Novel SFRP2 DNA Methylation Profile Following Neoadjuvant Therapy in Colorectal Cancer Patients with Different Grades of BMI

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Biochemical Determination
2.3. Human Colorectal CarcinomaCell Lines and SFRP2 Expression Assay by RT-qPCR
2.4. DNA Extraction, Bisulfite Treatment, and Pyrosequencing
2.5. Statistical Analysis
3. Results
3.1. Clinical, Anthropometrical, and Biochemical Characteristics of the CRC Patients
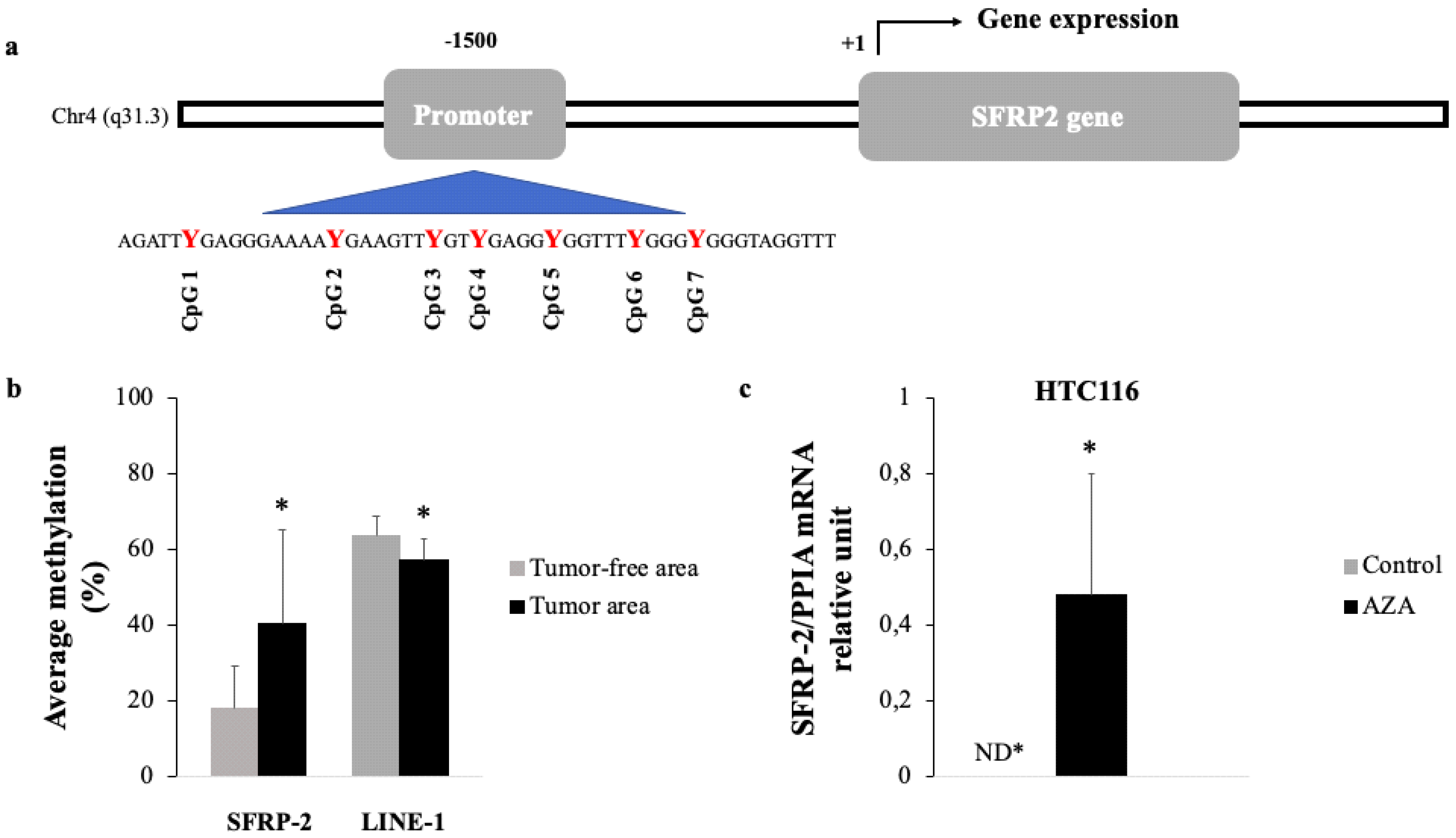
3.2. Methylation Status of SFRP2 in CRC Tissue
3.3. Baseline Characteristics of Colorectal Cancer Patients with Regards to their BMI Category
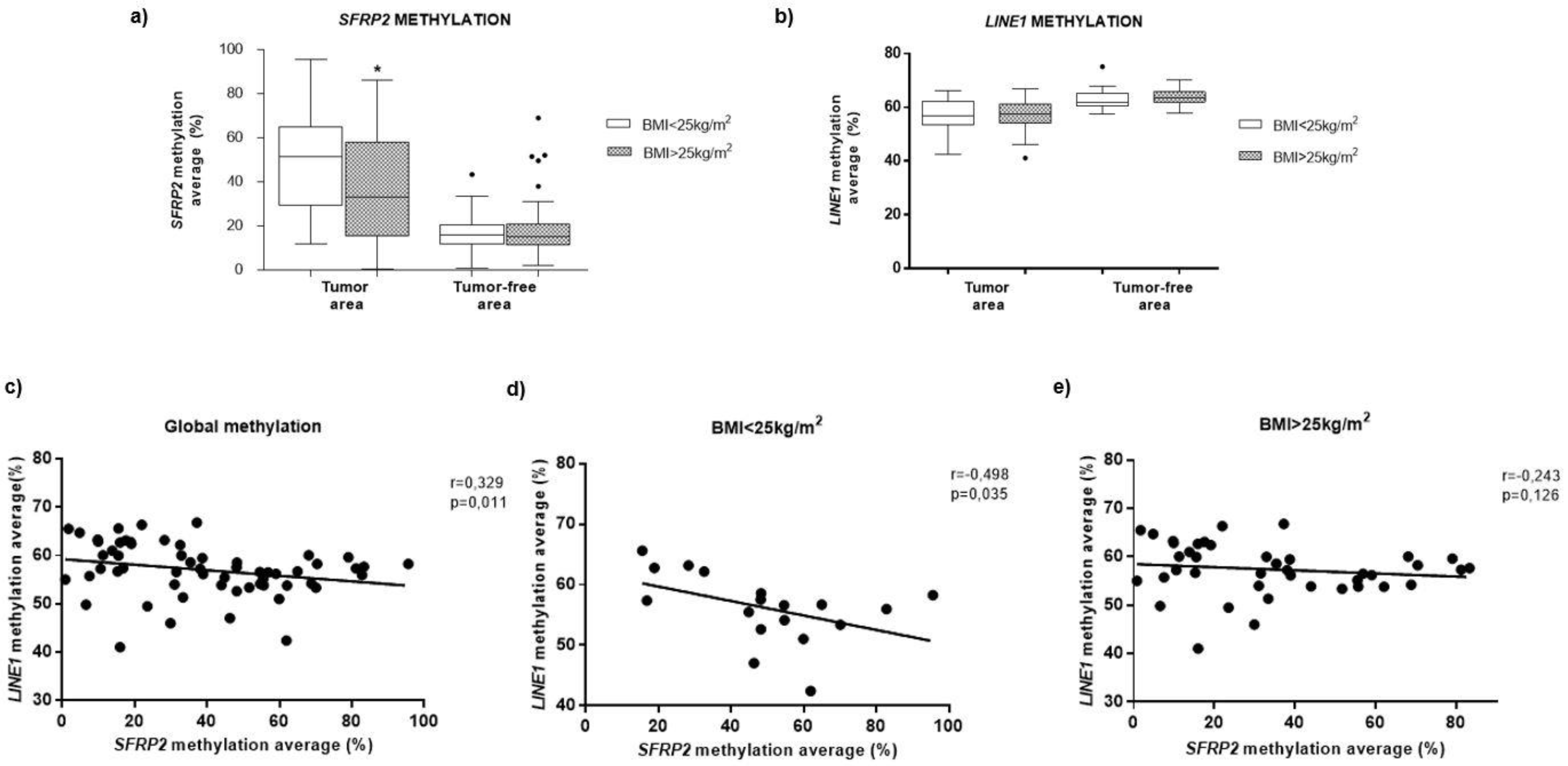
3.4. Methylation of SFRP2 in CRC Tissue is Associated with BMI
3.5. SFRP2Methylation in CRC Patients with and without Neoadjuvant Treatment
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Availability of Data and Materials
Abbreviations
| CRC | colorectal cancer |
| SFRP2 | secreted frizzled-related protein 2 |
| LINE-1 | long Interspersed Element-1 |
| BMI | body mass index |
| HDL-c | high density lipoprotein cholesterol |
| LDL-c | low density lipoprotein cholesterol |
| FFPE | formalin-fixed paraffin-embedded |
References
- Siegel, R.L.; Miller, K.D.; Jemal, A.; Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2016. CA Cancer J. Clin. 2016, 66, 7–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Serra, P.; Esteller, M. {DNA} methylation-associated silencing of tumor-suppressor {microRNAs} in cancer. Oncogene 2012, 31, 1609–1622. [Google Scholar] [CrossRef] [PubMed]
- Chimonidou, M.; Strati, A.; Tzitzira, A.; Sotiropoulou, G.; Malamos, N.; Georgoulias, V.; Lianidou, E.S. DNA methylation of tumor suppressor and metastasis suppressor genes in circulating tumor cells. Clin. Chem. 2011, 57, 1169–1177. [Google Scholar] [CrossRef]
- Kondo, Y.; Issa, J.P.J. Epigenetic changes in colorectal cancer. Cancer Metastasis Rev. 2004, 23, 29–39. [Google Scholar] [CrossRef]
- Sakai, E.; Nakajima, A.; Kaneda, A. Accumulation of aberrant DNA methylation during colorectal cancer development. World J. Gastroenterol. 2014, 20, 978–987. [Google Scholar] [CrossRef]
- Kumar, A.; Gosipatala, S.B.; Pandey, A.; Singh, P. Prognostic Relevance of Sfrp1 Gene Promoter Methylation in Colorectal Carcinoma. Asian Pac. J. Cancer Prev. 2019, 20, 1571–1577. [Google Scholar] [CrossRef]
