
Unraveling the Specific Ischemic Core and Penumbra Transcriptome in the Permanent Middle Cerebral Artery Occlusion Mouse Model Brain Treated with the Neuropeptide PACAP38

Abstract
:1. Introduction
2. Results and Discussion
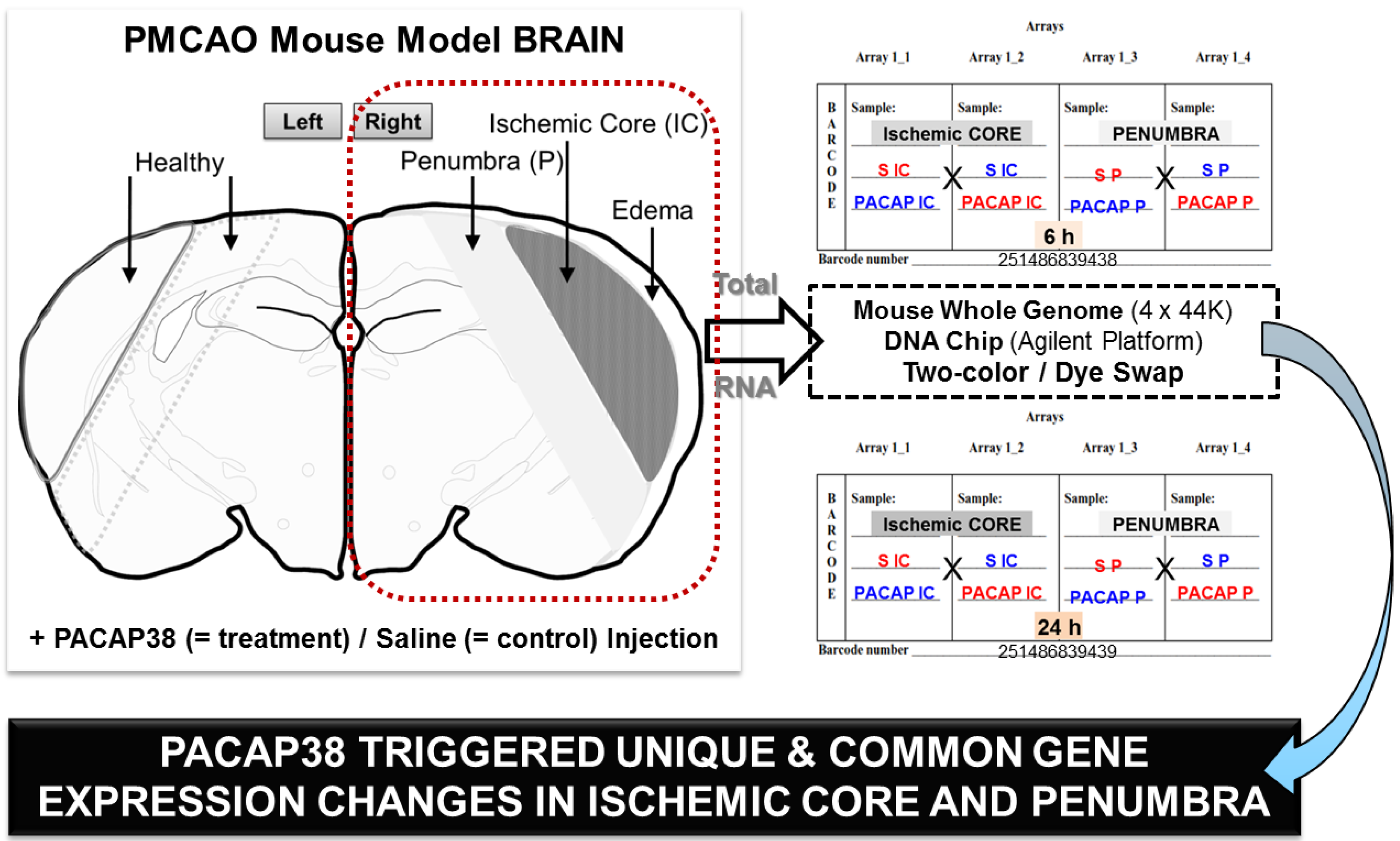
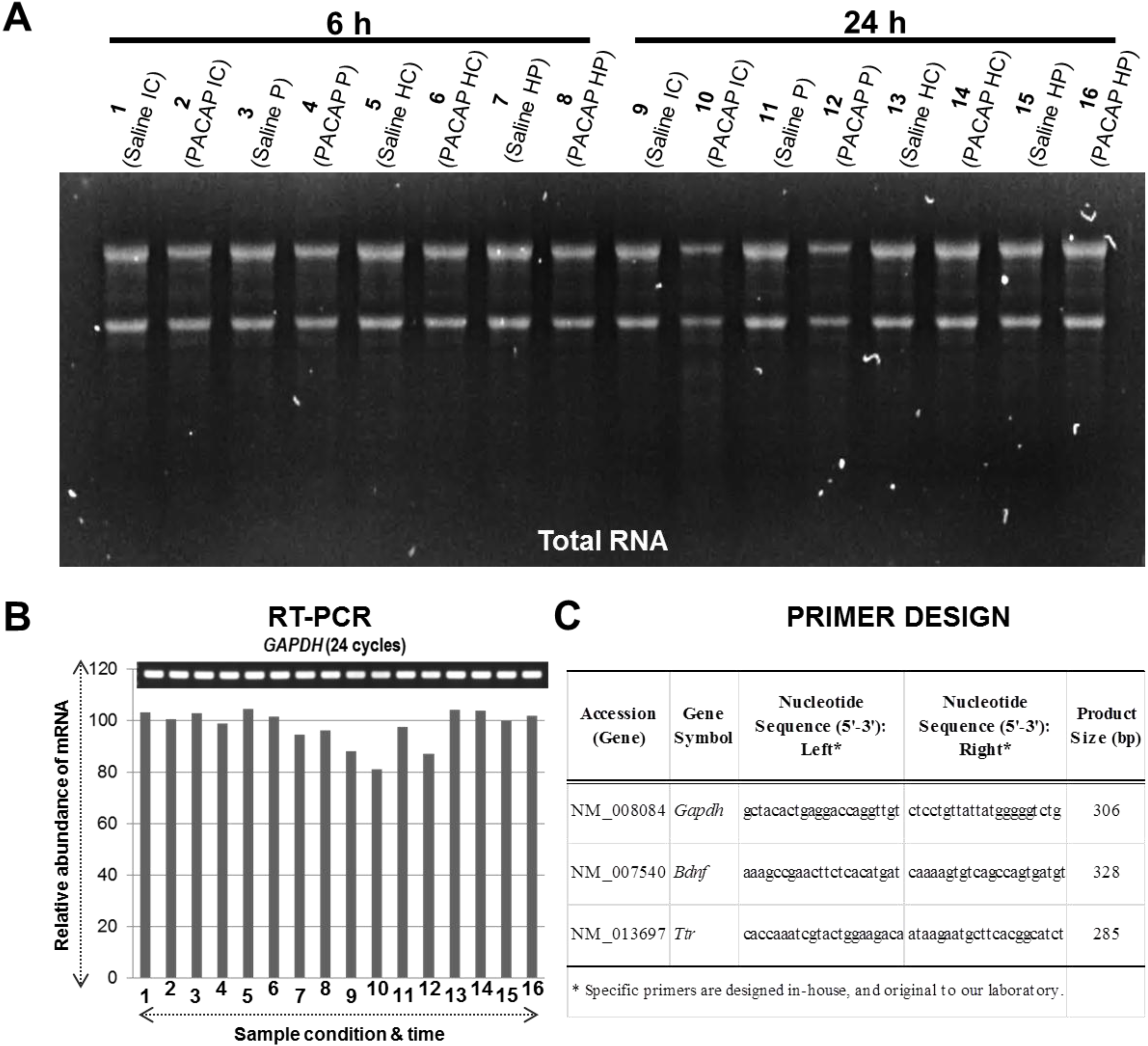
2.1. Experimental Design for Targeted Brain Region Dissection of Ischemic Core (IC) and Penumbra (P), Total RNA Extraction and Quality Check, cDNA Synthesis and RT-PCR, and DNA Microarray Analysis


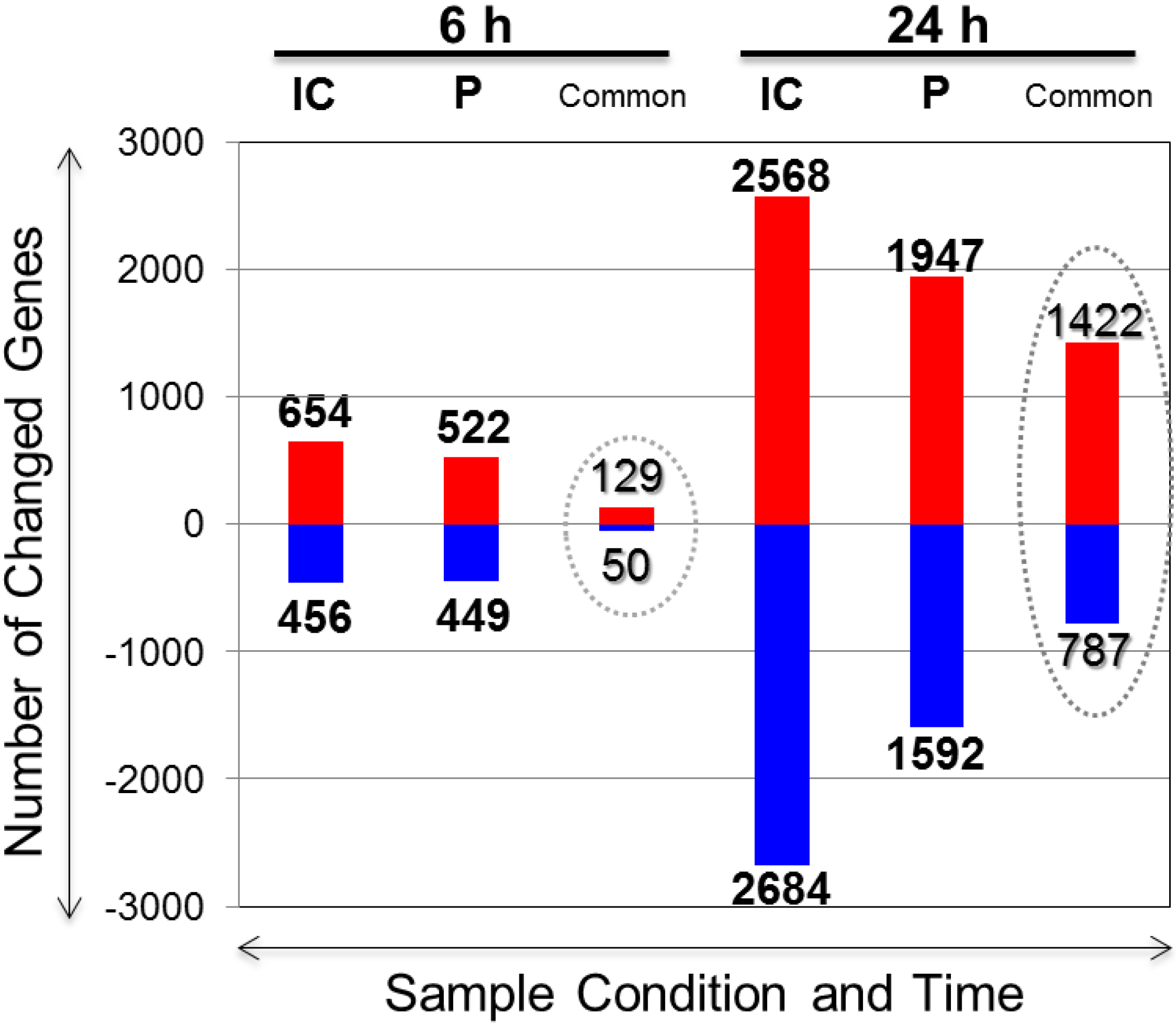
2.2. Overview of the Ischemic Core (IC) and Penumbra (P) Differentially Expressed Genes upon PACAP38 Treatment

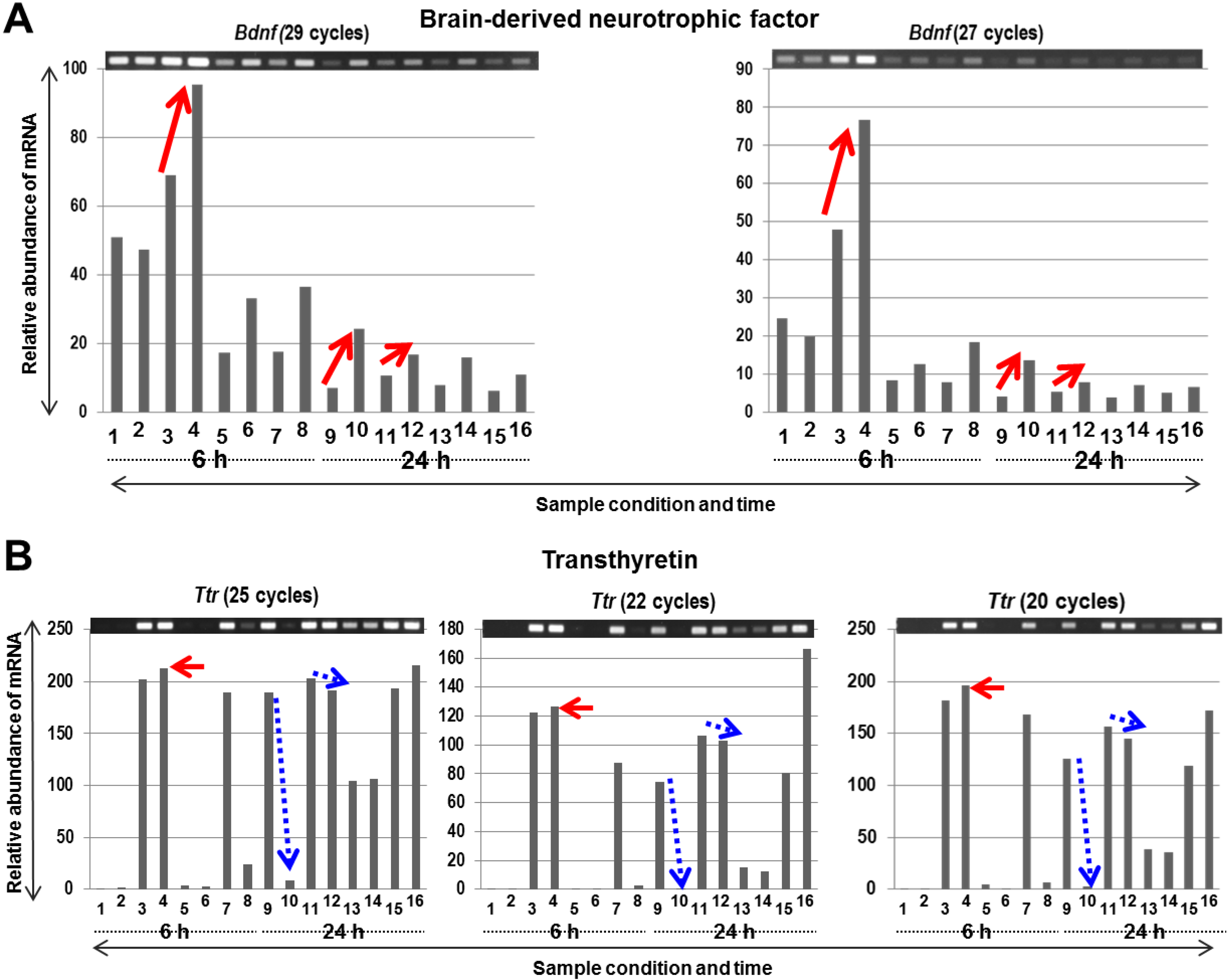
2.3. Identification of Two PACAP38 Induced Genes, Brain-Derived Neurotrophic Factor (Bdnf) and Transthyretin (Ttr) in the Ischemic Core (IC) and Penumbra (P) and Their Validation by RT-PCR as Examples

2.4. Functional Categorization/Biological Function of the Differentially Expressed Genes by PACAP38 Treatment in Ischemic Core (IC) and Penumbra (P) Regions Reveals Activation of Neuroprotective Mechanisms
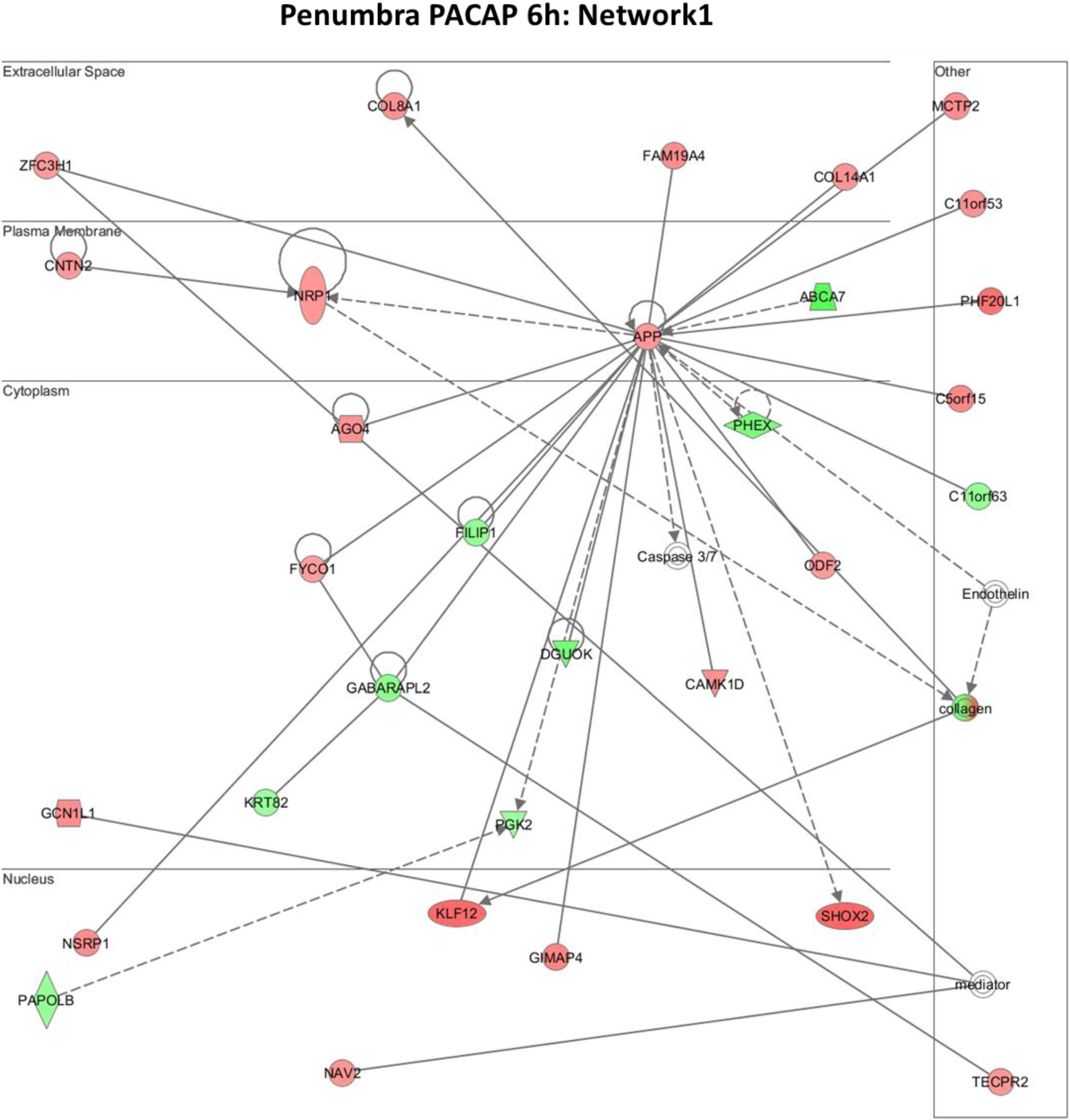
2.5. Biological Functions of PACAP38 Affected Genes by Ingenuity Pathway Analysis (IPA) Reveals Novel Molecular Networks Operating within the Ischemic Brain

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Entrez Gene Name | Agilent | Fold Change | Networks | Location | Type(s) |
|---|---|---|---|---|---|---|
| COMMD7 | COMM domain containing 7 | A_51_P459550 | −2.27 | 1 | Other | other |
| MROH8 | maestro heat-like repeat family member 8 | A_52_P263962 | −2.13 | 1 | Other | other |
| DDX43 | DEAD (Asp-Glu-Ala-Asp) box polypeptide 43 | A_52_P98287 | −2.00 | 1 | Other | enzyme |
| DRAM1 | DNA-damage regulated autophagy modulator 1 | A_51_P481482 | −1.85 | 1 | Cytoplasm | other |
| HSPB7 | heat shock 27kDa protein family, member 7 (cardiovascular) | A_51_P346445 | −1.79 | 1 | Cytoplasm | other |
| DNAJB1 | DnaJ (Hsp40) homolog, subfamily B, member 1 | A_51_P153486 | −1.61 | 1 | Nucleus | other |
| FANCM | Fanconi anemia, complementation group M | A_51_P476030 | −1.61 | 1 | Nucleus | enzyme |
| C1S | complement component 1, s subcomponent | A_52_P39505 | −1.59 | 1 | Extracellular Space | peptidase |
| KCNE3 | potassium voltage-gated channel, Isk-related family, member 3 | A_51_P336599 | −1.59 | 1 | Plasma Membrane | ion channel |
| GOLM1 | golgi membrane protein 1 | A_51_P171200 | −1.52 | 1 | Cytoplasm | other |
| ZC3HAV1L | zinc finger CCCH-type, antiviral 1-like | A_52_P260747 | −1.52 | 1 | Other | other |
