Towards Understanding Therapeutic Failures in Masquelet Surgery: First Evidence that Defective Induced Membrane Properties are Associated with Clinical Failures

 ,
,
Abstract
:1. Introduction
2. Experimental Section
2.1. Study Design
2.2. Histology and Immunohistochemical Analysis
2.3. Isolation and Characterization of Mesenchymal Stromal Cells (MSC) from IM Fragments
2.4. Real-Time PCR Analysis
2.5. Statistics
3. Results
3.1. Demographic and Clinical Features of Patients
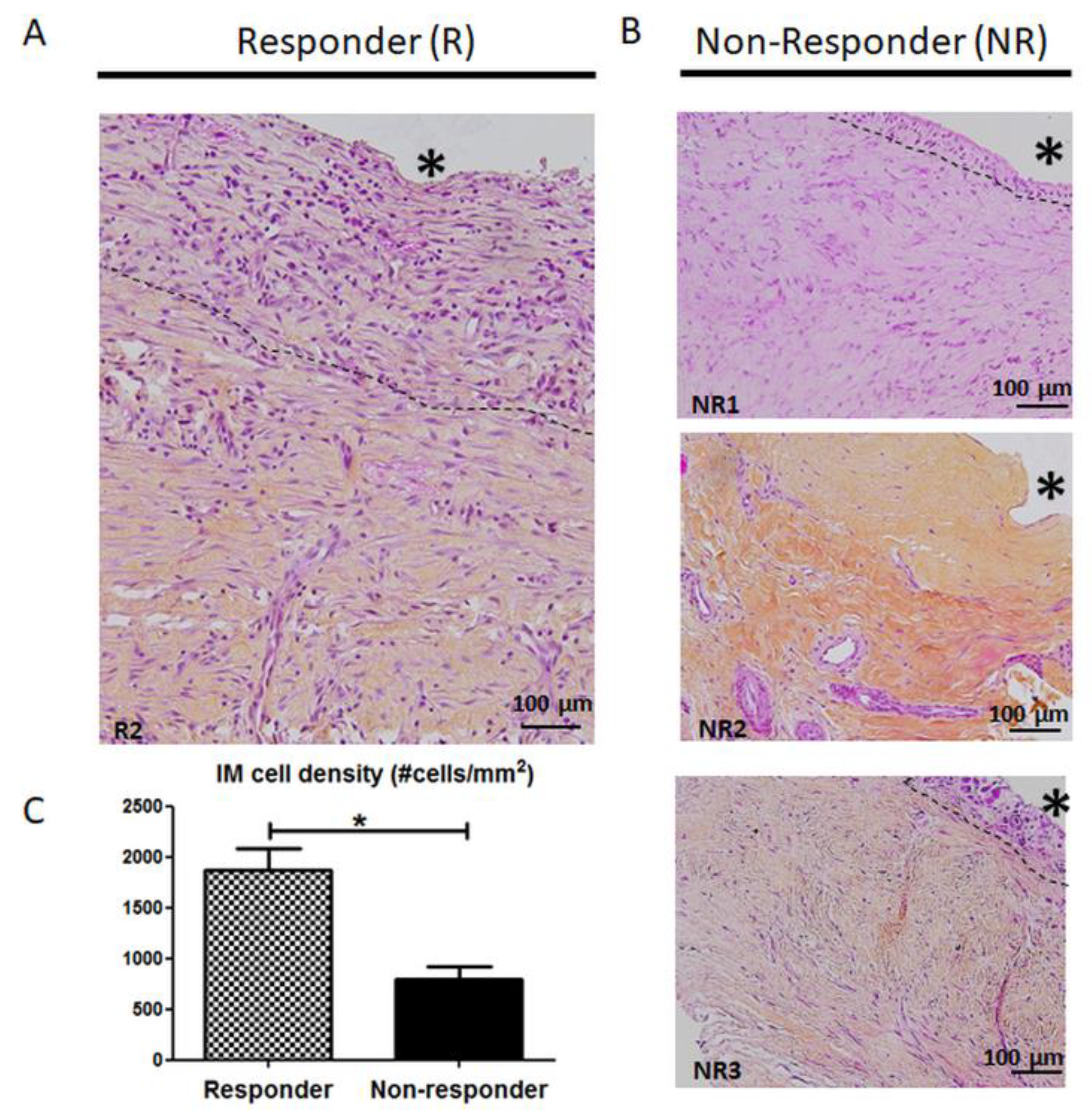
3.2. Histology and Cellularity are Disrupted in Non-Responder Induced Membranes (IM)
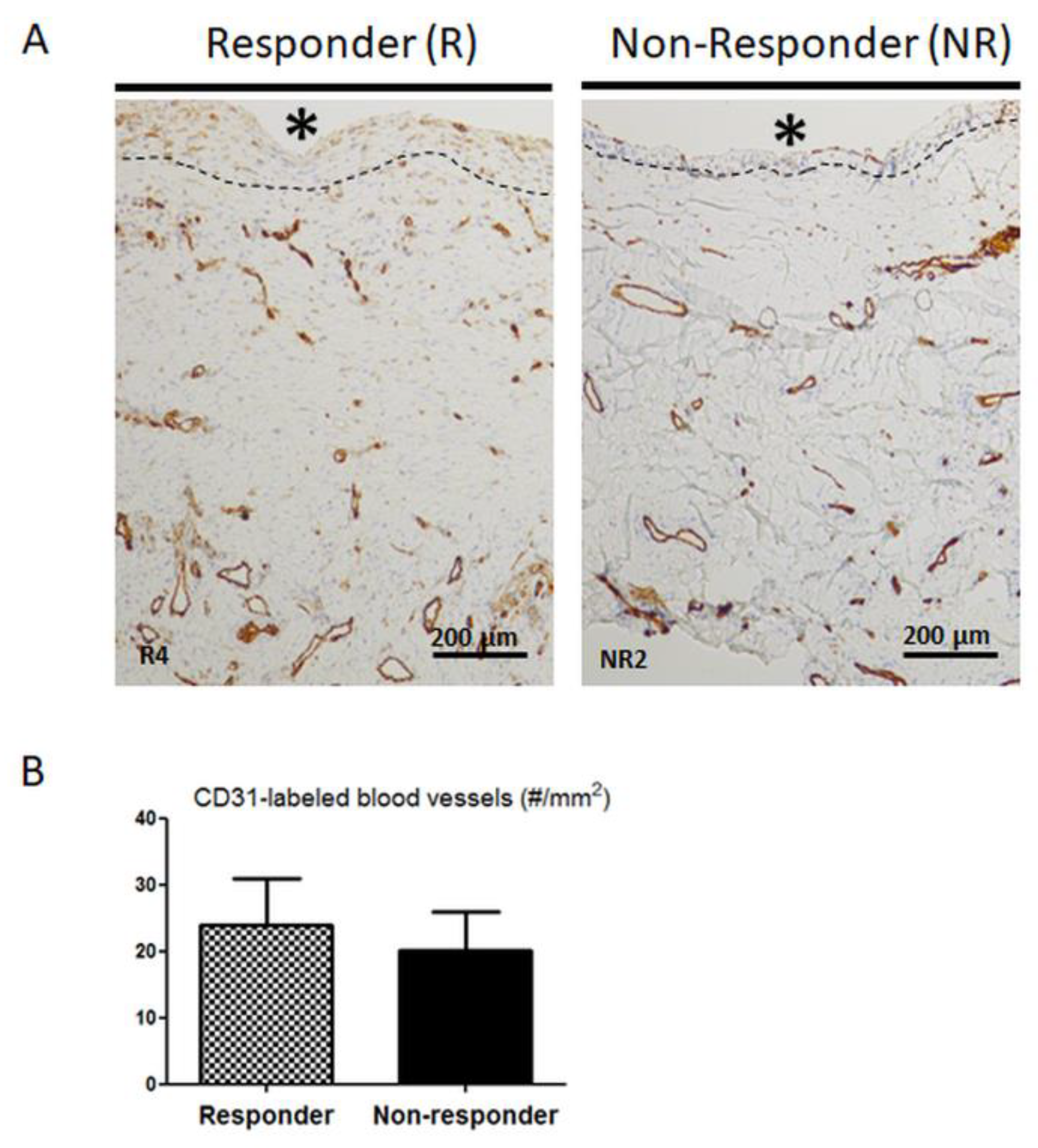
3.3. Distribution and Number of CD31-Positive Blood Vessels are not Altered in Non-Responder Induced Membranes
3.4. Mesenchymal Stromal Cell Content is Altered in Non-Responder Membranes
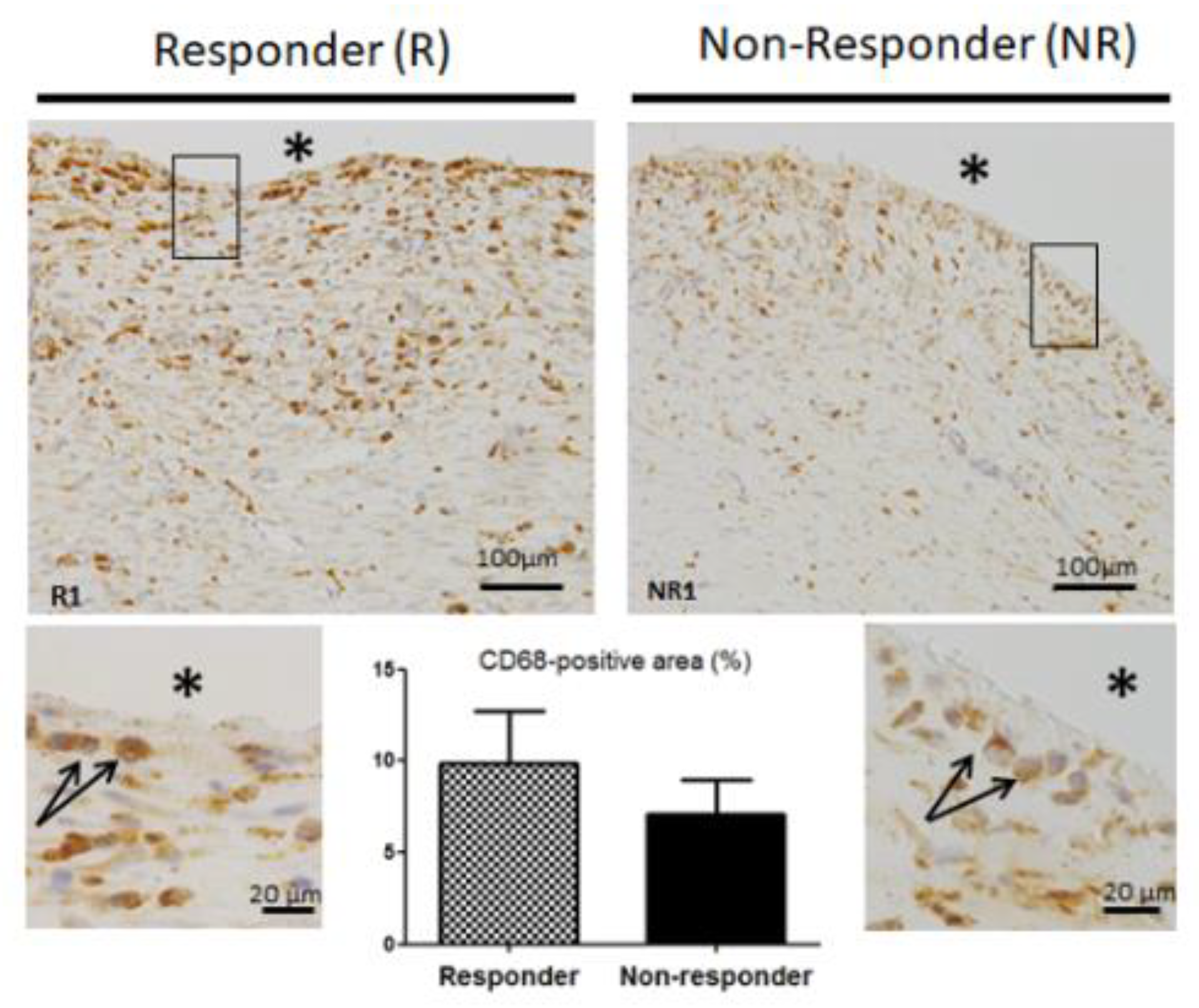
3.5. CD68 Pan Macrophage Marker Localization and Gene Expression are Similar in both Responder and Non-Responder Induced Membranes
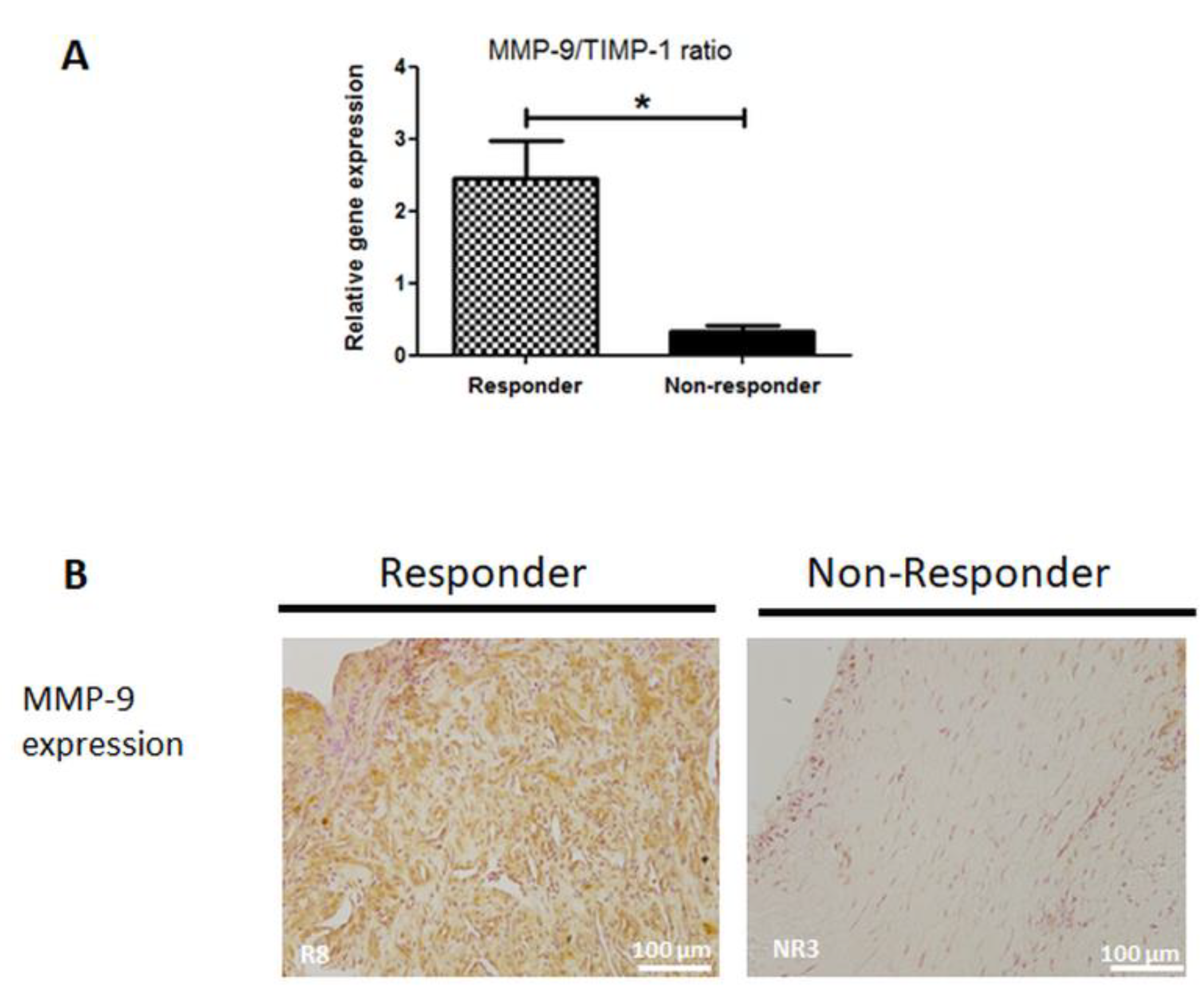
3.6. MMP-9 Expression is Disrupted in Non-Responder Induced Membranes
3.7. Collagen Matrix Organization is Disrupted in Non-Responder Induced Membranes
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
| Gene | Gene Symbol | Accession Number | 5’3’-Primer Sequence | Product Size (bp) | Annealing Temperature (°C) | Primers Concentrations (µM) | |
| REFERENCE GENES | ribosomal protein, large, P0 | rplp0 3’ set | NM_001002 | F GGCGACCTGGAAGTCCAACTAC | 113 | 54 | 0.5 |
| R CGGATCTGCTGCATCTGCTTG | |||||||
| rplp0 5’ set | NM_001002 | F GCATCTACAACCCTGAAGTGCTTG | 93 | 57 | 0.5 | ||
| R GCAGACAGACACTGGCAACATTG | |||||||
| peptidylpropyl isomerase A | ppia | XM_193409 | F TATCTGCACTGCCAAGACTGAGTG | 126 | 57 | 0.5 | |
| R CTTCTTGCTGGTCTTGCCATTCC | |||||||
| tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein zeta | ywhaz | NM_145690 | F CAGCACGCTAATAATGCAATTACTG | 129 | 54 | 0.5 | |