- Tokarz, P.; Pawlowska, E.; Bialkowska-Warzecha, J.; Blasiak, J. The significance of DNA methylation profile in metastasis-related genes for the progression of colorectal cancer. Cell. Mol. Biol. 2017, 63, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Bishnupuri, K.S.; Mishra, M.K. Epigenetics of colorectal cancer. In Epigenetic Advancements in Cancer; Springer International Publishing: Cham, Switzerland, 2016; ISBN 9783319249513. [Google Scholar]
- Portela, A.E.M. Epigenetic modifications and human disease. Nat. Biotechnol. 2010, 10, 1057–1068. [Google Scholar] [CrossRef]
- Baba, Y.; Huttenhower, C.; Nosho, K.; Tanaka, N.; Shima, K.; Hazra, A.; Schernhammer, E.S.; Hunter, D.J.; Giovannucci, E.L.; Fuchs, C.S.; et al. Epigenomic diversity of colorectal cancer indicated by LINE-1 methylation in a database of 869 tumors. Mol. Cancer 2010, 9, 125. [Google Scholar] [CrossRef]
- Sui, C.; Ma, J.; Chen, Q.; Yang, Y. The variation trends of {SFRP}2 methylation of tissue, feces, and blood detection in colorectal cancer development. Eur. J. Cancer Prev. 2016, 25, 288–298. [Google Scholar] [CrossRef]
- Hu, H.; Wang, T.; Pan, R.; Yang, Y.; Li, B.; Zhou, C.; Zhao, J.; Huang, Y.; Duan, S. Hypermethylated Promoters of Secreted Frizzled-Related Protein Genes are Associated with Colorectal Cancer. Pathol. Oncol. Res. 2019, 25, 567–575. [Google Scholar] [CrossRef]
- Bagci, B.; Sari, M.; Karadayi, K.; Turan, M.; Ozdemir, O.; Bagci, G. KRAS, BRAF oncogene mutations and tissue specific promoter hypermethylation of tumor suppressor SFRP2, DAPK1, MGMT, HIC1 and p16 genes in colorectal cancer patients. Cancer Biomark. 2016, 17, 133–143. [Google Scholar] [CrossRef]
- Crujeiras, A.B.; Morcillo, S.; Diaz-Lagares, A.; Sandoval, J.; Castellano-Castillo, D.; Torres, E.; Hervas, D.; Moran, S.; Esteller, M.; Macias-Gonzalez, M.; et al. Identification of an episignature of human colorectal cancer associated with obesity by genome-wide DNA methylation analysis. Int. J. Obes. 2018, 43, 176–188. [Google Scholar] [CrossRef]
- Li, H.; Wang, Z.; Zhao, G.; Ma, Y.; Chen, Y.; Xue, Q.; Zheng, M.; Fei, S. Performance of a MethyLight assay for methylated SFRP2 DNA detection in colorectal cancer tissue and serum. Int. J. Biol. Markers 2019, 34, 54–59. [Google Scholar] [CrossRef]
