Anti-Inflammatory Activity of Chrysophanol through the Suppression of NF-kB/Caspase-1 Activation in Vitro and in Vivo

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
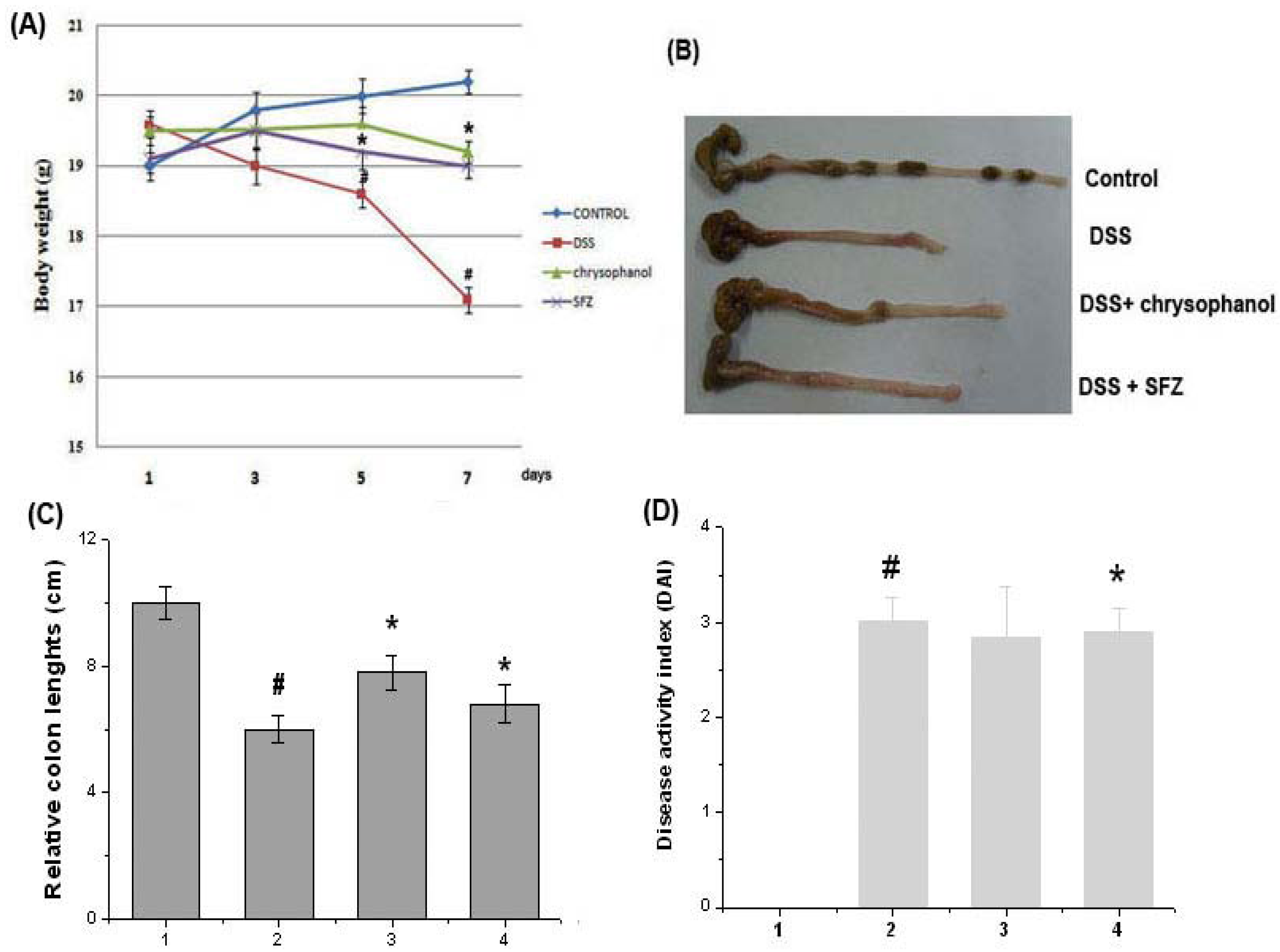
2.1. Effect of Chrysophanol on Clinical Signs in DSS-Induced Colitis
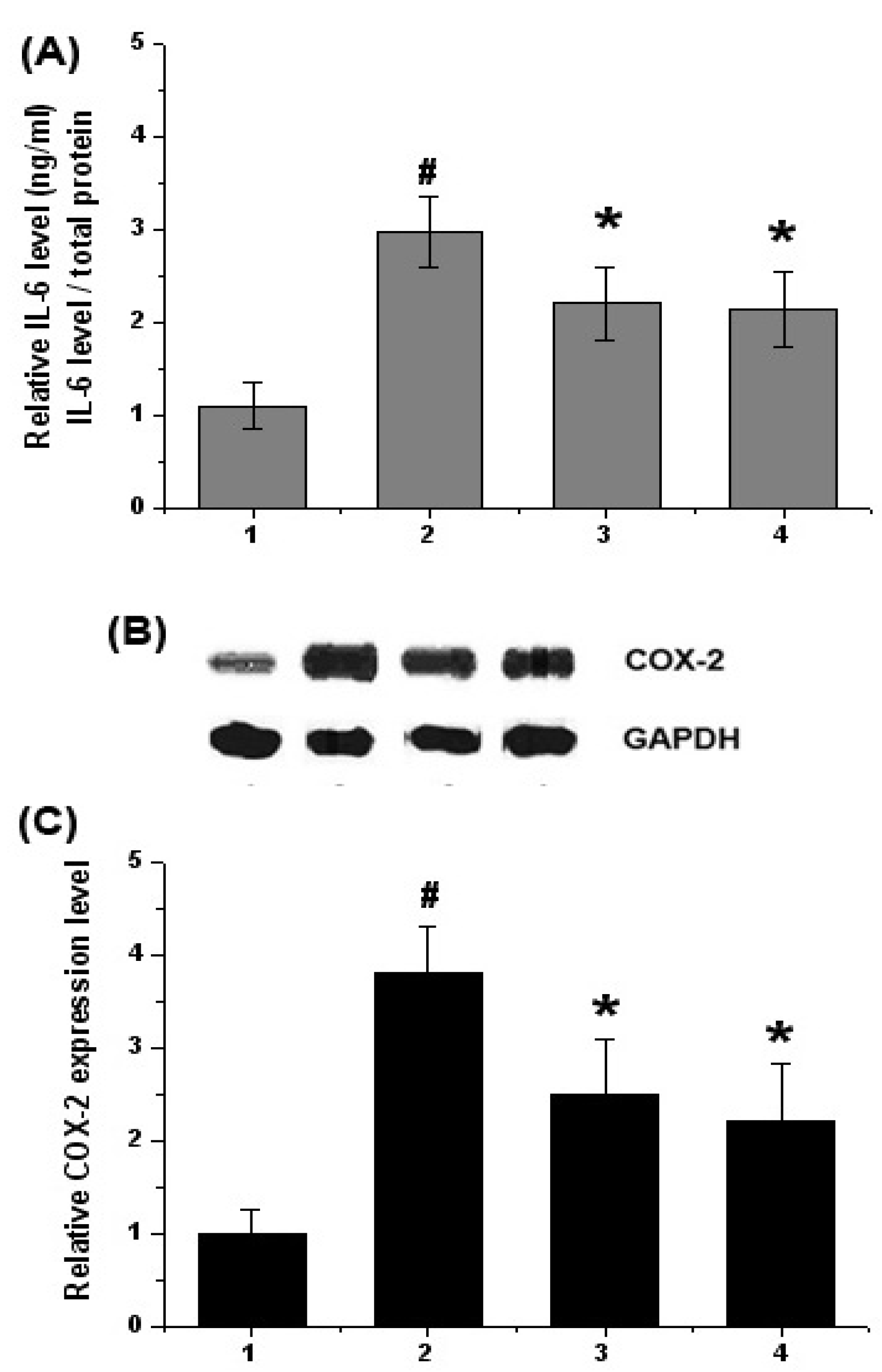
2.2. Effect of Chrysophanol on Levels of Inflammatory Mediators (IL-6 and COX-2) in DSS-Induced Colitis


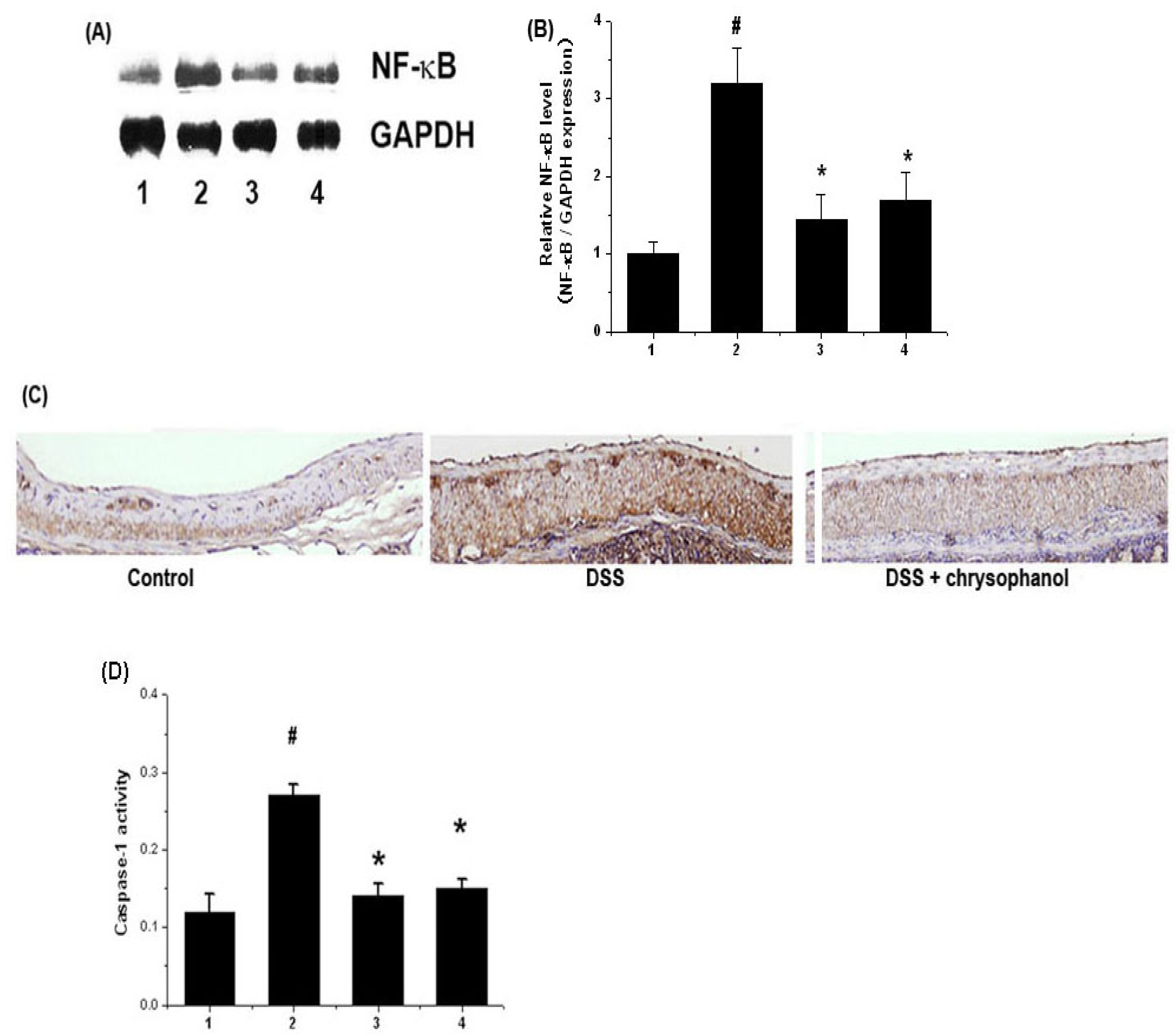
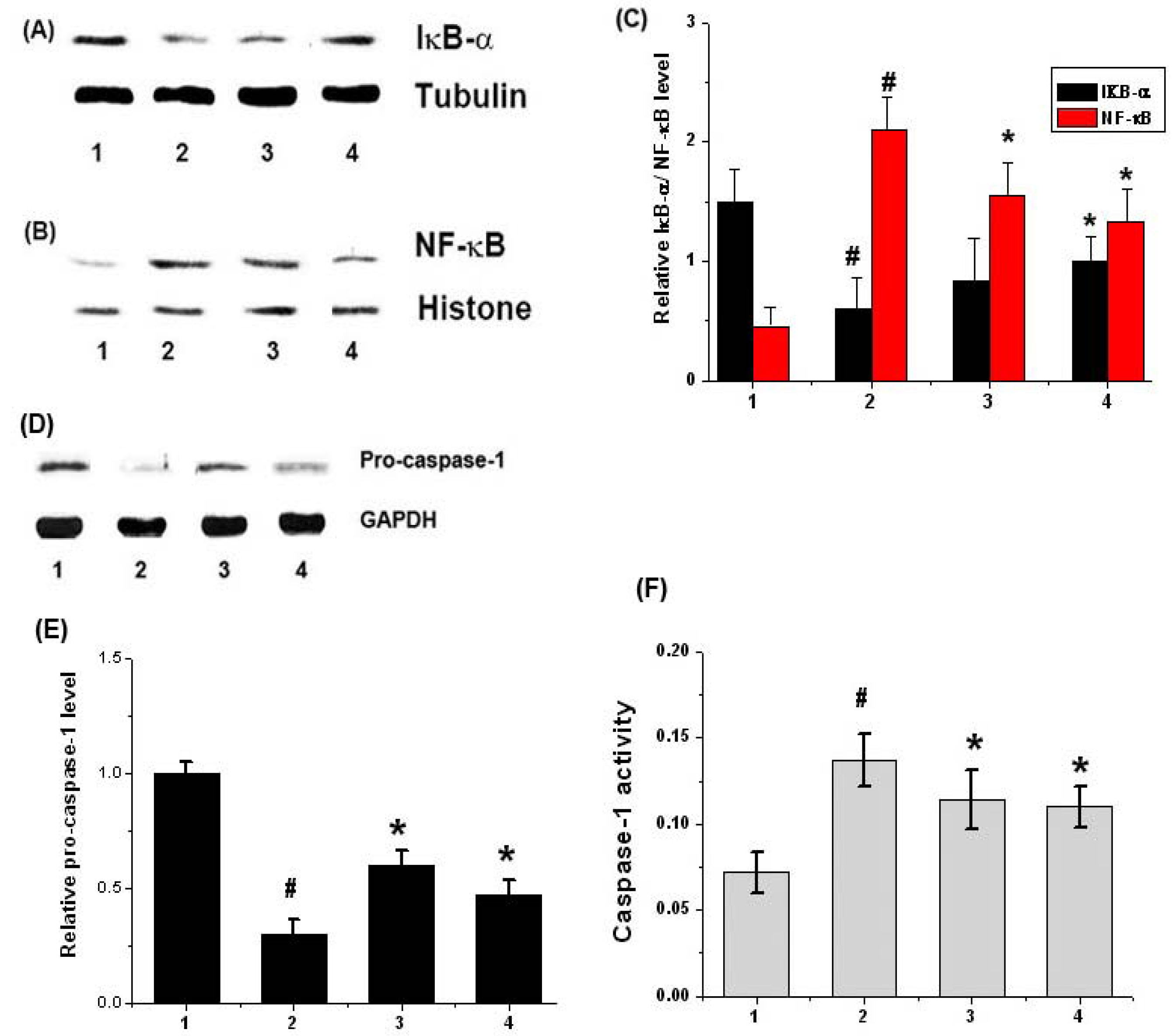
2.3. Effect of Chrysophanol on Activation of NF-κB (p65) and Caspase-1 in DSS-Induced Colitis

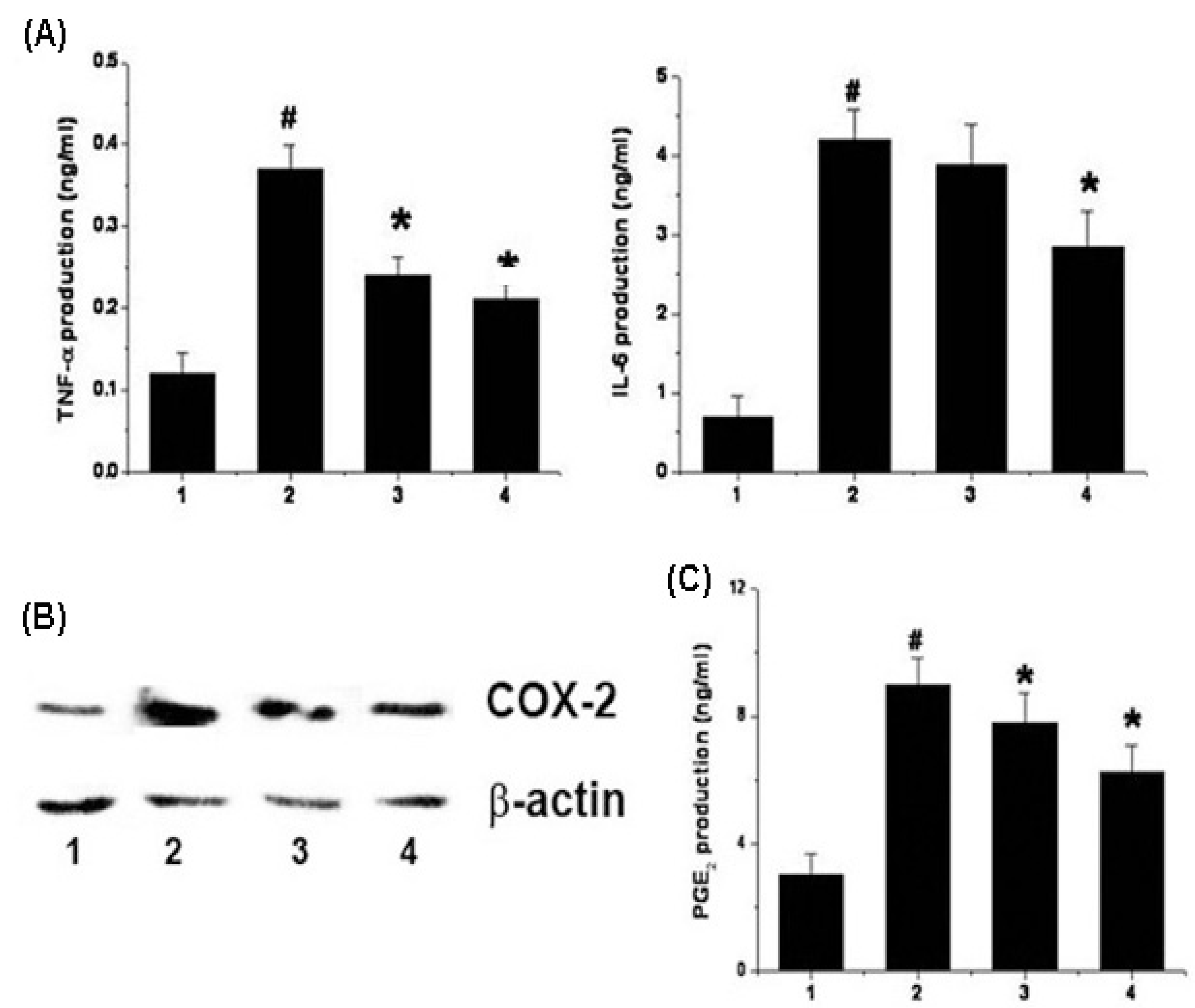
2.4. Effects of Chrysophanol on the Production of Cytokine, PGE2 and Expression of COX-2 in LPS-Stimulated Mouse Peritoneal Macrophage

2.5. Effects of Chrysophanol on NF-κB and Caspase-1 Activation in LPS-Stimulated Mouse Peritoneal Macrophage
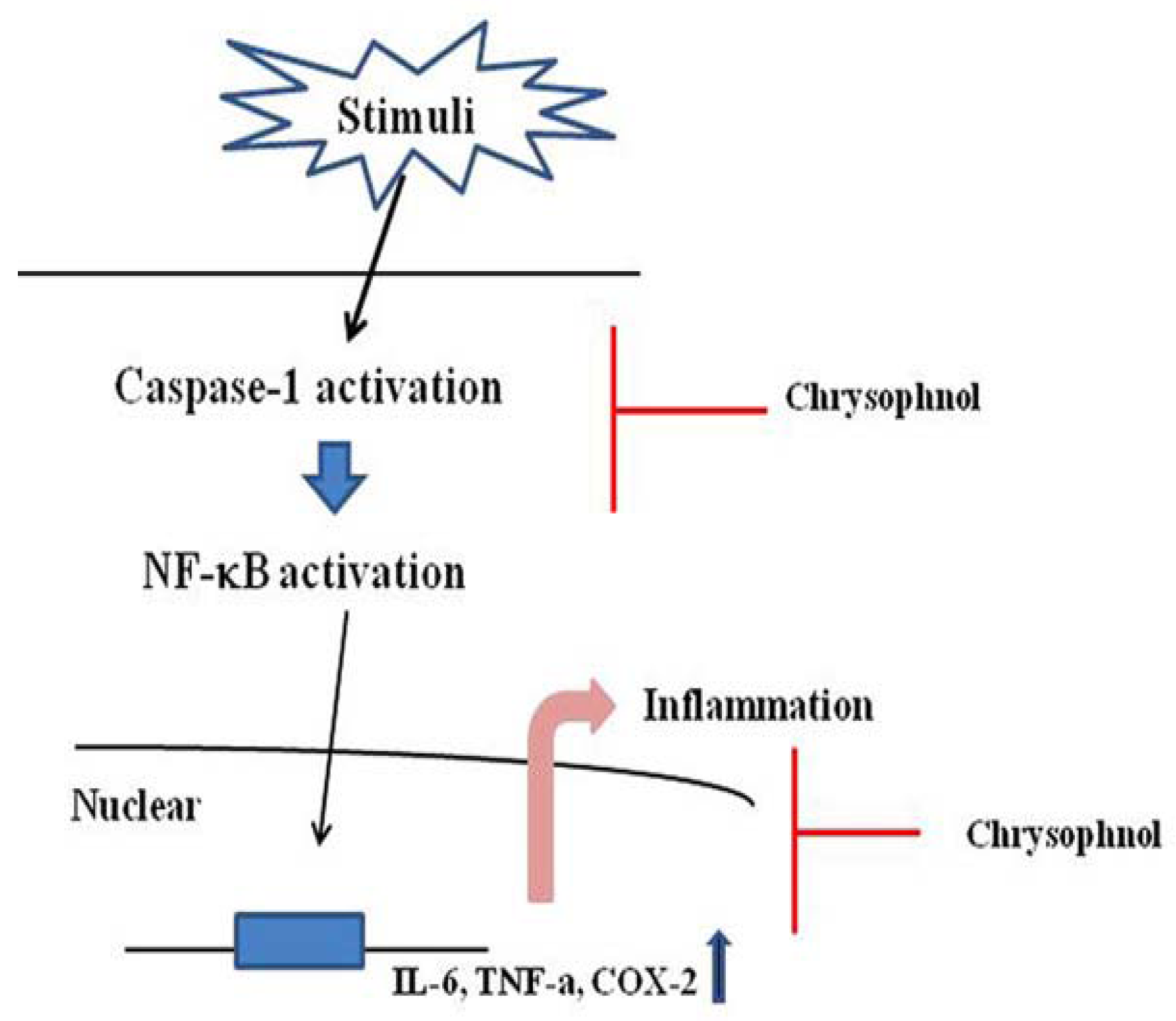
2.6. Discussion


3. Experimental
3.1. Reagents
3.2. Animals
3.3. Peritoneal Macrophage Cultures
3.4. MTT Assay
3.5. Induction of Colitis by DSS
3.6. DAI
3.7. Cytokines and Prostaglandins E2 (PGE2) Assay
3.8. Preparation of Cytoplasmic and Nuclear Extract
