Endometriosis and the Fallopian Tubes: Theories of Origin and Clinical Implications

 , , , and
, , , and 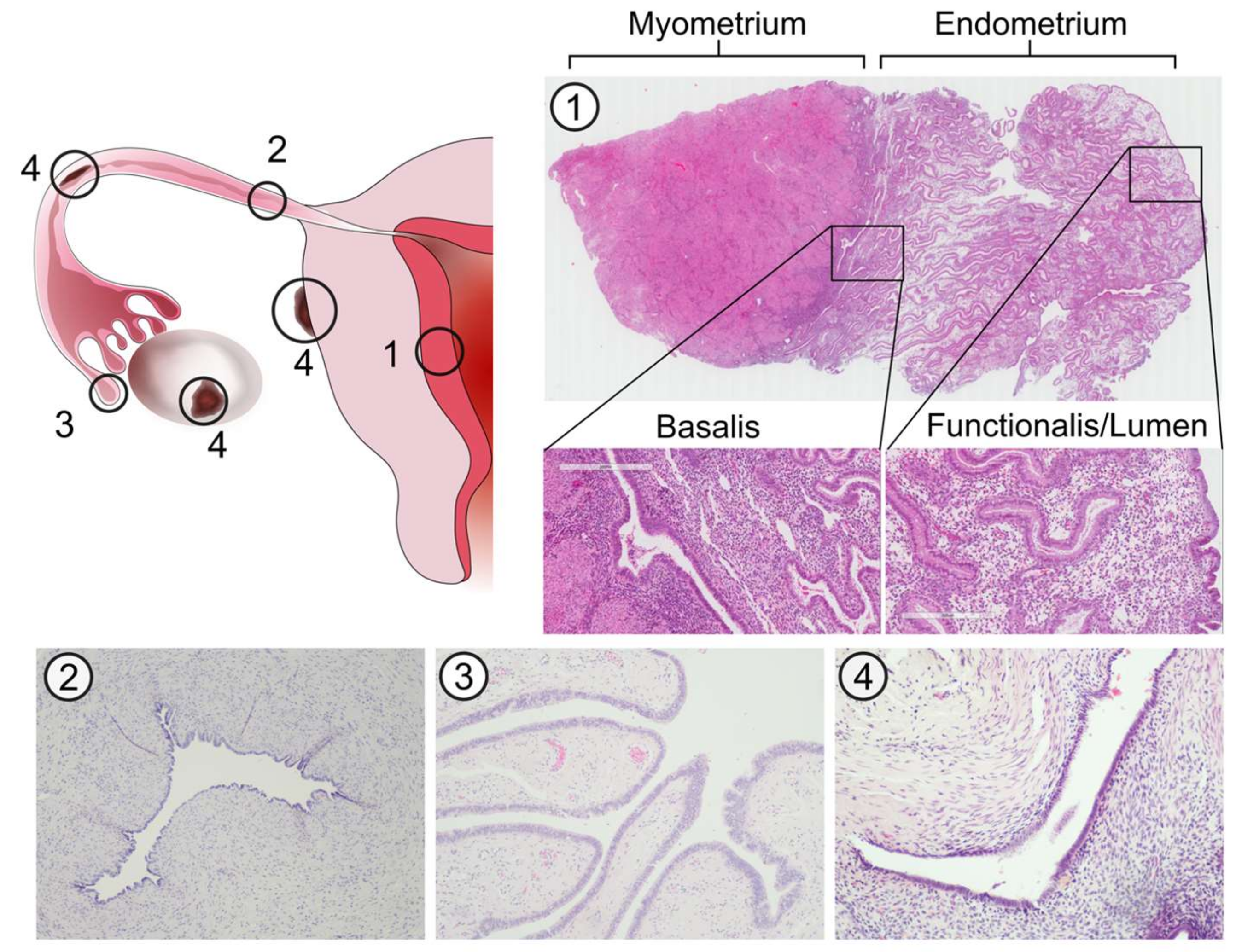{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Pathophysiology and Pathogenesis
2.1. Common Embryological Origin of the Endometrium and Fallopian Tubes
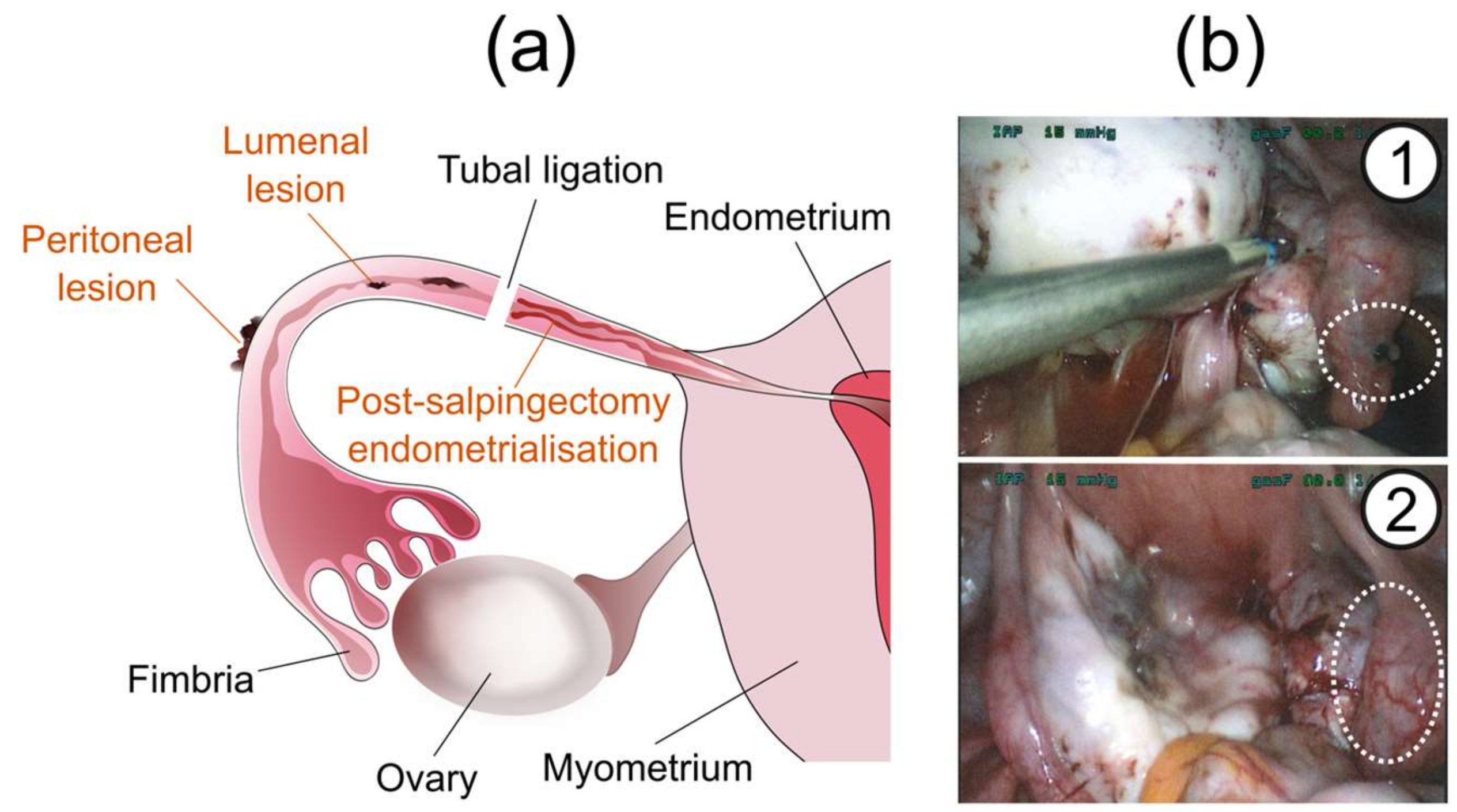
2.2. Types of Tubal Endometriosis
2.3. Asymmetric Presentation of Tubal Endometriosis
2.4. Prevalence of Tubal Endometriosis
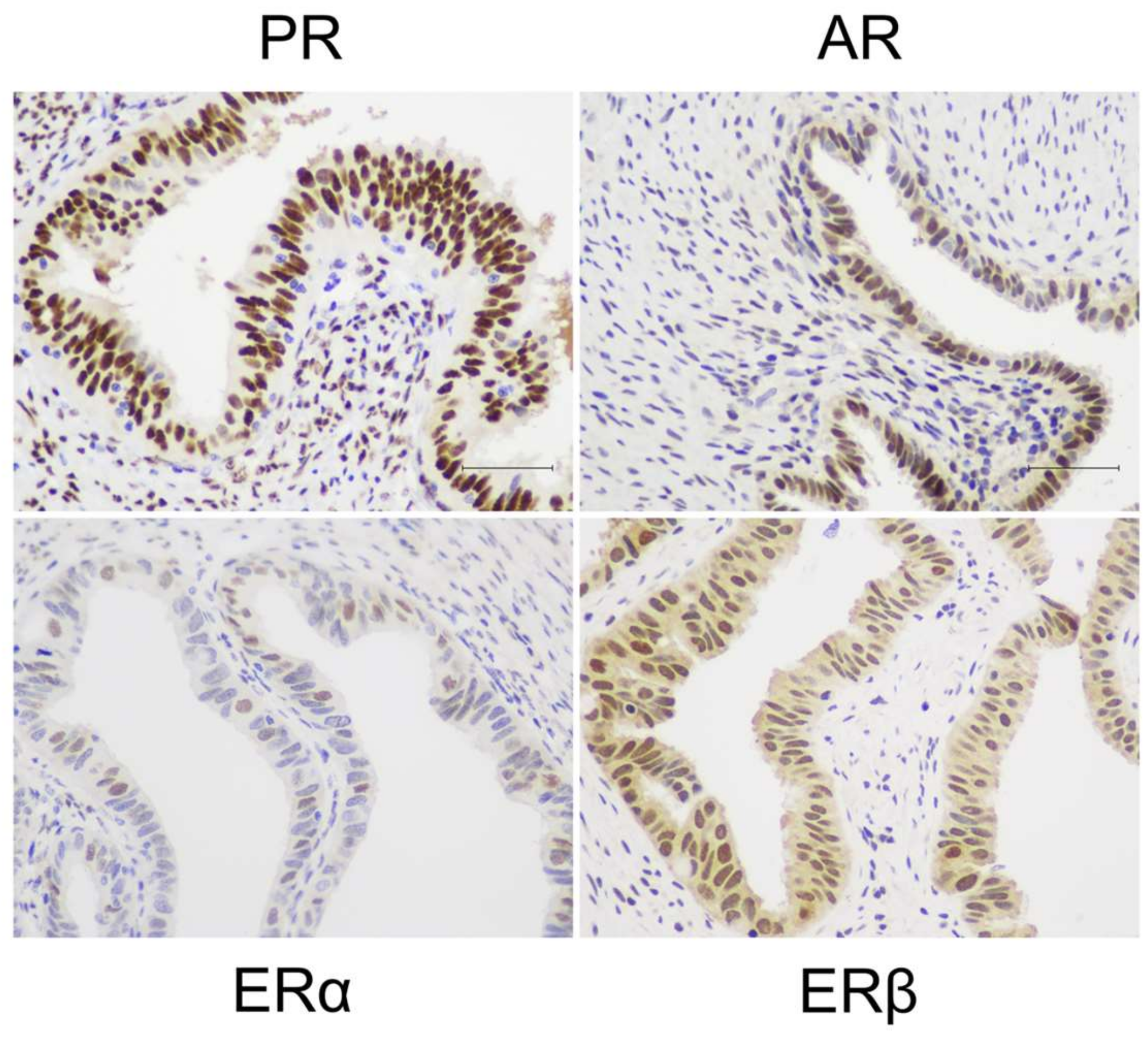
2.5. Shared Features of Hormone Responsiveness in Endometriotic Lesions and Fallopian Tubes
3. Tubal Origins of Endometriosis
3.1. Stem Cell Origin Theories
3.2. Tubal Stem Cells
4. Causes of Tubal Dysfunction in Endometriosis
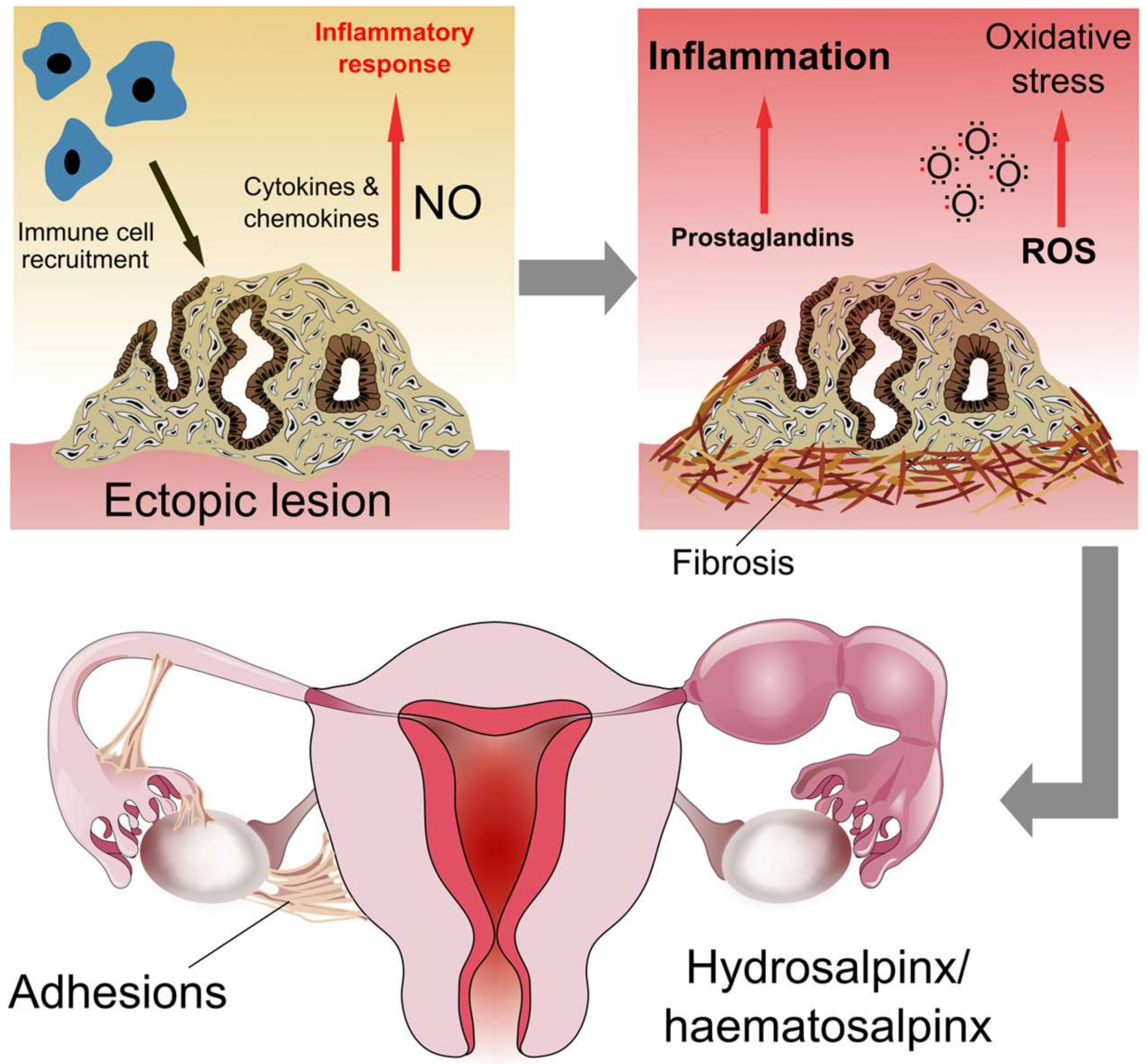
4.1. Inflammation
4.2. Tubal Obstruction, Adhesions and Hydrosalpinx
5. Consequences of Endometriosis-Associated Tubal Dysfunction
5.1. Subfertility
5.2. Association with Pelvic Pain
5.3. Ectopic Pregnancy
6. Association with Cancer
6.1. Incidence of Ovarian Cancer and Association with Endometriosis
6.2. Endometriosis-Associated Ovarian Cancer (EAOC)
6.3. Tubal Origins of Ovarian Cancer
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Shafrir, A.L.; Farland, L.V.; Shah, D.K.; Harris, H.R.; Kvaskoff, M.; Zondervan, K.; Missmer, S.A. Risk for and consequences of endometriosis: A critical epidemiologic review. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 51, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zondervan, K.T.; Becker, C.M.; Missmer, S.A. Endometriosis. N. Engl. J. Med. 2020, 382, 1244–1256. [Google Scholar] [CrossRef] [PubMed]
- Carter, J.E. Combined hysteroscopic and laproscopic findings in patients with chronic pelvic pain. J. Am. Assoc. Gynecol. Laparosc. 1994, 2, 43–47. [Google Scholar] [CrossRef]
- Sourial, S.; Tempest, N.; Hapangama, D.K. Theories on the pathogenesis of endometriosis. Int. J. Reprod. Med. 2014, 2014, 179515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.J.; Park, Y.M.; Jee, B.C.; Kim, Y.B.; Suh, C.S. Various anatomic locations of surgically proven endometriosis: A single-center experience. Obstet. Gynecol. Sci. 2015, 58, 53–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leyendecker, G.; Herbertz, M.; Kunz, G.; Mall, G. Endometriosis results from the dislocation of basal endometrium. Hum. Reprod. 2002, 17, 2725–2736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampson, J.A. Peritoneal endometriosis due to the menstrual dissemination of endometrial tissue into the peritoneal cavity. Am. J. Obstet. Gynecol. 1927, 14, 422–469. [Google Scholar] [CrossRef]
- Rock, J.A.; Markham, S.M. Pathogenesis of endometriosis. Lancet 1992, 340, 1264–1267. [Google Scholar] [CrossRef]
- Nap, A.W.; Groothuis, P.G.; Demir, A.Y.; Evers, J.L.; Dunselman, G.A. Pathogenesis of endometriosis. Best Pract. Res. Clin. Obstet. Gynaecol. 2004, 18, 233–244. [Google Scholar] [CrossRef]
- Tempest, N.; Maclean, A.; Hapangama, D.K. Endometrial stem cell markers: Current concepts and unresolved questions. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cousins, F.L.; Dorien, F.O.; Gargett, C.E. Endometrial stem/progenitor cells and their role in the pathogenesis of endometriosis. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 50, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Leyendecker, G.; Kunz, G.; Herbertz, M.; Beil, D.; Huppert, P.; Mall, G.; Kissler, S.; Noe, M.; Wildt, L. Uterine peristaltic activity and the development of endometriosis. Ann. N. Y. Acad. Sci. 2004, 1034, 338–355. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, T.; Yoshimura, Y. Stem cell theory for the pathogenesis of endometriosis. Front. Biosci. 2012, 4, 2754–2763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audebert, A.; Petousis, S.; Margioula-Siarkou, C.; Ravanos, K.; Prapas, N.; Prapas, Y. Anatomic distribution of endometriosis: A reappraisal based on series of 1101 patients. Eur. J. Obstet. Gynecol. Reprod. Biol. 2018, 230, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Rock, J.A.; Parmley, T.H.; King, T.M.; Laufe, L.E.; Su, B.S. Endometriosis and the development of tuboperitoneal fistulas after tubal ligation. Fertil. Steril. 1981, 35, 16–20. [Google Scholar] [CrossRef]
