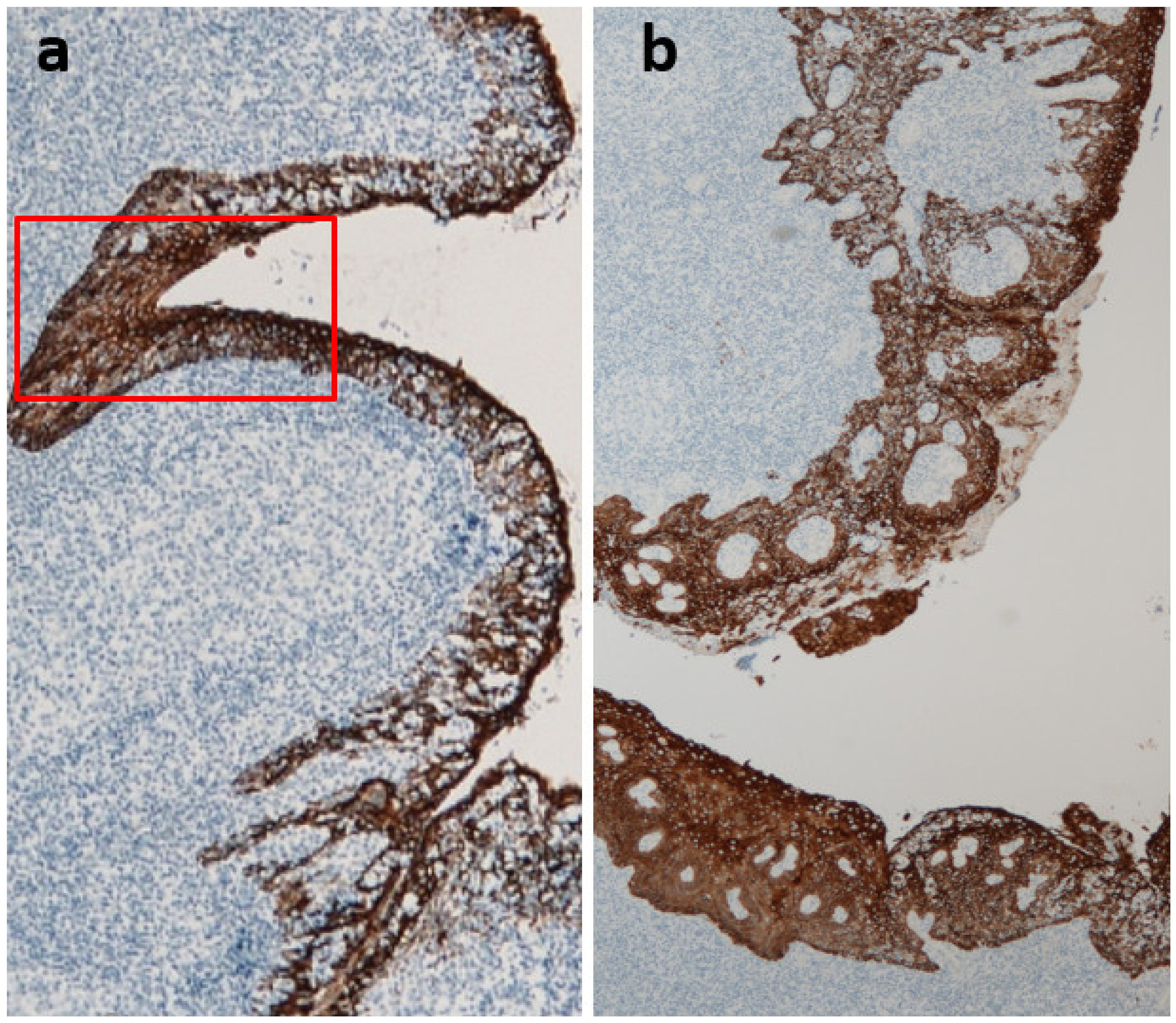
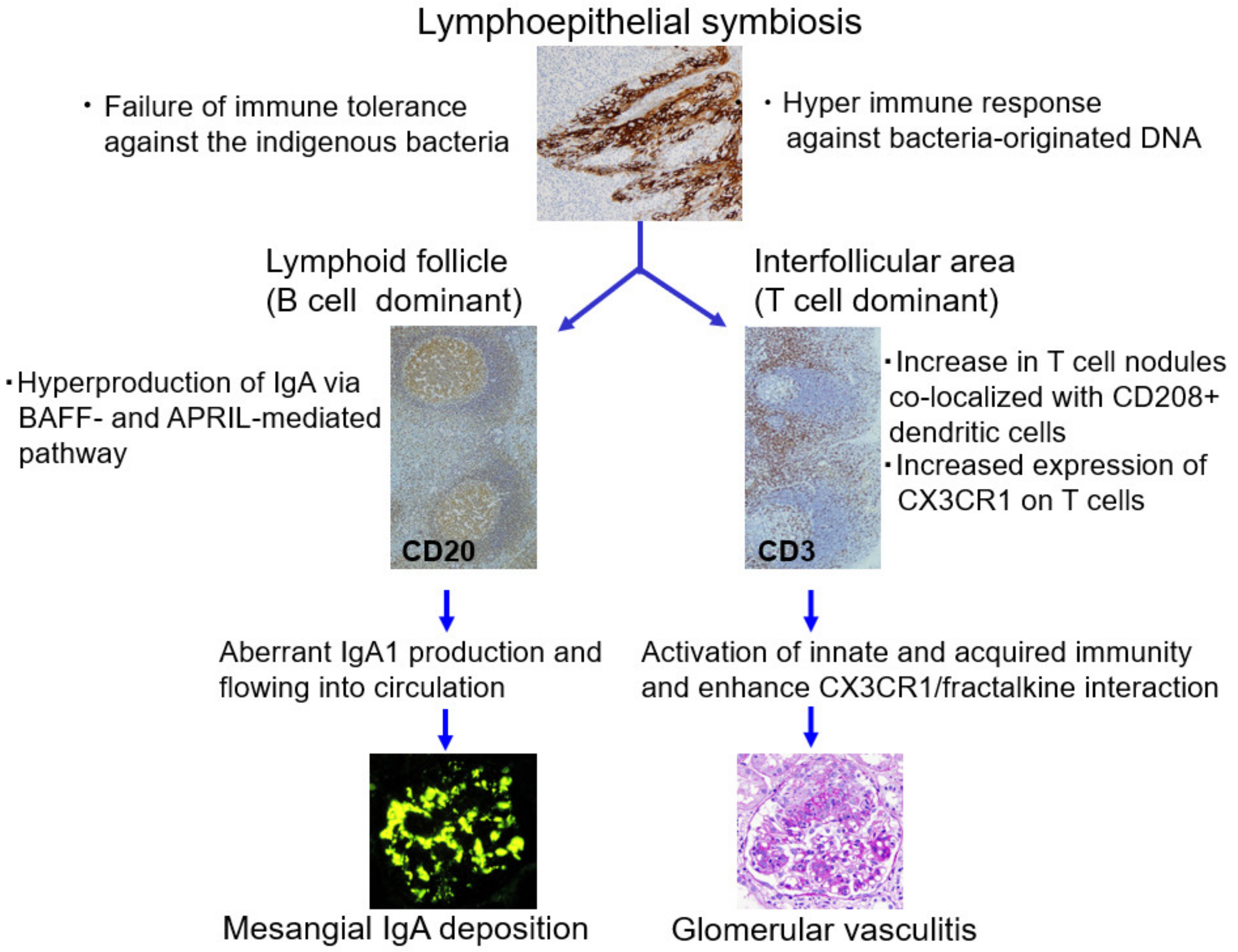
2.2. Role of Tonsils in Mesangial IgA Deposition
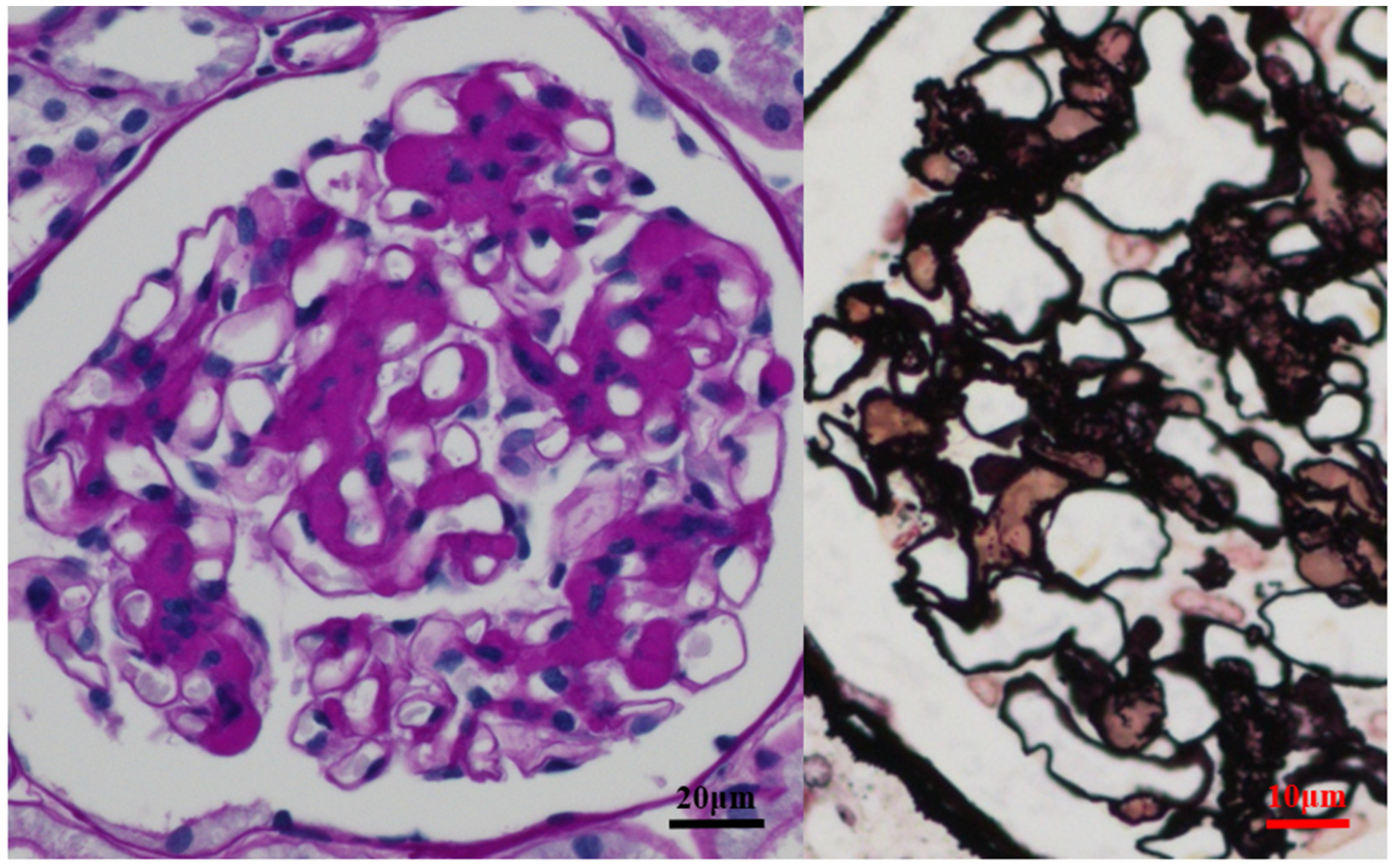
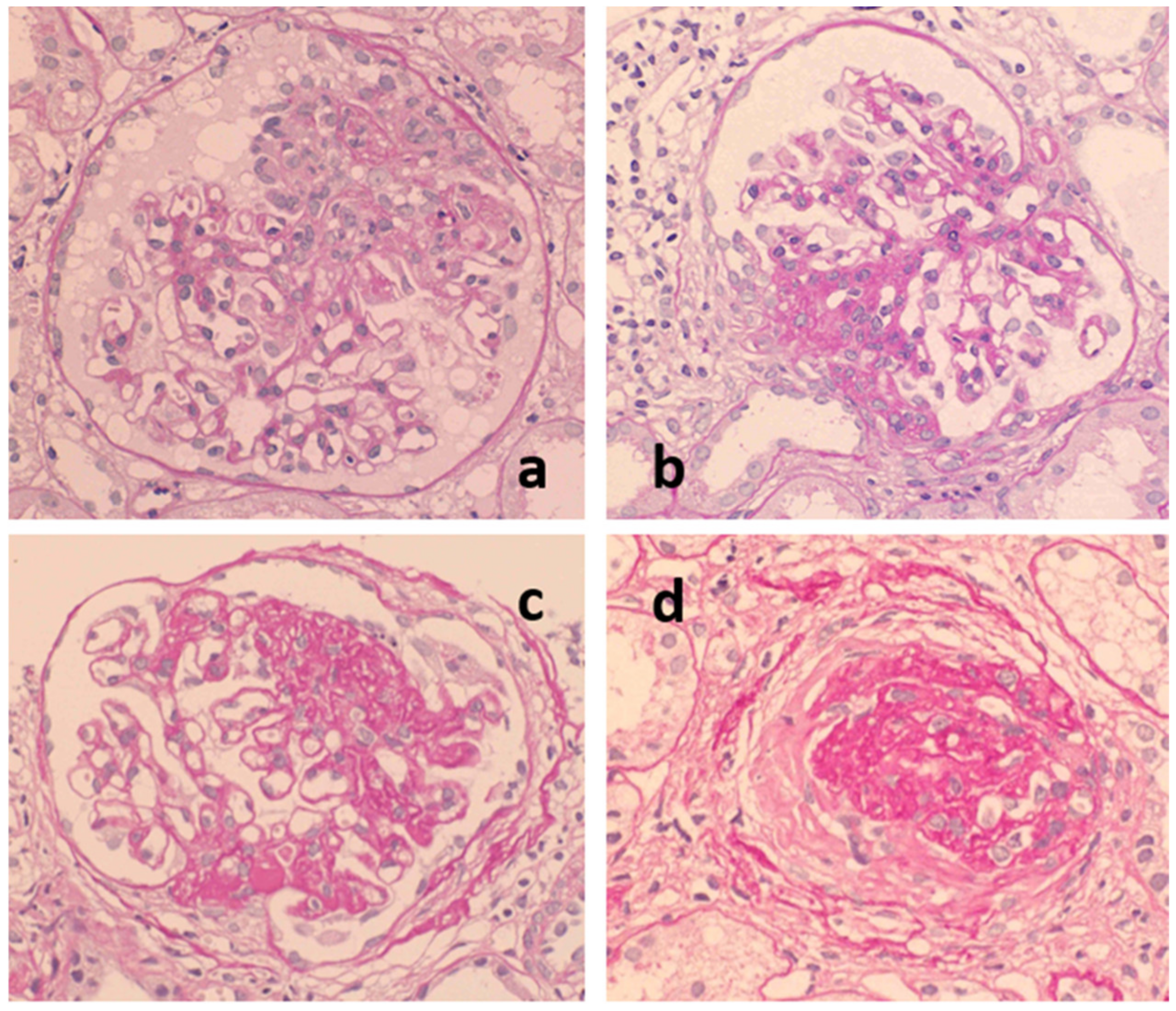
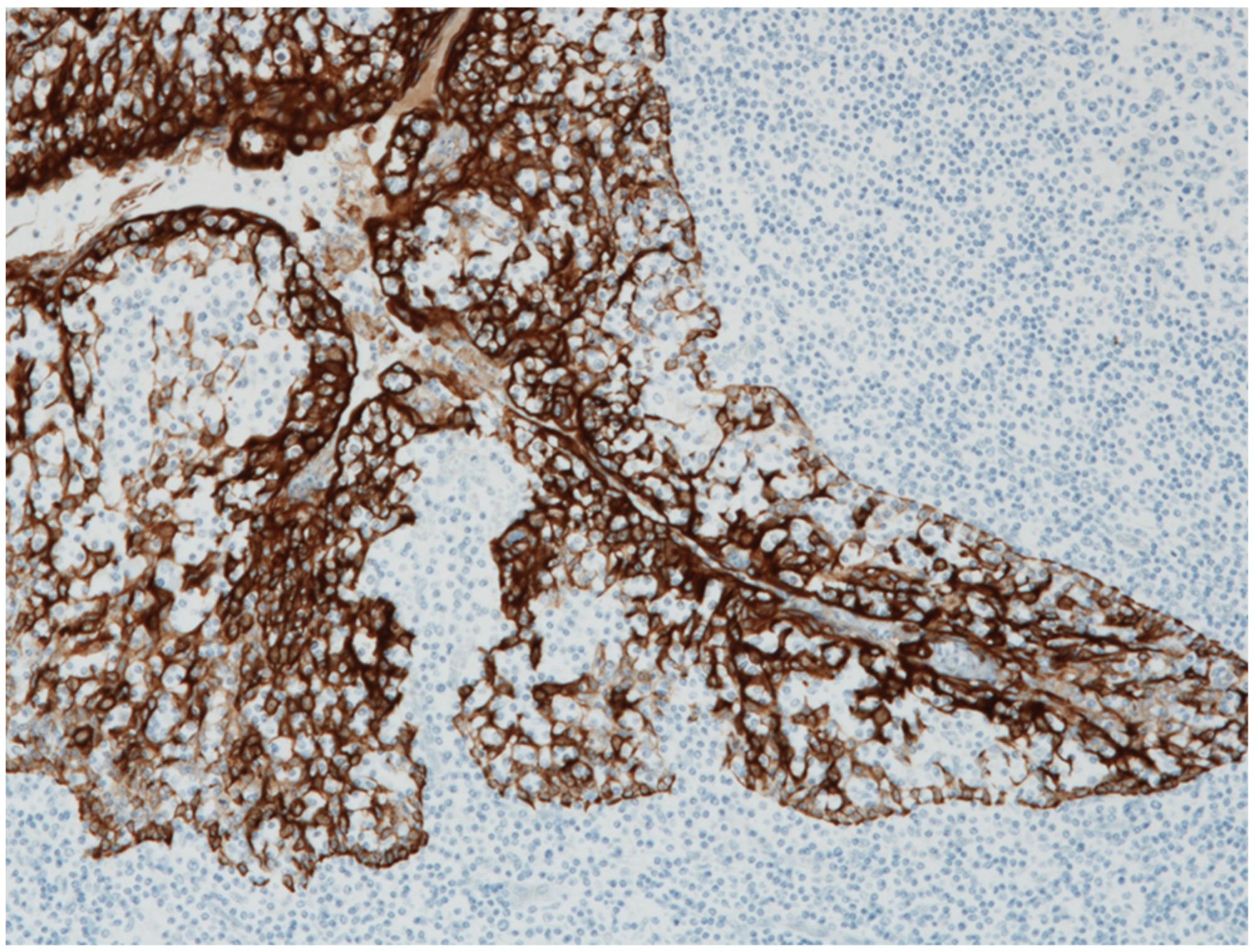
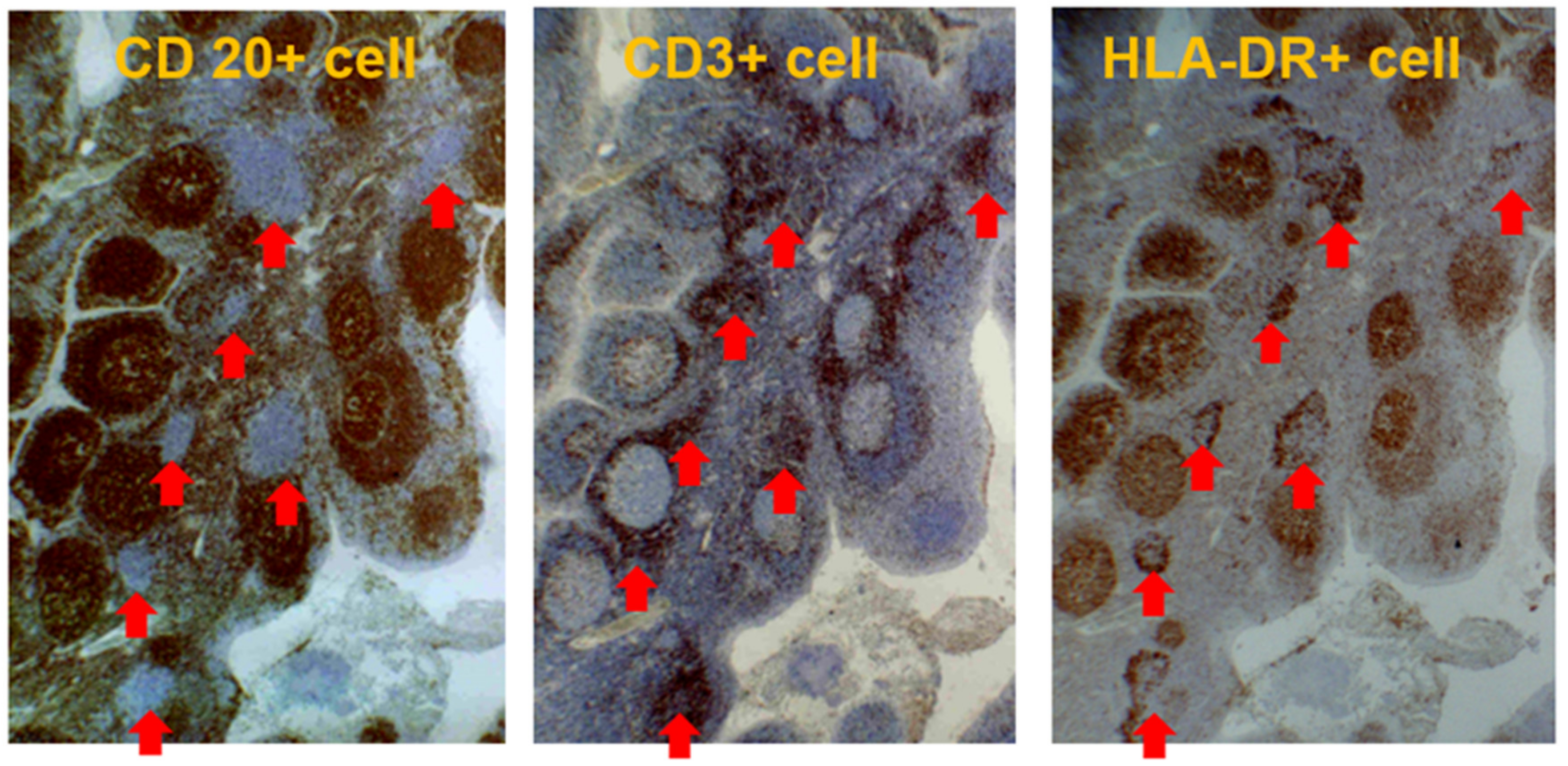
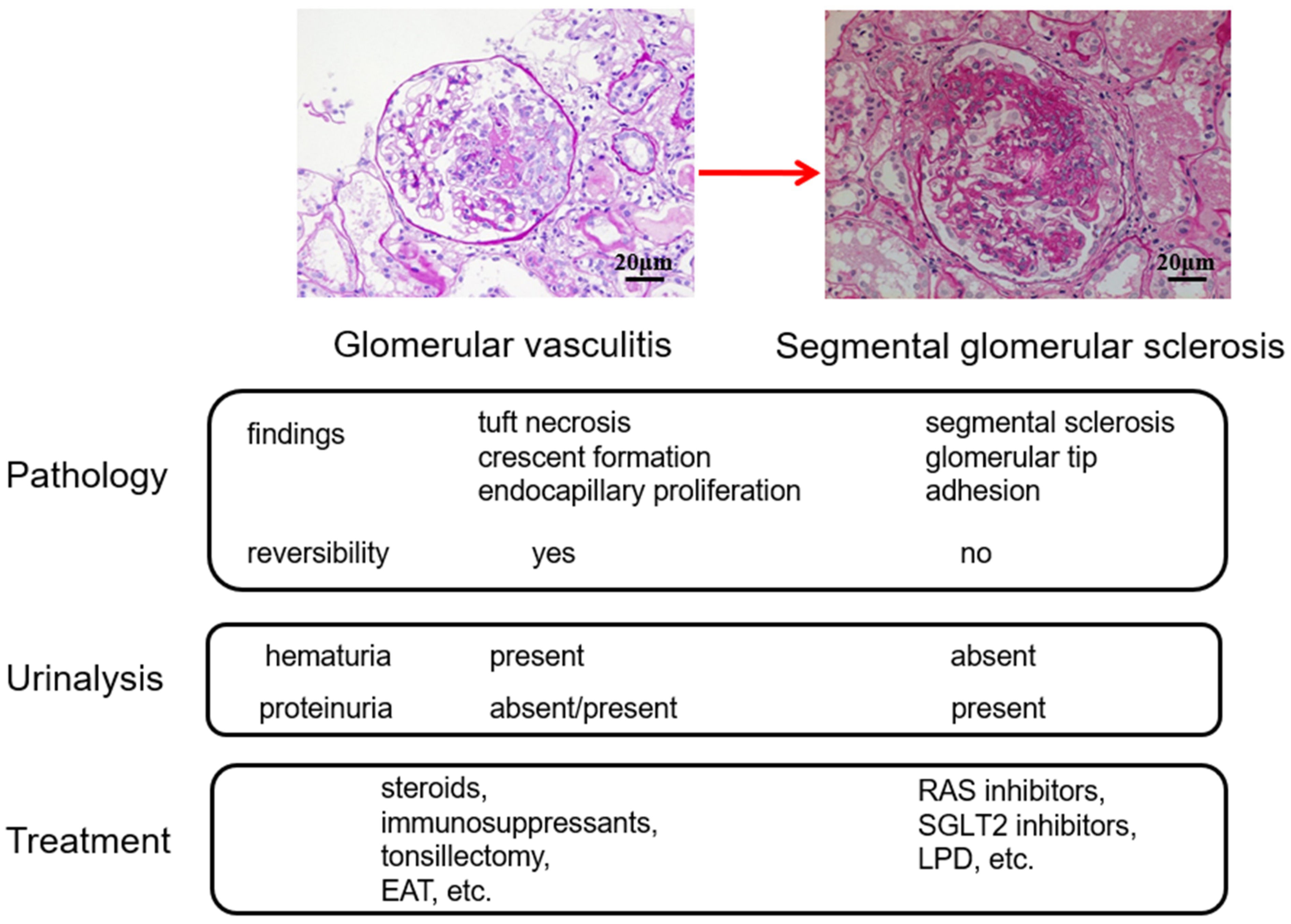
2.3. Role of Tonsils in Glomerular Vasculitis
3. Role of the Epipharyngeal Lymphoid Tissue in IgAN
3.1. Anatomical, Physiological, and Immunological Characteristics of the Epipharynx
3.2. Epipharyngeal Response to Airborne Infections
3.3. Role of the Epipharynx–Kidney Axis in Glomerular Vasculitis
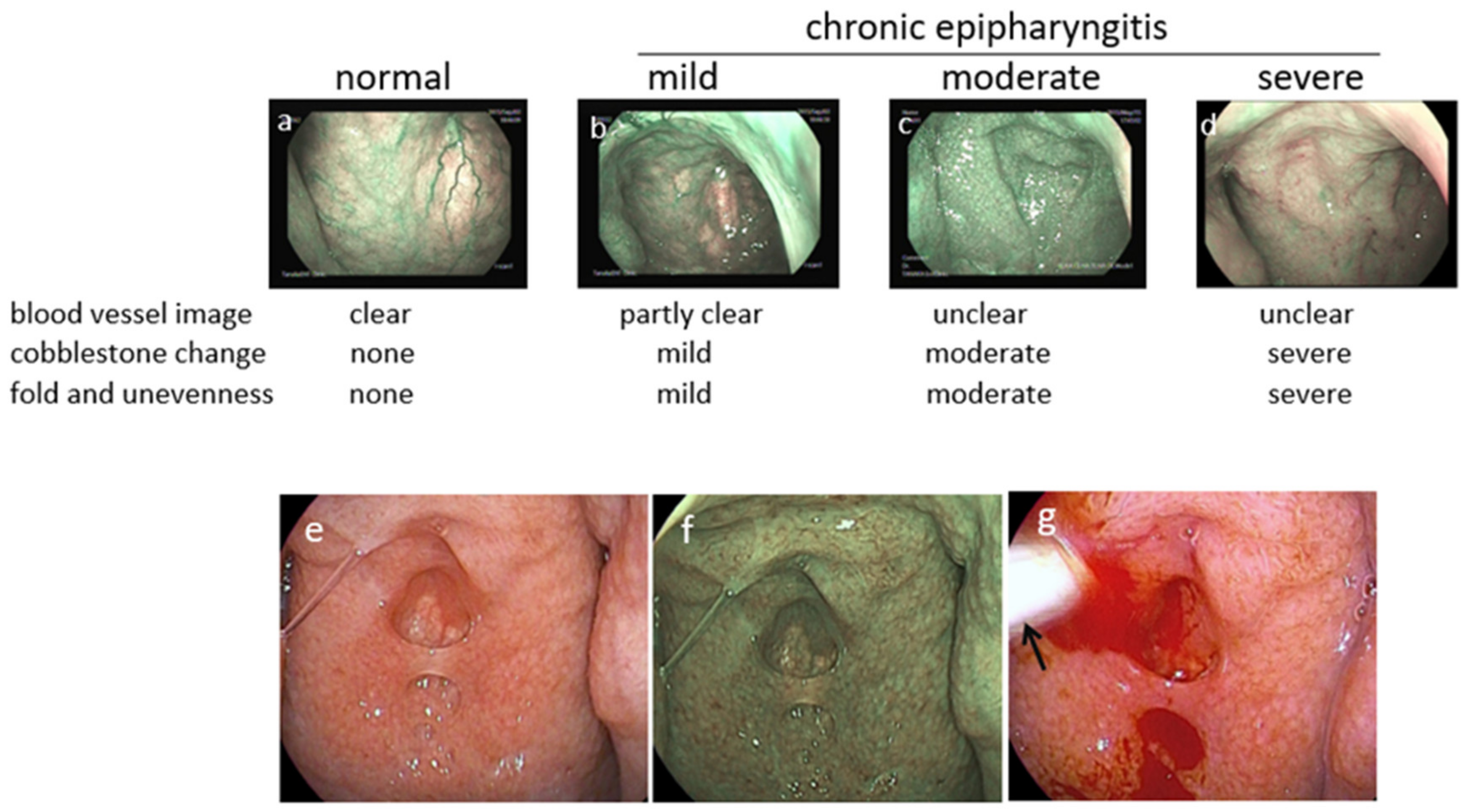
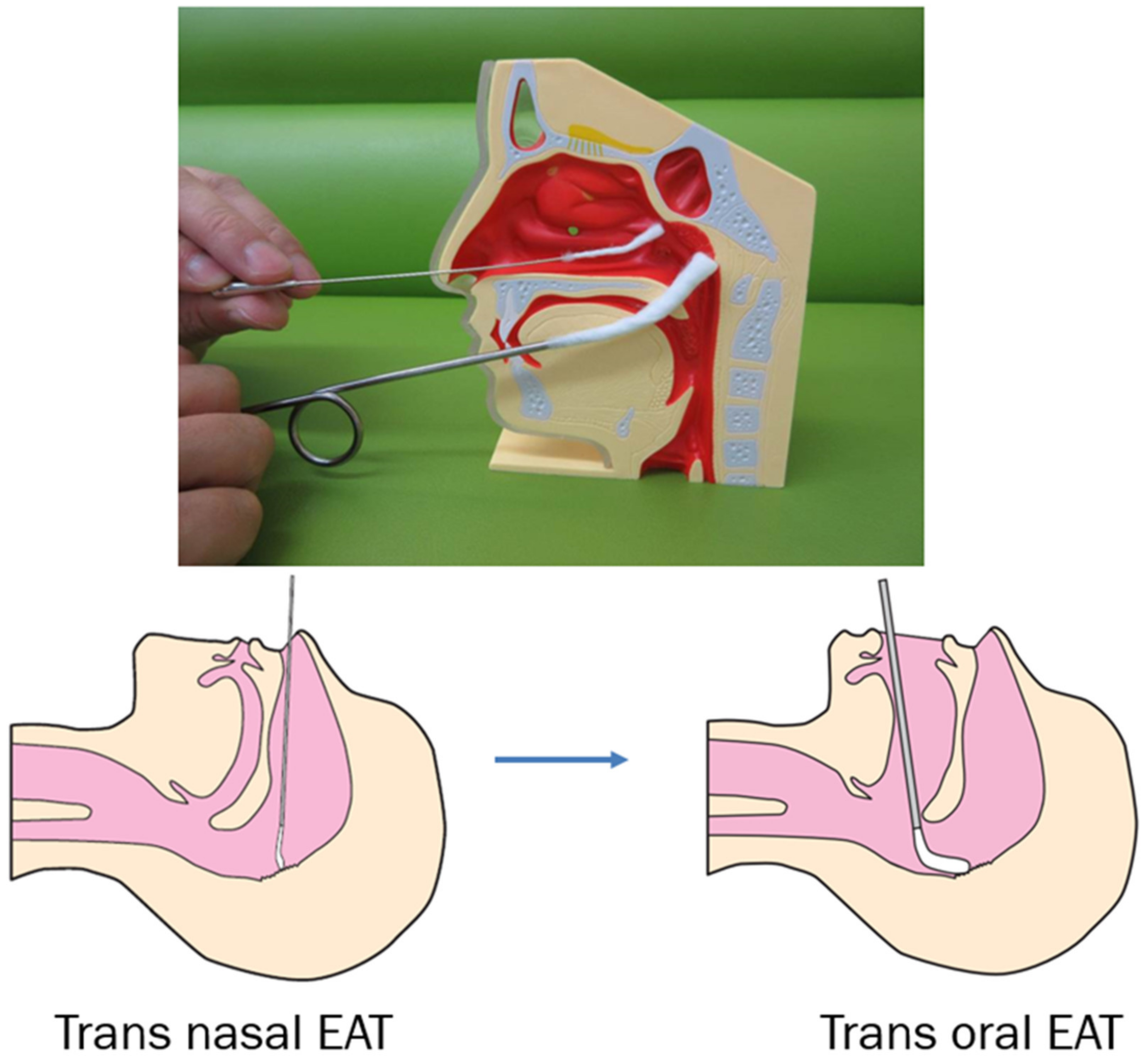
3.4. Diagnosis of Epipharyngitis
3.5. Treatment of Chronic Epipharyngitis
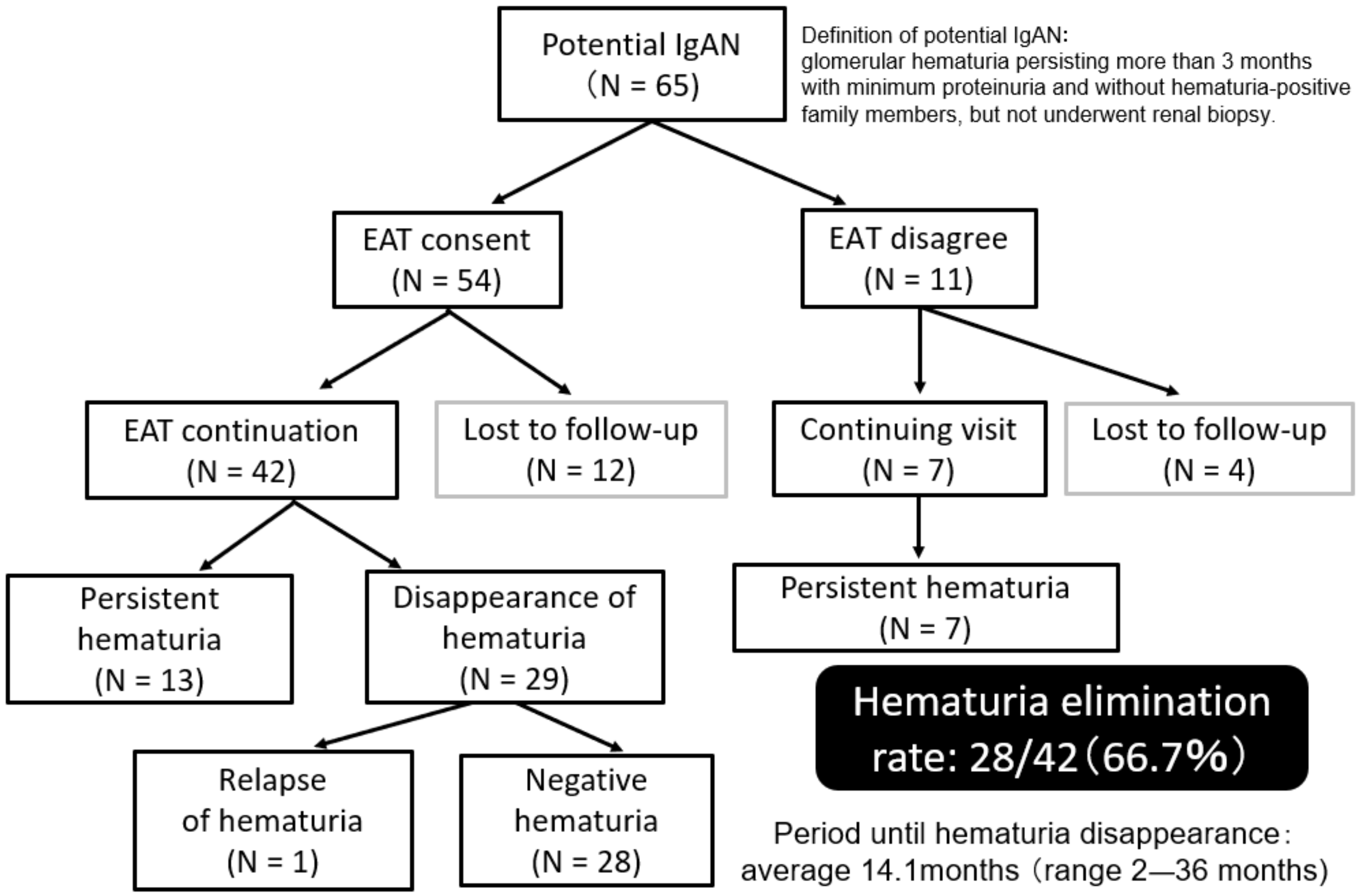
3.6. Effects of EAT on Refractory IgAN and Potential IgAN
4. Treatment Options Based on the Glomerular vasculitis Related Conditions of IgAN
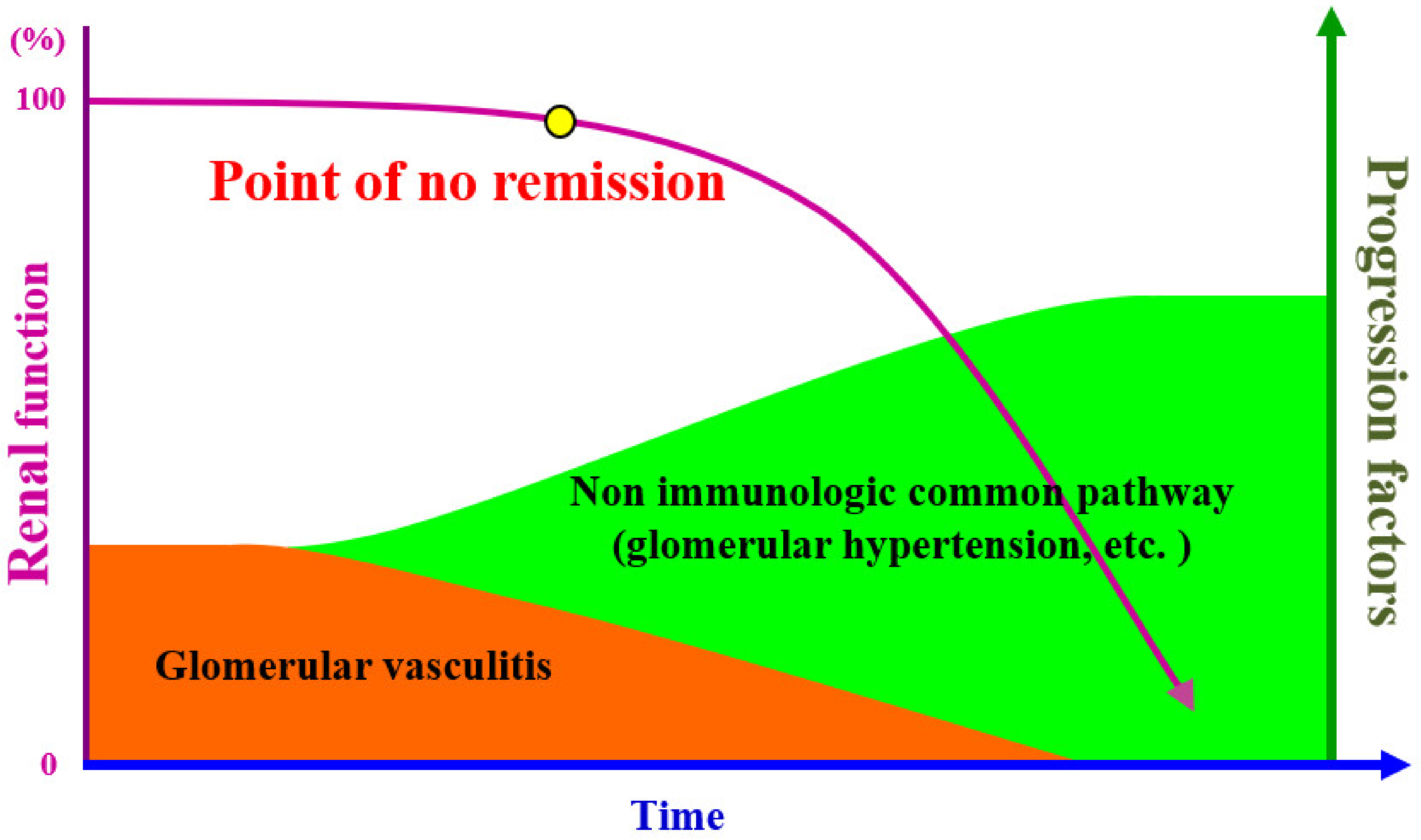
5. Conclusions and Future Direction
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Suzuki, K.; Honda, K.; Tanabe, K.; Toma, H.; Nihei, H.; Yamaguchi, Y. Incidence of latent mesangial IgA deposition in renal allograft donors in Japan. Kidney Int. 2003, 63, 2286–2294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellur, S.S.; Troyanov, S.; Cook, H.T.; Roberts, I.S.; Working Group of International IgA Nephropathy Network and Renal Pathology Society. Immunostaining findings in IgA nephropathy: Correlation with histology and clinical outcome in the Oxford classification patient cohort. Nephrol. Dial. Transplant. 2011, 26, 2533–2536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholls, K.; Walker, R.G.; Dowling, J.P.; Kincaid-Smith, P. “Malignant” IgA nephropathy. Am. J. Kidney Dis. 1985, 5, 42–46. [Google Scholar] [CrossRef]
- Kincaid-Smith, P.; Nicholls, K.; Birchall, I. Polymorphs infiltrate glomeruli in mesangial IgA glomerulonephritis. Kidney Int. 1989, 36, 1108–1111. [Google Scholar] [CrossRef] [Green Version]
- Tewari, R.; Nada, R.; Kaur, M.; Dudeja, P.; Rayat, C.S.; Sakhuja, V.; Joshi, K. Correlates of hematuria on glomerular histology and electron microscopy in IgA nephropathy. Med. J. Armed Forces India 2016, 72, 120–124. [Google Scholar] [CrossRef] [Green Version]
- Nagai, M.; Saitoh, S.; Takaki, T.; Ohbayashi, T.; Hotta, O.; Ohno, N.; Joh, K. Glomerular cellular interactions following disruption of the glomerular basement membrane in IgA nephropathy: Ultrastructural analyses by 3-dimensional serial block-face scanning electron microscopy. Kidney Med. 2020, 2, 222–225. [Google Scholar] [CrossRef]
