Evaluation of the Effects of Mitragyna speciosa Alkaloid Extract on Cytochrome P450 Enzymes Using a High Throughput Assay

Abstract
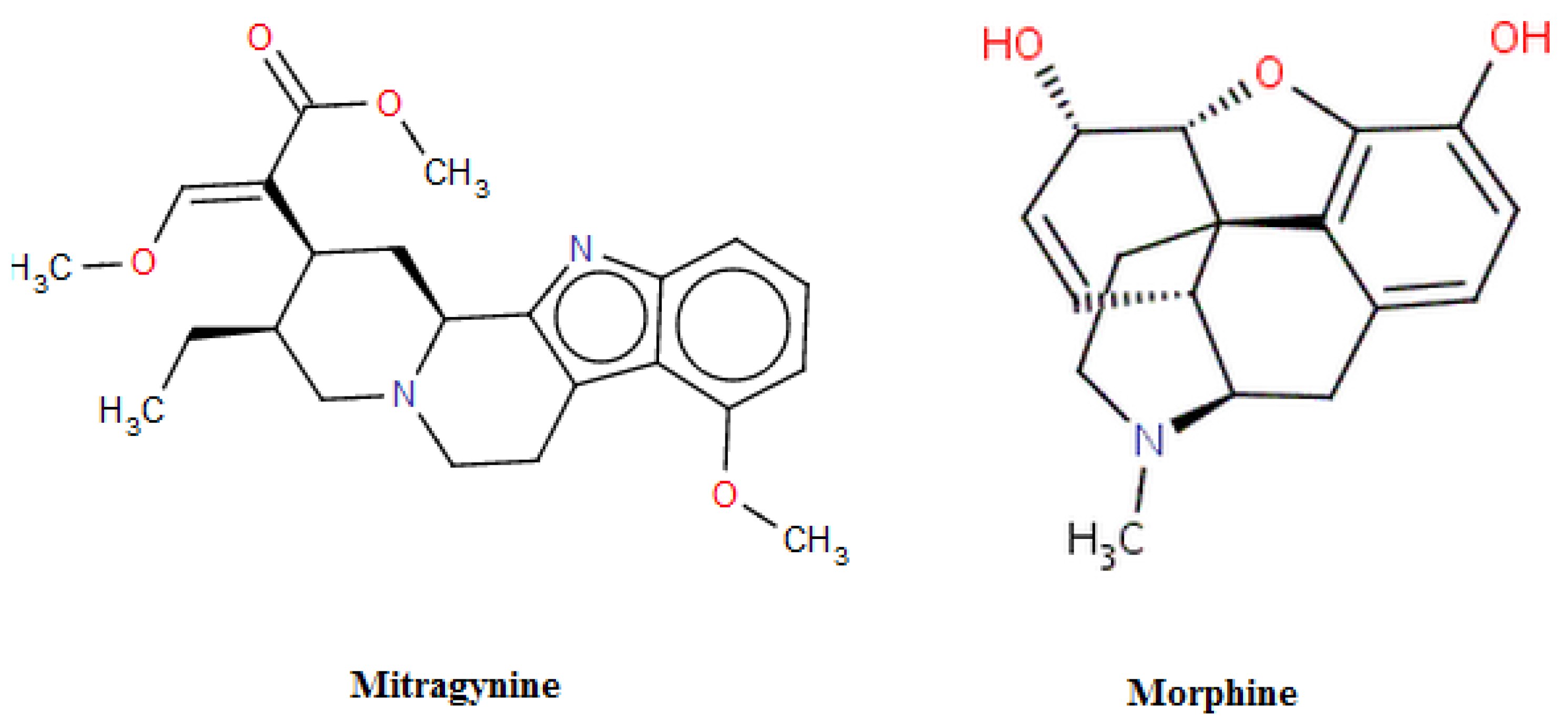
:1. Introduction
2. Results and Discussion
2.1. Determination of Optimal Incubation Time
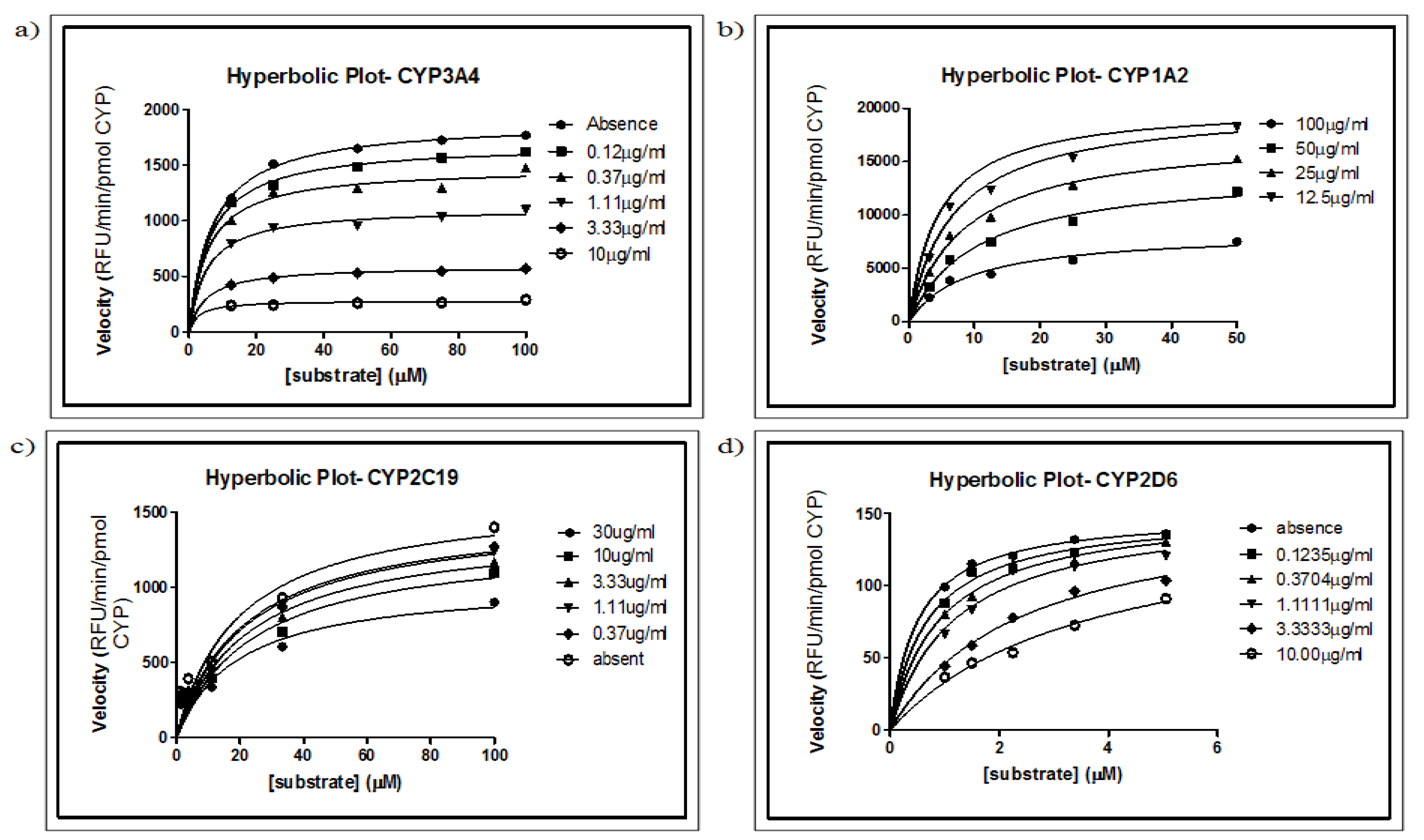
2.2. Determination of Km and Vmax Values
2.3. Determination of IC50 for Standard Inhibitors
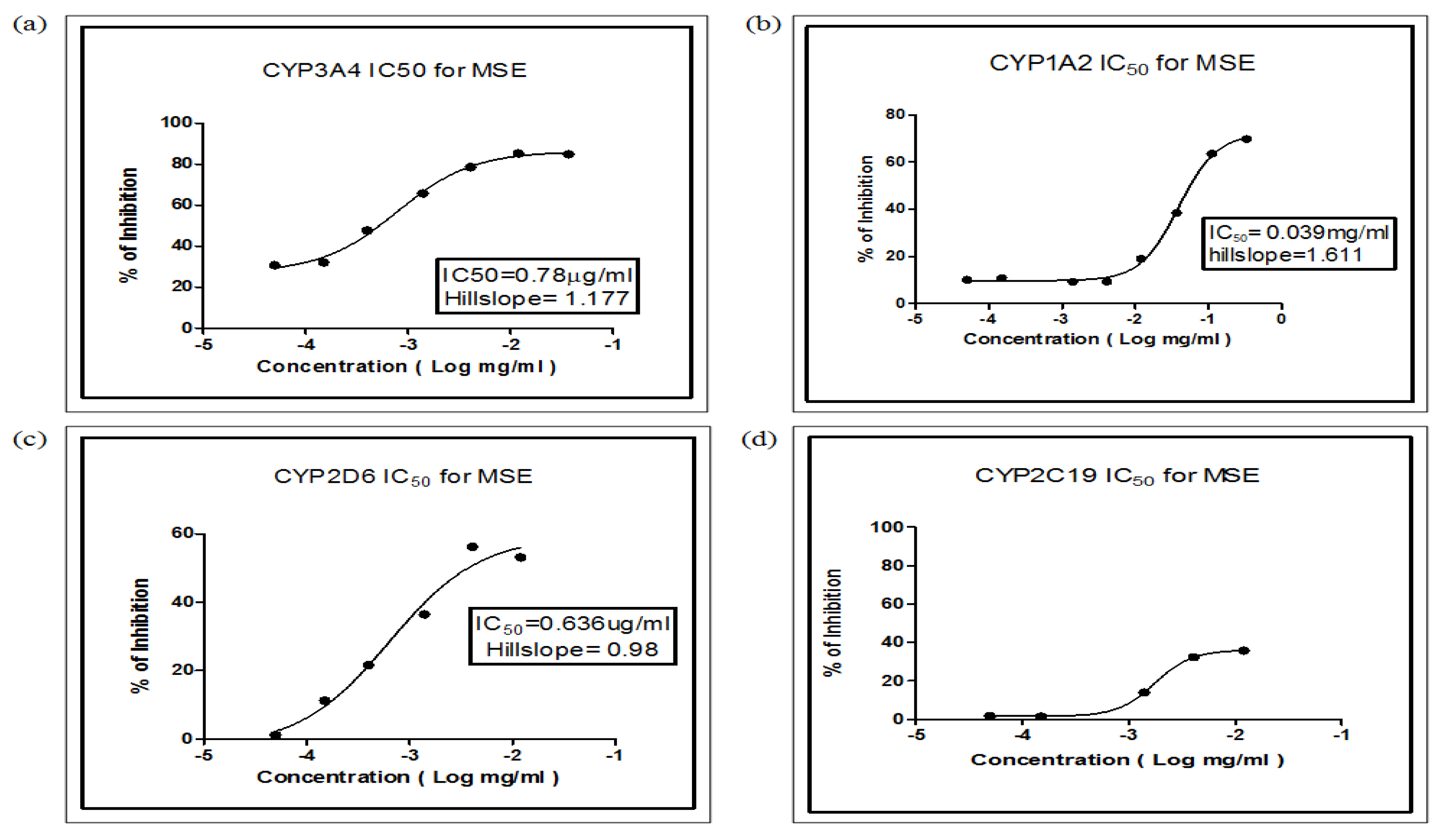
2.4. Determination of IC50 for MSE
2.5. Determination of Ki Values and Modes of Inhibition for MSE
3. Experimental
3.1. Chemicals
3.2. Plant Extraction
3.3. Fluorometric Enzyme Inhibition Assays
3.3.1. Time and Concentration Linearity
3.3.2. Determination of Km and Vmax Values
3.3.3. Half-Maximal Inhibitory Concentration (IC50) Determination of Inhibitors and Plant Extracts
3.3.4. Determination of Ki Values and Modes Of Inhibition
4. Conclusions
Acknowledgments
Conflict of Interest
Abbreviations
References
- Saidin, N.; Gooderham, N.J. In vitro toxicology of extract of Mitragyna speciosa Korth, a Malaysian phytopharmaceutical of abuse. Toxicology 2007, 240, 166–167. [Google Scholar] [CrossRef]
- Babu, K.M.; McCurdy, C.R.; Boyer, E.W. Opioid receptors and legal highs: Salvia divinorum and Kratom. Clin. Toxicol. 2008, 46, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Thongpradichote, S.; Matsumoto, K.; Tohda, M.; Takayama, H.; Aimi, N.; Sakai, S.; Watanabe, H. Identification of opioid receptor subtypes in antinociceptive action of supraspinal-administered mitragynine in mice. Life Sci. 1998, 62, 1371–1378. [Google Scholar] [CrossRef]