- Yang, Q.; Huang, T.; Ye, G.; Wang, B.; Zhang, X. Methylation of SFRP2 gene as a promising noninvasive biomarker using feces in colorectal cancer diagnosis: A systematic meta-analysis. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef]
- Bardou, M.; Barkun, A.N.; Martel, M. Obesity and colorectal cancer. Gut 2013, 62, 933–947. [Google Scholar] [CrossRef]
- Wahl, S.; Drong, A.; Lehne, B.; Loh, M.; Scott, W.R.; Kunze, S.; Tsai, P.C.; Ried, J.S.; Zhang, W.; Yang, Y.; et al. Epigenome-wide association study of body mass index, and the adverse outcomes of adiposity. Nature 2017, 541, 81–86. [Google Scholar] [CrossRef]
- Dayde, D.; Tanaka, I.; Jain, R.; Tai, M.C.; Taguchi, A. Predictive and Prognostic Molecular Biomarkers for Response to Neoadjuvant Chemoradiation in Rectal Cancer. Int. J. Mol. Sci. 2017, 18, 573. [Google Scholar] [CrossRef]
- Sun, Y.; Xu, Z.; Lin, H.; Lu, X.; Huang, Y.; Huang, S.; Wang, X.; Chi, P. Impact of body mass index on treatment outcome of neoadjuvant chemoradiotherapy in locally advanced rectal cancer. Eur. J. Surg. Oncol. 2017, 43, 1828–1834. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, Y.; Tan, L.; Fu, X. The pivotal role of DNA methylation in the radio-sensitivity of tumor radiotherapy. Cancer Med. 2018, 7, 3812–3819. [Google Scholar] [CrossRef]
- Sood, S.; Patel, F.D.; Srinivasan, R.; Dhaliwal, L.K. Chemoradiation therapy induces in vivo changes in gene promoter methylation & gene transcript expression in patients with invasive cervical cancer. Indian J. Med. Res. 2018, 147, 151–157. [Google Scholar]
- Stone, L. Radiotherapy induces epigenetic changes. Nat. Rev. Urol. 2016, 13, 241. [Google Scholar] [CrossRef]
- Bosnan, F.T.; Carneiro, F.; Hruban, R.H.; Theise, N.D. WHO Classification of Tumors of the Digestive System, 4th ed.; International Agency for Research on Cancer (IARC): Lyon, France, 2010; ISBN 978-92-832-2432-7. [Google Scholar]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef]
- Castellano-Castillo, D.; Morcillo, S.; Crujeiras, A.B.; Sánchez-Alcoholado, L.; Clemente-Postigo, M.; Torres, E.; Tinahones, F.J.; Macias-Gonzalez, M. Association between serum 25-hydroxyvitamin D and global DNA methylation in visceral adipose tissue from colorectal cancer patients. BMC Cancer 2019. [Google Scholar] [CrossRef]