- Takaki, T.; Ohno, N.; Saitoh, S.; Nagai, M.; Joh, K. Podocyte penetration of the glomerular basement membrane to contact on the mesangial cell at the lesion of mesangial interposition in lupus nephritis: A three-dimensional analysis by serial block-face scanning electron microscopy. Clin. Exp. Nephrol. 2019, 23, 773–781. [Google Scholar] [CrossRef]
- Hotta, O.; Taguma, Y.; Sudo, K.; Kurosawa, K. Limitation of kidney biopsy in detecting crescentic lesions in IgA nephropathy. Nephron 1993, 65, 472–473. [Google Scholar] [CrossRef]
- Sevillano, A.M.; Gutiérrez, E.; Yuste, C.; Cavero, T.; Mérida, E.; Rodríguez, P.; García, A.; Morales, E.; Fernández, C.; Martínez, M.A.; et al. Remission of hematuria Improves Renal Survival in IgA Nephropathy. J. Am. Soc. Nephrol. 2017, 28, 3089–3099. [Google Scholar] [CrossRef]
- Yu, G.Z.; Guo, L.; Dong, J.F.; Shi, S.F.; Liu, L.J.; Wang, J.W.; Sui, G.L.; Zhou, X.J.; Xing, Y.; Li, H.X. Persistent Hematuria and Kidney Disease Progression in IgA Nephropathy: A Cohort Study. Am. J. Kidney Dis. 2020, 76, 90–99. [Google Scholar] [CrossRef]
- Trimarchi, H.; Barratt, J.; Cattran, D.C.; Cook, H.T.; Coppo, R.; Haas, M.; Liu, Z.H.; Roberts, I.S.; Yuzawa, Y.; Zhang, H.; et al. Oxford Classification of IgA nephropathy 2016: An update from the IgA Nephropathy Classification Working Group. Kidney Int. 2017, 91, 1014–1021. [Google Scholar] [CrossRef] [Green Version]
- Hotta, O.; Furuta, T.; Chiba, S.; Tomioka, S.; Taguma, Y. Regression of IgA nephropathy: A repeat biopsy study. Am. J. Kidney Dis. 2002, 39, 493–502. [Google Scholar] [CrossRef]
- Ieiri, N.; Hotta, O.; Sato, T.; Taguma, Y. Significance of the duration of nephropathy for achieving clinical remission in patients with IgA nephropathy treated by tonsillectomy and steroid pulse therapy. Clin. Exp. Nephrol. 2012, 16, 122–129. [Google Scholar] [CrossRef]
- Sato, Y.; Hotta, O.; Taguma, Y.; Takasaka, T.; Nose, M. IgA nephropathy with poorly developed lymphoepithelial symbiosis of the palatine tonsils. Nephron 1996, 74, 301–308. [Google Scholar] [CrossRef]
- Joh, K.; Hotta, O.; Watanebe, K. Characteristics of IgA nephropathy—Associated tonsillitis and its relation to glomerulonephritis. Stomato-Pharyngology 2014, 27, 109–114. [Google Scholar]
- Takechi, H.; Oda, T.; Hotta, O.; Yamamoto, K.; Oshima, N.; Matsunobu, T.; Shiotani, A.; Nagura, H.; Shimazaki, H.; Tamai, S.; et al. Clinical and immunological implications of increase in CD208+ dendritic cells in tonsils of patients with immunoglobulin A nephropathy. Nephrol. Dial. Transplant. 2013, 28, 3004–3013. [Google Scholar] [CrossRef] [Green Version]
