
Concise Review: Comparison of Culture Membranes Used for Tissue Engineered Conjunctival Epithelial Equivalents

Abstract
:1. Conjunctiva


2. Conjunctival Stem Cells
3. Conjunctival Scarring Diseases
4. Tissue Engineered Conjunctival Equivalents
4.1. Biological Membranes

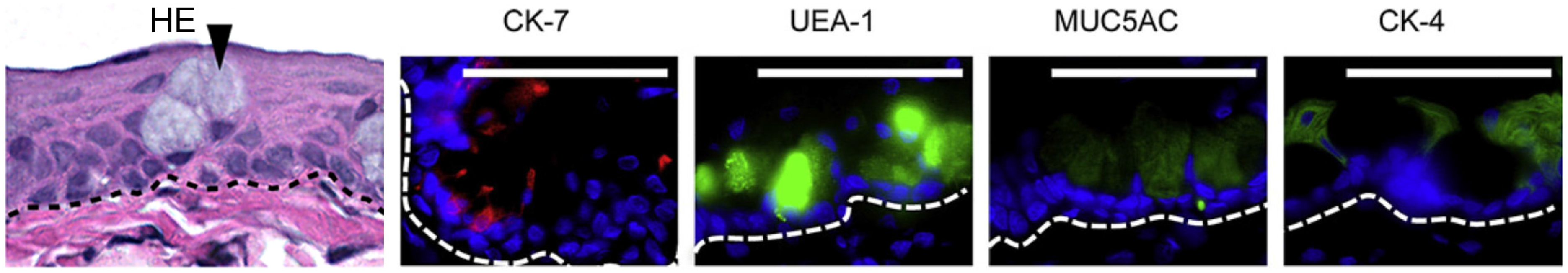{kind=link}
{kind=link}
| Substrate (s) | Cell Species | Explant/Suspension Culture | Culture Time (Days) | Feeder Cells | Air-Lifting | High Calcium | Basal Medium | Serum | Conjunctival Donor Site | Goblet Cells | Comment | Authors |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AM | Human | Explant | 21 | – | – | – | CNT50 | FBS/AS | – | Yes (with both serum type) | Rivas et al. 2014 | |
| AM | Human | Explant | 14 | No | No | No | DMEM:F12 | 5% FBS | Fornix/bulbus | Yes (<25% to 75% MUC5AC+) | Stratified culture | Eidet et al. 2014 |
| AM | Human | Explant | 12 | No | Yes | No | DMEM:F12 | 5% FBS | Bulbus | Yes (MUC5AC+, fever than in native conjunctiva) | Stratified culture | Tan et al. 2014 |
| AM | Rabbit | Explant | 8–15 | 3T3/No | Yes | – | – | – | Fornix | No MUC5AC− | Stratified culture (Ck3+/Ck12−) | Cho et al. 2014 |
| dAM | Human | Explant | 9–11 | 3T3 | Yes | – | DMEM:F12 | 10% FBS | – | Yes (PAS+, increased by γSI) | Stratified culture | Tian et al. 2014 |
| dAM | Human | Explant | – | No | No | – | DMEM:F12 | FBS | Fornix | Yes (PAS+) | Stratified culture | Silber et al. 2014 |
| dAM | Human | Suspension | 5 | No | No | – | KM (serum free or DMEM:F12) | 0%–20% FBS | Palpebra | Yes (100% Ck7+; best preserved by 10% FBS) | Martinez-Osorio et al. 2009 | |
| dAM | Human | Suspension | 21 | 3T3 | Yes | Yes | KM (serum free or DMEM:F12) | FBS | – | No (MUC5AC−) | Stratified culture | Tanioka et al. 2006 |
| dAM | Human | Explant | 14 | No | – | Yes | DMEM:F12 | FBS/HS | Bulbus | – | Stratified culture | Ang et al. 2005 |
| dAM | Human | Explant | 12–22 | No | Yes/No | Yes/No | KGM or DMEM/F12 | 0 or 10% FBS | Bulbus | Yes (MUC5AC detected by PCR in all groups) | Stratified culture | Ang et al. 2004 |
| dAM | Human | Explant | 11–15 | No | No | – | KGM:F12 | 10% FBS | Bulbus | – | Monolayer culture | Sangwan et al. 2003 |
| dAM | Rabbit | Suspension | <28 | RCF | Yes/No | No | DMEM:F12 | 5% FBS | – | Yes (scattered MUC5AC+ cells with/without AL and RCF) | Stratified culture (increased in AL) | Meller et al. 1999 |
| AlloDerm coated with COL4 | Human | Suspension | 18 | No | Yes | Yes | MCDB 153 | No | – | – | Stratified culture | Yoshizawa et al. 2004 |
4.2. Extracellular Matrix Protein-Containing Membranes
| Substrate (s) | Cell Species | Normal Cells/Cells Line | Explant/Suspension Culture | Culture Time (Days) | Feeder Cells | Air-Lifting | High Calcium | Basal Medium | Serum | Conjunctival Donor Site | Goblet Cells | Comm ent | Authors |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BSA:COL mix | Rabbit | Normal | Suspension | – | No | No | No | PC-1 (serum free) | No | – | – | Scholz et al. 2002 | |
| COL | Rabbit | Normal | Suspension | 6 | No | Yes/No | No | PC-1 (serum free) or DMEM:F12 | No | – | Yes (3% to 4% PAS + in AL group) | Yang et al. 2000 | |
| COL:FN mix | Human | Normal | Suspension | – | No | No | No | KGM (serum free) | No | Bulbus | – | Monolayer culture | Cook et al. 1998 |
| COL:FN mix | Human | Normal | Suspension | – | No | No | No | EpiLife | No | – | – | Gordan et al. 2005 | |
| COL1 | Bovine | Normal | Suspension | 12 | No | Yes/No | – | DMEM:F12 | 10% FBS | Bulbus | Yes (PAS + in both AL and submerged cultures) | Stratified culture (increased by AL) | Civiale et al. 2003 |
| COL1 gel | Rabbit | Normal | Suspension | 7–14 | No | No | No | DMEM:F12 | 10% FBS | Bulbus | Yes (PAS + cell within gel, PAS− on the gel surface) | Stratified culture within gel, monolayer on the gel surface | Niiya et al. 1997 |
