Terpenoids as Potential Anti-Alzheimer’s Disease Therapeutics

1
Department of Biological Sciences, College of Advanced Science, Dankook University, San#29, Anseo-dong, Dongnam-gu, Cheonan 330-714, Korea
2
Laboratory of Pharmacognosy, College of Pharmacy, Dankook University, San#29, Anseo-dong, Dongnam-gu, Cheonan 330-714, Korea
*
Author to whom correspondence should be addressed.
Molecules 2012, 17(3), 3524-3538; https://doi.org/10.3390/molecules17033524
Submission received: 7 December 2011
/
Revised: 12 March 2012
/
Accepted: 16 March 2012
/
Published: 19 March 2012
(This article belongs to the Special Issue Terpenoids)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Alzheimer’s disease (AD) is one of the most well-known neurodegenerative diseases and explains 50–60% of dementia in patients. The prevalence rate of AD is positively correlated with age and AD affects ≥40% of those over 85 years old. The major AD therapeutics available on the market are acetylcholinesterase inhibitors, such as tacrine and donepezil. New therapeutic agents that can block the disease-inducing mechanisms are essential. Diverse efforts have been made to discover anti-AD agents from natural sources. In this review article, we describe some representative terpenoids such as ginsenosides, gingkolides, and canabinoids as potential anti-AD agents. These compounds exhibit promising in vitro and in vivo biological activities, but are still waiting clinical trials. Additionally, we also discuss some terpenoids including cornel iridoid glycoside, oleanolic acid, tenuifolin, cryptotanshinone, and ursolic acid, which are under investigation for their in vitro and in vivo animal studies.
1. Introduction
The development of medicine and science has contributed to a dramatic increase in life expectancy worldwide and the average life span may reach 120 years old by 2050. The increasing proportion of the elderly will also be reflected in a marked increase in the number of age-related diseases, including neurodegenerative diseases.
Alzheimer’s disease (AD) is one of the most well-known neurodegenerative diseases, and explains 50–60% of patients with dementia. The prevalence rate of AD is positively correlated with age, and AD occurs in ≥40% of the elderly over 85 years old [1]. Patients with AD decline in cognitive function and find it difficult to remember recent events during the early stage (short-term memory loss). Once the disease progresses, patients experience difficulties in speech, speaking, and cognitive thinking, which is accompanied by long-term memory loss. Patients suffer from language deficits, depression, aggressive behavior, and psychosis during the late stage and eventually need total care from caregivers.
One of the pathologic hallmarks of AD is senile plaques (SPs), and the major constituent of SPs is beta-amyloid (Aβ), which is surrounded by dystrophic neurites and microglia and accumulates outside of neurons. Aβ is a product of sequential proteolytic cleavage of amyloid precursor protein (APP) by β-secretase and γ-secretase [2]. Aβ accumulates in the brain of patients with AD due to increased production or decreased clearance of Aβ. The overproduction of Aβ found in patients with AD who have genetic mutations in APP (familiar AD) is correlated with early onset (beginning in the 30s) of the disease. An increased amount of Aβ (soluble monomeric form) in the brain self-aggregates into Aβ oligomers (2–6 Aβ peptides) [3,4], which is more toxic to cells than the fibrillar or monomeric form [5]. Therefore, the excess toxic Aβ is the major cause of AD pathology (amyloid hypothesis) [6]. Particularly, the levels of Aβ oligomers correlate with the severity of cognitive impairment in patients with AD and play a critical role in AD pathology [7]. Aggregated Aβ oligomers lead to synaptic dysfunction due to oxidative stress and inflammation [8,9]. A recent study reported that Aβ induces neuronal death by binding to nerve growth factor receptors [10] such as pan neurotrophin receptor (p75NTR) and activation of downstream c-Jun N-terminal kinase signal [11]. In addition, activation of the N-methyl-D-aspartate (NMDA)-type glutamate receptor (NMDAR) disrupts calcium homeostasis, eventually inducing oxidative stress and synaptic loss [12,13]. Aβ oligomers bind and modulate presynaptic P/Q-type calcium channels at glutaminergic and gamma-amino butyric acid-ergic synapses and eventually impair P/Q current, which is important for neurotransmission and synaptic plasticity [14,15].
Hyperphosphorylated tau proteins accumulate inside of neurons as a form of paired helical filaments (PHFs), which are known as neurofibrillary tangles (NFTs) [16,17]. NFTs are another pathological hallmark of AD. Tau protein, normally present in neurons, binds to microtubules to promote microtubule assembly and stabilizes microtubules and vesicle transport. Conversely, abnormal hyperphosphorylation to tau significantly reduces the affinity of tau protein to microtubules, so hyperphosphorylated tau aggregates and forms PHFs [18]. Inhibited Wnt signaling following binding to the Frizzled receptor, a Wnt protein acceptor, induces neurotoxic hyper-phosphorylated tau proteins, which are found in NFTs [19]. A report that levels of hyperphosphorylated tau protein in cerebrospinal fluid correlate with the degree of cognitive impairment in patients with AD [20] supports the importance of NFTs in AD pathology. Disease-relevant phosphorylation of tau protein and the aggregation to PHF is induced by Aβ [21,22,23,24]. Additionally, tau phosphorylation is the limiting factor in Aβ-induced cell death [25]. These results support the suggestion that tau phosphorylation plays a critical role in AD progression induced by Aβ.