3.9. Western Blot Analysis
3.10. Assay of Caspase-1 Activity
3.11. Statistical Analysis
4. Conclusions
Acknowledgements
References
- Medzhitov, R.; Janeway, C.A.Jr. Innate immunity: the virtues of a nonclonal system of recognition. Cell 1997, 9, 295–298. [Google Scholar] [CrossRef]
- Beutler, B. TNF, Tlr4: central component of the sole mammalian LPS sensor. Curr. Opin. Immunol. 2000, 12, 20–26. [Google Scholar]
- Beutler, B. TNF, immunity and inflammatory disease: lessons of the past decade. J. Investig. Med. 1995, 43, 227–235. [Google Scholar]
- Parrillo, J.E. Pathogenetic mechanisms of septic shock. N. Engl. J. Med. 1993, 328, 1471–1477. [Google Scholar] [CrossRef]
- Papadakis, K.A.; Targan, S.R. Role of cytokines in the pathogenesis of inflammatory bowel disease. Ann. Rev. Med. 2000, 51, 289–298. [Google Scholar] [CrossRef]
- Rogler, G.; Andus T. Cytokines in inflammatory bowel disease. World J. Surg. 1998, 22, 382–389. [Google Scholar] [CrossRef]
- Ogata, H.; Hibi, T. Cytokine and anti-cytokine therapies for inflammatory bowel disease. Curr. Pharm. Des. 2003, 9, 1107–1113. [Google Scholar] [CrossRef]