| COL1 gel with/without 3T3 | Rabbit | Normal | Suspension | 6 | 3T3/No | Yes/No | No | DMEM:F12 | 10% FBS | – | No (PAS−, MUC5AC−) | Stratified culture (increased by AL and 3T3) | Chen et al. 1994 |
| COL1 gel with/without 3T3 or HCF | Human | Normal | Suspension | 14 | 3T3/HCF/no | Yes | No | DMEM:F12 | 5% FBS | Bulbus | Yes (only with HCF) | Stratified culture (with feeder cells) | Tsai et al. 1994 |
| COL1 or Matrigel | Human | ConjEp-1/p53DD/cdk4R/TERT cell line | Suspension | (3–7 days in high Ca) | 3T3/no | No | Yes | KM (serum free) or DMEM:F12 | 10% FBS | Bulbus | Yes (MUC5AC RNA highest with COL1) | Stratified culture (COL1), aggregates (Matrigel) | Gipson et al. 2003 |
| COL1, COL1: COL3 mix, LN, FN or Matrigel | Rabbit | Normal | Suspension | <14 | No | No | No | PC-1 (serum free) | 0 or 1% FBS | All conjunctiva | Yes (5% PAS + in serum free cultures on COL1:COL3 mix) | Stratified culture (COL1:COL3 mix) | Saha et al. 1996 |
| COL1, Matrigel or COL1:Matrigel mix | Rabbit | Normal | Suspension | – | No | No | No | DMEM:F12 | 5% FBS | All conjunctiva | Yes (PAS + cell in cultures on COL1 and in globules on Matrigel) | Monolayer culture (COL1), aggregates (Matrigel) | Tsai et al. 1988 |
| COL1:FN mix | Rabbit | Normal | Suspension | – | No | No | – | PC-1 (serum free) | No | – | – | Basu et al. 1998 | |
| COL4 | Rat | Normal | Suspension | 10 | – | Yes | – | KM (serum free) or DMEM:F12 | No | Palpebra | – | Yu et al. 2012 | |
| Compressed COL | Human | Normal | Suspension | 14 | No | No | No | DMEM:F12 | 10% FBS | – | – | Stratified culture | Drechsler et al. 2015 |
| Elastin-like polymer | Human | IOBA-NHC cell line | Suspension | 5 | No | No | – | DMEM:F12 | – | – | Yes (Ck7+) | – | Martinez-Osorio et al. 2009 |
| Gelatin-chitosan | Rabbit | Normal | Explant | 14 | No | No | – | DMEM:F12 | 10% FBS | – | – | Stratified culture (Ck4+) | Zhu et al. 2006 |
| LN-1, LN-β2 or COL1 gel with BCF | Bovine | Normal | Explant | 14 | BCF/no | No | No | KBM (serum free) or DMEM (serum) | 0 or 10% FBS | Bulbus | – | Stratified culture (DMEM/10% FCS and cultures on COL1 with BCF) | Kurpakus et al. 1999 |
| LN-1, LN-β2 or poly-I-Iysine | Bovine | Normal | Suspension | – | No | No | No | KBM (serum free) | No | – | – | Lin et al. 1999 | |
| LN-10 | Human | HC0597 cell line | Suspension | – | No | No | No | KBM (serum free) | No | – | – | Lin et al. 2002 |
4.3. Synthetic Polymer Membranes
| Substrate (s) | Cell Species | Normal Cells/Cells Line | Culture Time (Days) | Explant/Suspension Culture | Feeder Cells | Air-Lifting | High Calcium | Culture Medium | Serum | Conjunctival Donor Site | Goblet Cells | Comment | Authors |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 50:50 PDLGA, 85:15 PDLGA or Inion GTRTM | Human | IOBA-NHC cell line | – | Suspension | No | No | – | DMEM:F12 | 10% FBS | – | – | Extract studies showing lowest to highest viability with 50:50 PDLGA; 85:15 PDLGA; Inion GTRTM | Huhtala, et al. 2008 |
| 50:50 PDLGA, 85:15 PDLGA or Inion GTRTM | Human | IOBA-NHC cell line | 3 | Suspension | No | No | – | DMEM:F12 | 10% FBS | – | – | High viability with all types of polymer | Huhtala, et al. 2007 |
| P(EA-co-HEA) or 90:10 P(EA-co-MAAc) copolymers | Human | IOBA-NHC cell line | – | Suspension | No | – | – | DMEM:F12 | 10% FBS | – | – | All polymers were non-toxic, hydrophobicity increased adhesion, proliferation and viability | Campillo-Fernandez, et al. 2007 |
| Ultrathin PCL | Rabbit | Normal | – | Explant/suspension | No | No | Yes | KGM (serum free) | No | – | Yes (MUC5AC+ comparable to AM) | Stratified culture (increased by NaOH); NaOH surface modification increased hydrophilicity and proliferation | Ang, et al. 2006 |