- Donnez, J.; Casanas-Roux, F.; Ferin, J.; Thomas, K. Tubal polyps, epithelial inclusions, and endometriosis after tubal sterilization. Fertil. Steril. 1984, 41, 564–568. [Google Scholar] [CrossRef]
- Samimi, M.; Pourhanifeh, M.H.; Mehdizadehkashi, A.; Eftekhar, T.; Asemi, Z. The role of inflammation, oxidative stress, angiogenesis, and apoptosis in the pathophysiology of endometriosis: Basic science and new insights based on gene expression. J. Cell. Physiol. 2019, 234, 19384–19392. [Google Scholar] [CrossRef]
- Qi, H.; Zhang, H.; Zhao, X.; Qin, Y.; Liang, G.; He, X.; Zhang, J. Integrated analysis of mRNA and protein expression profiling in tubal endometriosis. Reproduction 2020, 159, 601–614. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.; Wang, L.; Wang, Y.; Zhang, T.; Li, L.; Cragun, J.M.; Chambers, S.K.; Kong, B.; Zheng, W. Tubal origin of ovarian endometriosis. Mod. Pathol. 2014, 27, 1154–1162. [Google Scholar] [CrossRef] [Green Version]
- Tortora, G.J.; Derrickson, B. Principles of Anatomy and Physiology; Wiley: Hoboken, NJ, USA, 2011. [Google Scholar]
- Maclean, A.; Kamal, A.; Adishesh, M.; Alnafakh, R.; Tempest, N.; Hapangama, D.K. Human uterine biopsy: Research value and common pitfalls. Int. J. Reprod. Med. 2020, 2020, 9275360. [Google Scholar] [CrossRef]
- Rezvani, M.; Shaaban, A.M. Fallopian tube disease in the nonpregnant patient. Radiographics 2011, 31, 527–548. [Google Scholar] [CrossRef] [Green Version]
- Foti, P.V.; Ognibene, N.; Spadola, S.; Caltabiano, R.; Farina, R.; Palmucci, S.; Milone, P.; Ettorre, G.C. Non-neoplastic diseases of the fallopian tube: MR imaging with emphasis on diffusion-weighted imaging. Insights Imaging 2016, 7, 311–327. [Google Scholar] [CrossRef] [Green Version]
- Qi, H.; Zhang, H.; Zhang, D.; Li, J.; Huang, Z.; Zhao, X.; Zhang, J. Reassessment of prevalence of tubal endometriosis, and its associated clinicopathologic features and risk factors in premenopausal women received salpingectomy. Eur. J. Obstet. Gynecol. Reprod. Biol. X 2019, 4, 100074. [Google Scholar] [CrossRef]
- Chakrabarti, I.; Ghosh, N. Post-salpingectomy endometriosis: An under-recognized entity. J. Midlife Health 2010, 1, 91–92. [Google Scholar] [CrossRef]
- Clement, P.B. The pathology of endometriosis: A survey of the many faces of a common disease emphasizing diagnostic pitfalls and unusual and newly appreciated aspects. Adv. Anat. Pathol. 2007, 14, 241–260. [Google Scholar] [CrossRef]
- Kurman, R.J.; Vang, R.; Junge, J.; Hannibal, C.G.; Kjaer, S.K.; Shih Ie, M. Papillary tubal hyperplasia: The putative precursor of ovarian atypical proliferative (borderline) serous tumors, noninvasive implants, and endosalpingiosis. Am. J. Surg. Pathol. 2011, 35, 1605–1614. [Google Scholar] [CrossRef] [PubMed]
- Tao, X.; Xie, Y.; Wang, L.; Gu, W.; Yu, X.; Zhou, X. The expression of Cox-2, NF-κB, and VEGF in ectopic endometrial tissues within fallopian tubes suggests different etiologies. Int. J. Gynecol. Pathol. 2014, 33, 411–417. [Google Scholar] [CrossRef]
- Jenkins, S.; Olive, D.L.; Haney, A.F. Endometriosis: Pathogenetic implications of the anatomic distribution. Obstet. Gynecol. 1986, 67, 335–338. [Google Scholar]