- Roberts, P.J.; Morgan, K.; Miller, R.; Hunter, J.O.; Middleton, S.J. Neuronal COX-2 expression in human myenteric plexus in active inflammatory bowel disease. Gut 2001, 48, 468–472. [Google Scholar] [CrossRef]
- Agoff, S.N.; Brentnall, T.A.; Crispin, D.A.; Taylor, S.L.; Raaka, S.; Haggitt, R.C.; Reed, M.W.; Afonina, I.A.; Rabinovitch, P.S.; Stevens, A.C.; Feng, Z.; Bronner, M.P. The role of cyclooxygenase 2 in ulcerative colitis-associated neoplasia. Am. J. Pathol. 2000, 157, 737–745. [Google Scholar] [CrossRef]
- Baldwin, A.S.Jr. The NF-kappa B and I kappa B proteins: new discoveries and insights. Ann. Rev. Immunol. 1996, 14, 649–683. [Google Scholar] [CrossRef]
- Finco, T.; Baldwin, A.S., Jr. Mechanistic aspects of NF-kB regulation: the emerging role of phosphorylation and proteolysis. Immunity 1995, 3, 263–272. [Google Scholar] [CrossRef]
- Neurath, M.F.; Fuss, I.; Schurmann, G.; Pettersson, S.; Arnold, K.; Müller-Lobeck, H.; Strober, W.; Herfarth, C.; Büschenfelde, K.H. Cytokine gene transcription by NF-kB family members in patients with inflammatory bowel disease. Ann. NY Acad. Sci. 1998, 859, 149–159. [Google Scholar]
