
Transcriptional Basis of Psoriasis from Large Scale Gene Expression Studies: The Importance of Moving towards a Precision Medicine Approach


Abstract
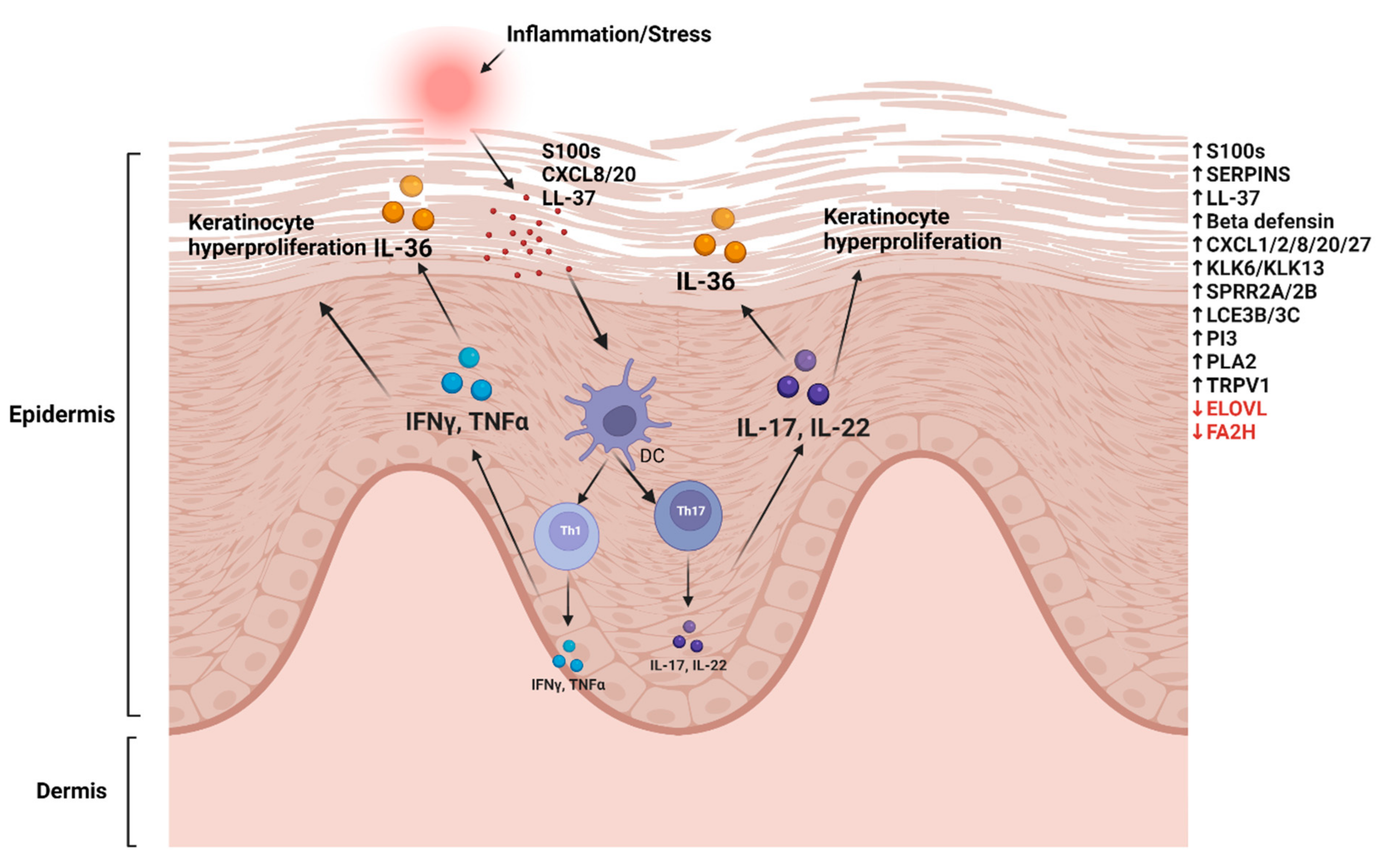
:1. Introduction
2. Large Cohort Transcriptome Analysis of Psoriasis
3. Alternative Splicing Variants: Psoriasis Isoforms
4. Neutrophils and Blood Transcriptome Signature in Psoriasis
5. Precision Medicine in Psoriasis Management
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mehrmal, S.; Uppal, P.; Nedley, N.; Giesey, R.L.; Delost, G.R. The global, regional, and national burden of psoriasis in 195 countries and territories, 1990 to 2017: A systematic analysis from the Global Burden of Disease Study 2017. J. Am. Acad. Dermatol. 2021, 84, 46–52. [Google Scholar] [CrossRef]
- Michalek, I.M.; Loring, B.; John, S. A systematic review of worldwide epidemiology of psoriasis. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 205–212. [Google Scholar] [CrossRef]
- Sahi, F.M.; Masood, A.; Danawar, N.A.; Mekaiel, A.; Malik, B.H. Association between Psoriasis and Depression: A Traditional Review. Cureus 2020, 12, e9708. [Google Scholar] [CrossRef]
- Henseler, T.; Christophers, E. Psoriasis of early and late onset: Characterization of two types of psoriasis vulgaris. J. Am. Acad. Dermatol. 1985, 13, 450–456. [Google Scholar] [CrossRef]
- Farber, E.M.; Nall, M.L.; Watson, W. Natural History of Psoriasis in 61 Twin Pairs. Arch. Dermatol. 1974, 109, 207–211. [Google Scholar] [CrossRef]
- Duffy, D.L.; Spelman, L.S.; Martin, N.G. Psoriasis in Australian twins. J. Am. Acad. Dermatol. 1993, 29, 428–434. [Google Scholar] [CrossRef] [Green Version]
- Rioux, G.; Ridha, Z.; Simard, M.; Turgeon, F.; Guérin, S.L.; Pouliot, R. Transcriptome Profiling Analyses in Psoriasis: A Dynamic Contribution of Keratinocytes to the Pathogenesis. Genes 2020, 11, 1155. [Google Scholar] [CrossRef]
- Benhadou, F.; Mintoff, D.; Del Marmol, V. Psoriasis: Keratinocytes or Immune Cells—Which Is the Trigger? Dermatology 2019, 235, 91–100. [Google Scholar] [CrossRef]
- Stuart, P.E.; Nair, R.P.; Ellinghaus, E.; Ding, J.; Tejasvi, T.; Gudjonsson, S.A.; Li, Y.; Weidinger, S.; Eberlein, B.; Gieger, C.; et al. Genome-wide association analysis identifies three psoriasis susceptibility loci. Nat. Genet. 2010, 42, 1000–1004. [Google Scholar] [CrossRef]
- Sun, L.-D.; Cheng, H.; Wang, Z.-X.; Zhang, A.-P.; Wang, P.-G.; Xu, J.-H.; Zhu, Q.-X.; Zhou, H.-S.; Ellinghaus, E.; Zhang, F.-R.; et al. Association analyses identify six new psoriasis susceptibility loci in the Chinese population. Nat. Genet. 2010, 42, 1005–1009. [Google Scholar] [CrossRef] [Green Version]
- Ellinghaus, E.; Ellinghaus, D.; Stuart, P.E.; Nair, R.P.; Debrus, S.; Raelson, J.V.; Belouchi, M.; Fournier, H.; Reinhard, C.; Ding, J.; et al. Genome-wide association study identifies a psoriasis susceptibility locus at TRAF3IP2. Nat. Genet. 2010, 42, 991–995. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.P.; Duffin, K.C.; Helms, C.; Ding, J.; Stuart, P.E.; Goldgar, D.; Gudjonsson, J.E.; Li, Y.; Tejasvi, T.; Feng, B.J.; et al. Genome-wide scan reveals association of psoriasis with IL-23 and NF-κB pathways. Nat. Genet. 2009, 41, 199–204. [Google Scholar] [CrossRef] [Green Version]
- Tsoi, L.C.; Stuart, P.E.; Tian, C.; Gudjonsson, J.E.; Das, S.; Zawistowski, M.; Ellinghaus, E.; Barker, J.N.; Chandran, V.; Dand, N.; et al. Large scale meta-analysis characterizes genetic architecture for common psoriasis associated variants. Nat. Commun. 2017, 8, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsoi, L.C.; Spain, S.L.; Knight, J.; Ellinghaus, E.; Stuart, P.E.; Capon, F.; Ding, J.; Li, Y.; Tejasvi, T.; Gudjonsson, J.E.; et al. Identification of 15 new psoriasis susceptibility loci highlights the role of innate immunity. Nat. Genet. 2012, 44, 1341–1348. [Google Scholar] [CrossRef] [Green Version]