| R AATGAGGCAGACAAAAGTTGGAAG | |||||||
| TARGET GENES | monocyte chemoattractant protein 1 | mcp1 | NM_002982 | F AGAAGAATCACCAGCAGCAAGTG | 86 | 55 | 0.5 |
| R TGCTTGGGGTCAGCACAGAT | |||||||
| transforming growth factor beta 2 | tgfβ2 | NM_001135599.3 | F GCCCGTATTTATGGAGTTCAGACAC | 93 | 58 | 0.5 | |
| R CGCAGCAAGGAGAAGCAGATG | |||||||
| metalloproteinase inhibitor 1 | timp | NM_003254.2 | F GGGGCTTCACCAAGACCTACAC | 108 | 58 | 0.5 | |
| R GGTCCGTCCACAAGCAATGAG | |||||||
| runt related transcription factor | runx2 | NM_001024630.3 | F GCCTTCCACTCTCAGTAAGA | 122 | 57 | 0.4 | |
| R GCATTCGTGGGTTGGAGAA | |||||||
| CD68 molecule | cd68 | NM_001251.2 | F GGTTGTCTACCTGAGCTACA | 74 | 56 | 0.4 | |
| R CCGAGAATGTCCACTGTG | |||||||
| von Willebrand factor | vwf | NM_000552.4 | F CAAGGTGGGAAGCTGTAAG | 105 | 53 | 0.5 | |
| R TCCTGCACATCGTTGATG | |||||||
| matrix metalloproteinase-9 | mmp9 | NM_004994.2 | F GGGAAGATGCTGCTGTTCA | 127 | 59 | 0.5 |
References
- Wiese, A.; Pape, H.C. Bone Defects Caused by High-energy Injuries, Bone Loss, Infected Nonunions, and Nonunions. Orthop. Clin. North Am. 2010, 41, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Chimutengwende-Gordon, M.; Mbogo, A.; Khan, W.; Wilkes, R. Limb reconstruction after traumatic bone loss. Injury 2017, 48, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Tong, K.; Zhong, Z.; Peng, Y.; Lin, C.; Cao, S.; Yang, Y.; Wang, G. Masquelet technique versus Ilizarov bone transport for reconstruction of lower extremity bone defects following posttraumatic osteomyelitis. Injury 2017, 48, 1616–1622. [Google Scholar] [CrossRef] [PubMed]