- Xue, R.H.; Li, J.; Huang, Z.; Li, Z.Z.; Chen, L.; Lin, Q.; Huang, H.F. Is tubal endometriosis an asymmetric disease? A 17-year retrospective study. Arch. Gynecol. Obstet. 2020, 301, 721–727. [Google Scholar] [CrossRef]
- Ulukus, M.; Yeniel, A.; Ergenoglu, A.M.; Mermer, T. Right endometrioma is related with more extensive obliteration of the Douglas pouch. Arch. Gynecol. Obstet. 2012, 285, 1483–1486. [Google Scholar] [CrossRef]
- Qi, H.; Liang, G.; Yu, J.; Wang, X.; Liang, Y.; He, X.; Feng, T.; Zhang, J. Genome-wide profiling of miRNA expression patterns in tubal endometriosis. Reproduction 2019, 157, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Zhang, D.; Ouyang, J.; Liang, Y.; Zhang, H.; Huang, Z.; Liang, G.; Zhu, Q.; Guan, X.; Zhang, J. Effects of pelvic endometriosis and adenomyosis on ciliary beat frequency and muscular contractions in the human fallopian tube. Reprod. Biol. Endocrinol. 2018, 16, 48. [Google Scholar] [CrossRef] [Green Version]
- Krikun, G.; Schatz, F.; Taylor, R.; Critchley, H.O.; Rogers, P.A.; Huang, J.; Lockwood, C.J. Endometrial endothelial cell steroid receptor expression and steroid effects on gene expression. J. Clin. Endocrinol. Metab. 2005, 90, 1812–1818. [Google Scholar] [CrossRef] [Green Version]
- Hapangama, D.K.; Kamal, A.M.; Bulmer, J.N. Estrogen receptor β: The guardian of the endometrium. Hum. Reprod. Update 2015, 21, 174–193. [Google Scholar] [CrossRef] [Green Version]
- Maybin, J.A.; Critchley, H.O. Progesterone: A pivotal hormone at menstruation. Ann. N. Y. Acad. Sci. 2011, 1221, 88–97. [Google Scholar] [CrossRef]
- George, S.H.; Milea, A.; Shaw, P.A. Proliferation in the normal FTE is a hallmark of the follicular phase, not BRCA mutation status. Clin. Cancer Res. 2012, 18, 6199–6207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maclean, A.; Bhullar, J.; Miwa, S.; Glinicki, T.; Morais, C.; Martin, F.; Valentijn, A.; Vasieva, O.; Tempest, N.; Drury, J.; et al. Multi-methodological approach defines sub-populations of epithelial cells in human endometrium. In Proceedings of the Society for Reproductive Investigation, 66th Annual Scientific Meeting, Paris, France, 15 March 2019; p. 103A. [Google Scholar]
- Maclean, A.; Bunni, E.; Makrydima, S.; Withington, A.; Kamal, A.; Valentijin, A.; Hapangama, D.K. Fallopian epithelial cells express androgen receptor and have a distinct hormonal responsiveness when compared with endometrial epithelium. Hum. Reprod. 2020, in press. [Google Scholar]
- Horne, A.W.; King, A.E.; Shaw, E.; McDonald, S.E.; Williams, A.R.; Saunders, P.T.; Critchley, H.O. Attenuated sex steroid receptor expression in fallopian tube of women with ectopic pregnancy. J. Clin. Endocrinol. Metab. 2009, 94, 5146–5154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, R.; Norström, A.; Weijdegard, B.; Egecioglu, E.; Fernandez-Rodriguez, J.; Feng, Y.; Stener-Victorin, E.; Brännström, M.; Billig, H. Distinct expression pattern of Dicer1 correlates with ovarian-derived steroid hormone receptor expression in human Fallopian tubes during ovulation and the midsecretory phase. J. Clin. Endocrinol. Metab. 2011, 96, E869–E877. [Google Scholar] [CrossRef] [Green Version]
- Hapangama, D.K.; Drury, J.; Da Silva, L.; Al-Lamee, H.; Earp, A.; Valentijn, A.J.; Edirisinghe, D.P.; Murray, P.A.; Fazleabas, A.T.; Gargett, C.E. Abnormally located SSEA1+/SOX9+ endometrial epithelial cells with a basalis-like phenotype in the eutopic functionalis layer may play a role in the pathogenesis of endometriosis. Hum. Reprod. 2019, 34, 56–68. [Google Scholar] [CrossRef]