- Schreiber, S.; Nikolaus, S.; Hampe, J. Activation of nuclear factor kappa B in inflammatory bowel disease. Gut 1998, 42, 477–484. [Google Scholar] [CrossRef]
- Lee, S.H.; Stehlik, C.; Reed, J.C. Cop, a caspase recruitment domaincontaining protein and inhibitor of caspase-1 activation processing. J. Biol. Chem. 2001, 276, 34495–34500. [Google Scholar]
- Wang, X.; Wang, H.Y.; Bryan, E.F.; Zhang, W.H.; Huo, C.F.; Guan, Y.J.; Zhang, Y.; Bruey, J.M.; John, C.R.; Robert, M.F. Dysregulation of receptor interacting protein-2 and caspase recruitment domain only protein mediates aberrant caspase-1 activation in Huntington’s disease. J. Neurosci. 2005, 25, 11645–11654. [Google Scholar]
- Faubel, S.; Lewis, E.C.; Reznikov, L.; Ljubanovic, D.; Hoke, T.S.; Somerset, H.; Oh, D.J.; Lu, L.; Klein, C.L.; Dinarello, C.A.; Edelstein, C.L. Cisplatin-induced acute renal failure is associated with an increase in the cytokines interleukin (IL)-1beta, IL-18, IL-6, and neutrophil infiltration in the kidney. J. Pharmacol. Exp. Ther. 2007, 322, 8–15. [Google Scholar]
- Siegmund, B.; Lehr, H.A.; Fantuzzi, G.; Dinarello, C.A. IL-1 beta -converting enzyme (caspase-1) in intestinal inflammation. Proc. Natl. Acad. Sci. USA 2001, 6, 13249–13254. [Google Scholar]