- Masquelet, A.C.; Begue, T. The concept of induced membrane for reconstruction of long bone defects. Orthop. Clin. North Am. 2010, 41, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Karger, C.; Kishi, T.; Schneider, L.; Fitoussi, F.; Masquelet, A.-C. Treatment of posttraumatic bone defects by the induced membrane technique. Orthop. Traumatol. Surg. Res. 2012, 98, 97–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelissier, P.; Masquelet, A.C.; Bareille, R.; Pelissier, S.M.; Amedee, J. Induced membranes secrete growth factors including vascular and osteoinductive factors and could stimulate bone regeneration. J. Orthop. Res. 2004, 22, 73–79. [Google Scholar] [CrossRef]
- Christou, C.; Oliver, R.A.; Yu, Y.; Walsh, W.R. The Masquelet technique for membrane induction and the healing of ovine critical sized segmental defects. PLoS ONE 2014, 9, e114122. [Google Scholar] [CrossRef] [Green Version]
- Zwetyenga, N.; Catros, S.; Emparanza, A.; Deminiere, C.; Siberchicot, F.; Fricain, J.-C. Mandibular reconstruction using induced membranes with autologous cancellous bone graft and HA-betaTCP: Animal model study and preliminary results in patients. Int. J. Oral Maxillofac. Surg. 2009, 38, 1289–1297. [Google Scholar] [CrossRef]
- Wang, X.; Wei, F.; Luo, F.; Huang, K.; Xie, Z. Induction of granulation tissue for the secretion of growth factors and the promotion of bone defect repair. J. Orthop. Surg. Res. 2015, 10, 147. [Google Scholar] [CrossRef] [Green Version]
- Cuthbert, R.J.; Churchman, S.M.; Tan, H.B.; McGonagle, D.; Jones, E.; Giannoudis, P.V. Induced periosteum a complex cellular scaffold for the treatment of large bone defects. Bone 2013, 57, 484–492. [Google Scholar] [CrossRef]
- Tang, Q.; Tong, M.; Zheng, G.; Shen, L.; Shang, P.; Liu, H. Masquelet’s induced membrane promotes the osteogenic differentiation of bone marrow mesenchymal stem cells by activating the Smad and MAPK pathways. Am. J. Transl. Res. 2018, 10, 1211–1219. [Google Scholar] [PubMed]
- Morelli, I.; Drago, L.; George, D.A.; Gallazzi, E.; Scarponi, S.; Romanò, C.L. Masquelet technique: Myth or reality? A systematic review and meta-analysis. Injury 2016, 47 (Suppl. 6), S68–S76. [Google Scholar] [CrossRef]