- Castellano-Castillo, D.; Morcillo, S.; Clemente-Postigo, M.; Crujeiras, A.B.; Fernandez-García, J.C.; Torres, E.; Tinahones, F.J.; Macias-Gonzalez, M. Adipose tissue inflammation and VDR expression and methylation in colorectal cancer. Clin. Epigenet. 2018. [Google Scholar] [CrossRef]
- Ashktorab, H.; Brim, H. DNA Methylation and Colorectal Cancer. Curr. Colorectal Cancer Rep. 2014, 10, 425–430. [Google Scholar] [CrossRef] [Green Version]
- Leygo, C.; Williams, M.; Jin, H.C.; Chan, M.W.Y.; Chu, W.K.; Grusch, M.; Cheng, Y.Y. DNA Methylation as a Noninvasive Epigenetic Biomarker for the Detection of Cancer. Dis. Markers 2017. [Google Scholar] [CrossRef]
- Bovolenta, P.; Esteve, P.; Ruiz, J.M.; Cisneros, E.; Lopez-Rios, J. Beyond Wnt inhibition: New functions of secreted Frizzled-related proteins in development and disease. J. Cell Sci. 2008, 121, 737–746. [Google Scholar] [CrossRef]
- Morikawa, T.; Kuchiba, A.; Yamauchi, M.; Meyerhardt, J.A.; Shima, K.; Nosho, K.; Chan, A.T.; Giovannucci, E.; Fuchs, C.S.; Ogino, S. Association of CTNNB1 (β-Catenin) alterations, body mass index, and physical activity with survival in patients with colorectal cancer. J. Am. Med. Assoc. 2011, 305, 1685–1694. [Google Scholar] [CrossRef]
- Bai, Y.; Sun, Q. Macrophage recruitment in obese adipose tissue. Obes. Rev. 2015, 16, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Chen, N.; Wang, J. Wnt/β-Catenin Signaling and Obesity. Front. Physiol. 2018, 9, 792. [Google Scholar] [CrossRef]
- Gomez-Millan, J.; Queipo, M.I.; Delgado, M.M.; Perez-Villa, L.; Roman, A.; Portilla, D.F.; Torres, E.; Luque, D.V.; Bayo, E.; Medina, J.A. The impact of body mass index and nuclear β-catenin on survival in locally advanced rectal cancer treated with preoperative radiochemotherapy. J. Surg. Oncol. 2017, 115, 301–306. [Google Scholar] [CrossRef]
- Veeck, J.; Noetzel, E.; Bektas, N.; Jost, E.; Hartmann, A.; Knüchel, R.; Dahl, E. Promoter hypermethylation of the SFRP2 gene is a high-frequent alteration and tumor-specific epigenetic marker in human breast cancer. Mol. Cancer 2008. [Google Scholar] [CrossRef]
- Tang, D.; Liu, J.; Wang, D.; Yu, H.; Li, Y.; Zhang, J. Diagnostic and prognostic value of the methylation status of secreted frizzled-related protein 2 in colorectal cancer. Clin. Investig. Med. 2011, 34, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Herman, J.G. Hypermethylation of tumor suppressor genes in cancer. Semin. Cancer Biol. 1999, 9, 359–367. [Google Scholar] [CrossRef]