| INTS12 | integrator complex subunit 12 | A_52_P1155474 | −1.47 | 1 | Nucleus | other |
| APP | amyloid beta (A4) precursor protein | A_52_P381311 | 1.44 | 1 | Plasma Membrane | other |
| BRWD1 | bromodomain and WD repeat domain containing 1 | A_52_P298237 | 1.44 | 1 | Nucleus | transcription regulator |
| APEX2 | APEX nuclease (apurinic/apyrimidinic endonuclease) 2 | A_52_P417148 | 1.50 | 1 | Nucleus | enzyme |
| INTS3 | integrator complex subunit 3 | A_52_P228684 | 1.50 | 1 | Nucleus | other |
| ARFGAP1 | ADP-ribosylation factor GTPase activating protein 1 | A_52_P207361 | 1.54 | 1 | Cytoplasm | transporter |
| FYCO1 | FYVE and coiled-coil domain containing 1 | A_52_P265666 | 1.59 | 1 | Cytoplasm | other |
| HEATR5A | HEAT repeat containing 5A | A_52_P601569 | 1.59 | 1 | Other | other |
| CAMK1D | calcium/calmodulin-dependent protein kinase ID | A_52_P804224 | 1.63 | 1 | Cytoplasm | kinase |
| PRICKLE2 | prickle homolog 2 (Drosophila) | A_52_P201482 | 1.63 | 1 | Nucleus | other |
| ULK2 | unc-51 like autophagy activating kinase 2 | A_52_P226137 | 1.63 | 1 | Cytoplasm | kinase |
| ANKS6 | ankyrin repeat and sterile alpha motif domain containing 6 | A_51_P298933 | 1.69 | 1 | Cytoplasm | other |
| FBXL20 | F-box and leucine-rich repeat protein 20 | A_52_P35477 | 1.72 | 1 | Cytoplasm | other |
| PHF8 | PHD finger protein 8 | A_51_P117369 | 1.72 | 1 | Nucleus | enzyme |
| FCHO1 | FCH domain only 1 | A_51_P228777 | 1.77 | 1 | Plasma Membrane | other |
| LMAN1 | lectin, mannose-binding, 1 | A_51_P264984 | 1.77 | 1 | Cytoplasm | other |
| ODF2 | outer dense fiber of sperm tails 2 | A_51_P113162 | 1.85 | 1 | Cytoplasm | other |
| ANKS4B | ankyrin repeat and sterile alpha motif domain containing 4B | A_51_P318618 | 1.86 | 1 | Nucleus | transcription regulator |
| ARCN1 | archain 1 | A_52_P1004491 | 2.03 | 1 | Cytoplasm | other |
| ORC3 | origin recognition complex, subunit 3 | A_52_P577438 | 2.04 | 1 | Nucleus | other |
| TBX22 | T-box 22 | A_52_P39481 | 2.19 | 1 | Nucleus | transcription regulator |
| ADAD2 | adenosine deaminase domain containing 2 | A_51_P140042 | 2.43 | 1 | Other | other |
| C1q | -- | -- | 1 | Plasma Membrane | complex |
| Symbol | Entrez Gene Name | Agilent | Fold Change | Networks | Location | Type(s) |
|---|---|---|---|---|---|---|
| ABCA7 | ATP-binding cassette, sub-family A (ABC1), member 7 | A_52_P4928 | −2.56 | 1 | Plasma Membrane | transporter |
| DGUOK | deoxyguanosine kinase | A_51_P385237 | −2.08 | 1 | Cytoplasm | kinase |