The major AD therapeutics available on the market are acetylcholinesterase (AChE) inhibitors. A strong correlation exists between the degree of cognitive impairment and a shortage of acetylcholine (ACh) in patients with AD [26]. AChE inhibitors such as tacrine, donepezil, rivastigmine, and galantamine have been developed as pharmacotherapy for AD. Although AChE inhibitors help alleviate AD symptoms, they do not delay disease progression. Therefore, new therapeutic agents that block the disease-inducing mechanisms are essential. Memantine, a NMDA receptor antagonist, has been approved by the U.S. Food and Drug Administration (FDA) to treat AD [27]. This drug improves language function and overall cognitive ability significantly in patients with moderate to severe AD [28,29].
Natural products have been used for medicinal purposes for a long time. The effort to develop natural products as potential therapeutics and advances in extraction and isolation techniques lead to the development of 63% of the natural product-derived drugs from 1981–2006 [30]. Much research effort has been devoted to the development of anti-AD agents from natural sources [31]. Galantamine isolated from bulbs and flowers of snowdrop Galanthus woronowii (Amaryllidaceae) has been approved by the FDA as an anti-AD medication due to its inhibitory effect against AChE. Terpenoids such as ginsenosides in Panax ginseng (Araliaceae) have been extensively studied to understand their beneficial effects on AD. Among many natural products, the terpenoids are the largest and the most diverse group of naturally occurring organic compounds, which increases the chance that a terpenoid will be identified as having activity against AD. This review article describes some terpenoids as possible therapeutic agents to treat AD.
2. Terpenoids
2.1. Ginsenosides from Panax ginseng CA Meyer (Araliaceae)
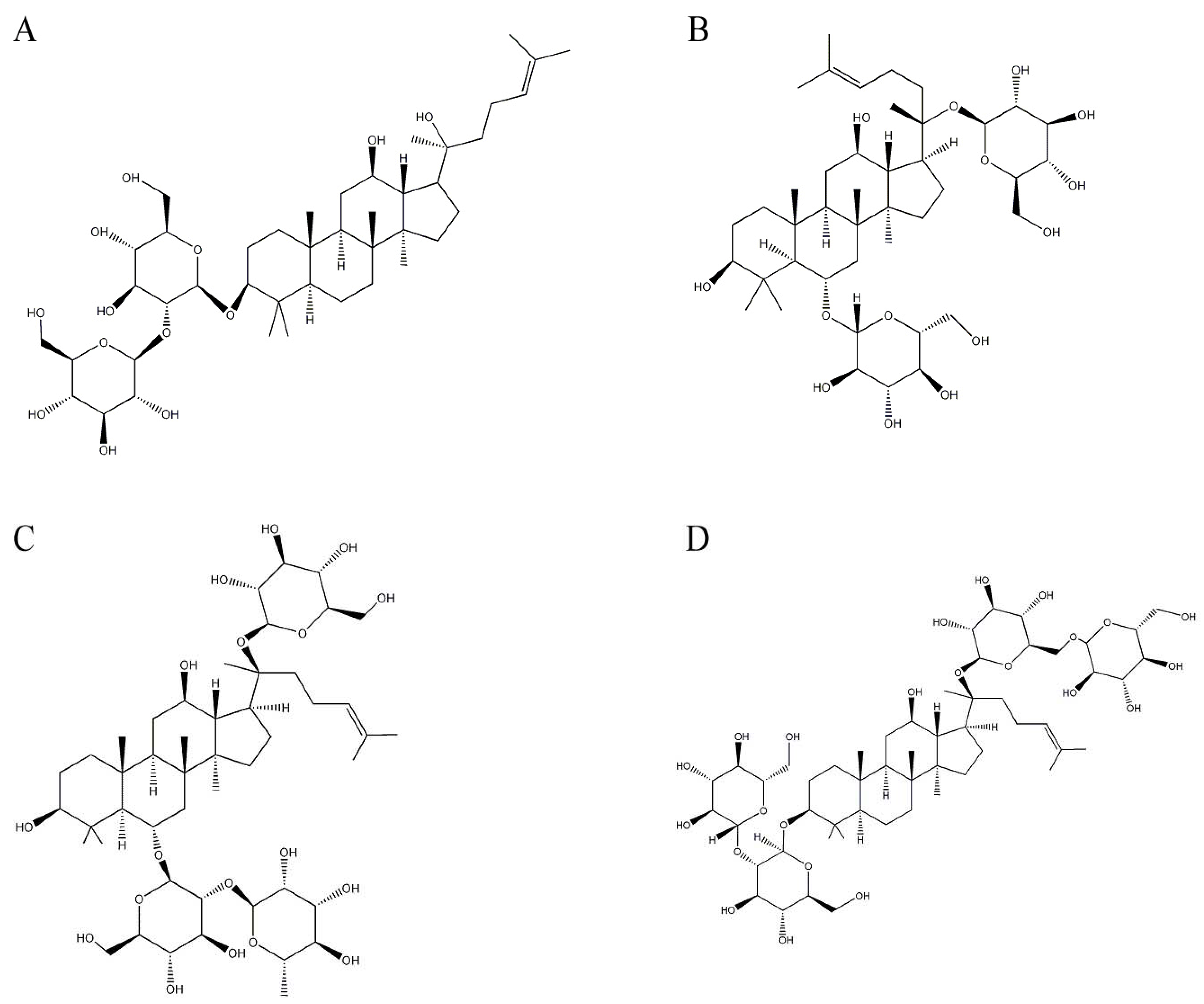
Ginsenosides are a series of derivatives of the dammarane-type triterpenes with some sugar moieties attached [32,33], which are the major active components in ginseng isolated from P. ginseng. P. ginseng is a well-known traditional medicinal plant that has been used as a representative tonic for thousands of years to promote health and longevity. Among diverse ginsenosides in ginseng extract, ginsenoside Rg3 (Figure 1A) significantly reduces the production of Aβ in CHO2B7 cells by 84% and in Tg2576 transgenic mice by 31% [34]. Ginsenoside Rg3 reduces Aβ levels by promoting Aβ degradation and by enhancing neprilysin gene expression, which is a rate-limiting enzyme in Aβ degradation [35]. Furthermore, ginseng attenuates learning deficits in the damaged or aging brains of rodents [34,36]. Additionally, ginsenoside Rg1 (Figure 1B) attenuates the amount of accumulated Aβ and improves cognitive performance in a transgenic mouse model by activating the protein kinase A/cAMP response element binding protein signaling pathway [37,38]. Ginsenoside Rg1 also reduces Aβ production by modulating the APP process, which is accompanied by an improvement in cognitive function [39,40].