- Takahara, M.; Kishibe, K.; Nozawa, H.; Harabuchi, Y. Increase of activated T-cells and up-regulation of Smad7 without elevation of TGF-beta expression in tonsils from patients with pustulosis palmaris et plantaris. Clin. Immunol. 2005, 115, 192–199. [Google Scholar] [CrossRef] [Green Version]
- Hotta, O.; Miyazaki, M.; Furuta, T.; Tomioka, S.; Chiba, S.; Horigome, I.; Abe, K.; Taguma, Y. Tonsillectomy and steroid pulse therapy significantly impact on clinical remission in patients with IgA nephropathy. Am. J. Kidney Dis. 2001, 38, 736–743. [Google Scholar] [CrossRef]
- Kawamura, T.; Yoshimura, M.; Miyazaki, Y.; Okamoto, H.; Kimura, K.; Hirano, K.; Matsushima, M.; Utsunomiya, Y.; Ogura, M.; Yokoo, T.; et al. A multicenter randomized controlled trial of tonsillectomy combined with steroid pulse therapy in patients with immunoglobulin A nephropathy. Nephrol. Dial. Transplant. 2014, 29, 1546–1553. [Google Scholar] [CrossRef]
- Komatsu, H.; Sato, Y.; Miyamoto, T.; Tamura, M.; Nakata, T.; Tomo, T.; Nishino, T.; Miyazaki, M.; Fujimoto, S. Significance of tonsillectomy combined with steroid pulse therapy for IgA nephropathy with mild proteinuria. Clin. Exp. Nephrol. 2016, 20, 94–102. [Google Scholar] [CrossRef] [Green Version]
- Yuzawa, Y.; Yamamoto, R.; Takahashi, K.; Katafuchi, R.; Tomita, M.; Fujigaki, Y.; Kitamura, H.; Goto, M.; Yasuda, T.; Sato, M.; et al. Evidence-based clinical practice guidelines for IgA nephropathy 2014. Clin. Exp. Neph. 2016, 20, 511–535. [Google Scholar] [CrossRef] [Green Version]
- Harabuchi, Y.; Takahara, M. Recent advances in the immunological understanding of association between tonsil and immunoglobulin A nephropathy as a tonsil-induced autoimmune/inflammatory syndrome. Immun. Inflamm. Dis. 2019, 7, 86–92. [Google Scholar] [CrossRef] [Green Version]
- Goto, T.; Bandoh, N.; Yoshizaki, T.; Nozawa, H.; Takahara, M.; Ueda, S.; Hayashi, T.; Harabuchi, Y. Increase in B-cell-activation factor (BAFF) and IFN-gamma productions by tonsillar mononuclear cells stimulated with deoxycytidyl-deoxyguanosine oligodeoxynucleotides (CpG-ODN) in patients with IgA nephropathy. Clin. Immunol. 2008, 126, 260–269. [Google Scholar] [CrossRef] [Green Version]
- Muto, M.; Manfroi, B.; Suzuki, H.; Joh, K.; Nagai, M.; Wakai, S.; Righini, C.; Maiguma, M.; Izui, S.; Tomino, Y.; et al. Toll-like receptor 9 stimulation induces aberrant expression of a proliferation-inducing ligand by tonsillar germinal center B cells in IgA nephropathy. J. Am. Soc. Nephrol. 2017, 28, 1227–1238. [Google Scholar] [CrossRef] [Green Version]
- Takahara, M.; Nagato, T.; Nozaki, Y.; Kumai, T.; Katada, A.; Hayashi, T.; Harabuchi, Y. A proliferation-inducing ligand (APRIL) induced hyper-production of IgA from tonsillar mononuclear cells in patients with IgA nephropathy. Cell. Immunol. 2019, 341, 103925. [Google Scholar] [CrossRef]