- Park, J.; Morley, T.S.; Kim, M.; Clegg, D.J.; Scherer, P.E. Obesity and cancer—Mechanisms underlying tumour progression and recurrence. Nat. Rev. Endocrinol. 2014, 10, 455–465. [Google Scholar] [CrossRef]
- Swets, M.; Zaalberg, A.; Boot, A.; Wezel, V.T.; Frouws, M.A.; Bastiaannet, E.; Gelderblom, H.; Velde, C.J.H.; Kuppen, P.J.K. Tumor LINE-1 Methylation Level in Association with Survival of Patients with Stage II Colon Cancer. Int. J. Mol. Sci. 2016, 18, 36. [Google Scholar] [CrossRef]
- Moretto, R.; Cremolini, C.; Rossini, D.; Pietrantonio, F.; Battaglin, F.; Mennitto, A.; Bergamo, F.; Loupakis, F.; Marmorino, F.; Berenato, R.; et al. Location of Primary Tumor and Benefit From Anti-Epidermal Growth Factor Receptor Monoclonal Antibodies in Patients With RAS and BRAF Wild-Type Metastatic Colorectal Cancer. Oncologist. 2016, 21, 988–994. [Google Scholar] [CrossRef]
- Berger, N.A.; Scacheri, P.C. Targeting Epigenetics to Prevent Obesity Promoted Cancers. Cancer Prev. Res. (Phila.) 2018, 11, 125–128. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.; Ma, L.; Ma, G.H.; Ren, H. Genome-wide Analysis Reveals DNA Methylation Alterations in Obesity Associated with High Risk of Colorectal Cancer. Sci. Rep. 2019, 9, 5100. [Google Scholar] [CrossRef] [PubMed]
- Bultman, S.J. A Reversible Epigenetic Link between Obesity and Cancer Risk. Trends Endocrinol. Metab. 2018, 29, 529–531. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Variables | BMI < 25 kg/m2 (n = 22) | BMI > 25 kg/m2 (n = 53) | p-Value |
|---|---|---|---|
| Age (years) | 62.91 ± 11.21 | 66.64 ± 7.91 | NS |
| Male/female | 14/8 | 38/15 | NS |
| BMI (kg/m2) | 23.33 ± 1.32 | 29.20 ± 3.43 | p < 0.05 |
| Waist circumference (cm) | 89.80 ± 12.01 | 101.39 ± 12.93 | NS |
| Glucose (mg/dL) | 122.57 ± 48.51 | 120.00 ± 40.40 | NS |
| Insulin (µUI/mL) | 4.07 ± 2.30 | 7.16 ± 5.63 | p < 0.05 |
| HOMA-IR | 1.42 ± 1.38 | 2.25 ± 2.17 | NS |
| Diabetes Mellitus (%) | 30 | 29.2 | NS |
| Triglycerides (mg/dL) | 146.71 ± 76.36 | 171.92 ± 82.52 | NS |
| Total cholesterol (mg/dL) | 180.76 ± 39.90 | 169.13 ± 36.60 | NS |