- Chittrakarn, S.; Sawangjaroen, K.; Prasettho, S.; Janchawee, B.; Keawpradub, N. Inhibitory effects of kratom leaf extract (Mitragyna speciosa Korth.) on the rat gastrointestinal tract. J. Ethnopharmacol. 2008, 116, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Porrogi, P.; Kóbori, L.; Kõhalmy, K.; Gulyás, J.; Vereczkey, L.; Monostory, K. Limited applicability of 7-methoxy-4-trifluoromethylcoumarin as a CYP2C9-selective substrate. Pharmacol. Rep. 2008, 60, 972–979. [Google Scholar] [PubMed]
- Hanapi, N.A.; Azizi, J.; Ismail, S.; Mansor, S.M. Evaluation of selected Malaysian medicinal plants on phase I drug metabolizing enzymes, CYP2C9, CYP2D6 and CYP3A4 activities in vitro. Int. J. Pharmacol. 2010, 6, 490–495. [Google Scholar] [CrossRef]
- Ogu, C.C.; Maxa, J.L. Drug interactions due to cytochrome P450. Proc. Bay. Univ. Med. Cent 2000, 13, 421–423. [Google Scholar] [CrossRef]
- Ito, K.; Ogihara, K.; Kanamitsu, S.; Itoh, T. Prediction of the in vivo interaction between midazolam and macrolides based on in vitro studies using human liver microsomes. Drug Metab. Dispos. 2003, 31, 945–954. [Google Scholar] [CrossRef] [PubMed]
- U.S. Food and Drug Administration. Guidance for Industry: Drug Metabolism/Drug Interaction Studies in the Drug Development Process: Studies in Vitro; U.S. Food and Drug Administration: Rockville, MD, USA, 1997. Available online: http://www.fda.gov/cder/guidance.htm (accessed on 19 August 2011).
- Fowler, S.; Zhang, H. In vitro evaluation of reversible and irreversible cytochrome P450 inhibition: Current status on methodologies and their utility for predicting drug-drug interactions. AAPS J. 2008, 10, 410–424. [Google Scholar] [CrossRef] [PubMed]
- Crespi, C.L.; Charles, L.; Miller, V.P.; Penman, B.W. Microtiter plate assays for inhibition of human, drug-metabolizing cytochromes P450. Anal. Biochem. 1997, 248, 188–190. [Google Scholar] [CrossRef] [PubMed]