- Takechi, H.; Nagura, H.; Houzawa, K.; Adachi, M.; Hotta, O. Clinical and immunological study of the tonsils in IgA nephropathy. Contrib. Nephrol. 2007, 157, 197. [Google Scholar]
- Kawaguchi, M.; Sakai, T.; Sakamaki, A.; Ishizawa, S.; Baba, Y.; Masuda, Y.; Koizumi, F. Expanded primary T nodules in the palatine tonsils from patients with IgA nephropathy. Acta Oto-Laryngol. Suppl. 1993, 50, 36–42. [Google Scholar] [CrossRef]
- Salaun, B.; de Saint-Vis, B.; Pacheco, N.; Pacheco, Y.; Riesler, A.; Isaac, S.; Leroux, C.; Clair-Moninot, V.; Pin, J.J.; Griffith, J.; et al. CD208/dendritic cell-lysosomal associated membrane protein is a marker of normal and transformed type II pneumocytes. Am. J. Pathol. 2004, 164, 861–871. [Google Scholar] [CrossRef] [Green Version]
- Imai, T.; Hieshima, K.; Haskell, C.; Baba, M.; Nagira, M.; Nishimura, M.; Kakizaki, M.; Takagi, S.; Nomiyama, H.; Schall, T.J.; et al. Identification and molecular characterization of fractalkine receptor CX3CR1, which mediates both leukocyte migration and adhesion. Cell 1997, 91, 521–530. [Google Scholar] [CrossRef] [Green Version]
- Jamieson, W.L.; Shimizu, S.; D’Ambrosio, J.A.; Meucci, O.; Fatatis, A. CX3CR1 is expressed by prostate epithelial cells and androgens regulate the levels of CX3CL1/fractalkine in the bone marrow: Potential role in prostate cancer bone tropism. Cancer Res. 2008, 68, 1715–1722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, G.E.; Xia, Y.; Chen, S.; Wang, Y.; Ye, R.D.; Harrison, J.K.; Bacon, K.B.; Zerwes, H.G.; Feng, L. NF-kappaB-dependent fractalkine induction in rat aortic endothelial cells by IL-1beta, TNF-alpha, and LPS. J. Leukoc. Biol. 2000, 67, 577–584. [Google Scholar] [CrossRef]
- Bjerkeli, V.; Halvorsen, B.; Damas, J.K.; Nordøy, I.; Yndestad, A.; Aukrust, P.; Frøland, S.S.; Halvorsen, B.; Damas, J.K.; Nordøy, I.; et al. Increased expression of fractalkine (CX3CL1) and its receptor, CX3CR1, in Wegener’s granulomatosis--possible role in vascular inflammation. Rheumatology 2007, 46, 1422–1427. [Google Scholar] [CrossRef] [Green Version]
- Cox, S.N.; Sallustio, F.; Serino, G.; Loverre, A.; Pesce, F.; Gigante, M.; Zaza, G.; Stifanelli, P.F.; Ancona, N.; Schena, F.P. Activated innate immunity and the involvement of CX3CR1—Fractalkine in promoting hematuria in patients with IgA nephropathy. Kidney Int. 2012, 82, 548–560. [Google Scholar] [CrossRef] [Green Version]
- Iwatani, H.; Nagasawa, Y.; Yamamoto, R.; Iio, K.; Mizui, M.; Horii, A.; Kitahara, T.; Inohara, H.; Kumanogoh, A.; Imai, E.; et al. CD16+CD56+ cells are a potential culprit for hematuria in IgA nephropathy. Clin. Exp. Nephrol. 2015, 19, 216–224. [Google Scholar] [CrossRef]