- Giannoudis, P.V.; Harwood, P.J.; Tosounidis, T.; Kanakaris, N.K. Restoration of long bone defects treated with the induced membrane technique: Protocol and outcomes. Injury 2016, 47 (Suppl. 6), S53–S61. [Google Scholar] [CrossRef]
- El-Alfy, B.S.; Ali, A.M. Management of segmental skeletal defects by the induced membrane technique. Indian J. Orthop. 2015, 49, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Giotikas, D.; Tarazi, N.; Spalding, L.; Nabergoj, M.; Krkovic, M. Results of the Induced Membrane Technique in the Management of Traumatic Bone Loss in the Lower Limb: A Cohort Study. J. Orthop. Trauma 2019, 33, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Siboni, R.; Joseph, E.; Blasco, L.; Barbe, C.; Bajolet, O.; Diallo, S.; Ohl, X. Management of septic non-union of the tibia by the induced membrane technique. What factors could improve results? Orthop. Traumatol. Surg. Res. 2018, 104, 911–915. [Google Scholar] [CrossRef] [PubMed]
- Pugniere, P.; Banzet, S.; Chaillou, T.; Mouret, C.; Peinnequin, A. Pitfalls of reverse transcription quantitative polymerase chain reaction standardization: Volume-related inhibitors of reverse transcription. Anal. Biochem. 2011, 415, 151–157. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034-1. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J. A power primer. Psychol. Bull. 1992, 112, 155–159. [Google Scholar] [CrossRef]
- Tarchala, M.; Harvey, E.J.; Barralet, J. Biomaterial-Stabilized Soft Tissue Healing for Healing of Critical-Sized Bone Defects: The Masquelet Technique. Adv. Healthc. Mater. 2016, 5, 630–640. [Google Scholar] [CrossRef] [PubMed]