- Pelkonen, O.; Mäenpää, J.; Taavitsainen, P.; Rautio, A.; Raunio, H. Inhibition and induction of human cytochrome P450 (CYP) enzymes. Xenobiotica 1998, 28, 1203–1253. [Google Scholar] [CrossRef] [PubMed]
- Donato, M.T.; Jiménez, N.; Castell, J.V.; Gómez-Lechón, M.J. Fluorescence-based assays for screening nine cytochrome P450 (P450) activities in intact cells expressing individual human P450 enzymes. Drug Metab. Dispos. 2004, 32, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Harkey, M.R.; Henderson, G.L. Effects of herbal components on cDNA-expressed cytochrome P450 enzyme catalytic activity. Life Sci. 2002, 71, 1579–1589. [Google Scholar] [CrossRef]
- Saxena, A.; Tripathi, K.P.; Roy, S.; Khan, F.; Sharma, A. Pharmacovigilance: Effects of herbal components on human drugs interactions involving cytochrome P450. Bioinformation 2008, 3, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Reanmongkol, W.; Keawpradub, N.; Sawangjaroen, K. Effects of the extracts from Mitragyna speciosa Korth. Leaves on analgesic and behavioral activities in experimental animals. Songklanakarin J. Sci. Technol. 2007, 29, 39–48. [Google Scholar]
- Ruan, X. Drug-related side effects of long-term intrathecal morphine therapy. Pain Physician 2007, 10, 357–366. [Google Scholar] [PubMed]
- Matsumoto, K.; Horie, S.; Ishikawa, H.; Takayama, H.; Aimi, N.; Ponglux, D.; Watanabe, K. Antinociceptive effect of 7-hydroxymitragynine in mice: Discovery of an orally active opioid analgesic from the Thai medicinal herb Mitragyna speciosa. Life Sci. 2004, 74, 2143–2155. [Google Scholar] [CrossRef] [PubMed]