- Hotta, O.; Taguma, Y.; Yusa, N.; Ooyama, M. Analysis of mononuclear cells in urine using flow cytometry in glomerular diseases. Kidney Int. Suppl. 1994, 47, S117–S121. [Google Scholar]
- Otaka, R.; Takahara, M.; Ueda, S.; Nagato, T.; Kishibe, K.; Nomura, K.; Katada, A.; Hayashi, T.; Harabuchi, Y. Up-regulation of CX3CR1 on tonsillar CD8-positive cells in patients with IgA nephropathy. Hum. Immunol. 2017, 78, 375–383. [Google Scholar] [CrossRef]
- Vergano, L.; Loiacono, E.; Albera, R.; Coppo, R.; Camilla, R.; Peruzzi, L.; Amore, A.; Donadio, M.E.; Chiale, F.; Boido, A.; et al. Can tonsillectomy modify the innate and adaptive immunity pathways involved in IgA nephropathy? J. Nephrol. 2015, 28, 51–58. [Google Scholar] [CrossRef]
- Bourges, D.; Wang, C.H.; Chevaleyre, C.; Salmon, H. T and IgA B lymphocytes of the pharyngeal and palatine tonsils: Differential expression of adhesion molecules and chemokines. Scand. J. Immunol. 2004, 60, 338–350. [Google Scholar] [CrossRef]
- Sugita, R. A diagnostic method of the nasopharyngitis and a follow-up by cytology. Stomato-Pharyngology 2010, 23, 23–35. [Google Scholar]
- Hotta, O.; Tanaka, A.; Torigoe, A.; Imai, K.; Ieiri, N.; Japanese Focal Inflammation Research Group. Involvement of chronic epipharyngitis in autoimmune (auto-inflammatory) syndrome induced by adjuvants (ASIA). Immunol. Res. 2017, 65, 66–71. [Google Scholar] [CrossRef]
- Takano, K.; Kojima, T.; Go, M.; Murata, M.; Ichimiya, S.; Himi, T.; Sawada, N. HLA-DR- and CD11c-positive dendritic cells penetrate beyond well-developed epithelial tight junctions in human nasal mucosa of allergic rhinitis. J. Histochem. Cytochem. 2005, 53, 611–619. [Google Scholar] [CrossRef] [Green Version]
- Fujimura, Y. Evidence of M cells as portals of entry for antigens in the nasopharyngeal lymphoid tissue of humans. Virchows Arch. 2000, 436, 560–566. [Google Scholar] [CrossRef]
- Boyaka, P.N.; Wright, P.F.; Marinaro, M.; Kiyono, H.; Johnson, J.E.; Gonzales, R.A.; Ikizler, M.R.; Werkhaven, J.A.; Jackson, R.J.; Fujihashi, K.; et al. Human nasopharyngeal-associated lymphoreticular tissues. Functional analysis of subepithelial and intraepithelial B and T cells from adenoids and tonsils. Am. J. Pathol. 2000, 157, 2023–2035. [Google Scholar] [CrossRef]
- Hotta, O. Chronic epipharyngitis and its possible focal infection role. Stomato-Pharyngology 2010, 23, 37–42. [Google Scholar]
- Iwasaki, A.; Medzhitov, R. Toll-like receptor control of the adaptive immune responses. Nat. Immun. 2004, 5, 987–995. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [Green Version]