- Aho, O.-M.; Lehenkari, P.; Ristiniemi, J.; Lehtonen, S.; Risteli, J.; Leskelä, H.-V. The mechanism of action of induced membranes in bone repair. J. Bone Jt. Surg. Am. 2013, 95, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Aurégan, J.-C.; Bégué, T.; Rigoulot, G.; Glorion, C.; Pannier, S. Success rate and risk factors of failure of the induced membrane technique in children: A systematic review. Injury 2016, 47 (Suppl. 6), S62–S67. [Google Scholar] [CrossRef]
- Gruber, H.E.; Ode, G.; Hoelscher, G.; Ingram, J.; Bethea, S.; Bosse, M.J. Osteogenic, stem cell and molecular characterisation of the human induced membrane from extremity bone defects. Bone Jt. Res. 2016, 5, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, Z.; Brooks, P.J.; Barzilay, O.; Fine, N.; Glogauer, M. Macrophages, Foreign Body Giant Cells and Their Response to Implantable Biomaterials. Materials 2015, 8, 5671–5701. [Google Scholar] [CrossRef] [Green Version]
- Azi, M.L.; Teixeira, A.D.; Cotias, R.B.; Joeris, A.; Kfuri, M. Induced-Membrane Technique in the Management of Posttraumatic Bone Defects. JBJS Essent. Surg. Tech. 2019, 9, e22. [Google Scholar] [CrossRef]
- Newman, A.C.; Nakatsu, M.N.; Chou, W.; Gershon, P.D.; Hughes, C.C.W. The requirement for fibroblasts in angiogenesis: Fibroblast-derived matrix proteins are essential for endothelial cell lumen formation. Mol. Biol. Cell 2011, 22, 3791–3800. [Google Scholar] [CrossRef]
- Anderson, J.M.; Rodriguez, A.; Chang, D.T. Foreign body reaction to biomaterials. Semin. Immunol. 2008, 20, 86–100. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Jennings, T.A.; Eaton, J.W. Mast cells mediate acute inflammatory responses to implanted biomaterials. Proc. Natl. Acad. Sci. USA 1998, 95, 8841–8846. [Google Scholar] [CrossRef] [Green Version]
- Kao, W.J.; McNally, A.K.; Hiltner, A.; Anderson, J.M. Role for interleukin-4 in foreign-body giant cell formation on a poly(etherurethane urea) in vivo. J. Biomed. Mater. Res. 1995, 29, 1267–1275. [Google Scholar] [CrossRef]