- Philipp, A.A.; Wissenbach, D.K.; Zoerntlein, S.W.; Klein, O.N.; Kanogsunthornrat, J.; Maurer, H.H. Studies on the metabolism of mitragynine, the main alkaloid of the herbal drug Kratom, in rat and human urine using liquid chromatography-linear ion trap mass spectrometry. J. Mass Spectrom. 2009, 44, 1249–1261. [Google Scholar] [CrossRef] [PubMed]
- Beckett, A.H.; Shellard, E.J.; Phillipson, J.D.; Lee, L.M. Alkaloids from Mitragyna speciosa Korth. J. Pharm. Pharmacol. 1965, 17, 753–755. [Google Scholar] [CrossRef] [PubMed]
- Cornish-Bowden, A. A simple graphical method for determining the inhibition constants of mixed uncompetitive and non-competitive inhibitors. Biochem. J. 1974, 137, 143–144. [Google Scholar] [CrossRef] [PubMed]
- Henderson, L.; Yue, Q.Y.; Bergquist, C.; Gerden, B.; Arlett, P. St John’s wort (Hypericum perforatum): Drug interactions and clinical outcomes. J. Clin. Pharmacol. 2002, 54, 349–356. [Google Scholar] [CrossRef]
- Azizi, J.; Ismail, S.; Mordi, M.N.; Ramanathan, S.; Said, M.I.; Mansor, S.M. In vitro and in vivo effects of three different Mitragyna speciosa Korth leaf extracts on phase II drug metabolizing enzymes—glutathione transferases (GSTs). Molecules 2010, 15, 432–441. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cytochrome/Substrate | This study | Reference | ||||
|---|---|---|---|---|---|---|
| Km (µM) | IC50 (µM) | Km (µM) a | IC50 (µM) a,b | |||
| 3A4/BFC | 48.94 | 0.045 | 50 | 0.06 ± 0.016 b 0.09 ± 0.03 a | ||
| 2D6/AMMC | 1.016 | 0.005 | 1.5 | 0.0069 ± 0.0006 b 0.5 ± 0.2 a | ||
| 1A2/CEC | 23.69 | 0.755 | 25 | 0.58 ± 0.05 b 1.4 ± 0.3 a | ||
| 2C19/CEC | 4.627 | 2.511 | 5 | 2 ± 0.5 b 3.2 ± 1.2 a | ||
| Cytochrome P450 | IC50 (µg/mL) | Ki (µg/mL) | Mode of inhibition |
|---|---|---|---|