- Greene, A.D.; Lang, S.A.; Kendziorski, J.A.; Sroga-Rios, J.M.; Herzog, T.J.; Burns, K.A. Endometriosis: Where are we and where are we going? Reproduction 2016, 152, R63–R78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulun, S.E.; Monsavais, D.; Pavone, M.E.; Dyson, M.; Xue, Q.; Attar, E.; Tokunaga, H.; Su, E.J. Role of estrogen receptor-β in endometriosis. Semin. Reprod. Med. 2012, 30, 39–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, R.K.; Bulmer, J.N.; Searle, R.F. Phenotypic and functional studies of leukocytes in human endometrium and endometriosis. Hum. Reprod. Update 1998, 4, 702–709. [Google Scholar] [CrossRef] [Green Version]
- Shao, R.; Cao, S.; Wang, X.; Feng, Y.; Billig, H. The elusive and controversial roles of estrogen and progesterone receptors in human endometriosis. Am. J. Transl. Res. 2014, 6, 104–113. [Google Scholar]
- Carneiro, M.M.; Morsch, D.M.; Camargos, A.F.; Reis, F.M.; Spritzer, P.M. Androgen receptor and 5alpha-reductase are expressed in pelvic endometriosis. BJOG 2008, 115, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Alderman, M.H., 3rd; Yoder, N.; Taylor, H.S. The systemic effects of endometriosis. Semin. Reprod. Med. 2017, 35, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Schüring, A.N.; Dahlhues, B.; Korte, A.; Kiesel, L.; Titze, U.; Heitkötter, B.; Ruckert, C.; Götte, M. The endometrial stem cell markers notch-1 and numb are associated with endometriosis. Reprod. Biomed. Online 2018, 36, 294–301. [Google Scholar] [CrossRef]
- Halme, J.; Hammond, M.G.; Hulka, J.F.; Raj, S.G.; Talbert, L.M. Retrograde menstruation in healthy women and in patients with endometriosis. Obstet. Gynecol. 1984, 64, 151–154. [Google Scholar] [PubMed]
- Tempest, N.; Jansen, M.; Baker, A.M.; Hill, C.J.; Hale, M.; Magee, D.; Treanor, D.; Wright, N.A.; Hapangama, D.K. Histological 3D reconstruction and in vivo lineage tracing of the human endometrium. J. Pathol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Valentijn, A.J.; Palial, K.; Al-Lamee, H.; Tempest, N.; Drury, J.; Von Zglinicki, T.; Saretzki, G.; Murray, P.; Gargett, C.E.; Hapangama, D.K. SSEA-1 isolates human endometrial basal glandular epithelial cells: Phenotypic and functional characterization and implications in the pathogenesis of endometriosis. Hum. Reprod. 2013, 28, 2695–2708. [Google Scholar] [CrossRef] [Green Version]
- Nabeshima, H.; Murakami, T.; Yoshinaga, K.; Sato, K.; Terada, Y.; Okamura, K. Analysis of the clonality of ectopic glands in peritoneal endometriosis using laser microdissection. Fertil. Steril. 2003, 80, 1144–1150. [Google Scholar] [CrossRef]
- Tempest, N.; Baker, A.M.; Wright, N.A.; Hapangama, D.K. Does human endometrial LGR5 gene expression suggest the existence of another hormonally regulated epithelial stem cell niche? Hum. Reprod. 2018, 33, 1052–1062. [Google Scholar] [CrossRef] [PubMed]
- Schuijers, J.; Clevers, H. Adult mammalian stem cells: The role of Wnt, Lgr5 and R-spondins. EMBO J. 2012, 31, 2685–2696. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.M.; Graham, T.A.; Elia, G.; Wright, N.A.; Rodriguez-Justo, M. Characterization of LGR5 stem cells in colorectal adenomas and carcinomas. Sci. Rep. 2015, 5, 8654. [Google Scholar] [CrossRef] [Green Version]
- Jaks, V.; Barker, N.; Kasper, M.; van Es, J.H.; Snippert, H.J.; Clevers, H.; Toftgård, R. Lgr5 marks cycling, yet long-lived, hair follicle stem cells. Nat. Genet. 2008, 40, 1291–1299. [Google Scholar] [CrossRef]
- Barker, N.; Rookmaaker, M.B.; Kujala, P.; Ng, A.; Leushacke, M.; Snippert, H.; van de Wetering, M.; Tan, S.; Van Es, J.H.; Huch, M.; et al. Lgr5(+ve) stem/progenitor cells contribute to nephron formation during kidney development. Cell. Rep. 2012, 2, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Beumer, J.; Clevers, H. Regulation and plasticity of intestinal stem cells during homeostasis and regeneration. Development 2016, 143, 3639–3649. [Google Scholar] [CrossRef] [Green Version]