Another ginsenoside in ginseng extract, ginsenoside Re (Figure 1C), protects PC12 cells against Aβ-induced neurotoxicity [39]. In addition, ginsenoside Rb1 (Figure 1D) reverses Aβ-induced memory loss in rats by attenuating neuroinflammation markers in the hippocampus [41]. Ginsenoside Rb1 also exhibits beneficial effects on spatial learning by increasing synaptic density in the brain [42].
Figure 1.
Structures of ginsenosides in Panax ginseng. (A) Ginsenoside Rg3; (B) ginsenoside Rg1; (C) ginsenoside Re; and (D) ginsenoside Rb1.
Figure 1.
Structures of ginsenosides in Panax ginseng. (A) Ginsenoside Rg3; (B) ginsenoside Rg1; (C) ginsenoside Re; and (D) ginsenoside Rb1.

Ginseng extracts have been studied in clinical trials for their biological efficacy. However, the clinical trial results are complicated by different extraction methods and even different ginseng species used in the different trials. Clinical studies with a placebo-controlled, double-blind, balanced, crossover design have identified both positive and negative effects of ginseng. A 400 mg dose of ginseng extract provides the most beneficial effects in terms of enhanced cognitive function (quality of memory) in a placebo-controlled, double-blind, balanced, crossover design of 20 young healthy adults who are administered a single dose of 200, 400, or 600 mg ginseng (G115) [43,44]. Although these studies have reported significant benefits of ginseng extracts on cognitive function, the small sample sizes limit the certainty of the results. Another clinical trial administers ginseng powder (4.5 g/d) daily for 12 weeks to 58 patients with AD as the treatment group and 39 patients with AD as a placebo control group [45]. Cognitive performance is monitored using the Mini-Mental State Examination (MMSE) score and the Alzheimer’s Disease Assessment Scale (ADAS) during 12 weeks of ginseng treatment. The ginseng group shows gradually improved MMSE and ADAS scores over the 12 weeks of treatment, whereas the control group shows gradually declined MMSE and ADAS scores, suggesting the beneficial effect of ginseng extracts on cognitive function and memory enhancement. However, the beneficial effect of ginseng extract on memory declines gradually to the level of the control group during a 12 week follow-up period without treatment.
2.2. Ginkgolides and Bilobalide from Gingko biloba L. (Ginkgoaceae)
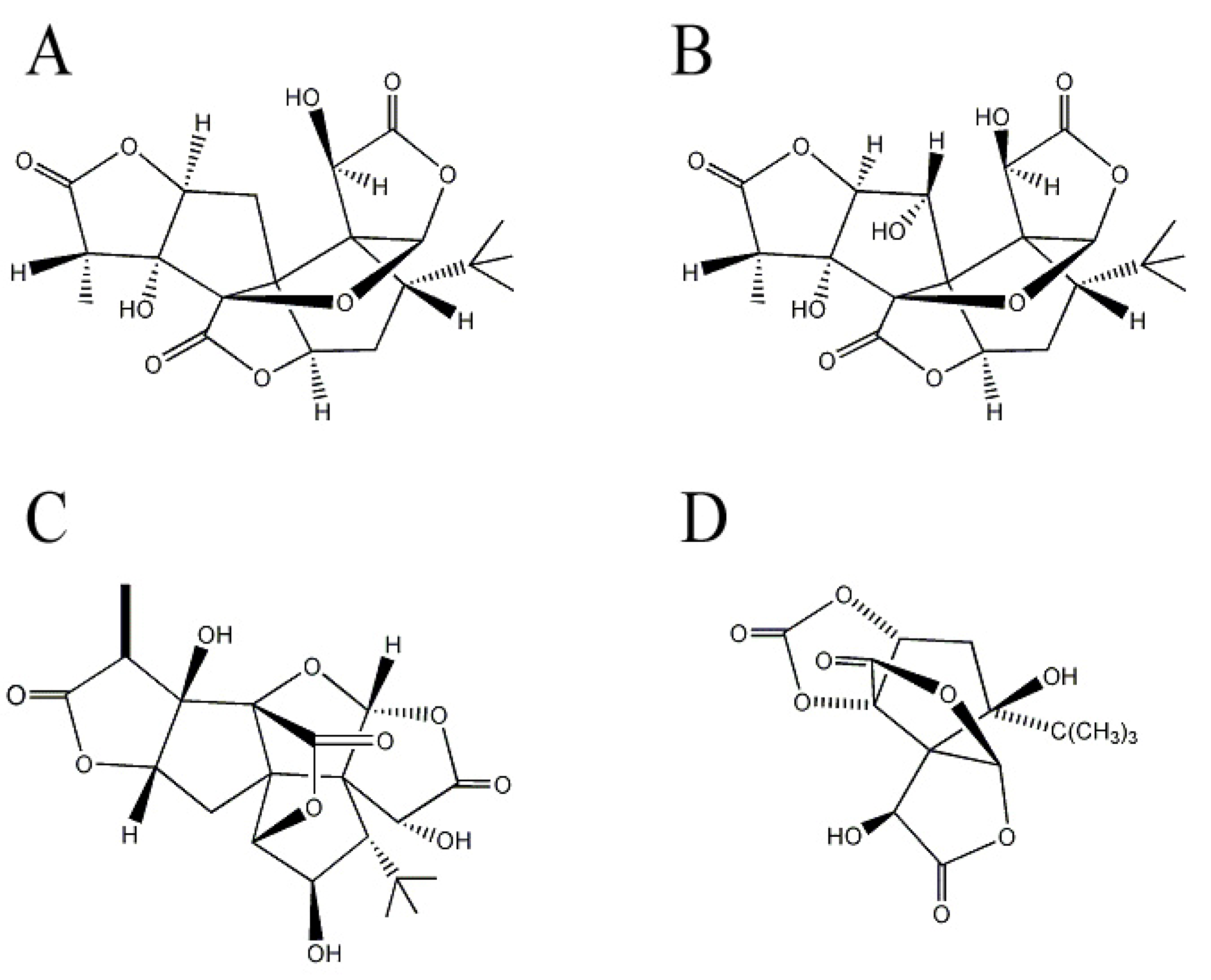