| PHEX | phosphate regulating endopeptidase homolog, X-linked | A_51_P468249 | −2.04 | 1 | Cytoplasm | peptidase |
| GABARAPL2 | GABA(A) receptor-associated protein-like 2 | A_52_P521475 | −1.75 | 1 | Cytoplasm | other |
| FILIP1 | filamin A interacting protein 1 | A_51_P333438 | −1.67 | 1 | Cytoplasm | other |
| C11orf63 | chromosome 11 open reading frame 63 | A_51_P258473 | −1.61 | 1 | Other | other |
| PAPOLB | poly(A) polymerase beta (testis specific) | A_51_P310333 | −1.59 | 1 | Nucleus | enzyme |
| KRT82 | keratin 82 | A_51_P239367 | −1.54 | 1 | Cytoplasm | other |
| PGK2 | phosphoglycerate kinase 2 , PGK-2 | A_51_P125487 | −1.52 | 1 | Cytoplasm | kinase |
| FYCO1 | FYVE and coiled-coil domain containing 1 | A_52_P265666 | 1.44 | 1 | Cytoplasm | other |
| ODF2 | outer dense fiber of sperm tails 2 | A_51_P113162 | 1.49 | 1 | Cytoplasm | other |
| ZFC3H1 | zinc finger, C3H1-type containing | A_52_P122393 | 1.50 | 1 | Extracellular Space | other |
| CNTN2 | contactin 2 (axonal) | A_52_P651870 | 1.52 | 1 | Plasma Membrane | other |
| APP | amyloid beta (A4) precursor protein | A_52_P110982 | 1.55 | 1 | Plasma Membrane | other |
| NRP1 | neuropilin 1 | A_51_P469285 | 1.55 | 1 | Plasma Membrane | transmembrane receptor |
| TECPR2 | tectonin beta-propeller repeat containing 2 | A_51_P130282 | 1.56 | 1 | Other | other |
| NAV2 | neuron navigator 2 | A_52_P551829 | 1.57 | 1 | Nucleus | other |
| COL14A1 | collagen, type XIV, alpha 1 | A_51_P141467 | 1.59 | 1 | Extracellular Space | other |
| AGO4 | argonaute RISC catalytic component 4 | A_52_P17422 | 1.61 | 1 | Cytoplasm | translation regulator |
| C11orf53 | chromosome 11 open reading frame 53 | A_51_P173107 | 1.62 | 1 | Other | other |
| CAMK1D | calcium/calmodulin-dependent protein kinase ID | A_52_P804224 | 1.65 | 1 | Cytoplasm | kinase |
| COL8A1 | collagen, type VIII, alpha 1 | A_52_P282058 | 1.65 | 1 | Extracellular Space | other |
| GCN1L1 | GCN1 general control of amino-acid synthesis 1-like 1 (yeast) | A_52_P285100 | 1.67 | 1 | Cytoplasm | translation regulator |
| FAM19A4 | family with sequence similarity 19 (chemokine (C-C motif)-like), member A4 | A_52_P54770 | 1.69 | 1 | Extracellular Space | other |
| MCTP2 | multiple C2 domains, transmembrane 2 | A_51_P154913 | 1.70 | 1 | Other | other |
| NSRP1 (=Ccdc55) | nuclear speckle splicing regulatory protein 1 | A_52_P172798 | 1.71 | 1 | Nucleus | other |
| C5orf15 | chromosome 5 open reading frame 15 | A_52_P33041 | 1.78 | 1 | Other | other |