- Gankovskaya, L.V.; Bykova, V.P.; Namasova-Baranova, L.S.; Karaulov, A.V.; Rahmanova, I.V.; Gankovskii, V.A.; Merkushova, C.D.; Svitich, O.A. Innate immunity gene expression by epithelial cells of upper respiratory tract in children with adenoid hypertrophy. Auris Nasus Larynx 2018, 45, 753–759. [Google Scholar] [CrossRef]
- Suzuki, H.; Suzuki, Y.; Narita, I.; Aizawa, M.; Kihara, M.; Yamanaka, T.; Kanou, T.; Tsukaguchi, H.; Novak, J.; Horikoshi, S.; et al. Toll-like receptor 9 affects severity of IgA nephropathy. J. Am. Soc. Nephrol. 2008, 19, 2384–2395. [Google Scholar] [CrossRef] [Green Version]
- Coppo, R.; Camilla, R.; Amore, A.; Peruzzi, L.; Daprà, V.; Loiacono, E.; Vatrano, S.; Rollino, C.; Sepe, V.; Rampino, T.; et al. Toll-like receptor 4 expression is increased in circulating mononuclear cells of patients with immunoglobulin A nephropathy. Clin. Exp. Immunol. 2009, 159, 73–81. [Google Scholar] [CrossRef]
- Chang, H.; Yu, D.S.; Liu, X.Q.; Zhang, Q.Y.; Cheng, N.; Zhang, S.Q.; Qu, Z.H. Clinical significance of TLR3 and TLR4 in peripheral blood mononuclear cells from children with Henoch-Schönlein purpura nephritis. Exp. Ther. Med. 2014, 7, 1703–1707. [Google Scholar] [CrossRef] [Green Version]
- Donadio, M.E.; Loiacono, E.; Peruzzi, L.; Amore, A.; Camilla, R.; Camilla, R.; Chiale, F.; Vergano, L.; Boido, A.; Conrieri, M.; et al. Toll-like receptors, immunoproteasome and regulatory T cells in children with Henoch-Schönlein purpura and primary IgA nephropathy. Pediatr. Nephrol. 2014, 29, 1545–1551. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Peng, X.; Liu, Y.; Liu, H.; Liu, F.; He, L.; Liu, Y.; Zhang, F.; Guo, C.; Chen, G.; et al. TLR9 and BAFF: Their expression in patients with IgA nephropathy. Mol. Med. Rep. 2014, 10, 1469–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, A.; Komatsuda, A.; Kaga, H.; Sato, R.; Togashi, M.; Okuyama, S.; Wakui, H.; Takahashi, N. Different expression patterns of Toll-like receptor mRNAs in blood mononuclear cells of IgA Nephropathy and IgA Vasculitis with Nephritis. Tohoku J. Exp. Med. 2016, 240, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Hotta, O.; Tanaka, A.; Oda, T. Chronic epipharyngitis: A missing background of IgA nephropathy. Autoimmun. Rev. 2019, 18, 835–836. [Google Scholar] [CrossRef]
- Hotta, O.; Oda, T. The epipharynx-kidney axis triggers glomerular vasculitis in immunoglobulin nephropathy. Immunol. Res. 2019, 67, 304–309. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, W.; Kolls, J.K.; Zheng, Y. The biological functions of T helper 17 cell effector cytokines in inflammation. Immunity 2008, 28, 454–467. [Google Scholar] [CrossRef] [Green Version]
- Laan, M.; Cui, Z.H.; Hoshino, H.; Lötvall, J.; Sjöstrand, M.; Gruenert, D.C.; Skoogh, B.E.; Lindén, A. Neutrophil recruitment by human IL-17 via C-X-C chemokine release in the airways. J. Immunol. 1999, 162, 2347–2352. [Google Scholar]