| Temperature-responsive polymer, poly(N-isopropyl-acrylamide; PIPAAm) | Rabbit | Normal | 10 | Suspension | No | No | No | DMEM:F12 | 10% FBS | Fornix/ palpebra | Yes (21.5% MUC5AC+, PAS+) | Stratified culture (4–5 cell layers); proliferation rate of 38.4%; high viability; Ck4 mRNA increased with time | Yao, et al. 2015 |
5. Future Avenues for Developing Tissue Engineered Conjunctival Epithelial Equivalents
5.1. Comparative Studies of the Effect of Different Substrates on Cultured Conjunctival Epithelial Cells
5.2. Storage and Transportation of Cultured Conjunctival Epithelial Cells
6. Conclusion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wagoner, M.D. Chemical injuries of the eye: Current concepts in pathophysiology and therapy. Surv. Ophthalmol. 1997, 41, 275–313. [Google Scholar] [CrossRef]
- Gipson, I.K.; Joyce, N.; Zieske, J. The Anatomy and Cell Biology of the Human Cornea, Limbus, Conjunctiva, and Adnexa. In The Cornea; Foster, C.A.D., Dohlman, C., Eds.; Lippincott Williams & Wilkens: Philadelphia, PA, USA, 2005; pp. 1–35. [Google Scholar]
- Steven, P.; Gebert, A. Conjunctiva-associated lymphoid tissue—Current knowledge, animal models and experimental prospects. Ophthalmic Res. 2009, 42, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Schrader, S.; Notara, M.; Beaconsfield, M.; Tuft, S.J.; Daniels, J.T.; Geerling, G. Tissue engineering for conjunctival reconstruction: Established methods and future outlooks. Curr. Eye Res. 2009, 34, 913–924. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Thelin, W.R.; Rogers, T.D.; Stutts, M.J.; Randell, S.H.; Grubb, B.R.; Boucher, R.C. Regional differences in rat conjunctival ion transport activities. Am. J. Physiol. Cell Physiol. 2012, 303, C767–C780. [Google Scholar] [CrossRef] [PubMed]
- Gendler, S.J.; Spicer, A.P. Epithelial mucin genes. Annu. Rev. Physiol. 1995, 57, 607–634. [Google Scholar] [CrossRef] [PubMed]
- Fostad, I.G.; Eidet, J.R.; Shatos, M.A.; Utheim, T.P.; Utheim, O.A.; Raeder, S.; Dartt, D.A. Biopsy harvesting site and distance from the explant affect conjunctival epithelial phenotype ex vivo. Exp. Eye Res. 2012, 104, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Dartt, D.A. Control of mucin production by ocular surface epithelial cells. Exp. Eye Res. 2004, 78, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Spurr-Michaud, S.; Argueso, P.; Gipson, I. Assay of mucins in human tear fluid. Exp. Eye Res. 2007, 84, 939–950. [Google Scholar] [CrossRef] [PubMed]
- Argueso, P.; Gipson, I.K. Epithelial mucins of the ocular surface: Structure, biosynthesis and function. Exp. Eye Res. 2001, 73, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Miranda, A.; Nakatsu, M.N.; Zarei-Ghanavati, S.; Nguyen, C.V.; Deng, S.X. Keratin 13 is a more specific marker of conjunctival epithelium than keratin 19. Mol. Vis. 2011, 17, 1652–1661. [Google Scholar] [PubMed]
- Nichols, K.K.; Foulks, G.N.; Bron, A.J.; Glasgow, B.J.; Dogru, M.; Tsubota, K.; Lemp, M.A.; Sullivan, D.A. The international workshop on meibomian gland dysfunction: Executive summary. Invest. Ophthalmol. Vis. Sci. 2011, 52, 1922–1929. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.G.; Lin, T.; Sun, T.T.; Lavker, R.M. Clonal analysis of the in vivo differentiation potential of keratinocytes. Invest. Ophthalmol. Vis. Sci. 1997, 38, 753–761. [Google Scholar] [PubMed]
- Mason, S.L.; Stewart, R.M.; Kearns, V.R.; Williams, R.L.; Sheridan, C.M. Ocular epithelial transplantation: Current uses and future potential. Regen. Med. 2011, 6, 767–782. [Google Scholar] [CrossRef] [PubMed]