| GIMAP4 | GTPase, IMAP family member 4 | A_52_P205001 | 1.83 | 1 | Nucleus | other |
| PHF20L1 | PHD finger protein 20-like 1 | A_52_P180283 | 2.10 | 1 | Other | other |
| KLF12 | Kruppel-like factor 12 | A_51_P118877 | 2.23 | 1 | Nucleus | transcription regulator |
| SHOX2 | short stature homeobox 2 | A_52_P356698 | 2.28 | 1 | Nucleus | transcription regulator |
| Caspase 3/7 | -- | -- | 1 | Cytoplasm | group | |
| collagen | -- | -- | 1 | Other | group | |
| Endothelin | -- | -- | 1 | Other | group | |
| mediator | -- | -- | 1 | Other | complex |


3. Concluding Remarks
4. Materials and Methods
4.1. Animals and Husbandry
4.2. Permanent Middle Cerebral Artery Occlusion (PMCAO), PACAP38 and Saline Treatments
4.3. Dissection of Brain, Sampling, and Storage
4.4. Total RNA Extraction, Synthesis of cDNA, and Reverse Transcription-Polymerase Chain Reaction
4.5. DNA Microarray Analysis
4.6. Access to Gene Array Data
4.7. Functional Categorization
4.8. Ingenuity Pathway Analysis (IPA)
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gusev, E.I.; Skvortsova, V.I. Brain Ischemia; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2003. [Google Scholar]
- Kimura, C.; Ohkubo, S.; Ogi, K.; Hosoya, M.; Itoh, Y.; Onda, H.; Miyata, A.; Jian, L.; Dahl, R.R.; Stibbs, H.H.; et al. A novel peptide which stimulates adenylate cyclase: molecular cloning and characterization of the ovine and human cDNAs. Biochem. Biophys. Res. Commun. 1990, 166, 81–89. [Google Scholar] [CrossRef]
- Arimura, A. Perspectives on pituitary adenylate cyclase activating polypeptide (PACAP) in the neuroendocrine, endocrine, and nervous systems. Jpn. J. Physiol. 1998, 48, 301–331. [Google Scholar] [CrossRef] [PubMed]
- Shioda, S.; Ozawa, H.; Dohi, K.; Mizushima, H.; Matsumoto, K.; Nakajo, S.; Takaki, A.; Zhou, C.J.; Nakai, Y.; Arimura, A. PACAP protects hippocampal neurons against apoptosis: Involvement of JNK/SAPK signaling pathway. Ann. N. Y. Acad. Sci. 1998, 865, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Ohtaki, H.; Funahashi, H.; Dohi, K.; Oguro, T.; Horai, R.; Asano, M.; Iwakura, Y.; Yin, L.; Matsunaga, M.; Goto, N.; et al. Suppression of oxidative neuronal damage after transient middle cerebral artery occlusion in mice lacking interleukin-1. Neurosci. Res. 2003, 45, 313–324. [Google Scholar] [CrossRef]
- Ohtaki, H.; Nakamachi, T.; Dohi, K.; Aizawa, Y.; Takaki, A.; Hodoyama, K.; Yofu, S.; Hashimoto, H.; Shintani, N.; Baba, A.; et al. Pituitary adenylate cyclase-activating polypeptide (PACAP) decreases ischemic neuronal cell death in association with IL-6. Proc. Natl. Acad. Sci. USA 2006, 103, 7488–7493. [Google Scholar] [CrossRef]