Ginkgolides are a cyclic diterpenes of labdane type commonly isolated from G. biloba. EGb761, extract of G. biloba leaves which contains 24% flavonoid glycosides, 6% terpenoids, and 5–10% organic acids [46] has been extensively evaluated for its neuroprotective effects [47], and terpene trilactones ginkgolides are the major pharmacologically active constituents in EGb761. For example, pre-treatment of neuronal cells with ginkgolide A and B (Figure 2A,B) protects neuronal cells from synaptic damage evaluated by the loss of synaptophysin, a presynaptic synaptic marker [48] and increases neuronal survival against Aβ-induced toxicity [49]. Ginkgolide B rescues hippocampal neurons from Aβ-induced apoptosis by increasing the production of brain-derived neurotrophic factor [50] and reduces apoptotic death of neuronal cells in hemorrhagic rat brain [51]. In transgenic Caenorhabditis elegans, ginkgolide A alleviates Aβ-induced adverse behavior including paralysis [52]. Ginkgolide B reverses the Aβ-induced reduction of ACh release from hippocampal brain slices, suggesting potential improvements in learning and memory deteriorated by Aβ [53]. Furthermore, Vitolo et al. reports that ginkgolide J (Figure 2C) is the most inhibitor of Aβ-induced hippocampal neuronal cell death among the ginkgolides in EGb761 [54]. Additionally, bilobalide (Figure 2D) reduces Aβ-induced synaptic loss and subsequently enhances hippocampal neurogenesis and synaptogenesis [55]. Bilobalide also rescues chick embryonic neurons from apoptosis induced by serum deprivation or staurosporine treatment [56,57].
Figure 2.
Structures of ginkgolides and bilobalide in Gingko biloba. (A) Ginkgolide A; (B) ginkgolide B; (C) ginkgolide J; and (D) bilobalide.
Figure 2.
Structures of ginkgolides and bilobalide in Gingko biloba. (A) Ginkgolide A; (B) ginkgolide B; (C) ginkgolide J; and (D) bilobalide.

Despite these previous findings, regarding the neuroprotective effects of ginkgolides and bilobalides in EGB761 and EGB761 itself, its clinical efficacy is inconsistent and remains controversial. Therefore, clinical trials reflecting diverse races, dementia severity, and different doses of EGb761 should be performed to evaluate the consistency of the effects of EGb761 against AD.
2.3. Cannabinoids from Cannabis sativa L. (Cannabaceae)
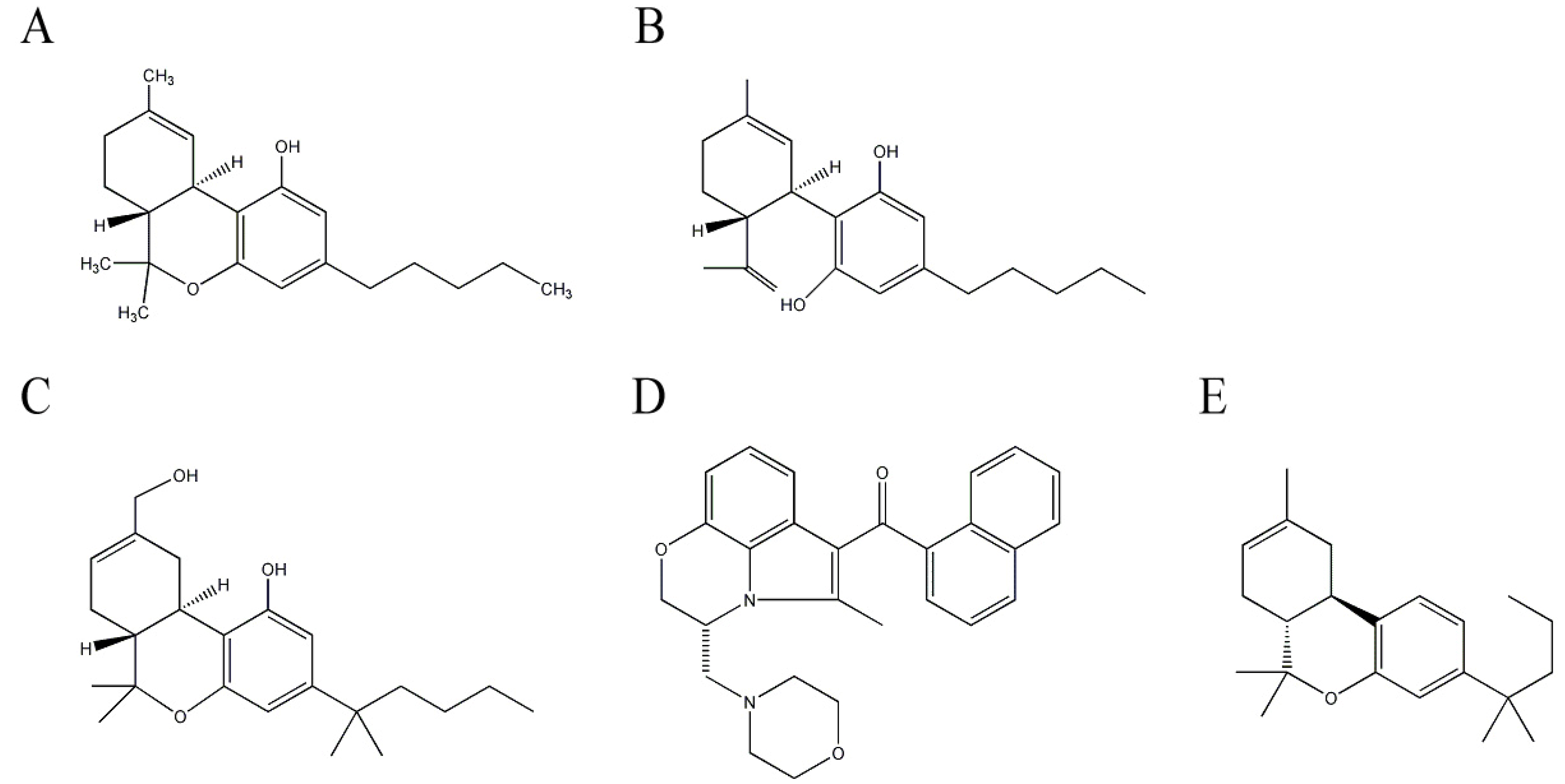
Cannabinoids are aromatic compounds containing a monoterpene moiety derived from isoprene units, which are isolated from C. sativa in the Cannabaceae family. The diverse pharmacological activities of cannabinoids are mediated by activating specific cannabinoid receptors including CB1 and CB2 [58]. CB1 receptors, which mediate the psychoactive properties, are expressed in the nervous system such as neurons and glial cells [59], whereas CB2 receptors are mainly located in immune