- Ng, A.; Tan, S.; Singh, G.; Rizk, P.; Swathi, Y.; Tan, T.Z.; Huang, R.Y.; Leushacke, M.; Barker, N. Lgr5 marks stem/progenitor cells in ovary and tubal epithelia. Nat. Cell. Biol. 2014, 16, 745–757. [Google Scholar] [CrossRef]
- Paik, D.Y.; Janzen, D.M.; Schafenacker, A.M.; Velasco, V.S.; Shung, M.S.; Cheng, D.; Huang, J.; Witte, O.N.; Memarzadeh, S. Stem-like epithelial cells are concentrated in the distal end of the fallopian tube: A site for injury and serous cancer initiation. Stem Cells 2012, 30, 2487–2497. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y.; Ning, G.; Howitt, B.E.; Mehra, K.; Wu, L.; Wang, X.; Hong, Y.; Kern, F.; Wei, T.S.; Zhang, T.; et al. In vitro and in vivo correlates of physiological and neoplastic human Fallopian tube stem cells. J. Pathol. 2016, 238, 519–530. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.H.; Chu, T.Y.; Ding, D.C. Human fallopian tube epithelial cells exhibit stemness features, self-renewal capacity, and Wnt-related organoid formation. J. Biomed. Sci. 2020, 27, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.H.; Ding, D.C.; Chu, T.Y. Estradiol and progesterone induced differentiation and increased stemness gene expression of human fallopian tube epithelial cells. J. Cancer 2019, 10, 3028–3036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tai, F.W.; Chang, C.Y.; Chiang, J.H.; Lin, W.C.; Wan, L. Association of pelvic inflammatory disease with risk of endometriosis: A nationwide cohort study involving 141,460 individuals. J. Clin. Med. 2018, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, J.F.; Ryan, I.P.; Milam, T.D.; Murai, J.T.; Schriock, E.D.; Landers, D.V.; Taylor, R.N. Interleukin-6 secretion in vitro is up-regulated in ectopic and eutopic endometrial stromal cells from women with endometriosis. J. Clin. Endocrinol. Metab. 1996, 81, 1118–1122. [Google Scholar] [CrossRef] [Green Version]
- Kessler, M.; Hoffmann, K.; Fritsche, K.; Brinkmann, V.; Mollenkopf, H.J.; Thieck, O.; Teixeira da Costa, A.R.; Braicu, E.I.; Sehouli, J.; Mangler, M.; et al. Chronic Chlamydia infection in human organoids increases stemness and promotes age-dependent CpG methylation. Nat. Commun. 2019, 10, 1194. [Google Scholar] [CrossRef]
- Du, H.; Taylor, H.S. Contribution of bone marrow-derived stem cells to endometrium and endometriosis. Stem Cells 2007, 25, 2082–2086. [Google Scholar] [CrossRef]
- Makiyan, Z. Endometriosis origin from primordial germ cells. Organogenesis 2017, 13, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Bulun, S.E.; Yilmaz, B.D.; Sison, C.; Miyazaki, K.; Bernardi, L.; Liu, S.; Kohlmeier, A.; Yin, P.; Milad, M.; Wei, J. Endometriosis. Endocr. Rev. 2019, 40, 1048–1079. [Google Scholar] [CrossRef]
- Hess, A.P.; Talbi, S.; Hamilton, A.E.; Baston-Buest, D.M.; Nyegaard, M.; Irwin, J.C.; Barragan, F.; Kruessel, J.S.; Germeyer, A.; Giudice, L.C. The human oviduct transcriptome reveals an anti-inflammatory, anti-angiogenic, secretory and matrix-stable environment during embryo transit. Reprod. Biomed. Online 2013, 27, 423–435. [Google Scholar] [CrossRef] [Green Version]
- Matsushima, T.; Kaseki, H.; Ishihara, K.; Araki, T. Assessment of fallopian tube cytology for the diagnosis of endometriosis and hydrosalpinx. J. Nippon Med. Sch. 2002, 69, 445–450. [Google Scholar] [CrossRef] [Green Version]
- Haney, A.F.; Misukonis, M.A.; Weinberg, J.B. Macrophages and infertility: Oviductal macrophages as potential mediators of infertility. Fertil. Steril. 1983, 39, 310–315. [Google Scholar] [CrossRef]