- Ohtaki, H.; Nakamachi, T.; Dohi, K.; Shioda, S. Role of PACAP in ischemic neural death. J. Mol. Neurosci. 2007, 36, 16–25. [Google Scholar] [CrossRef]
- Ohtaki, H.; Takeda, T.; Dohi, K.; Yofu, S.; Nakamachi, T.; Satoh, K.; Hiraizumi, Y.; Miyaoka, H.; Matsunaga, M.; Shioda, S. Increased mitochondrial DNA oxidative damage after transient middle cerebral artery occlusion in mice. Neurosci. Res. 2008, 58, 349–355. [Google Scholar] [CrossRef]
- Ohtaki, H.; Ylostalo, J.H.; Foraker, J.E.; Robinson, A.P.; Reger, R.L.; Shioda, S.; Prockop, D.J. Stem/progenitor cells from bone marrow decrease neuronal death in global ischemia by modulation of inflammatory/immune responses. Proc. Natl. Acad. Sci. USA 2008, 105, 14638–14643. [Google Scholar] [CrossRef] [PubMed]
- Nakamachi, T.; Endo, S.; Ohtaki, H.; Yin, L.; Kenji, D.; Kudo, Y.; Funahashi, H.; Matsuda, K.; Shioda, S. Orexin-1 receptor expression after global ischemia in mice. Regul. Pep. 2005, 126, 49–54. [Google Scholar] [CrossRef]
- Nakamachi, T.; Ohtaki, H.; Yofu, S.; Dohi, K.; Watanabe, J.; Mori, H.; Sato, A.; Hashimoto, H.; Shintani, N.; Baba, A.; et al. Endogenous pituitary adenylate cyclase activating polypeptide is involved in suppression of edema in the ischemic brain. Acta. Neurochir. Suppl. 2010, 106, 43–46. [Google Scholar]
- Hori, M.; Nakamachi, T.; Rakwal, R.; Shibato, J.; Nakamura, K.; Wada, Y.; Tsuchikawa, D.; Yoshikawa, A.; Tamaki, K.; Shioda, S. Unraveling the ischemic brain transcriptome in a permanent middle cerebral artery occlusion model by DNA microarray analysis. Dis. Model. Mech. 2012, 5, 270–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hori, M.; Nakamachi, T.; Rakwal, R.; Shibato, J.; Ogawa, T.; Aiuchi, T.; Tsuruyama, T.; Tamaki, K.; Shioda, S. Transcriptomics and proteomics analyses of the PACAP38 influenced ischemic brain in permanent middle cerebral artery occlusion model mice. J. Neuroinflammation 2012, 9, 256. [Google Scholar] [CrossRef] [PubMed]
- Hori, M.; Nakamachi, T.; Rakwal, R.; Shibato, J.; Ogawa, T.; Aiuchi, T.; Tsuruyama, T.; Tamaki, K.; Shioda, S. CORRECTION: Transcriptomics and proteomics analyses of the PACAP38 influenced ischemic brain in permanent middle cerebral artery occlusion model mice. J. Neuroinflammation 2013, 10, 18. [Google Scholar] [CrossRef]
- Tamas, A.; Reglodi, D.; Szanto, Z.; Borsiczky, B.; Nemeth, J.; Lengvari, I. Comparative neuroprotective effects of preischemic PACAP and VIP administration in permanent occlusion of the middle cerebral artery in rats. Neuroendocrinol. Letts. 2002, 23, 249–254. [Google Scholar]