cells [60]. Δ9-Tetrahydrocannabinol (THC) (Figure 3A) mainly binds to CB1 receptors. This plant contains about 60 cannabinoids, and one of the potential anti-AD agents is THC, the major cannabinoid in C. sativa. THC is one of the widely-studied natural products and has anti-emetic, anti-convulsive, anti-inflammatory, and analgesic effects [61]. A protective effect of THC against AD has been reported. THC comparatively inhibits AChE and increases the availability of ACh. In addition, THC reduces the inhibition of AChE-induced Aβ aggregation, and subsequently reduces Aβ-induced toxicity [62] and is more efficient than commercially available AChE inhibitors such as tacrine and donepezil. Furthermore, THC reduces behavioral and circadian disturbances in patients with severe dementia [63].
Another major cannabinoid having neuroprotective effects against AD is cannabidiol (CBD) (Figure 3B). CBD comprises about 40% of cannabis extracts and is the principle non-psychotrophic constituent. The strong antioxidant effect of CBD provides neuroprotection by reducing oxidative damage such as lipid peroxidation [64,65]. Furthermore, CBD alleviates Aβ-induced inflammatory signals by reducing nitric oxide production by inhibiting p38 and nuclear factor-κB signaling pathways [66]. Tau hyperphosphorylation, one of pathological hallmarks of AD, is also reduced by CBD treatment, as it reduces glycogen synthase kinase-3β, an enzyme responsible for tau hyperphosphorylation in patients with AD [67]. The neuroprotective effects of CBD have been confirmed in an AD-mouse model induced with an intrahippocampal injection of Aβ (1–42) by a reduction in glial activated pro-inflammatory mediators [68,69]. Because CBD lacks psychoactive properties, it is one of the attractive potential anti-AD targets.
Additionally, some synthetic cannabinoids such as HU-210, WIN55, 212-2, and JWH-133 (Figure 3C–E), greatly reduce microglial activation and cytokine production in Aβ-administered rats. Consequently, these synthetic compounds alleviate cognitive impairment by reducing the decrease in neuronal marker levels [69,70,71]. These reports suggest that cannabinoids, particularly THC and CBD, have potential to be developed as anti-AD therapeutics.
Figure 3.
Structures of cannabinoids in Cannabis sativa and synthetic cannabinoid derivatives. (A) Δ9-Tetrahydrocannabinol (THC); (B) cannabidiol (CBD); (C) HU-210; (D) WIN55,212-2; and (E) JWH-133.
Figure 3.
Structures of cannabinoids in Cannabis sativa and synthetic cannabinoid derivatives. (A) Δ9-Tetrahydrocannabinol (THC); (B) cannabidiol (CBD); (C) HU-210; (D) WIN55,212-2; and (E) JWH-133.