- Nagasaki, T.; Zhao, J. Uniform distribution of epithelial stem cells in the bulbar conjunctiva. Invest. Ophthalmol. Vis. Sci. 2005, 46, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.G.; Cotsarelis, G.; Sun, T.T.; Lavker, R.M. Label-retaining cells are preferentially located in fornical epithelium: Implications on conjunctival epithelial homeostasis. Invest. Ophthalmol. Vis. Sci. 1995, 36, 236–246. [Google Scholar] [PubMed]
- Lavker, R.M.; Wei, Z.G.; Sun, T.T. Phorbol ester preferentially stimulates mouse fornical conjunctival and limbal epithelial cells to proliferate in vivo. Invest. Ophthalmol. Vis. Sci. 1998, 39, 301–307. [Google Scholar] [PubMed]
- Pe’er, J.; Zajicek, G.; Greifner, H.; Kogan, M. Streaming conjunctiva. Anat. Rec. 1996, 245, 36–40. [Google Scholar] [CrossRef]
- Chen, W.; Ishikawa, M.; Yamaki, K.; Sakuragi, S. Wistar rat palpebral conjunctiva contains more slow-cycling stem cells that have larger proliferative capacity: Implication for conjunctival epithelial homeostasis. Jpn. J. Ophthalmol. 2003, 47, 119–128. [Google Scholar] [CrossRef]
- Wei, Z.G.; Wu, R.L.; Lavker, R.M.; Sun, T.T. In vitro growth and differentiation of rabbit bulbar, fornix, and palpebral conjunctival epithelia. Implications on conjunctival epithelial transdifferentiation and stem cells. Invest. Ophthalmol. Vis. Sci. 1993, 34, 1814–1828. [Google Scholar] [PubMed]
- Wirtschafter, J.D.; Ketcham, J.M.; Weinstock, R.J.; Tabesh, T.; McLoon, L.K. Mucocutaneous junction as the major source of replacement palpebral conjunctival epithelial cells. Invest. Ophthalmol. Vis. Sci. 1999, 40, 3138–3146. [Google Scholar] [PubMed]
- Pellegrini, G.; Golisano, O.; Paterna, P.; Lambiase, A.; Bonini, S.; Rama, P.; de Luca, M. Location and clonal analysis of stem cells and their differentiated progeny in the human ocular surface. J. Cell Biol. 1999, 145, 769–782. [Google Scholar] [CrossRef] [PubMed]
- Qi, H.; Zheng, X.; Yuan, X.; Pflugfelder, S.C.; Li, D.Q. Potential localization of putative stem/progenitor cells in human bulbar conjunctival epithelium. J. Cell Physiol. 2010, 225, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Stewart, R.M.; Sheridan, C.M.; Hiscott, P.S.; Czanner, G.; Kaye, S.B. Human conjunctival stem cells are predominantly located in the medial canthal and inferior forniceal areas. Invest. Ophthalmol. Vis. Sci. 2015, 56, 2021–2030. [Google Scholar] [CrossRef] [PubMed]
- Eidet, J.R.; Fostad, I.G.; Shatos, M.A.; Utheim, T.P.; Utheim, O.A.; Raeder, S.; Dartt, D.A. Effect of biopsy location and size on proliferative capacity of ex vivo expanded conjunctival tissue. Invest. Ophthalmol. Vis. Sci. 2012, 53, 2897–2903. [Google Scholar] [CrossRef] [PubMed]
- Ramos, T.; Scott, D.; Ahmad, S. An update on ocular surface epithelial stem cells: Cornea and conjunctiva. Stem Cells Int. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Schofield, R. The stem cell system. Biomed. Pharmacother. Biomed. Pharmacother. 1983, 37, 375–380. [Google Scholar] [PubMed]
- Watt, F.M.; Hogan, B.L. Out of eden: Stem cells and their niches. Science 2000, 287, 1427–1430. [Google Scholar] [CrossRef] [PubMed]
- Schlotzer-Schrehardt, U.; Dietrich, T.; Saito, K.; Sorokin, L.; Sasaki, T.; Paulsson, M.; Kruse, F.E. Characterization of extracellular matrix components in the limbal epithelial stem cell compartment. Exp. Eye Res. 2007, 85, 845–860. [Google Scholar] [CrossRef] [PubMed]
- Radford, C.F.; Rauz, S.; Williams, G.P.; Saw, V.P.; Dart, J.K. Incidence, presenting features, and diagnosis of cicatrising conjunctivitis in the United Kingdom. Eye 2012, 26, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Sobolewska, B.; Deuter, C.; Zierhut, M. Current medical treatment of ocular mucous membrane pemphigoid. Ocul. Surf. 2013, 11, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Solomon, A.; Espana, E.M.; Tseng, S.C. Amniotic membrane transplantation for reconstruction of the conjunctival fornices. Ophthalmology 2003, 110, 93–100. [Google Scholar] [CrossRef]