- Oestreicher, J.L.; Walters, I.B.; Kikuchi, T.; Gilleaudeau, P.; Surette, J.; Schwertschlag, U.; Dorner, A.J.; Krueger, J.G.; Trepicchio, W.L. Molecular classification of psoriasis disease-associated genes through pharmacogenomic expression profiling. Pharm. J. 2001, 1, 272–287. [Google Scholar] [CrossRef] [PubMed]
- Keermann, M.; Kõks, S.; Reimann, E.; Prans, E.; Abram, K.; Kingo, K. Transcriptional landscape of psoriasis identifies the involvement of IL36 and IL36RN. BMC Genom. 2015, 16, 322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reich, K.; Papp, K.A.; Matheson, R.T.; Tu, J.H.; Bissonnette, R.; Bourcier, M.; Gratton, D.; Kunynetz, R.A.; Poulin, Y.; Rosoph, L.A.; et al. Evidence that a neutrophil–keratinocyte crosstalk is an early target of IL -17A inhibition in psoriasis. Exp. Dermatol. 2015, 24, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Sotiropoulou, P.A.; Blanpain, C. Development and Homeostasis of the Skin Epidermis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008383. [Google Scholar] [CrossRef] [Green Version]
- Weinstein, G.D.; Van Scott, E.J. Autoradiographic analysis of turnover time and protein synthesis. J. Invest. Dermatol. 1965, 45, 257–262. [Google Scholar] [CrossRef] [Green Version]
- Nograles, K.E.; Zaba, L.C.; Guttman-Yassky, E.; Fuentes-Duculan, J.; Suárez-Fariñas, M.; Cardinale, I.; Khatcherian, A.; Gonzalez, J.; Pierson, K.C.; White, T.R.; et al. Th17 cytokines interleukin (IL)-17 and IL-22 modulate distinct inflammatory and keratinocyte-response pathways. Br. J. Dermatol. 2008, 159, 1092–1102. [Google Scholar] [CrossRef] [Green Version]
- Brembilla, N.C.; Senra, L.; Boehncke, W.-H. The IL-17 Family of Cytokines in Psoriasis: IL-17A and Beyond. Front. Immunol. 2018, 9, 1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harden, J.L.; Krueger, J.G.; Bowcock, A.M. The immunogenetics of Psoriasis: A comprehensive review. J. Autoimmun. 2015, 64, 66–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furue, K.; Yamamura, K.; Tsuji, G.; Mitoma, C.; Uchi, H.; Nakahara, T.; Kido-Nakahara, M.; Kadono, T.; Furue, M. Highlighting Interleukin-36 Signalling in Plaque Psoriasis and Pustular Psoriasis. Acta Derm. Venereol. 2018, 98, 5–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahn, M.; Frey, S.; Hueber, A.J. The novel interleukin-1 cytokine family members in inflammatory diseases. Curr. Opin. Rheumatol. 2017, 29, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Iznardo, H.; Puig, L. Exploring the Role of IL-36 Cytokines as a New Target in Psoriatic Disease. Int. J. Mol. Sci. 2021, 22, 4344. [Google Scholar] [CrossRef]
- Iznardo, H.; Puig, L. The interleukin-1 family cytokines in psoriasis: Pathogenetic role and therapeutic perspectives. Expert Rev. Clin. Immunol. 2021, 17, 187–199. [Google Scholar] [CrossRef]
- Madonna, S.; Girolomoni, G.; Dinarello, C.A.; Albanesi, C. The Significance of IL-36 Hyperactivation and IL-36R Targeting in Psoriasis. Int. J. Mol. Sci. 2019, 20, 3318. [Google Scholar] [CrossRef] [Green Version]
- Capon, F.; Semprini, S.; Novelli, G.; Chimenti, S.; Fabrizi, G.; Zambruno, G.; Murgia, S.; Carcassi, C.; Fazio, M.; Mingarelli, R.; et al. Fine Mapping of the PSORS4 Psoriasis Susceptibility Region on Chromosome 1q21. J. Investig. Dermatol. 2001, 116, 728–730. [Google Scholar] [CrossRef] [Green Version]