2.4. Other Terpenoids Having Potential Anti-AD Effects
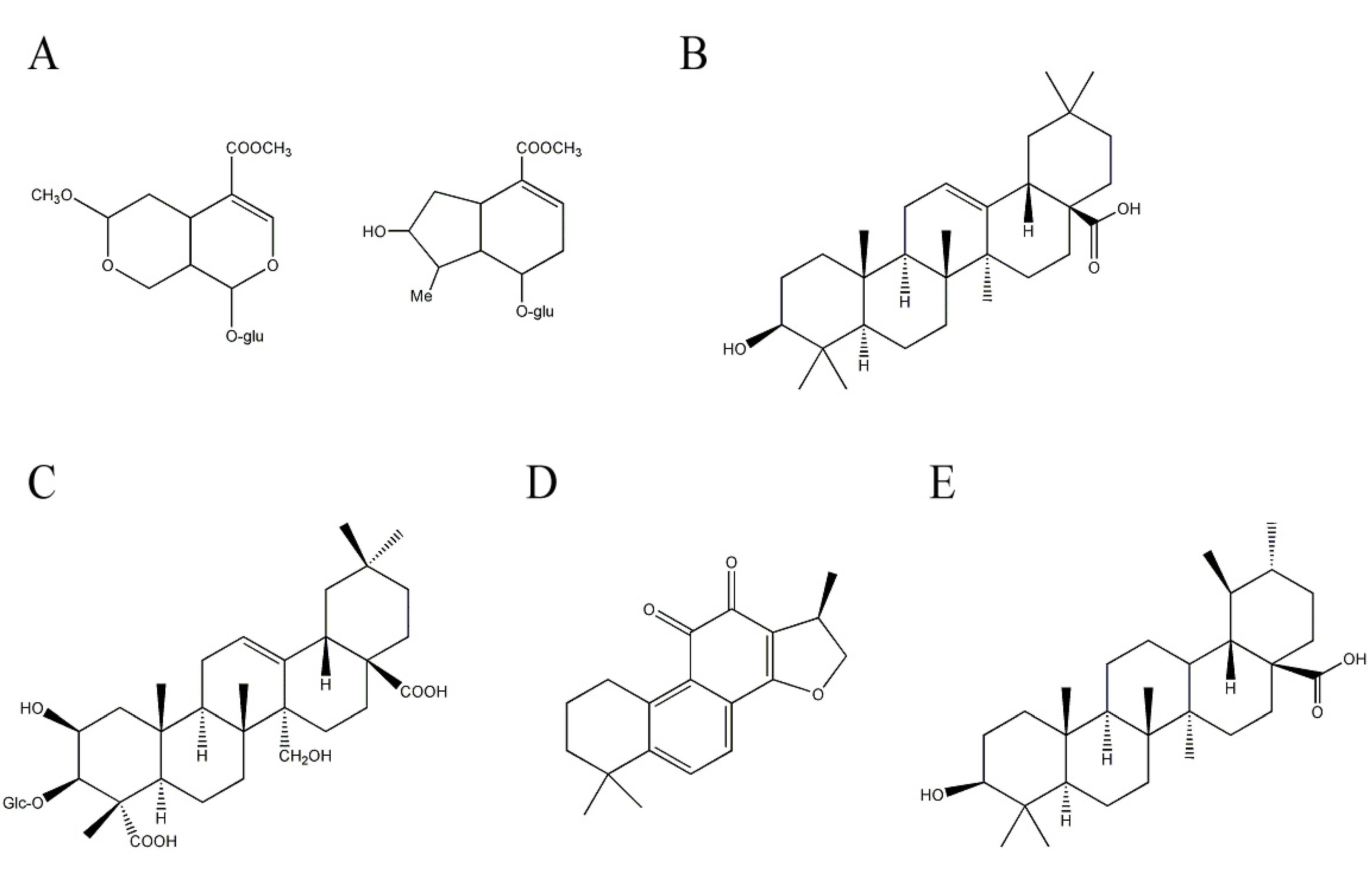
Cornel iridoid glycoside, mainly including morroniside and loganin (Figure 4A) as major compounds in fruits of Cornus officinalis Sieb. Et Zucc. (Cornaceae), has a neuroprotective effect by increasing neurological function and decreasing cerebral infarct size in cerebral ischemic rats [72]. Cornel iridoid glycoside improves memory deficits and attenuates hippocampal neuronal loss by improving the brain environment for repair and promoting neuronal survival in fimbria-fornix transfected rats [73].
Oleanolic acid is a triterpene which has been identified as neuroprotective constituents in Aralia cordata Thunb. (Araliaceae). A. cordata is distributed in eastern Asia such as China, Japan, and Korea and some biological activities including anti-nociceptive, anti-diabetic, anti-oxidant, and anti-inflammatory activities have been reported. Due to its anti-oxidant and anti-inflammatory activities, extracts of A. cordata are tested for their neuroprotective effect against Aβ. An extract of A. cordata rescues neuronal death induced by Aβ in cultured rat cortical neurons and improves Aβ-induced memory deficit in mice. The neuroprotective constituent included in A. cordata is revealed as a triterpene, oleanolic acid (Figure 4B) [74,75].
Tenuifolin is another triterpene that has been reported to be beneficial for AD. It is isolated from Polygala tenuifolia Willd. (Polygalaceae). P. tenuifolia is a well-known traditional Chinese medicine that is frequently used to improve cognitive function. An extract of P. tenuifolia decreases the production of Aβ in in vitro cultured cells [76,77]. Additional effort to identify the responsible constituents in P. tenuifolia leads to the isolation of tenuifolin (Figure 4C [77]. Tenuifolin reduces Aβ secretion by inhibiting β-secretase, one of the enzymes responsible for cleaving APP to Aβ. Furthermore, tenuifolin improves learning and memory in aged mice by decreasing AChE activity accompanied by increased neurotransmitters levels such as norepinephrine and dopamine [78].
Figure 4.
Structures of other terpenoids with anti-Alzheimer’s disease activity. (A) morroniside and loganin (B) oleanolic acid; (C) tenuifolin; (D) cryptotanshinone; and (E) ursolic acid.
Figure 4.
Structures of other terpenoids with anti-Alzheimer’s disease activity. (A) morroniside and loganin (B) oleanolic acid; (C) tenuifolin; (D) cryptotanshinone; and (E) ursolic acid.