- Wyatt, L.E.; Sinow, J.D.; Wollman, J.S.; Sami, D.A.; Miller, T.A. The influence of time on human breast capsule histology: Smooth and textured silicone-surfaced implants. Plast. Reconstr. Surg. 1998, 102, 1922–1931. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Wang, W.; Nassiri, S.; Kwan, T.; Dang, C.; Liu, W.; Spiller, K.L. Temporal and spatial distribution of macrophage phenotype markers in the foreign body response to glutaraldehyde-crosslinked gelatin hydrogels. J. Biomater. Sci. Polym. Ed. 2016, 27, 721–742. [Google Scholar] [CrossRef] [PubMed]
- Jham, B.C.; Nikitakis, N.G.; Scheper, M.A.; Papadimitriou, J.C.; Levy, B.A.; Rivera, H. Granulomatous foreign-body reaction involving oral and perioral tissues after injection of biomaterials: A series of 7 cases and review of the literature. J. Oral Maxillofac. Surg. 2009, 67, 280–285. [Google Scholar] [CrossRef]
- Wanat, K.A.; Rosenbach, M.; Zoiber, A.F.; Zhang, P.J.; Schaffer, A. E-cadherin is expressed by mono-and multinucleated histiocytes in cutaneous sarcoidal and foreign body granulomas. Am. J. Derm. 2014, 36, 651–654. [Google Scholar] [CrossRef] [Green Version]
- Nagaoka, I.; Hirota, S. Increased expression of matrix metalloproteinase-9 in neutrophils in glycogen-induced peritoneal inflammation of guinea pigs. Inflamm. Res. 2000, 49, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Colnot, C.; Thompson, Z.; Miclau, T.; Werb, Z.; Helms, J.A. Altered fracture repair in the absence of MMP9. Development 2003, 130, 4123–4133. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Yu, Y.Y.; Lieu, S.; Yang, F.; Lang, J.; Lu, C.; Werb, Z.; Hu, D.; Miclau, T.; Marcucio, R.; et al. MMP9 regulates the cellular response to inflammation after skeletal injury. Bone 2013, 52, 111–119. [Google Scholar] [CrossRef] [Green Version]