- Patel, B.C.; Sapp, N.A.; Collin, R. Standardized range of conformers and symblepharon rings. Ophthalmic Plast. Reconstr. Surg. 1998, 14, 144–145. [Google Scholar] [CrossRef]
- Fein, W. Repair of total and subtotal symblepharons. Ophthalmic Surg. 1979, 10, 44–47. [Google Scholar] [PubMed]
- Donnenfeld, E.D.; Perry, H.D.; Wallerstein, A.; Caronia, R.M.; Kanellopoulos, A.J.; Sforza, P.D.; D’Aversa, G. Subconjunctival mitomycin C for the treatment of ocular cicatricial pemphigoid. Ophthalmology 1999, 106, 72–79. [Google Scholar] [CrossRef]
- Thoft, R.A. Conjunctival transplantation. Arch. Ophthalmol. 1977, 95, 1425–1427. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Inatomi, T.; Cooper, L.J.; Rigby, H.; Fullwood, N.J.; Kinoshita, S. Phenotypic investigation of human eyes with transplanted autologous cultivated oral mucosal epithelial sheets for severe ocular surface diseases. Ophthalmology 2007, 114, 1080–1088. [Google Scholar] [CrossRef] [PubMed]
- Burillon, C.; Huot, L.; Justin, V.; Nataf, S.; Chapuis, F.; Decullier, E.; Damour, O. Cultured autologous oral mucosal epithelial cell sheet (caomecs) transplantation for the treatment of corneal limbal epithelial stem cell deficiency. Invest. Ophthalmol. Vis. Sci. 2012, 53, 1325–1331. [Google Scholar] [CrossRef] [PubMed]
- Homma, R.; Yoshikawa, H.; Takeno, M.; Kurokawa, M.S.; Masuda, C.; Takada, E.; Tsubota, K.; Ueno, S.; Suzuki, N. Induction of epithelial progenitors in vitro from mouse embryonic stem cells and application for reconstruction of damaged cornea in mice. Invest. Ophthalmol. Vis. Sci. 2004, 45, 4320–4326. [Google Scholar] [CrossRef] [PubMed]
- Reinshagen, H.; Auw-Haedrich, C.; Sorg, R.V.; Boehringer, D.; Eberwein, P.; Schwartzkopff, J.; Sundmacher, R.; Reinhard, T. Corneal surface reconstruction using adult mesenchymal stem cells in experimental limbal stem cell deficiency in rabbits. Acta Ophthalmol. 2011, 89, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Gomes, J.A.; Geraldes Monteiro, B.; Melo, G.B.; Smith, R.L.; Cavenaghi Pereira da Silva, M.; Lizier, N.F.; Kerkis, A.; Cerruti, H.; Kerkis, I. Corneal reconstruction with tissue-engineered cell sheets composed of human immature dental pulp stem cells. Invest. Ophthalmol. Vis. Sci. 2010, 51, 1408–1414. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Blazejewska, E.A.; Call, M.K.; Yamanaka, O.; Liu, H.; Schlotzer-Schrehardt, U.; Kruse, F.E.; Kao, W.W. From hair to cornea: Toward the therapeutic use of hair follicle-derived stem cells in the treatment of limbal stem cell deficiency. Stem Cells 2011, 29, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Reza, H.M.; Ng, B.Y.; Gimeno, F.L.; Phan, T.T.; Ang, L.P. Umbilical cord lining stem cells as a novel and promising source for ocular surface regeneration. Stem Cell Rev. 2011, 7, 935–947. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Zhang, X.; Huang, D.; Huang, B.; Gao, N.; Wang, Z.; Ge, J. Conjunctival reconstruction with progenitor cell-derived autologous epidermal sheets in rhesus monkey. PLoS One 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.P.; Yang, X.Z.; Cao, G.P. Conjunctiva reconstruction by induced differentiation of human amniotic epithelial cells. Genet. Mol. Res. 2015, 14, 13823–13834. [Google Scholar] [CrossRef] [PubMed]
- Wenkel, H.; Rummelt, V.; Naumann, G.O. Long term results after autologous nasal mucosal transplantation in severe mucus deficiency syndromes. Br. J. Ophthalmol. 2000, 84, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Tseng, S.C. Amniotic membrane transplantation for ocular surface reconstruction. Biosci. Rep. 2001, 21, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Touhami, A.; Grueterich, M.; Tseng, S.C. The role of NGF signaling in human limbal epithelium expanded by amniotic membrane culture. Invest. Ophthalmol. Vis. Sci. 2002, 43, 987–994. [Google Scholar] [PubMed]