- Xia, C.; Braunstein, Z.; Toomey, A.C.; Zhong, J.; Rao, X. S100 Proteins As an Important Regulator of Macrophage Inflammation. Front. Immunol. 2017, 8, 1908. [Google Scholar] [CrossRef] [Green Version]
- Eckert, R.L.; Broome, A.-M.; Ruse, M.; Robinson, N.; Ryan, D.; Lee, K. S100 Proteins in the Epidermis. J. Investig. Dermatol. 2004, 123, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Tsoi, L.C.; Swindell, W.R.; Gudjonsson, J.E.; Tejasvi, T.; Johnston, A.; Ding, J.; Stuart, P.E.; Xing, X.; Kochkodan, J.J.; et al. Transcriptome Analysis of Psoriasis in a Large Case–Control Sample: RNA-Seq Provides Insights into Disease Mechanisms. J. Investig. Dermatol. 2014, 134, 1828–1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowcock, A.M.; Shannon, W.; Du, F.; Duncan, J.; Cao, K.; Aftergut, K.; Catier, J.; Fernandez-Vina, M.A.; Menter, A. Insights into psoriasis and other inflammatory diseases from large-scale gene expression studies. Hum. Mol. Genet. 2001, 10, 1793–1805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, I.; Gao, B.; Boguniewicz, M.; Darst, M.A.; Travers, J.B.; Leung, D.Y. Distinct patterns of gene expression in the skin lesions of atopic dermatitis and psoriasis: A gene microarray analysis. J. Allergy Clin. Immunol. 2003, 112, 1195–1202. [Google Scholar] [CrossRef] [PubMed]
- Nattkemper, L.A.; Tey, H.L.; Valdes-Rodriguez, R.; Lee, H.; Mollanazar, N.K.; Albornoz, C.; Sanders, K.M.; Yosipovitch, G. The Genetics of Chronic Itch: Gene Expression in the Skin of Patients with Atopic Dermatitis and Psoriasis with Severe Itch. J. Investig. Dermatol. 2018, 138, 1311–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Krueger, J.G.; Kao, M.-C.J.; Lee, E.; Du, F.; Menter, A.; Wong, W.H.; Bowcock, A.M. Novel mechanisms of T-cell and dendritic cell activation revealed by profiling of psoriasis on the 63,100-element oligonucleotide array. Physiol. Genom. 2003, 13, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Kulski, J.K.; Kenworthy, W.; Bellgard, M.; Taplin, R.; Okamoto, K.; Oka, A.; Mabuchi, T.; Ozawa, A.; Tamiya, G.; Inoko, H. Gene expression profiling of Japanese psoriatic skin reveals an increased activity in molecular stress and immune response signals. J. Mol. Med. 2005, 83, 964–975. [Google Scholar] [CrossRef]
- Gudjonsson, J.E.; Ding, J.; Johnston, A.; Tejasvi, T.; Guzman, A.M.; Nair, R.P.; Voorhees, J.J.; Abecasis, G.R.; Elder, J.T. Assessment of the Psoriatic Transcriptome in a Large Sample: Additional Regulated Genes and Comparisons with In Vitro Models. J. Investig. Dermatol. 2010, 130, 1829–1840. [Google Scholar] [CrossRef] [Green Version]
- Mee, J.B.; Johnson, C.M.; Morar, N.; Burslem, F.; Groves, R.W. The Psoriatic Transcriptome Closely Resembles That Induced by Interleukin-1 in Cultured Keratinocytes: Dominance of Innate Immune Responses in Psoriasis. Am. J. Pathol. 2007, 171, 32–42. [Google Scholar] [CrossRef] [Green Version]
- Bando, M.; Hiroshima, Y.; Kataoka, M.; Shinohara, Y.; Herzberg, M.C.; Ross, K.F.; Nagata, T.; Kido, J. Interleukin-1αregulates antimicrobial peptide expression in human keratinocytes. Immunol. Cell Biol. 2007, 85, 532–537. [Google Scholar] [CrossRef]
- Haider, A.S.; Lowes, M.A.; Suàrez-Fariñas, M.; Zaba, L.C.; Cardinale, I.; Blumenberg, M.; Krueger, J.G. Cellular Genomic Maps Help Dissect Pathology in Human Skin Disease. J. Investig. Dermatol. 2008, 128, 606–615. [Google Scholar] [CrossRef] [Green Version]