- MacLauchlan, S.; Skokos, E.A.; Meznarich, N.; Zhu, D.H.; Raoof, S.; Shipley, J.M.; Senior, R.M.; Bornstein, P.; Kyriakides, T.R. Macrophage fusion, giant cell formation, and the foreign body response require matrix metalloproteinase 9. J. Leukoc. Biol. 2009, 85, 617–626. [Google Scholar] [CrossRef]
- Haubruck, P.; Heller, R.; Apitz, P.; Kammerer, A.; Alamouti, A.; Daniel, V.; Schmidmaier, G.; Moghaddam, A. Evaluation of matrix metalloproteases as early biomarkers for bone regeneration during the applied Masquelet therapy for non-unions. Injury 2018, 49, 1732–1738. [Google Scholar] [CrossRef]
- Arpino, V.; Brock, M.; Gill, S.E. The role of TIMPs in regulation of extracellular matrix proteolysis. Matrix Biol. 2015, 44–46, 247–254. [Google Scholar] [CrossRef]
- Walton, K.L.; Johnson, K.E.; Harrison, C.A. Targeting TGF-β Mediated SMAD Signaling for the Prevention of Fibrosis. Front. Pharm. 2017, 8, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gindraux, F.; Loisel, F.; Bourgeois, M.; Oudina, K.; Melin, M.; de Billy, B.; Sergent, P.; Leclerc, G.; Petite, H.; Auber, F.; et al. Induced membrane maintains its osteogenic properties even when the second stage of Masquelet’s technique is performed later. Eur. J. Trauma Emerg. Surg. 2019, 1–12. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient Identification | Age (yrs) | Gender (F/M) | Smoker (Y/N) | Bone Defect Location | Bone Defect Lenght (cm) | Fracture Fixation | Infection (Y/N) | Duration Spacer in Site (wks) | |
|---|---|---|---|---|---|---|---|---|---|
| Responder (R) | R1 | 30 | M | N | Left tibia | 12 | Ex.Fix | N | 20 |
| R2 | 65 | M | N | Left radius | 7 | Plate | Y | 12 | |
| R3 | 87 | M | N | Left tibia | 5 | Nail | Y | 6 | |
| R4 | 40 | M | N | Left femur | 9 | Nail | N | 8 | |
| R5 | 46 | M | Y | Left tibia | 8 | K-wires | Y | 8 | |
| R6 | 55 | M | N | Left tibia | 4 | Ex.Fix | Y | 10 | |
| R7 | 42 | M | N | Right tibia | 15 | Ex. Fix | Y | 8 | |
| R8 | 40 | M | Y | Right radius | 6 | Plate | Y | 8 | |