| 3A4/BFC | 0.78 | 1.526 a | Non competitive |
| 2D6/AMMC | 0.636 | 2.6 a | Competitive |
| 1A2/CEC | 39 | 18.57 a | Non competitive |
| 2C19/CEC | Nd b | 84.88 a | Non competitive |
| CYP3A4 | CYP1A2 | CYP2C19 | CYP2D6 | |
|---|---|---|---|---|
| Substrate (final) | BFC: 50 µM | CEC: 5 µM | CEC: 25 µM | AMMC: 1.5 µM |
| Enzyme | 1.0 pmol/well | 0.5 pmol/well | 0.5 pmol/well | 1.5 pmol/well |
| Standard inhibitor | Ketoconazole | Furafylline | Tranylcypromine | Quinidine |
| Phosphate buffer | 200 mM | 100 mM | 50 mM | 100 mM |
| Fluorescence Filter | Ex: 409 nm | Ex: 409 nm | Ex: 409 nm | Ex: 390 nm |
| Em: 530 nm | Em: 460 nm | Em: 460 nm | Em: 460 nm | |
| Temperature | 30 °C | 25 °C | 30 °C | 30 °C |
| Incubation time | 30 min | 20 min | 30 min | 30 min |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kong, W.M.; Chik, Z.; Ramachandra, M.; Subramaniam, U.; Aziddin, R.E.R.; Mohamed, Z. Evaluation of the Effects of Mitragyna speciosa Alkaloid Extract on Cytochrome P450 Enzymes Using a High Throughput Assay. Molecules 2011, 16, 7344-7356. https://doi.org/10.3390/molecules16097344
Kong WM, Chik Z, Ramachandra M, Subramaniam U, Aziddin RER, Mohamed Z. Evaluation of the Effects of Mitragyna speciosa Alkaloid Extract on Cytochrome P450 Enzymes Using a High Throughput Assay. Molecules. 2011; 16(9):7344-7356. https://doi.org/10.3390/molecules16097344
Chicago/Turabian StyleKong, Wai Mun, Zamri Chik, Murali Ramachandra, Umarani Subramaniam, Raja Elina Raja Aziddin, and Zahurin Mohamed. 2011. "Evaluation of the Effects of Mitragyna speciosa Alkaloid Extract on Cytochrome P450 Enzymes Using a High Throughput Assay" Molecules 16, no. 9: 7344-7356. https://doi.org/10.3390/molecules16097344