- Gudjonsson, J.E.; Ding, J.; Li, X.; Nair, R.P.; Tejasvi, T.; Qin, Z.; Ghosh, D.; Aphale, A.; Gumucio, D.L.; Voorhees, J.J.; et al. Global Gene Expression Analysis Reveals Evidence for Decreased Lipid Biosynthesis and Increased Innate Immunity in Uninvolved Psoriatic Skin. J. Investig. Dermatol. 2009, 129, 2795–2804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasquali, L.; Srivastava, A.; Meisgen, F.; Das Mahapatra, K.; Xia, P.; Landén, N.; Pivarcsi, A.; Sonkoly, E. The Keratinocyte Transcriptome in Psoriasis: Pathways Related to Immune Responses, Cell Cycle and Keratinization. Acta Derm. Venereol. 2019, 99, 196–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ainali, C.; Valeyev, N.; Perera, G.; Williams, A.; Gudjonsson, J.E.; Ouzounis, C.A.; Nestle, F.O.; Tsoka, S. Transcriptome classification reveals molecular subtypes in psoriasis. BMC Genom. 2012, 13, 472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suárez-Fariñas, M.; Lowes, M.A.; Zaba, L.C.; Krueger, J.G. Evaluation of the Psoriasis Transcriptome across Different Studies by Gene Set Enrichment Analysis (GSEA). PLoS ONE 2010, 5, e10247. [Google Scholar] [CrossRef] [Green Version]
- Mimoso, C.; Blumenberg, M.; Perel-Man, R. Looking within the lesion: Large scale transcriptional profiling of psoriatic plaques. World J. Dermatol. 2014, 3, 28–35. [Google Scholar] [CrossRef]
- Suárez-Fariñas, M.; Li, K.; Fuentes-Duculan, J.; Hayden, K.; Brodmerkel, C.; Krueger, J.G. Expanding the Psoriasis Disease Profile: Interrogation of the Skin and Serum of Patients with Moderate-to-Severe Psoriasis. J. Investig. Dermatol. 2012, 132, 2552–2564. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Richman, L.; Morehouse, C.; Reyes, M.D.L.; Higgs, B.; Boutrin, A.; White, B.; Coyle, A.; Krueger, J.; Kiener, P.A.; et al. Type I Interferon: Potential Therapeutic Target for Psoriasis? PLoS ONE 2008, 3, e2737. [Google Scholar] [CrossRef]
- Eriksen, K.W.; Lovato, P.; Skov, L.; Krejsgaard, T.; Kaltoft, K.; Geisler, C.; Ødum, N. Increased Sensitivity to Interferon-α in Psoriatic T Cells. J. Investig. Dermatol. 2005, 125, 936–944. [Google Scholar] [CrossRef] [Green Version]
- Johnson-Huang, L.M.; Suárez-Fariñas, M.; Pierson, K.C.; Fuentes-Duculan, J.; Cueto, I.; Lentini, T.; Sullivan-Whalen, M.; Gilleaudeau, P.; Krueger, J.G.; Haider, A.S.; et al. A Single Intradermal Injection of IFN-γ Induces an Inflammatory State in Both Non-Lesional Psoriatic and Healthy Skin. J. Investig. Dermatol. 2012, 132, 1177–1187. [Google Scholar] [CrossRef] [Green Version]
- Swindell, W.R.; Xing, X.; Stuart, P.E.; Chen, C.S.; Aphale, A.; Nair, R.P.; Voorhees, J.J.; Elder, J.T.; Johnston, A.; Gudjonsson, J.E. Heterogeneity of Inflammatory and Cytokine Networks in Chronic Plaque Psoriasis. PLoS ONE 2012, 7, e34594. [Google Scholar] [CrossRef]
- Swindell, W.R.; Johnston, A.; Voorhees, J.J.; Elder, J.T.; Gudjonsson, J.E. Dissecting the psoriasis transcriptome: Inflammatory- and cytokine-driven gene expression in lesions from 163 patients. BMC Genom. 2013, 14, 527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reischl, J.; Schwenke, S.; Beekman, J.M.; Mrowietz, U.; Stürzebecher, S.; Heubach, J.F. Increased Expression of Wnt5a in Psoriatic Plaques. J. Investig. Dermatol. 2007, 127, 163–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, S.; Krueger, J.G.; Li, K.; Jabbari, A.; Brodmerkel, C.; Lowes, M.A.; Suárez-Fariñas, M. Meta-Analysis Derived (MAD) Transcriptome of Psoriasis Defines the “Core” Pathogenesis of Disease. PLoS ONE 2012, 7, e44274. [Google Scholar] [CrossRef] [Green Version]
- Ewald, D.A.; Malajian, D.; Krueger, J.G.; Workman, C.T.; Wang, T.; Tian, S.; Litman, T.; Guttman-Yassky, E.; Suárez-Fariñas, M. Meta-analysis derived atopic dermatitis (MADAD) transcriptome defines a robust AD signature highlighting the involvement of atherosclerosis and lipid metabolism pathways. BMC Med. Genom. 2015, 8, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, D.; Yao, S.; Wu, H.; Ke, X.; Zhou, X.; Geng, S.; Dong, S.; Chen, H.; Yang, T.; Cheng, Y.; et al. A transcriptome-wide association study identifies novel susceptibility genes for psoriasis. Hum. Mol. Genet. 2021, 31, 300–308. [Google Scholar] [CrossRef]