- Chen, Y.; Samal, B.; Hamelink, C.R.; Xiang, C.C.; Chen, Y.; Chen, M.; Vaudry, D.; Brownstein, M.J.; Hallenbeck, J.M.; Eiden, L.E. Neuroprotection by endogenous and exogenous PACAP following stroke. Regul. Pept. 2006, 137, 4–19. [Google Scholar] [CrossRef] [PubMed]
- Dejda, A.; Seaborn, T.; Bourgault, S.; Touzani, O.; Fournier, A.; Vaudry, H.; Vaudry, D. PACAP and a novel stable analog protect rat brain from ischemia: Insight into the mechanisms of action. Peptides 2011, 32, 1207–1216. [Google Scholar] [CrossRef] [PubMed]
- Reglodi, D.; Kiss, P.; Szabadfi, K.; Atlasz, T.; Gabriel, R.; Horvath, G.; Szakaly, P.; Sandor, B.; Lubics, A.; Laszlo, E.; et al. PACAP is an endogenous protective factor-insights from PACAP-deficient mice. J. Mol. Neurosci. 2012, 48, 482–492. [Google Scholar] [CrossRef]
- Tamas, A.; Reglodi, D.; Farkas, O.; Kovesdi, E.; Pal, J.; Povlishock, J.T.; Schwarcz, A.; Czeiter, E.; Szanto, Z.; Doczi, T.; et al. Effect of PACAP in central and peripheral nerve injuries. Int. J. Mol. Sci. 2012, 13, 8430–8448. [Google Scholar] [CrossRef]
- Hori, M.; Nakamachi, T.; Shibato, J.; Rakwal, R.; Tsuchida, M.; Shioda, S.; Numazawa, S. PACAP38 differentially effects genes and CRMP2 protein expression in ischemic core and penumbra regions of permanent middle cerebral artery occlusion model mice brain. Int. J. Mol. Sci. 2014, 15, 17014–17034. [Google Scholar] [CrossRef] [PubMed]
- DeRisi, J.L.; Iyer, V.R.; Brown, P.O. Exploring the metabolic and genetic control of gene expression on a genomic scale. Science 1997, 278, 680–686. [Google Scholar] [CrossRef]
- Ogawa, T.; Rakwal, R.; Shibato, J.; Sawa, C.; Saito, T.; Murayama, A.; Kuwagata, M.; Kageyama, H.; Yagi, M.; Satoh, K.; et al. Seeking gene candidates responsible for developmental origins of health and disease. Congenit. Anom. (Kyoto) 2011, 51, 110–125. [Google Scholar] [CrossRef]
- Ogawa, T.; Shibato, J.; Rakwal, R.; Saito, T.; Tamura, G.; Kuwagata, M.; Shioda, S. Seeking gene candidates responsible for developmental origins of health and disease from the fetal mouse liver following maternal food restriction. Congenit. Anom. (Kyoto) 2014, 54, 195–219. [Google Scholar]
- Suzuki, T.; Higgins, P.J.; Crawford, D.R. Control selection for RNA quantitation. Biotechniques 2000, 29, 332–337. [Google Scholar] [PubMed]
- Series GSE62884. Available online: http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE62884 (accessed on 20 January 2015).