| Non responder (NR) | NR1 | 47 | F | N | Left tibia | 10 | Ex. Fix | Y | 20 |
| NR2 | 40 | M | N | Left femur | 9 | Nail | N | 40 | |
| NR3 | 60 | M | Y | Right humerus | 7 | Plate | Y | 9 | |
| Mean +/− SEM | Responder | 50.63 ± 6.40 | 8.25 ± 1.30 | 10.00 ± 1.55 | |||||
| Non-responder | 49.00 ± 5.85 | 8.66 ± 0.88 | 23.00 ± 9.07 | ||||||
| P-value | 0.88 | 0.85 | 0.047 | ||||||
| Patient Identification | Presence or Absence of IM-Derived MSC (CD45- CD90+ CD73+ CD105+ Cells) | |
|---|---|---|
| Responder (R) | R1 | Presence |
| R2 | Not tested | |
| R3 | Presence | |
| R4 | Presence | |
| R5 | Presence | |
| R6 | Presence | |
| R7 | Presence | |
| R8 | Presence | |
| Non-Responder (NR) | NR1 | Not tested |
| NR2 | Absence | |
| NR3 | Absence |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Durand, M.; Barbier, L.; Mathieu, L.; Poyot, T.; Demoures, T.; Souraud, J.-B.; Masquelet, A.-C.; Collombet, J.-M. Towards Understanding Therapeutic Failures in Masquelet Surgery: First Evidence that Defective Induced Membrane Properties are Associated with Clinical Failures. J. Clin. Med. 2020, 9, 450. https://doi.org/10.3390/jcm9020450
Durand M, Barbier L, Mathieu L, Poyot T, Demoures T, Souraud J-B, Masquelet A-C, Collombet J-M. Towards Understanding Therapeutic Failures in Masquelet Surgery: First Evidence that Defective Induced Membrane Properties are Associated with Clinical Failures. Journal of Clinical Medicine. 2020; 9(2):450. https://doi.org/10.3390/jcm9020450
Chicago/Turabian StyleDurand, Marjorie, Laure Barbier, Laurent Mathieu, Thomas Poyot, Thomas Demoures, Jean-Baptiste Souraud, Alain-Charles Masquelet, and Jean-Marc Collombet. 2020. "Towards Understanding Therapeutic Failures in Masquelet Surgery: First Evidence that Defective Induced Membrane Properties are Associated with Clinical Failures" Journal of Clinical Medicine 9, no. 2: 450. https://doi.org/10.3390/jcm9020450