- Wu, M.Y.; Chao, K.H.; Yang, J.H.; Lee, T.H.; Yang, Y.S.; Ho, H.N. Nitric oxide synthesis is increased in the endometrial tissue of women with endometriosis. Hum. Reprod. 2003, 18, 2668–2671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekerhovd, E.; Brännström, M.; Alexandersson, M.; Norström, A. Evidence for nitric oxide mediation of contractile activity in isolated strips of the human Fallopian tube. Hum. Reprod. 1997, 12, 301–305. [Google Scholar] [CrossRef] [Green Version]
- Symons, L.K.; Miller, J.E.; Tyryshkin, K.; Monsanto, S.P.; Marks, R.M.; Lingegowda, H.; Vanderbeck, K.; Childs, T.; Young, S.L.; Lessey, B.A.; et al. Neutrophil recruitment and function in endometriosis patients and a syngeneic murine model. FASEB J. 2020, 34, 1558–1575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osuga, Y.; Koga, K.; Hirota, Y.; Hirata, T.; Yoshino, O.; Taketani, Y. Lymphocytes in endometriosis. Am. J. Reprod. Immunol. 2011, 65, 1–10. [Google Scholar] [CrossRef]
- Smith, J.M.; Wira, C.R.; Fanger, M.W.; Shen, L. Human fallopian tube neutrophils—A distinct phenotype from blood neutrophils. Am. J. Reprod. Immunol. 2006, 56, 218–229. [Google Scholar] [CrossRef]
- Donnez, J.; Binda, M.M.; Donnez, O.; Dolmans, M.M. Oxidative stress in the pelvic cavity and its role in the pathogenesis of endometriosis. Fertil. Steril. 2016, 106, 1011–1017. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Agarwal, A.; Krajcir, N.; Alvarez, J.G. Role of oxidative stress in endometriosis. Reprod. Biomed. Online 2006, 13, 126–134. [Google Scholar] [CrossRef]
- Czernobilsky, B.; Silverstein, A. Salpingitis in ovarian endometriosis. Fertil. Steril. 1978, 30, 45–49. [Google Scholar] [CrossRef]
- Nabekura, H.; Koike, H.; Ohtsuka, T.; Yamaguchi, M.; Miyakawa, I.; Mori, N. Fallopian tube prostaglandin production with and without endometriosis. Int. J. Fertil. Menopausal Stud. 1994, 39, 57–63. [Google Scholar]
- Sacco, K.; Portelli, M.; Pollacco, J.; Schembri-Wismayer, P.; Calleja-Agius, J. The role of prostaglandin E2 in endometriosis. Gynecol. Endocrinol. 2012, 28, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.J.; Yang, J.; Liu, Z.; Yang, G.; Shen, Z.J. Telocytes damage in endometriosis-affected rat oviduct and potential impact on fertility. J. Cell. Mol. Med. 2015, 19, 452–462. [Google Scholar] [CrossRef] [PubMed]
- Volodarsky-Perel, A.; Buckett, W.; Tulandi, T. Treatment of hydrosalpinx in relation to IVF outcome: A systematic review and meta-analysis. Reprod. Biomed. Online 2019, 39, 413–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, N.; van Voorst, S.; Sowter, M.C.; Strandell, A.; Mol, B.W. Surgical treatment for tubal disease in women due to undergo in vitro fertilisation. Cochrane Database Syst. Rev. 2010, 2010, CD002125. [Google Scholar] [CrossRef]
- Papaioannou, S. A hypothesis for the pathogenesis and natural history of proximal tubal blockage. Hum. Reprod. 2004, 19, 481–485. [Google Scholar] [CrossRef] [Green Version]
- Guan, J.; Watrelot, A. Fallopian tube subtle pathology. Best Pract. Res. Clin. Obstet. Gynaecol. 2019, 59, 25–40. [Google Scholar] [CrossRef]
- Baggish, M.S.; Karram, M.M. Atlas of Pelvic Anatomy and Gynecologic Surgery; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Fortier, K.J.; Haney, A.F. The pathologic spectrum of uterotubal junction obstruction. Obstet. Gynecol. 1985, 65, 93–98. [Google Scholar]
- Zegers-Hochschild, F.; Adamson, G.D.; Dyer, S.; Racowsky, C.; de Mouzon, J.; Sokol, R.; Rienzi, L.; Sunde, A.; Schmidt, L.; Cooke, I.D.; et al. The international glossary on infertility and fertility care, 2017. Fertil. Steril. 2017, 108, 393–406. [Google Scholar] [CrossRef] [Green Version]
- National Institute for Health and Care Excellence. Fertility Problems: Assessment and Treatment Clinical Guideline [CG156]. Available online: https://www.nice.org.uk/guidance/cg156/chapter/Context (accessed on 15 May 2020).