- Hofer, M.; Pagliusi, S.R.; Hohn, A.; Leibrock, J.; Barde, Y.A. Regional distribution of brain-derived neurotrophic factor mRNA in the adult mouse brain. EMBO J. 1990, 9, 2459–2464. [Google Scholar]
- Korte, M.; Carroll, P.; Wolf, E.; Brem, G.; Thoenen, H.; Bonhoeffer, T. Hippocampal long-term potentiation is impaired in mice lacking brain-derived neurotrophic factor. Proc. Natl. Acad. Sci. USA 1995, 92, 8856–8860. [Google Scholar] [CrossRef]
- Pettersson, L.M.; Geremia, N.M.; Ying, Z.; Verge, V.M. Injury-associated PACAP expression in rat sensory and motor neurons is induced by endogenous BDNF. PLoS One 2014, 9, e100730. [Google Scholar] [CrossRef] [PubMed]
- Santos, S.D.; Lambertsen, K.L.; Clausen, B.H.; Akinc, A.; Alvarez, R.; Finsen, B.; Sraiva, M.J. CSF transthyretin neuroprotection in a mouse model of brain ischemia. J. Neurochem. 2010, 115, 1434–1444. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cattaneo, F.; Ryno, L.; Hulleman, J.; Reixach, N.; Buxbaum, J.N. The systemic amyloid precursor transthyretin (TTR) behaves as a neuronal stress protein regulated by HSF1 in SH-SY5Y human neuroblastoma cells and APP23 Alzheimer’s disease model mice. J. Neurosci. 2014, 34, 7253–7265. [Google Scholar] [CrossRef] [PubMed]
- Vieira, M.; Gomes, J.R.; Saraiva, M.J. Transthyretin induces insulin-like growth factor 1 nuclear translocation regulating its levels in the Hippocampus. Mol. Neurobiol. 2014. [Google Scholar] [CrossRef]
- Lazarovici, P.; Cohen, G.; Arien-Zakay, H.; Chen, J.; Zhang, C.; Chopp, M.; Jiang, H. Multimodal neuroprotection induced by PACAP38 in oxygen-glucose deprivation and middle cerebral artery occlusion stroke models. J. Mol. Neurosci. 2012, 48, 526–540. [Google Scholar] [CrossRef] [PubMed]
- NCBI Reference Sequence: NM_009921.2. Available online: http://www.ncbi.nlm.nih.gov/nuccore/NM_009921.2 (accessed on 20 January 2015).
- Brandenburg, L.O.; Jansen, S.; Wruck, C.J.; Lucius, R.; Pufe, T. Antimicrobial peptide rCRAMP induced glial cell activation through P2Y receptor signalling pathways. Mol. Immunol. 2010, 47, 1905–1913. [Google Scholar] [CrossRef] [PubMed]
- NCBI Reference Sequence: NM_133245.1. Available online: http://www.ncbi.nlm.nih.gov/nuccore/NM_133245.1 (accessed on 20 January 2015).
- Kihm, A.J.; Kong, Y.; Hong, W.; Russell, J.E.; Rouda, S.; Adachi, K.; Simon, M.C.; Blobel, G.A.; Weiss, M.J. An abundant erythroid protein that stabilizes free alpha-haemoglobin. Nature 2002, 417, 758–763. [Google Scholar] [CrossRef]
- Kong, Y.; Zhou, S.; Kihm, A.J.; Katein, A.M.; Yu, X.; Gell, D.A.; Mackay, J.P.; Adachi, K.; Foster-Brown, L.; Louden, C.S.; et al. Loss of alpha-haemoglobin stabilizing protein impairs erythropoiesis and exacerbates beta-thalassemia. J. Clin. Invest. 2004, 114, 1457–1466. [Google Scholar] [CrossRef]
- Feng, L.; Zhou, S.; Gu, L.; Gell, D.A.; Mackay, J.P.; Weiss, M.J.; Gow, A.J.; Shi, Y. Structure of oxidized α-haemoglobin bound to AHSP reveals a protective mechanims for haem. Nature 2005, 435, 697–701. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Zhao, G.W.; Yu, W.; Xie, X.M.; Wang, W.T.; Yang, R.F.; Lv, X.; Liu, D.P. Activation of STAT3 stimulates AHSP expression in K562 cells. Sci. China Life Sci. 2014, 57, 488–494. [Google Scholar] [CrossRef] [PubMed]
- NCBI Reference Sequence: NM_010510.1. Available online: http://www.ncbi.nlm.nih.gov/nuccore/6754303 (accessed on 20 January 2015).