- Boehncke, W.-H.; Schön, M.P. Psoriasis. Lancet 2015, 386, 983–994. [Google Scholar] [CrossRef]
- Xu, X.; Tang, X.; Zhang, Y.; Pan, Z.; Wang, Q.; Tang, L.; Zhu, C.; Cheng, H.; Zhou, F. Chromatin accessibility and transcriptome integrative analysis revealed AP-1-mediated genes potentially modulate histopathology features in psoriasis. Clin. Epigenetics 2022, 14, 38. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, J.; Huang, B.O.; Xu, Y.-M.; Li, J.; Huang, L.-F.; Lin, J.; Zhang, J.; Min, Q.-H.; Yang, W.-M.; et al. Mechanism of alternative splicing and its regulation. Biomed. Rep. 2015, 3, 152–158. [Google Scholar] [CrossRef] [Green Version]
- Reixachs-Solé, M.; Eyras, E. Uncovering the impacts of alternative splicing on the proteome with current omics techniques. Wiley Interdiscip. Rev. RNA 2022, e1707. [Google Scholar] [CrossRef]
- Barbosa-Morais, N.L.; Irimia, M.; Pan, Q.; Xiong, H.Y.; Gueroussov, S.; Lee, L.J.; Slobodeniuc, V.; Kutter, C.; Watt, S.; Çolak, R.; et al. The Evolutionary Landscape of Alternative Splicing in Vertebrate Species. Science 2012, 338, 1587–1593. [Google Scholar] [CrossRef] [Green Version]
- Pan, Q.; Shai, O.; Lee, L.J.; Frey, B.J.; Blencowe, B.J. Deep surveying of alternative splicing complexity in the human transcriptome by high-throughput sequencing. Nat. Genet. 2008, 40, 1413–1415. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.T.; Sandberg, R.; Luo, S.; Khrebtukova, I.; Zhang, L.; Mayr, C.; Kingsmore, S.F.; Schroth, G.P.; Burge, C.B. Alternative isoform regulation in human tissue transcriptomes. Nature 2008, 456, 470–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Yu, P. Genome-wide transcriptome analysis identifies alternative splicing regulatory network and key splicing factors in mouse and human psoriasis. Sci. Rep. 2018, 8, 4124. [Google Scholar] [CrossRef] [Green Version]
- Kõks, S.; Keermann, M.; Reimann, E.; Prans, E.; Abram, K.; Silm, H.; Kõks, G.; Kingo, K. Psoriasis-Specific RNA Isoforms Identified by RNA-Seq Analysis of 173,446 Transcripts. Front. Med. 2016, 3, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, R.; Lewerenz, V.; Büchau, A.S.; Walz, M.; Ruzicka, T. Human S100A15 splice variants are differentially expressed in inflammatory skin diseases and regulated through Th1 cytokines and calcium. Exp. Dermatol. 2007, 16, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Büchau, A.S.; Hassan, M.; Kukova, G.; Lewerenz, V.; Kellermann, S.; Würthner, J.U.; Wolf, R.; Walz, M.; Gallo, R.L.; Ruzicka, T. S100A15, an Antimicrobial Protein of the Skin: Regulation by E. coli through Toll-Like Receptor 4. J. Investig. Dermatol. 2007, 127, 2596–2604. [Google Scholar] [CrossRef] [Green Version]
- Wolf, R.; Mirmohammadsadegh, A.; Walz, M.; Lysa, B.; Tartler, U.; Remus, R.; Hengge, U.; Michel, G.; Ruzicka, T. Molecular cloning and characterization of alternatively spliced mRNA isoforms from psoriatic skin encoding a novel member of the S100 family. FASEB J. 2003, 17, 1969–1971. [Google Scholar] [CrossRef] [Green Version]
- Kerkhoff, C.; Voss, A.; Scholzen, T.E.; Averill, M.M.; Zänker, K.S.; Bornfeldt, K.E. Novel insights into the role of S100A8/A9 in skin biology. Exp. Dermatol. 2012, 21, 822–826. [Google Scholar] [CrossRef] [Green Version]