- Huang, Q.; Lu, G.; Shen, H.M.; Chung, M.C.; Ong, C.N. Anti-cancer properties of anthraquinones from rhubarb. Med. Res. Rev. 2007, 27, 609–630. [Google Scholar] [CrossRef]
- Shoemaker, M.; Hamilton, B.; Dairkee, S.H.; Cohen, I.; Campbell, M.J. In vitro anticancer activity of twelve Chinese medicinal herbs. Phytother. Res. 2005, 19, 649–651. [Google Scholar] [CrossRef]
- Arosio, B.; Gagliano, N.; Fusaro, L.M.; Parmeggiani, L.; Tagliabue, J.; Galetti, P. Aloe-Emodin quinone pretreatment reduces acute liver injury induced by carbon tetrachloride. Pharmacol. toxicol. 2000, 87, 229–233. [Google Scholar]
- Fosse, C.; Le Texier, L.; Roy, S.; Delaforge, M.; Grégoire, S.; Neuwels, M.; Azerad, R. Parameters and mechanistic studies on the oxidative ring cleavage of synthetic heterocyclic naphthoquinones by Streptomyces strains. Appl. Microbiol. Biotechnol. 2004, 65, 446–456. [Google Scholar]
- Cooper, H.S.; Murthy, S.N.; Shah, R.S.; Sedergran, D.J. Clinicopathologic study of dextran sulfate sodium experimental murine colitis. Lab. Invest. 1993, 69, 238–249. [Google Scholar]