- Ambjorn, M.; Ejlerskov, P.; Liu, Y.; Lees, M.; Jaattela, M.; Issazadeh-Navikas, S. IFNB1/interferon-β-induced autophagy in MCF-7 breast cancer cells counteracts its proapoptotic function. Autophagy 2013, 9, 287–302. [Google Scholar] [CrossRef] [PubMed]
- Belardelli, F.; Gresser, I. The neglected role of type I interferon in the T-cell response: Implications for its clinal use. Immunol. Today 1996, 17, 369–372. [Google Scholar] [CrossRef] [PubMed]
- Morrell, C.N.; Srivastava, K.; Swaim, A.; Lee, M.T.; Chen, J.; Nagineni, C.; Hooks, J.J.; Detrick, B. Beta interferon suppresses the development of experimental cerebral malaria. Infect. Immun. 2011, 79, 1750–1758. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, Y.; Wang, L.; Zhnag, Z.; Lu, D.; Lu, M.; Chopp, M. Therapeutic benefit of intravenous administration of bone marrow stromal cells after cerebral ischemia in rats. Stroke 2001, 32, 1005–1011. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Hong, J.M.; Moon, G.J.; Lee, P.H.; Ahn, Y.H.; Bang, O.Y. A long-term follow-up study of intravenous autologous mesenchymal stem cell transplantation in patients with ischemic stroke. Stem Cells 2010, 28, 1099–1106. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, J.Y.; Wang, H.W.; Chang, S.J.; Liao, K.H.; Lee, I.H.; Lin, W.S.; Wu, C.H.; Lin, W.Y.; Cheng, S.M. Mesenchymal stem cells from human umbilical cord express preferentially secreted factors related to neuroprotection, neurogenesis, and angiogenesis. PLoS One 2013, 8, e72604. [Google Scholar] [CrossRef] [PubMed]
- Morales, I.; Farias, G.; Maccioni, R.B. Neuroimmunomodulation in the pathogenesis of Alzheimer’s disease. Neuroimmunomodulation 2010, 17, 202–204. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Dinarello, C.A.; Mier, J.W. IL-6 and APPs: Anti-inflammatory and immunosuppressive mediators. Immunol. Today 1997, 18, 482–432. [Google Scholar] [CrossRef]
- Loddick, S.A.; Turnbull, A.V.; Rothwell, N.J. Cerebral interleukin-6 is neuroprotective during permanent focal cerebral ischemia in the rat. J. Cereb. Blood Flow Metab. 1998, 18, 176–179. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.P.; Qiu, Y.H.; Lu, J.H.; Wang, J.J. Interleukin-6 protects cultured cerebellar granule neurons against glutamate-induced neurotoxicity. Neurosci. Lett. 2005, 374, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Shinozaki, Y.; Nomura, M.; Iwatsuki, K.; Moriyama, Y.; Gachet, C.; Koizumi, S. Microglia trigger astrocyte-mediated neuroprotection via purinergic gliotransmission. Sci. Rep. 2014, 4, 4329. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Benveniste, E.N. Immune functions of astrocytes. Glia 2001, 36, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.; Frederick, T.J.; Miller, S.D. Astrocytes in multiple sclerosis: A product of their environment. Cell. Mol. Life Sci. 2008, 65, 2702–2720. [Google Scholar] [CrossRef] [PubMed]
- Rostene, W.; Dansereau, M.A.; Godefroy, D.; Van Steenwinckel, J.; Goazigo, A.R.-L.; Melik-Parsadaniantz, S.; Apartis, E.; Hunot, S.; Beaudet, N.; Sarret, P. Neurochemokines: A menage a trois providing new insights on the functions of chemokines in the central nervous system. J. Neurochem. 2011, 118, 680–694. [Google Scholar] [CrossRef] [PubMed]
- Israelsson, C.; Kylberg, A.; Bengtsson, H.; Hillered, L.; Ebendal, T. Interacting chemokine signals regulate dendritic cells in acute brain injury. PLoS One 2014, 9, e104754. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Koo, E.H. Biology and pathophysiology of the amyloid precursor protein. Mol. Neurodegener. 2011, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wang, Z.; Sun, L.; Yang, L.; Hongmei, L.; Cole, A.L.; Rodriguez-Rivera, J.; Lu, H.-C.; Zheng, H. The amyloid precursor protein controls adult hippocampal neurogenesis through GABAergic interneurons. J. Neurosci. 2014, 34, 13314–13325. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Tessier-Lavinge, M. Neuropilin is a recepor for the axonal chemorepellent semaphorin III. Cell 1997, 90, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Oz-Levi, D.; Gelman, A.; Elazar, Z.; Lancer, D. TECPR2: A new authophagy link for neurodegeneration. Autophagy 2013, 95, 801–802. [Google Scholar] [CrossRef]
- McNeill, E.M.; Klockner-Bormann, M.; Roesler, E.C.; Talton, L.E.; Moechars, D.; Clagett-Dame, M. Nav2 hypomorphic mutant mice are ataxic and exhibit abnormalities in cerebellar development. Dev. Biol. 2011, 353, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Gene Expression Omnibus. Available online: http://www.ncbi.nlm.nih.gov/geo/ (accessed on 20 January 2015).
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hori, M.; Nakamachi, T.; Shibato, J.; Rakwal, R.; Shioda, S.; Numazawa, S. Unraveling the Specific Ischemic Core and Penumbra Transcriptome in the Permanent Middle Cerebral Artery Occlusion Mouse Model Brain Treated with the Neuropeptide PACAP38. Microarrays 2015, 4, 2-24. https://doi.org/10.3390/microarrays4010002
Hori M, Nakamachi T, Shibato J, Rakwal R, Shioda S, Numazawa S. Unraveling the Specific Ischemic Core and Penumbra Transcriptome in the Permanent Middle Cerebral Artery Occlusion Mouse Model Brain Treated with the Neuropeptide PACAP38. Microarrays. 2015; 4(1):2-24. https://doi.org/10.3390/microarrays4010002
Chicago/Turabian StyleHori, Motohide, Tomoya Nakamachi, Junko Shibato, Randeep Rakwal, Seiji Shioda, and Satoshi Numazawa. 2015. "Unraveling the Specific Ischemic Core and Penumbra Transcriptome in the Permanent Middle Cerebral Artery Occlusion Mouse Model Brain Treated with the Neuropeptide PACAP38" Microarrays 4, no. 1: 2-24. https://doi.org/10.3390/microarrays4010002