Cryptotanshinone (Figure 4D), a labdane-type diterpene, is an active compound that possesses anti-inflammatory, anti-oxidant, and anti-apoptotic activities [79,80,81]. The compound can be isolated from Salvia miltiorrhiza Bunge (Labiatae). Cryptotanshinone easily crosses the blood-brain barrier and affects cognitive function in mice [82]. Furthermore, cryptotanshinone reduces Aβ production by up-regulating α-secretase, which cleaves APPs in the middle of the Aβ sequence, which precludes Aβ production in vivo and in vitro by activating the PI3K pathway [83,84]. In addition, cryptotanshinone protects neuronal cell damage by inhibiting Aβ aggregation [85].
A screening effort to identify potent AChE inhibitors from medicinal herbs leads to the isolation of the triterpene, ursolic acid (Figure 4E) from Origanum majorana L. (Lamiaceae). Ursolic acid effectively inhibits AChE activity in a dose-dependent and competitive/non-competitive manner [86]. Ursolic acid also reduces Aβ-induced oxidative damage such as free radical formation and lipid peroxidation in in vitro assay systems [87]. Ursolic acid inhibits Aβ binding to microglia, reducing the production of proinflammatory cytokines and neurotoxic reactive oxygen species and leading to a neuroprotective effect against Aβ [88].
3. Conclusions
Natural products are attractive sources for developing anti-AD agents, because they can provide diverse structural characteristics and biological activities. Unfortunately, some AChE inhibitors and NMDA receptor antagonists are the only medications approved by the FDA to treat patients with AD. Therefore, this review discussed some natural products and their molecular targets, particularly terpenoids, which can be developed as potential anti-AD agents. The representative terpenoids with anti-AD effects are ginsenosides from P. ginseng, ginkgolides and bilobalide from G. biloba, and cannabinoids from C. sativa. The evaluation of biological activities by in vitro cell based assays and in vivo animal studies indicate the beneficial effects of these compounds against AD. However, their clinical efficacy is still controversial. Clinical trials should be designed to reflect diverse races, dementia severity, and different doses of biologically active compounds.
Other compounds such as cornel iridoid glycoside, oleanolic acid, tenuifolin, cryptotanshinone and ursolic acid have outstanding neuroprotective effects in in vitro assays. These compounds can exert beneficial effects on central nervous system directly or indirectly by acting on peripheral targets. Therefore, the methods to efficiently deliver the bioactive compounds to the brain should be considered to develop terpenoids as anti-AD agents. In addition, the supply of large quantities of biologically active compounds is essential to develop natural product-derived biologically active compounds as therapeutic agents. To overcome this restriction, a mixture of two or three bioactive compounds that act synergistically might be used as the alternative instead of a single compound. Furthermore, a continuous search for bioactive compounds including terpenoids is expected to lead to the discovery of therapeutic agents against AD from natural sources.
Acknowledgments
This research was supported by the Korea Research Foundation Grant funded by the Korean Government (KRF-2010-0003980) to S.-Y. Park.
Conflict of Interest
The authors declare no conflict of interest.
References
- Hebert, L.E.; Scherr, P.A.; Bienias, J.L.; Bennett, D.A.; Evans, D.A. Alzheimer disease in the US population: Prevalence estimates using the 2000 census. Arch. Neurol. 2003, 60, 1119–1122. [Google Scholar] [CrossRef]
- Selkoe, D.J. The cell biology of beta-amyloid precursor protein and presenilin in Alzheimer’s disease. Trends Cell. Biol. 1998, 8, 447–453. [Google Scholar] [CrossRef]
- Selkoe, D.J. Alzheimer’s disease: Genes, proteins, and therapy. Physiol. Rev. 2001, 81, 741–766. [Google Scholar]
- Tanzi, R.E.; Bertram, L. Twenty years of the Alzheimer’s disease amyloid hypothesis: A genetic perspective. Cell 2005, 120, 545–555. [Google Scholar] [CrossRef]
- Walsh, D.M.; Selkoe, D.J. A beta oligomers – a decade of discovery. J. Neurochem. 2007, 101, 1172–1184. [Google Scholar] [CrossRef]
- Kawahara, M.; Kuroda, Y. Molecular mechanism of neurodegeneration induced by Alzheimer’s beta-amyloid protein: Channel formation and disruption of calcium homeostasis. Brain Res. Bull. 2000, 53, 389–397. [Google Scholar] [CrossRef]
- Lue, L.F.; Kuo, Y.M.; Roher, A.E.; Brachova, L.; Shen, Y.; Sue, L.; Beach, T.; Kurth, J.H.; Rydel, R.E.; Rogers, J. Soluble amyloid beta peptide concentration as a predictor of synaptic change in Alzheimer’s disease. Am. J. Pathol. 1999, 155, 853–862. [Google Scholar] [CrossRef]
- Roberson, E.D.; Mucke, L. 100 years and counting: Prospects for defeating Alzheimer’s disease. Science 2006, 314, 781–784. [Google Scholar] [CrossRef]