- Okayasu, I.; Hatakeyama, S.; Yamada, M.; Ohkusa, T.; Inagaki, Y.; Nakaya, R. Novel method in the induction of reliable experimental acute and chronic ulcerative colitis in mice. Gastroenterology 1990, 98, 694–702. [Google Scholar]
- Murthy, S.N.; Cooper, H.S.; Shim, H.; Shah, R.S.; Ibarahim, S.A.; Sedergran, D.J. Treatment of dextran sulfate sodium-induced murine colitis by intracolonic cyclosporin. Dig. Dis. Sci. 1993, 38, 1722–1734. [Google Scholar] [CrossRef]
- Ghosh, S.; May, M.J.; Kopp, E.B. NF-kappa B and Rel proteins: evolutionarily conserved mediators of immune responses. Ann. Rev. Immunol. 1998, 16, 225–260. [Google Scholar] [CrossRef]
- Bouma, G.; Strober, W. Theimmunological and genetic basis of inflammatory bowel disease. Nat. Rev. Immunol. 2003, 3, 521–533. [Google Scholar] [CrossRef]
- Ishiguro, K.; Ando, T.; Maeda, O.; Hasegawa, M.; Kado-matsu, K.; Ohmiya, N.; Niwa, Y.; Xavier, R.; Goto, H. Paeonol attenuates TNBS-induced colitis by inhibiting NF-kappaB and STAT1 transactivation. Toxicol. Appl. Pharmacol. 2006, 217, 35–42. [Google Scholar] [CrossRef]
- Sandborn, W.J.; Targan, S.R. Biologic therapy of inflammatory bowel disease. Gastroenterology 2002, 122, 1592–1608. [Google Scholar] [CrossRef]