- Peng, M.Y.; Wang, Z.H.; Yao, C.Y.; Jiang, L.N.; Jin, Q.L.; Wang, J.; Li, B.Q. Interleukin 17-producing gamma delta T cells increased in patients with active pulmonary tuberculosis. Cell. Mol. Immunol. 2008, 5, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, N.; Vanlaere, I.; de Rycke, R.; Cauwels, A.; Joosten, L.A.; Lubberts, E.; van den Berg, W.B.; Libert, C. IL-17 produced by Paneth cells drives TNF-induced shock. J. Exp. Med. 2008, 205, 1755–1761. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Chen, B.; Shi, J.; Chen, F.; Zhang, J.; Sun, Z. Analysis of regulatory T cell subsets in the peripheral blood of immunoglobulin A nephropathy (IgAN) patients. Genet. Mol. Res. 2015, 14, 14088–14092. [Google Scholar] [CrossRef]
- Iwakura, Y.; Ishigame, H.; Saijo, S.; Nakae, S. Functional specialization of interleukin-17 family members. Immunity 2011, 34, 149–162. [Google Scholar] [CrossRef] [Green Version]
- Hidalgo, A.; Peired, A.J.; Wild, M.; Vestweber, D.; Frenette, P.S. Complete identification of E-selectin ligands on neutrophils reveals distinct functions of PSGL-1, ESL-1, and CD44. Immunity 2007, 26, 477–489. [Google Scholar] [CrossRef] [Green Version]
- Gong, Y.; Zhang, Y.; Feng, S.; Liu, X.; Lü, S.; Long, M. Dynamic contributions of P- and E-selectins to β2-integrin-induced neutrophil transmigration. FASEB J. 2017, 31, 212–223. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.; Hino, M.; Kutsuna, H.; Hato, F.; Sakamoto, C.; Takahashi, T.; Tatsumi, N.; Kitagawa, S. Selective activation of p38 mitogen-activated protein kinase cascade in human neutrophils stimulated by IL-1β. J. Immunol. 2001, 167, 5940–5947. [Google Scholar] [CrossRef] [Green Version]
- Horiguchi, S. The discovery of the nasopharyngitis and its influence on general diseases. Acta Oto-Laryngol. Suppl. 1975, 329, 1–120. [Google Scholar]
- Hotta, O.; Inoue, C.N.; Tanaka, A.; Ieiri, N. The possible mechanism underling epipharyngeal abrasive therapy (EAT) with ZnCl2 solution for neuroendocrine disorders and autoimmune diseases. J. Antivir. Antiretrovir. 2017, 9, 81–86. [Google Scholar] [CrossRef]
- Neumann, H.; Fujishiro, M.; Wilcox, C.M.; Mönkemüller, K. Present and future perspectives of virtual chromoendoscopy with i-scan and optical enhancement technology. Dig. Endosc. 2014, 26 (Suppl. S1), 43–51. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaki, K.; Suzuki, Y.; Nakata, J.; Sakamoto, N.; Horikoshi, S.; Kawamura, T.; Matsuo, S.; Tomino, Y. Nationwide survey on current treatments for IgA nephropathy in Japan. Clin. Exp. Nephrol. 2013, 17, 827–833. [Google Scholar] [CrossRef] [Green Version]