- Lee, S.B.; Li, D.Q.; Tan, D.T.; Meller, D.C.; Tseng, S.C. Suppression of TGF-β signaling in both normal conjunctival fibroblasts and pterygial body fibroblasts by amniotic membrane. Curr. Eye Res. 2000, 20, 325–334. [Google Scholar] [CrossRef]
- Ueta, M.; Kweon, M.N.; Sano, Y.; Sotozono, C.; Yamada, J.; Koizumi, N.; Kiyono, H.; Kinoshita, S. Immunosuppressive properties of human amniotic membrane for mixed lymphocyte reaction. Clin. Exp. Immunol. 2002, 129, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Solomon, A.; Rosenblatt, M.; Monroy, D.; Ji, Z.; Pflugfelder, S.C.; Tseng, S.C. Suppression of interleukin 1α and interleukin 1β in human limbal epithelial cells cultured on the amniotic membrane stromal matrix. Br. J. Ophthalmol. 2001, 85, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Kruse, F.E.; Joussen, A.M.; Rohrschneider, K.; You, L.; Sinn, B.; Baumann, J.; Volcker, H.E. Cryopreserved human amniotic membrane for ocular surface reconstruction. Graefe’s Arch. Clin. Exp. Ophthalmol. 2000, 238, 68–75. [Google Scholar] [CrossRef]
- Lee, S.H.; Tseng, S.C. Amniotic membrane transplantation for persistent epithelial defects with ulceration. Am. J. Ophthalmol. 1997, 123, 303–312. [Google Scholar] [CrossRef]
- Nakamura, T.; Yoshitani, M.; Rigby, H.; Fullwood, N.J.; Ito, W.; Inatomi, T.; Sotozono, C.; Nakamura, T.; Shimizu, Y.; Kinoshita, S. Sterilized, freeze-dried amniotic membrane: A useful substrate for ocular surface reconstruction. Invest. Ophthalmol. Vis. Sci. 2004, 45, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Russo, A.; Bonci, P.; Bonci, P. The effects of different preservation processes on the total protein and growth factor content in a new biological product developed from human amniotic membrane. Cell. Tissue Banking 2012, 13, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Von Versen-Hoeynck, F.; Steinfeld, A.P.; Becker, J.; Hermel, M.; Rath, W.; Hesselbarth, U. Sterilization and preservation influence the biophysical properties of human amnion grafts. Biol. J. Int. Assoc. Biol. Standard. 2008, 36, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Thomasen, H.; Pauklin, M.; Steuhl, K.P.; Meller, D. Comparison of cryopreserved and air-dried human amniotic membrane for ophthalmologic applications. Graefe’s Arch. Clin. Exp. Ophthalmol. 2009, 247, 1691–1700. [Google Scholar] [CrossRef] [PubMed]
- Henderson, H.W.; Collin, J.R. Mucous membrane grafting. Dev. Ophthalmol. 2008, 41, 230–242. [Google Scholar] [PubMed]
- Meller, D.; Tseng, S.C. Conjunctival epithelial cell differentiation on amniotic membrane. Invest. Ophthalmol. Vis. Sci. 1999, 40, 878–886. [Google Scholar] [PubMed]
- Eidet, J.R.; Utheim, O.A.; Raeder, S.; Dartt, D.A.; Lyberg, T.; Carreras, E.; Huynh, T.T.; Messelt, E.B.; Louch, W.E.; Roald, B.; et al. Effects of serum-free storage on morphology, phenotype, and viability of ex vivo cultured human conjunctival epithelium. Exp. Eye Res. 2012, 94, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Osorio, H.; Calonge, M.; Corell, A.; Reinoso, R.; Lopez, A.; Fernandez, I.; San Jose, E.G.; Diebold, Y. Characterization and short-term culture of cells recovered from human conjunctival epithelium by minimally invasive means. Mol. Vis. 2009, 15, 2185–2195. [Google Scholar] [PubMed]
- Fuchs, E.; Green, H. Regulation of terminal differentiation of cultured human keratinocytes by vitamin A. Cell 1981, 25, 617–625. [Google Scholar] [CrossRef]