- Loher, F.; Bauer, C.; Landauer, N.; Schmall, K.; Siegmund, B.; Lehr, H.A.; Dauer, M.; Schoenharting, M.; Endres, S.; Eigler, A. The interleukin-1 beta-converting enzyme inhibitor pralnacasan reduces dextran sulfate sodium-induced murine colitis and T helper 1 T-cell activation. J. Pharmacol. Exp. Ther. 2004, 308, 583–590. [Google Scholar]
- Lamkanfi, M.; Kalai, M.; Saelens, X.; Declercq, W.; Vandenabeele, P. Caspase-1 activates nuclear factor of the kappa-enhancer in B cells independently of its enzymatic activity. J. Biol. Chem. 2004, 4, 24785–24793. [Google Scholar]
- Narumi, S.; Finke, J.H.; Hamilton, T.A. Interferon gamma and interleukin 2 synergize to induce selective monokine expression in murine peritoneal macrophages. J. Biol. Chem. 1990, 265, 7036–7041. [Google Scholar]
- Hendrickson, B.A.; Gokhale, R.; Cho, J.H. Clinical aspects and pathophysiology of inflammatory bowel disease. Clin. Microbiol. Rev. 2002, 15, 79–94. [Google Scholar] [CrossRef]
- Kim, M.S.; Lim, W.K.; Cha, J.G.; An, N.H.; Yoo, S.J.; Park, J.H.; Kim, H.M.; Lee, Y. M. The activation of PI 3-K and PKC zeta in PMA-induced differentiation of HL-60 cells. Cancer Lett. 2001, 171, 79–85. [Google Scholar] [CrossRef]