- Macer, M.L.; Taylor, H.S. Endometriosis and infertility: A review of the pathogenesis and treatment of endometriosis-associated infertility. Obstet. Gynecol. Clin. N Am. 2012, 39, 535–549. [Google Scholar] [CrossRef] [Green Version]
- Halis, G.; Arici, A. Endometriosis and inflammation in infertility. Ann. N. Y. Acad. Sci. 2004, 1034, 300–315. [Google Scholar] [CrossRef]
- Lyons, R.A.; Djahanbakhch, O.; Saridogan, E.; Naftalin, A.A.; Mahmood, T.; Weekes, A.; Chenoy, R. Peritoneal fluid, endometriosis, and ciliary beat frequency in the human fallopian tube. Lancet 2002, 360, 1221–1222. [Google Scholar] [CrossRef]
- Yang, X.J.; Xu, J.Y.; Shen, Z.J.; Zhao, J. Immunohistochemical alterations of cajal-like type of tubal interstitial cells in women with endometriosis and tubal ectopic pregnancy. Arch. Gynecol. Obstet. 2013, 288, 1295–1300. [Google Scholar] [CrossRef] [PubMed]
- Kissler, S.; Zangos, S.; Wiegratz, I.; Kohl, J.; Rody, A.; Gaetje, R.; Doebert, N.; Wildt, L.; Kunz, G.; Leyendecker, G.; et al. Utero-tubal sperm transport and its impairment in endometriosis and adenomyosis. Ann. N. Y. Acad. Sci. 2007, 1101, 38–48. [Google Scholar] [CrossRef]
- Reeve, L.; Lashen, H.; Pacey, A.A. Endometriosis affects sperm-endosalpingeal interactions. Hum. Reprod. 2005, 20, 448–451. [Google Scholar] [CrossRef] [Green Version]
- Chronic Pelvic Pain: ACOG Practice Bulletin, Number 218. Obstet. Gynecol. 2020, 135, e98–e109. [CrossRef] [PubMed]
- Howard, F.M. The role of laparoscopy in chronic pelvic pain: Promise and pitfalls. Obstet. Gynecol. Surv. 1993, 48, 357–387. [Google Scholar] [CrossRef] [PubMed]
- Coxon, L.; Horne, A.W.; Vincent, K. Pathophysiology of endometriosis-associated pain: A review of pelvic and central nervous system mechanisms. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 51, 53–67. [Google Scholar] [CrossRef]
- McKinnon, B.D.; Bertschi, D.; Bersinger, N.A.; Mueller, M.D. Inflammation and nerve fiber interaction in endometriotic pain. Trends Endocrinol. Metab. 2015, 26, 1–10. [Google Scholar] [CrossRef]
- Howard, F.M. Endometriosis and mechanisms of pelvic pain. J. Minim Invasive Gynecol. 2009, 16, 540–550. [Google Scholar] [CrossRef]
- Chen, C.; Huang, L.; Liu, P.; Su, G.; Li, W.; Lu, L.; Wang, L.; Li, X.; Duan, H.; Zou, C.; et al. Neurovascular quantitative study of the uterosacral ligament related to nerve-sparing radical hysterectomy. Eur. J. Obstet. Gynecol. Reprod. Biol. 2014, 172, 74–79. [Google Scholar] [CrossRef]
- Biswas, A.; Manivannan, M.; Srinivasan, M.A. Vibrotactile sensitivity threshold: Nonlinear stochastic mechanotransduction model of the Pacinian Corpuscle. IEEE Trans. Haptics. 2015, 8, 102–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anaf, V.; Simon, P.; El Nakadi, I.; Fayt, I.; Simonart, T.; Buxant, F.; Noel, J.C. Hyperalgesia, nerve infiltration and nerve growth factor expression in deep adenomyotic nodules, peritoneal and ovarian endometriosis. Hum. Reprod. 2002, 17, 1895–1900. [Google Scholar] [CrossRef] [Green Version]
- Tokushige, N.; Markham, R.; Russell, P.; Fraser, I.S. Nerve fibres in peritoneal endometriosis. Hum. Reprod. 2006, 21, 3001–3007. [Google Scholar] [CrossRef] [Green Version]
- Tokushige, N.; Russell, P.; Black, K.; Barrera, H.; Dubinovsky, S.; Markham, R.; Fraser, I.S. Nerve fibers in ovarian endometriomas. Fertil. Steril. 2010, 94, 1944–1947. [Google Scholar] [CrossRef]
- Arnold, J.; Vercellino, G.F.; Chiantera, V.; Schneider, A.; Mechsner, S.; Barcena de Arellano, M.L. Neuroimmunomodulatory alterations in non-lesional peritoneum close to peritoneal endometriosis. Neuroimmunomodulation 2013, 20, 9–18. [Google Scholar] [CrossRef]
- Tulandi, T.; Felemban, A.; Chen, M.F. Nerve fibers and histopathology of endometriosis-harboring peritoneum. J. Am. Assoc. Gynecol. Laparosc. 2001, 8, 95–98. [Google Scholar] [CrossRef]
- Barcena de Arellano, M.L.; Arnold, J.; Vercellino, G.F.; Chiantera, V.; Ebert, A.D.; Schneider, A.; Mechsner, S. Influence of nerve growth factor in endometriosis-associated symptoms. Reprod. Sci. 2011, 18, 1202–1210. [Google Scholar] [CrossRef]
- McKinnon, B.; Bersinger, N.A.; Wotzkow, C.; Mueller, M.D. Endometriosis-associated nerve fibers, peritoneal fluid cytokine concentrations, and pain in endometriotic lesions from different locations. Fertil. Steril. 2012, 97, 373–380. [Google Scholar] [CrossRef]
- Mechsner, S.; Kaiser, A.; Kopf, A.; Gericke, C.; Ebert, A.; Bartley, J. A pilot study to evaluate the clinical relevance of endometriosis-associated nerve fibers in peritoneal endometriotic lesions. Fertil. Steril. 2009, 92, 1856–1861. [Google Scholar] [CrossRef]
- Kajitani, T.; Maruyama, T.; Asada, H.; Uchida, H.; Oda, H.; Uchida, S.; Miyazaki, K.; Arase, T.; Ono, M.; Yoshimura, Y. Possible involvement of nerve growth factor in dysmenorrhea and dyspareunia associated with endometriosis. Endocr. J. 2013, 60, 1155–1164. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Chen, J.; Sun, Y.; Huang, X.; Xu, H.; Zhang, X. Loss of nerve fibers in the oviduct isthmus in women with hydrosalpinx. Acta Histochem. 2013, 115, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, S.; Canis, M.; Pouly, J.L.; Wattiez, A.; Okamura, K.; Mage, G. Cyclooxygenase-2 expression in deep endometriosis and matched eutopic endometrium. Fertil. Steril. 2004, 82, 1309–1315. [Google Scholar] [CrossRef] [PubMed]
- Ota, H.; Igarashi, S.; Sasaki, M.; Tanaka, T. Distribution of cyclooxygenase-2 in eutopic and ectopic endometrium in endometriosis and adenomyosis. Hum. Reprod. 2001, 16, 561–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keenan, J.A.; Chen, T.T.; Chadwell, N.L.; Torry, D.S.; Caudle, M.R. IL-1 beta, TNF-alpha, and IL-2 in peritoneal fluid and macrophage-conditioned media of women with endometriosis. Am. J. Reprod. Immunol. 1995, 34, 381–385. [Google Scholar] [CrossRef]