- Sangwan, V.S.; Vemuganti, G.K.; Iftekhar, G.; Bansal, A.K.; Rao, G.N. Use of autologous cultured limbal and conjunctival epithelium in a patient with severe bilateral ocular surface disease induced by acid injury: A case report of unique application. Cornea 2003, 22, 478–481. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Qiu, F.; Qu, Y.L.; Li, C.; Shao, Y.; Xiao, Q.; Liu, Z.; Li, W. Amniotic membrane inhibits squamous metaplasia of human conjunctival epithelium. Am. J. Physiol. Cell Physiol. 2011, 301, C115–C125. [Google Scholar] [CrossRef] [PubMed]
- Yoshizawa, M.; Feinberg, S.E.; Marcelo, C.L.; Elner, V.M. Ex vivo produced human conjunctiva and oral mucosa equivalents grown in a serum-free culture system. J. Oral Maxillofac. Surg. 2004, 62, 980–988. [Google Scholar] [CrossRef] [PubMed]
- Tsai, R.J.; Tseng, S.C. Substrate modulation of cultured rabbit conjunctival epithelial cell differentiation and morphology. Invest. Ophthalmol. Vis. Sci. 1988, 29, 1565–1576. [Google Scholar] [PubMed]
- Chen, W.Y.; Mui, M.M.; Kao, W.W.; Liu, C.Y.; Tseng, S.C. Conjunctival epithelial cells do not transdifferentiate in organotypic cultures: Expression of K12 keratin is restricted to corneal epithelium. Curr. Eye Res. 1994, 13, 765–778. [Google Scholar] [CrossRef] [PubMed]
- Drechsler, C.C.; Kunze, A.; Kureshi, A.; Grobe, G.; Reichl, S.; Geerling, G.; Daniels, J.T.; Schrader, S. Development of a conjunctival tissue substitute on the basis of plastic compressed collagen. J. Tissue Eng. Regen. Med. 2015. [Google Scholar] [CrossRef] [PubMed]
- Gordon, Y.J.; Huang, L.C.; Romanowski, E.G.; Yates, K.A.; Proske, R.J.; McDermott, A.M. Human cathelicidin (LL-37), a multifunctional peptide, is expressed by ocular surface epithelia and has potent antibacterial and antiviral activity. Curr. Eye Res. 2005, 30, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Cook, E.B.; Stahl, J.L.; Miller, S.T.; Gern, J.E.; Sukow, K.A.; Graziano, F.M.; Barney, N.P. Isolation of human conjunctival mast cells and epithelial cells: Tumor necrosis factor-alpha from mast cells affects intercellular adhesion molecule 1 expression on epithelial cells. Invest. Ophthalmol. Vis. Sci. 1998, 39, 336–343. [Google Scholar] [PubMed]
- Saha, P.; Kim, K.J.; Lee, V.H. A primary culture model of rabbit conjunctival epithelial cells exhibiting tight barrier properties. Curr. Eye Res. 1996, 15, 1163–1169. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.K.; Haworth, I.S.; Bolger, M.B.; Lee, V.H. Proton-driven dipeptide uptake in primary cultured rabbit conjunctival epithelial cells. Invest. Ophthalmol. Vis. Sci. 1998, 39, 2365–2373. [Google Scholar] [PubMed]
- Kurpakus, M.A.; Lin, L. The lack of extracellular laminin β2 chain deposition correlates to the loss of conjunctival epithelial keratin K4 localization in culture. Curr. Eye Res. 1999, 18, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Kurpakus Wheater, M. Differential rapid adhesion of bovine ocular surface epithelial cells to laminin isoforms. Curr. Eye Res. 1999, 19, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Kurpakus-Wheater, M. Laminin α5 chain adhesion and signaling in conjunctival epithelial cells. Invest. Ophthalmol. Vis. Sci. 2002, 43, 2615–2621. [Google Scholar] [PubMed]
- Gipson, I.K.; Spurr-Michaud, S.; Argueso, P.; Tisdale, A.; Ng, T.F.; Russo, C.L. Mucin gene expression in immortalized human corneal-limbal and conjunctival epithelial cell lines. Invest. Ophthalmol. Vis. Sci. 2003, 44, 2496–2506. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Beuerman, R.W.; Chan-Park, M.B.; Cheng, Z.; Ang, L.P.; Tan, D.T. Enhancement of the mechanical and biological properties of a biomembrane for tissue engineering the ocular surface. Ann. Acad. Med. Singapore 2006, 35, 210–214. [Google Scholar] [PubMed]