| HDL-c (mg/dL) | 42.86 ± 15.07 | 39.51 ± 13.57 | NS |
| LDL-c (mg/dL) | 109.99 ± 30.94 | 100.72 ± 30.88 | NS |
| Corrected calcium (mg/dL) | 9.09±0.32 | 8.94 ± 0.54 | NS |
| Alkaline phosphatase (U/L) | 63.09 ± 26.76 | 72.57 ± 30.65 | NS |
| IGF-1 (ng/mL) | 137.26 ± 48.41 | 128.75 ± 73.92 | NS |
| Variables | BMI < 25 kg/m2 (n = 22) | BMI > 25 kg/m2 (n = 53) | p-Value |
|---|---|---|---|
| Location | 0.37 | ||
| Proximal (Cecum, Ascending, Transverse) | 2 | 12 | |
| Distal (Descending, Sigmoid) | 5 | 9 | |
| Rectum | 15 | 32 | |
| Stage | 0.43 | ||
| I | 6 | 13 | |
| II | 8 | 12 | |
| III | 7 | 20 | |
| IV | 1 | 8 | |
| Treatments (previous surgery) | |||
| Radiotherapy + Chemotherapy | 4 | 13 | 0.25 |
| Radiotherapy | 2 | 1 | |
| Chemotherapy | 0 | 4 | |
| Non-treatment | 16 | 35 |
| R2 = 0.280 | 95% IC | |||
|---|---|---|---|---|
| Variables | Standardized β | p Value | Lower Bound | Upper Bound |
| BMI (BMI < 25 vs. BMI > 25) | −0.339 | 0.022 | −32.166 | −2.700 |
| Neoadjuvant therapy (treated vs. non treated) | −0.419 | <0.001 | −33.657 | −5.860 |
| Diabetes mellitus (diabetic vs. no diabetic) | −0.297 | 0.064 | −34.535 | 1.013 |
| Age (years) | 0.188 | 0.218 | −0.284 | 1.201 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabrera-Mulero, A.; Crujeiras, A.B.; Izquierdo, A.G.; Torres, E.; Ayers, D.; Casanueva, F.F.; Tinahones, F.J.; Morcillo, S.; Macias-Gonzalez, M. Novel SFRP2 DNA Methylation Profile Following Neoadjuvant Therapy in Colorectal Cancer Patients with Different Grades of BMI. J. Clin. Med. 2019, 8, 1041. https://doi.org/10.3390/jcm8071041
Cabrera-Mulero A, Crujeiras AB, Izquierdo AG, Torres E, Ayers D, Casanueva FF, Tinahones FJ, Morcillo S, Macias-Gonzalez M. Novel SFRP2 DNA Methylation Profile Following Neoadjuvant Therapy in Colorectal Cancer Patients with Different Grades of BMI. Journal of Clinical Medicine. 2019; 8(7):1041. https://doi.org/10.3390/jcm8071041
Chicago/Turabian StyleCabrera-Mulero, Amanda, Ana B. Crujeiras, Andrea G. Izquierdo, Esperanza Torres, Duncan Ayers, Felipe F. Casanueva, Francisco J. Tinahones, Sonsoles Morcillo, and Manuel Macias-Gonzalez. 2019. "Novel SFRP2 DNA Methylation Profile Following Neoadjuvant Therapy in Colorectal Cancer Patients with Different Grades of BMI" Journal of Clinical Medicine 8, no. 7: 1041. https://doi.org/10.3390/jcm8071041