- Hotta, O. Use of corticosteroids, other immunosuppressive therapies, and tonsillectomy in the treatment of IgA nephropathy. Semin Nephrol. 2004, 24, 244–255. [Google Scholar] [CrossRef]
- Hotta, O.; Tanaka, A.; Tani, S. Epipharyngeal treatment in systemic disease: Anticipated breakthrough. Stomato-Pharyngology 2016, 29, 99–106. [Google Scholar]
- Hotta, O. Involvement of epipharynx-kidney axis in chronic kidney disease. Stomato-Pharyngology 2020, 33, 17–23. [Google Scholar]
- Heerspink, H.J.L.; Stefánsson, B.V.; Correa-Rotter, R.; Chertow, G.M.; Greene, T.; Hou, F.F.; Mann, J.F.E.; McMurray, J.J.V.; Lindberg, M.; Rossing, P.; et al. DAPA-CKD Trial Committees and Investigators. Dapagliflozin in patients with chronic kidney disease. N. Engl. J. Med. 2020, 383, 1436–1446. [Google Scholar] [CrossRef]
- Barratt, J.; Floege, J. SGLT-2 inhibition in IgA nephropathy: The new standard of care? Kidney Int. 2021, 100, 24–26. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Epipharyngeal Lymphocytes | |||||
|---|---|---|---|---|---|
| Healthy Control | Acute Pharyngitis a | IgA Nephropathy | Tonsillar Lymphocytes | Peripheral Blood b | |
| n = 9 | n = 8 | n = 32 | n = 25 | n = 7 | |
| CD4 (%) | 29.1 | 35.2 | 41.7 ** | 39.2 | 42.2 |
| CD8 (%) | 8.4 | 9.9 | 12.1 | 9.3 | 29.8 |
| CD4/CD8 | 4.28 | 4.12 | 4.25 | 4.63 | 1.48 |
| CD20 (%) | 57.8 | 54.6 | 43.6* | 51.4 | 18.4 |
| CD4+HLA-DR+/CD4+ (%) | 30.5 | 53.3 *** | 40.3 **,# | 29.5 | 10.1 |
| CD8+HLA-DR+/CD8+ (%) | 41.6 | 66.4 ** | 58.9 ** | 45.6 | 23.0 |
| CD19+CD69+/CD19+ (%) | 41.4 | 30.1 | 29.1 | 46.9 | 0.8 |
| CD19+CD5+/CD19+ (%) | 4.1 | 10.5 ** | 12.8 ** | 20.0 | 22.4 |
| CD56 (%) | 0.6 | 1.0 | 1.2 | 1.5 | 15.4 |
| CD57 (%) | 1.3 | 0.5 | 2.2 | 5.3 | 14.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hotta, O.; Ieiri, N.; Nagai, M.; Tanaka, A.; Harabuchi, Y. Role of Palatine Tonsil and Epipharyngeal Lymphoid Tissue in the Development of Glomerular Active Lesions (Glomerular vasculitis) in Immunoglobulin A Nephropathy. Int. J. Mol. Sci. 2022, 23, 727. https://doi.org/10.3390/ijms23020727
Hotta O, Ieiri N, Nagai M, Tanaka A, Harabuchi Y. Role of Palatine Tonsil and Epipharyngeal Lymphoid Tissue in the Development of Glomerular Active Lesions (Glomerular vasculitis) in Immunoglobulin A Nephropathy. International Journal of Molecular Sciences. 2022; 23(2):727. https://doi.org/10.3390/ijms23020727
Chicago/Turabian StyleHotta, Osamu, Norio Ieiri, Masaaki Nagai, Ayaki Tanaka, and Yasuaki Harabuchi. 2022. "Role of Palatine Tonsil and Epipharyngeal Lymphoid Tissue in the Development of Glomerular Active Lesions (Glomerular vasculitis) in Immunoglobulin A Nephropathy" International Journal of Molecular Sciences 23, no. 2: 727. https://doi.org/10.3390/ijms23020727