- Schoonbroodt, S.; Legrand-Poels, S.; Best-Belpomme, M.; Piette, J. Activation of the NF-kappaB transcription factor in a T-lymphocytic cell line by hypochlorous acid. Biochem. J. 1997, 321, 777–785. [Google Scholar]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kim, S.-J.; Kim, M.-C.; Lee, B.-J.; Park, D.-H.; Hong, S.-H.; Um, J.-Y. Anti-Inflammatory Activity of Chrysophanol through the Suppression of NF-kB/Caspase-1 Activation in Vitro and in Vivo. Molecules 2010, 15, 6436-6451. https://doi.org/10.3390/molecules15096436
Kim S-J, Kim M-C, Lee B-J, Park D-H, Hong S-H, Um J-Y. Anti-Inflammatory Activity of Chrysophanol through the Suppression of NF-kB/Caspase-1 Activation in Vitro and in Vivo. Molecules. 2010; 15(9):6436-6451. https://doi.org/10.3390/molecules15096436
Chicago/Turabian StyleKim, Su-Jin, Min-Cheol Kim, Byong-Joo Lee, Dae-Hee Park, Seung-Heon Hong, and Jae-Young Um. 2010. "Anti-Inflammatory Activity of Chrysophanol through the Suppression of NF-kB/Caspase-1 Activation in Vitro and in Vivo" Molecules 15, no. 9: 6436-6451. https://doi.org/10.3390/molecules15096436