- Scudiero, I.; Zotti, T.; Ferravante, A.; Vessichelli, M.; Vito, P.; Stilo, R. Alternative splicing of CARMA2/CARD14 transcripts generates protein variants with differential effect on NF-κB activation and endoplasmic reticulum stress-induced cell death. J. Cell. Physiol. 2011, 226, 3121–3131. [Google Scholar] [CrossRef] [Green Version]
- Jordan, C.T.; Cao, L.; Roberson, E.; Duan, S.; Helms, C.A.; Nair, R.P.; Duffin, K.C.; Stuart, P.E.; Goldgar, D.; Hayashi, G.; et al. Rare and Common Variants in CARD14, Encoding an Epidermal Regulator of NF-kappaB, in Psoriasis. Am. J. Hum. Genet. 2012, 90, 796–808. [Google Scholar] [CrossRef] [Green Version]
- Jordan, C.T.; Cao, L.; Roberson, E.D.; Pierson, K.C.; Yang, C.-F.; Joyce, C.E.; Ryan, C.; Duan, S.; Helms, C.A.; Liu, Y.; et al. PSORS2 Is Due to Mutations in CARD14. Am. J. Hum. Genet. 2012, 90, 784–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Wang, C.; Boisson, B.; Misra, S.; Rayman, P.; Finke, J.H.; Puel, A.; Casanova, J.-L.; Li, X. The Differential Regulation of Human ACT1 Isoforms by Hsp90 in IL-17 Signaling. J. Immunol. 2014, 193, 1590–1599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Széll, M.; Bata-Csörgő, Z.; Koreck, A.; Pivarcsi, A.; Polyánka, H.; Szeg, C.; Gaál, M.; Dobozy, A.; Kemény, L. Proliferating Keratinocytes Are Putative Sources of the Psoriasis Susceptibility-Related EDA+(Extra Domain A of Fibronectin) Oncofetal Fibronectin. J. Investig. Dermatol. 2004, 123, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Szlavicz, E.; Szabo, K.; Groma, G.; Bata-Csorgo, Z.; Pagani, F.; Kemeny, L.; Szell, M. Erratum to: Splicing factors differentially expressed in psoriasis alter mRNA maturation of disease-associated EDA+ fibronectin. Mol. Cell. Biochem. 2017, 436, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawat, A.; Rinchai, D.; Toufiq, M.; Marr, A.K.; Kino, T.; Garand, M.; Tatari-Calderone, Z.; Kabeer, B.S.A.; Krishnamoorthy, N.; Bedognetti, D.; et al. A Neutrophil-Driven Inflammatory Signature Characterizes the Blood Transcriptome Fingerprint of Psoriasis. Front. Immunol. 2020, 11, 587946. [Google Scholar] [CrossRef]
- Schön, M.P.; Broekaert, S.M.C.; Erpenbeck, L. Sexy again: The renaissance of neutrophils in psoriasis. Exp. Dermatol. 2017, 26, 305–311. [Google Scholar] [CrossRef] [Green Version]
- Chiang, C.-C.; Cheng, W.-J.; Korinek, M.; Lin, C.-Y.; Hwang, T.-L. Neutrophils in Psoriasis. Front. Immunol. 2019, 10, 2376. [Google Scholar] [CrossRef]
- Rodriguez-Rosales, Y.A.; Langereis, J.D.; Gorris, M.A.; Reek, J.M.V.D.; Fasse, E.; Netea, M.G.; de Vries, I.J.M.; Gomez-Muñoz, L.; van Cranenbroek, B.; Körber, A.; et al. Immunomodulatory aged neutrophils are augmented in blood and skin of psoriasis patients. J. Allergy Clin. Immunol. 2021, 148, 1030–1040. [Google Scholar] [CrossRef]
- Wolberink, E.A.; Peppelman, M.; Van De Kerkhof, P.C.; Van Erp, P.E.; Gerritsen, M.-J.P. Establishing the dynamics of neutrophil accumulation in vivo by reflectance confocal microscopy. Exp. Dermatol. 2014, 23, 184–188. [Google Scholar] [CrossRef]
- Wang, L.; Yu, X.; Wu, C.; Zhu, T.; Wang, W.; Zheng, X.; Jin, H. RNA sequencing-based longitudinal transcriptomic profiling gives novel insights into the disease mechanism of generalized pustular psoriasis. BMC Med. Genom. 2018, 11, 52. [Google Scholar] [CrossRef] [Green Version]
- Garshick, M.S.; Barrett, T.J.; Wechter, T.; Azarchi, S.; Scher, J.U.; Neimann, A.; Katz, S.; Fuentes-Duculan, J.; Cannizzaro, M.V.; Jelic, S.; et al. Inflammasome Signaling and Impaired Vascular Health in Psoriasis. Arter. Thromb. Vasc. Biol. 2019, 39, 787–798. [Google Scholar] [CrossRef]