- Niiya, A.; Matsumoto, Y.; Ishibashi, T.; Matsumoto, K.; Kinoshita, S. Collagen gel-embedding culture of conjunctival epithelial cells. Graefe’s Arch. Clin. Exp. Ophthalmol. 1997, 235, 32–40. [Google Scholar] [CrossRef]
- Martinez-Osorio, H.; Juarez-Campo, M.; Diebold, Y.; Girotti, A.; Alonso, M.; Arias, F.J.; Rodriguez-Cabello, J.C.; Garcia-Vazquez, C.; Calonge, M. Genetically engineered elastin-like polymer as a substratum to culture cells from the ocular surface. Curr. Eye Res. 2009, 34, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Ang, L.P.; Cheng, Z.Y.; Beuerman, R.W.; Teoh, S.H.; Zhu, X.; Tan, D.T. The development of a serum-free derived bioengineered conjunctival epithelial equivalent using an ultrathin poly(epsilon-caprolactone) membrane substrate. Invest. Ophthalmol. Vis. Sci. 2006, 47, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Campillo-Fernandez, A.J.; Pastor, S.; Abad-Collado, M.; Bataille, L.; Gomez-Ribelles, J.L.; Meseguer-Duenas, J.M.; Monleon-Pradas, M.; Artola, A.; Alio, J.L.; Ruiz-Moreno, J.M. Future design of a new keratoprosthesis. Physical and biological analysis of polymeric substrates for epithelial cell growth. Biomacromolecules 2007, 8, 2429–2436. [Google Scholar] [CrossRef] [PubMed]
- Huhtala, A.; Pohjonen, T.; Salminen, L.; Salminen, A.; Kaarniranta, K.; Uusitalo, H. In vitro biocompatibility of degradable biopolymers in cell line cultures from various ocular tissues: Direct contact studies. J. Biomed. Mater. Res. Part A 2007, 83, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Huhtala, A.; Pohjonen, T.; Salminen, L.; Salminen, A.; Kaarniranta, K.; Uusitalo, H. In vitro biocompatibility of degradable biopolymers in cell line cultures from various ocular tissues: Extraction studies. J. Mater. Sci. Mater. Med. 2008, 19, 645–649. [Google Scholar] [CrossRef] [PubMed]
- Rama, P.; Matuska, S.; Paganoni, G.; Spinelli, A.; de Luca, M.; Pellegrini, G. Limbal stem-cell therapy and long-term corneal regeneration. N Engl. J. Med. 2010, 363, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Daniels, J.T.; Secker, G.A.; Shortt, A.J.; Tuft, S.J.; Seetharaman, S. Stem cell therapy delivery: Treading the regulatory tightrope. Regen. Med. 2006, 1, 715–719. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Osei-Bempong, C.; Dana, R.; Jurkunas, U. The culture and transplantation of human limbal stem cells. J. Cell Physiol. 2010, 225, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Utheim, T.P.; Raeder, S.; Utheim, O.A.; de la Paz, M.; Roald, B.; Lyberg, T. Sterility control and long-term eye-bank storage of cultured human limbal epithelial cells for transplantation. Br. J. Ophthalmol. 2009, 93, 980–983. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, A.R.; Daniels, J.T. Concise review: Limbal epithelial stem cell therapy: Controversies and challenges. Stem Cells 2011, 29, 1923–1932. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eidet, J.R.; Dartt, D.A.; Utheim, T.P. Concise Review: Comparison of Culture Membranes Used for Tissue Engineered Conjunctival Epithelial Equivalents. J. Funct. Biomater. 2015, 6, 1064-1084. https://doi.org/10.3390/jfb6041064
Eidet JR, Dartt DA, Utheim TP. Concise Review: Comparison of Culture Membranes Used for Tissue Engineered Conjunctival Epithelial Equivalents. Journal of Functional Biomaterials. 2015; 6(4):1064-1084. https://doi.org/10.3390/jfb6041064
Chicago/Turabian StyleEidet, Jon Roger, Darlene A. Dartt, and Tor Paaske Utheim. 2015. "Concise Review: Comparison of Culture Membranes Used for Tissue Engineered Conjunctival Epithelial Equivalents" Journal of Functional Biomaterials 6, no. 4: 1064-1084. https://doi.org/10.3390/jfb6041064