- Rapkin, A.; Morgan, M.; Bonpane, C.; Martinez-Maza, O. Peritoneal fluid interleukin-6 in women with chronic pelvic pain. Fertil. Steril. 2000, 74, 325–328. [Google Scholar] [CrossRef]
- Punnonen, J.; Teisala, K.; Ranta, H.; Bennett, B.; Punnonen, R. Increased levels of interleukin-6 and interleukin-10 in the peritoneal fluid of patients with endometriosis. Am. J. Obstet. Gynecol. 1996, 174, 1522–1526. [Google Scholar] [CrossRef]
- Richter, O.N.; Dorn, C.; Rösing, B.; Flaskamp, C.; Ulrich, U. Tumor necrosis factor alpha secretion by peritoneal macrophages in patients with endometriosis. Arch. Gynecol. Obstet. 2005, 271, 143–147. [Google Scholar] [CrossRef]
- Arici, A.; Oral, E.; Attar, E.; Tazuke, S.I.; Olive, D.L. Monocyte chemotactic protein-1 concentration in peritoneal fluid of women with endometriosis and its modulation of expression in mesothelial cells. Fertil. Steril. 1997, 67, 1065–1072. [Google Scholar] [CrossRef]
- Moalem, G.; Tracey, D.J. Immune and inflammatory mechanisms in neuropathic pain. Brain. Res. Rev. 2006, 51, 240–264. [Google Scholar] [CrossRef]
- D’Hooghe, T.M.; Debrock, S.; Hill, J.A.; Meuleman, C. Endometriosis and subfertility: Is the relationship resolved? Semin. Reprod. Med. 2003, 21, 243–254. [Google Scholar] [CrossRef]
- Bulletti, C.; Coccia, M.E.; Battistoni, S.; Borini, A. Endometriosis and infertility. J. Assist. Reprod. Genet. 2010, 27, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Perkins, K.M.; Boulet, S.L.; Kissin, D.M.; Jamieson, D.J. Risk of ectopic pregnancy associated with assisted reproductive technology in the United States, 2001–2011. Obstet. Gynecol. 2015, 125, 70–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farland, L.V.; Prescott, J.; Sasamoto, N.; Tobias, D.K.; Gaskins, A.J.; Stuart, J.J.; Carusi, D.A.; Chavarro, J.E.; Horne, A.W.; Rich-Edwards, J.W.; et al. Endometriosis and risk of adverse pregnancy outcomes. Obstet. Gynecol. 2019, 134, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Yong, P.J.; Matwani, S.; Brace, C.; Quaiattini, A.; Bedaiwy, M.A.; Albert, A.; Allaire, C. Endometriosis and ectopic pregnancy: A meta-analysis. J. Minim. Invasive. Gynecol. 2020, 27, 352–361. [Google Scholar] [CrossRef] [Green Version]
- Shaw, J.L.; Dey, S.K.; Critchley, H.O.; Horne, A.W. Current knowledge of the aetiology of human tubal ectopic pregnancy. Hum. Reprod. Update 2010, 16, 432–444. [Google Scholar] [CrossRef] [Green Version]
- Patton, D.L.; Moore, D.E.; Spadoni, L.R.; Soules, M.R.; Halbert, S.A.; Wang, S.P. A comparison of the fallopian tube’s response to overt and silent salpingitis. Obstet. Gynecol. 1989, 73, 622–630. [Google Scholar]
- Kutluay, L.; Vicdan, K.; Turan, C.; Batioğlu, S.; Oğuz, S.; Gökmen, O. Tubal histopathology in ectopic pregnancies. Eur. J. Obstet. Gynecol. Reprod. Biol. 1994, 57, 91–94. [Google Scholar] [CrossRef]
- Hjordt Hansen, M.V.; Dalsgaard, T.; Hartwell, D.; Skovlund, C.W.; Lidegaard, O. Reproductive prognosis in endometriosis. A national cohort study. Acta Obstet. Gynecol. Scand. 2014, 93, 483–489. [Google Scholar] [CrossRef]
- Reid, B.M.; Permuth, J.B.; Sellers, T.A. Epidemiology of ovarian cancer: A review. Cancer Biol. Med. 2017, 14, 9–32. [Google Scholar] [CrossRef] [Green Version]
- National Institute for Health and Care Excellence. Ovarian Cancer. Available online: https://cks.nice.org.uk/ovarian-cancer#!topicSummary (accessed on 13 May 2020).
- Kim, H.S.; Kim, T.H.; Chung, H.H.; Song, Y.S. Risk and prognosis of ovarian cancer in women with endometriosis: A meta-analysis. Br. J. Cancer 2014, 110, 1878–1890. [Google Scholar] [CrossRef]
- Wang, C.; Liang, Z.; Liu, X.; Zhang, Q.; Li, S. The Association between Endometriosis, Tubal ligation, hysterectomy and epithelial ovarian cancer: Meta-analyses. Int. J. Environ. Res. Public Health 2016, 13. [Google Scholar] [CrossRef] [Green Version]
- Mutch, D.G.; Prat, J. 2014 FIGO staging for ovarian, fallopian tube and peritoneal cancer. Gynecol. Oncol. 2014, 133, 401–404. [Google Scholar] [CrossRef]
- Van Gorp, T.; Amant, F.; Neven, P.; Vergote, I.; Moerman, P. Endometriosis and the development of malignant tumours of the pelvis: A review of literature. Best Pract. Res. Clin. Obstet. Gynaecol. 2004, 18, 349–371. [Google Scholar] [CrossRef]
- Hermens, M.; van Altena, A.M.; Bulten, J.; Siebers, A.G.; Bekkers, R.L.M. Increased association of ovarian cancer in women with histological proven endosalpingiosis. Cancer Epidemiol. 2020, 65, 101700. [Google Scholar] [CrossRef]
- Singh, N.; Gilks, C.B.; Hirschowitz, L.; Kehoe, S.; McNeish, I.A.; Miller, D.; Naik, R.; Wilkinson, N.; McCluggage, W.G. Primary site assignment in tubo-ovarian high-grade serous carcinoma: Consensus statement on unifying practice worldwide. Gynecol. Oncol. 2016, 141, 195–198. [Google Scholar] [CrossRef] [Green Version]
- Long Roche, K.C.; Abu-Rustum, N.R.; Nourmoussavi, M.; Zivanovic, O. Risk-reducing salpingectomy: Let us be opportunistic. Cancer 2017, 123, 1714–1720. [Google Scholar] [CrossRef] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hill, C.J.; Fakhreldin, M.; Maclean, A.; Dobson, L.; Nancarrow, L.; Bradfield, A.; Choi, F.; Daley, D.; Tempest, N.; Hapangama, D.K. Endometriosis and the Fallopian Tubes: Theories of Origin and Clinical Implications. J. Clin. Med. 2020, 9, 1905. https://doi.org/10.3390/jcm9061905
Hill CJ, Fakhreldin M, Maclean A, Dobson L, Nancarrow L, Bradfield A, Choi F, Daley D, Tempest N, Hapangama DK. Endometriosis and the Fallopian Tubes: Theories of Origin and Clinical Implications. Journal of Clinical Medicine. 2020; 9(6):1905. https://doi.org/10.3390/jcm9061905
Chicago/Turabian StyleHill, Christopher J., Marwa Fakhreldin, Alison Maclean, Lucy Dobson, Lewis Nancarrow, Alice Bradfield, Fiona Choi, Diandra Daley, Nicola Tempest, and Dharani K. Hapangama. 2020. "Endometriosis and the Fallopian Tubes: Theories of Origin and Clinical Implications" Journal of Clinical Medicine 9, no. 6: 1905. https://doi.org/10.3390/jcm9061905