- Garshick, M.S.; Baumer, Y.; Dey, A.K.; Grattan, R.; Ng, Q.; Teague, H.L.; Yu, Z.-X.; Chen, M.Y.; Tawil, M.; Barrett, T.J.; et al. Characterization of PCSK9 in the Blood and Skin of Psoriasis. J. Investig. Dermatol. 2020, 141, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Krahel, J.A.; Baran, A.; Kamiński, T.W.; Maciaszek, M.; Flisiak, I. Methotrexate Decreases the Level of PCSK9—A Novel Indicator of the Risk of Proatherogenic Lipid Profile in Psoriasis. The Preliminary Data. J. Clin. Med. 2020, 9, 910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Wang, Y.; Zhao, J.; Zhou, D.; Lv, J.; Zhang, G.; Di, T.; Li, P. A study on the pathogenesis of blood-heat psoriasis with transcriptome analysis. Ann. Transl. Med. 2020, 8, 1523. [Google Scholar] [CrossRef]
- Coda, A.B.; Icen, M.; Smith, J.R.; Sinha, A.A. Global transcriptional analysis of psoriatic skin and blood confirms known disease-associated pathways and highlights novel genomic “hot spots” for differentially expressed genes. Genomics 2012, 100, 18–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.-W.; Yan, D.; Singh, R.; Bui, A.; Lee, K.; Truong, A.; Milush, J.M.; Somsouk, M.; Liao, W. Multi-omic Analysis of the Gut Microbiome in Psoriasis Reveals Distinct Host-Microbe Associations. JID Innov. 2022, 100115. [Google Scholar] [CrossRef]
- Da Rosa, J.C.; Kim, J.; Tian, S.; Tomalin, L.E.; Krueger, J.G.; Suárez-Fariñas, M. Shrinking the Psoriasis Assessment Gap: Early Gene-Expression Profiling Accurately Predicts Response to Long-Term Treatment. J. Investig. Dermatol. 2017, 137, 305–312. [Google Scholar] [CrossRef] [Green Version]
- Nishikawa, R.; Nagai, H.; Bito, T.; Ikeda, T.; Horikawa, T.; Adachi, A.; Matsubara, T.; Nishigori, C. Genetic prediction of the effectiveness of biologics for psoriasis treatment. J. Dermatol. 2016, 43, 1273–1277. [Google Scholar] [CrossRef] [Green Version]
- Tsoi, L.C.; Patrick, M.T.; Shuai, S.; Sarkar, M.K.; Chi, S.; Ruffino, B.; Billi, A.C.; Xing, X.; Uppala, R.; Zang, C.; et al. Cytokine responses in nonlesional psoriatic skin as clinical predictor to anti-TNF agents. J. Allergy Clin. Immunol. 2022, 149, 640–649.e5. [Google Scholar] [CrossRef]
- Foulkes, A.C.; Watson, D.S.; Carr, D.F.; Kenny, J.G.; Slidel, T.; Parslew, R.; Pirmohamed, M.; Anders, S.; Reynolds, N.J.; Griffiths, C.E.; et al. A Framework for Multi-Omic Prediction of Treatment Response to Biologic Therapy for Psoriasis. J. Investig. Dermatol. 2020, 139, 100–107. [Google Scholar] [CrossRef] [Green Version]
- Bagel, J.; Wang, Y.; Montgomery, I.P.; Abaya, C.; Andrade, E.; Boyce, C.; Tomich, T.; Lee, B.-I.; Pariser, D.; Menter, A.; et al. A Machine Learning-Based Test for Predicting Response to Psoriasis Biologics. Ski. J. Cutan. Med. 2021, 5, 621–638. [Google Scholar] [CrossRef]
- Tomalin, L.E.; Kim, J.; da Rosa, J.C.; Lee, J.; Fitz, L.J.; Berstein, G.; Valdez, H.; Wolk, R.; Krueger, J.G.; Suárez-Fariñas, M. Early Quantification of Systemic Inflammatory Proteins Predicts Long-Term Treatment Response to Tofacitinib and Etanercept. J. Investig. Dermatol. 2019, 140, 1026–1034. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Authors/Ref | Samples | Findings | Year |
|---|---|---|---|
| Oestreicher [15] | 24 psoriatic skin biopsies –lesional and uninvolved | 159 DEGs were generated including S100A7, S100A12, elafin, KRT16, KRT17, MMP12, FARP5 | 2001 |
| Nattkemper [34] | 25 patients with atopic dermatitis and 25 patients with psoriasis | 18,000 DEGs common between itchy, lesional atopic, and psoriatic skin identified, outofwhich 2000 genes were unique to both AD and psoriasis including CCL-2/3/17/18, IL-6/8/17A/22/23A, S100A9/A15, TRPV1, PLA2 | 2007 |
| Yao [47] | 26 paired nonlesional and lesional (all were plaque-type) skin biopsies from 26 psoriatic patients | Type 1 IFNS were significantly elevated in psoriatic lesions suggesting their active signaling in psoriasis | 2008 |
| Ainali [43] | 37 patients affected by chronic plaque psoriasis. | A comprehensive analysis of gene expression in paired lesional and non-lesional psoriatic tissue samples revealed different molecular subgroups associated with Wnt, Notch, TGF-beta, ErbB signaling pathways. | 2012 |
| Ewald [54] | Four microarray datasets including 54 LS and 43 NL samples | Differentially expressed in AD several genes involved in lipid metabolism including FA2H, critical in maintaining the permeability barrier of epidermis and ELOVL3, encoding a protein involved in the elongation of long chain fatty acids and essential in prevention of trans-epidermal water loss. | 2015 |
| Gudjonsson [41] | 58 psoriatic subjects | Uninvolved psoriatic skin exists in a a “prepsoriatic” gene expression signature and downregulation of PPARA, ESR2 and SREBBF1 suggesting decreased lipid biosynthesis and increased innate immunity in uninvolved psoriatic skin. | 2009 |
| Gudjonsson [37] | 58 psoriatic subjects | Identified over 600 new transcripts SERPINB4, PI3, DEFB4 and several S100 family members. Comparison of the psoriatic transcriptome to the transcriptomes of cytokine stimulated cultured keratinocytes revealed little overlap with the lesional psoriatic dysregulated transcriptome. | 2010 |
| Swindell [50] | 62 lesional skin samples obtained from patients with stable chronic plaque psoriasis. | Variability in cytokine signature was identified by whole genome transcriptional profiling | 2012 |
| Suez Farinas [46] | Skin biopsies from 85 paired lesional and non-lesional samples from a cohort of patients with moderate-to-severe psoriasis vulgaris who were not receiving active psoriasis therapy | Identified 2725 genes as being differentially expressed in psoriasis and link to functional pathways associated with metabolic diseases/diabetes and to cardiovascular risk pathways | 2012 |
| Li [31] | 92 psoriatic skin biopsies | RNA-seq analysis identified differentially expressed transcripts enriched for lymphoid and/or myeloid signature transcripts and genes induced by IL-17 in keratinocytes. | 2014 |
| Swindell [51] | 163 biopsies from psoriatic lesions | Identified 1233 psoriasis-increased DEGs attributing to keratinocyte activity, infiltration of lesions by T-cells, and macrophages (11%). | 2013 |
| Tian [53] | 5 microarray data sets, including 193 LS and NL pairs | Several new genes were identified that are involved in cardiovascular development and lipid metabolism. highlighting the relationship between psoriasis and systemic manifestations such as the metabolic syndrome and cardiovascular disease | 2012 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krishnan, V.S.; Kõks, S. Transcriptional Basis of Psoriasis from Large Scale Gene Expression Studies: The Importance of Moving towards a Precision Medicine Approach. Int. J. Mol. Sci. 2022, 23, 6130. https://doi.org/10.3390/ijms23116130
Krishnan VS, Kõks S. Transcriptional Basis of Psoriasis from Large Scale Gene Expression Studies: The Importance of Moving towards a Precision Medicine Approach. International Journal of Molecular Sciences. 2022; 23(11):6130. https://doi.org/10.3390/ijms23116130
Chicago/Turabian StyleKrishnan, Vidya S., and Sulev Kõks. 2022. "Transcriptional Basis of Psoriasis from Large Scale Gene Expression Studies: The Importance of Moving towards a Precision Medicine Approach" International Journal of Molecular Sciences 23, no. 11: 6130. https://doi.org/10.3390/ijms23116130