- Heneka, M.T.; O’Banion, M.K. Inflammatory processes in Alzheimer’s disease. J. Neuroimmunol. 2007, 184, 69–91. [Google Scholar] [CrossRef]
- Yamamoto, N.; Matsubara, E.; Maeda, S.; Minagawa, H.; Takashima, A.; Maruyama, W.; Michikawa, M.; Yanagisawa, K. A ganglioside-induced toxic soluble Abeta assembly. Its enhanced formation from Abeta bearing the Arctic mutation. J. Biol. Chem. 2007, 282, 2646–2655. [Google Scholar]
- Coulson, E.J. Does the p75 neurotrophin receptor mediate Abeta-induced toxicity in Alzheimer’s disease? J. Neurochem. 2006, 98, 654–660. [Google Scholar] [CrossRef]
- de Felice, F.G.; Velasco, P.T.; Lambert, M.P.; Viola, K.; Fernandez, S.J.; Ferreira, S.T.; Klein, W.L. Abeta oligomers induce neuronal oxidative stress through an N-methyl-D-aspartate receptor-dependent mechanism that is blocked by the Alzheimer drug memantine. J. Biol. Chem. 2007, 282, 11590–11601. [Google Scholar]
- Shankar, G.M.; Bloodgood, B.L.; Townsend, M.; Walsh, D.M.; Selkoe, D.J.; Sabatini, B.L. Natural oligomers of the Alzheimer amyloid-beta protein induce reversible synapse loss by modulating an NMDA-type glutamate receptor-dependent signaling pathway. J. Neurosci. 2007, 27, 2866–2875. [Google Scholar]
- Magdesian, M.H.; Carvalho, M.M.; Mendes, F.A.; Saraiva, L.M.; Juliano, M.A.; Juliano, L.; Garcia-Abreu, J.; Ferreira, S.T. Amyloid-beta binds to the extracellular cysteine-rich domain of Frizzled and inhibits Wnt/beta-catenin signaling. J. Biol. Chem. 2008, 283, 9359–9368. [Google Scholar]
- Nimmrich, V.; Grimm, C.; Draguhn, A.; Barghorn, S.; Lehmann, A.; Schoemaker, H.; Hillen, H.; Gross, G.; Ebert, U.; Bruehl, C. Amyloid beta oligomers (A beta(1-42) globulomer) suppress spontaneous synaptic activity by inhibition of P/Q-type calcium currents. J. Neurosci. 2008, 28, 788–797. [Google Scholar] [CrossRef]
- Mezler, M.; Barghorn, S.; Schoemaker, H.; Gross, G.; Nimmrich, V. Abeta oligomer directly modulates P/Q-type calcium currents in Xenopus oocytes. Br. J. Pharmacol. 2012, 165, 1572–1583. [Google Scholar] [CrossRef]
- Kosik, K.S.; Joachim, C.L.; Selkoe, D.J. Microtubule-associated protein tau (tau) is a major antigenic component of paired helical filaments in Alzheimer disease. Proc. Natl. Acad. Sci. USA 1986, 83, 4044–4048. [Google Scholar] [CrossRef]
- Kondo, J.; Honda, T.; Mori, H.; Hamada, Y.; Miura, R.; Ogawara, M.; Ihara, Y. The carboxyl third of tau is tightly bound to paired helical filaments. Neuron 1988, 1, 827–834. [Google Scholar] [CrossRef]
- Iqbal, K.; Liu, F.; Gong, C.X.; Alonso Adel, C.; Grundke-Iqbal, I. Mechanisms of tau-induced neurodegeneration. Acta Neuropathol. 2009, 118, 53–69. [Google Scholar] [CrossRef]
- Wallin, A.K.; Blennow, K.; Andreasen, N.; Minthon, L. CSF biomarkers for Alzheimer’s Disease: levels of beta-amyloid, tau, phosphorylated tau relate to clinical symptoms and survival. Dement. Geriatr. Cogn. Disord. 2006, 21, 131–138. [Google Scholar] [CrossRef]
- Busciglio, J.; Lorenzo, A.; Yeh, J.; Yankner, B.A. beta-amyloid fibrils induce tau phosphorylation and loss of microtubule binding. Neuron 1995, 14, 879–888. [Google Scholar] [CrossRef]
- Greenberg, S.M.; Kosik, K.S. Secreted beta-APP stimulates MAP kinase and phosphorylation of tau in neurons. Neurobiol. Aging 1995, 16, 403–407, discussion 407–408. [Google Scholar] [CrossRef]
- Takashima, A.; Honda, T.; Yasutake, K.; Michel, G.; Murayama, O.; Murayama, M.; Ishiguro, K.; Yamaguchi, H. Activation of tau protein kinase I/glycogen synthase kinase-3beta by amyloid beta peptide (25–35) enhances phosphorylation of tau in hippocampal neurons. Neurosci. Res. 1998, 31, 317–323. [Google Scholar] [CrossRef]
- Zheng, W.H.; Bastianetto, S.; Mennicken, F.; Ma, W.; Kar, S. Amyloid beta peptide induces tau phosphorylation and loss of cholinergic neurons in rat primary septal cultures. Neuroscience 2002, 115, 201–121. [Google Scholar] [CrossRef]
- Leschik, J.; Welzel, A.; Weissmann, C.; Eckert, A.; Brandt, R. Inverse and distinct modulation of tau-dependent neurodegeneration by presenilin 1 and amyloid-beta in cultured cortical neurons: Evidence that tau phosphorylation is the limiting factor in amyloid-beta-induced cell death. J. Neurochem. 2007, 101, 1303–1315. [Google Scholar] [CrossRef]
- Francis, P.T.; Palmer, A.M.; Sims, N.R.; Bowen, D.M.; Davison, A.N.; Esiri, M.M.; Neary, D.; Snowden, J.S.; Wilcock, G.K. Neurochemical studies of early-onset Alzheimer’s disease. Possible influence on treatment. N. Engl. J. Med. 1985, 313, 7–11. [Google Scholar] [CrossRef]
- Lipton, S.A. Pathologically-activated therapeutics for neuroprotection: Mechanism of NMDA receptor block by memantine and S-nitrosylation. Curr. Drug Targets 2007, 8, 621–632. [Google Scholar] [CrossRef]
- Ferris, S.; Ihl, R.; Robert, P.; Winblad, B.; Gatz, G.; Tennigkeit, F.; Gauthier, S. Treatment effects of Memantine on language in moderate to severe Alzheimer’s disease patients. Alzheimers Dement. 2009, 5, 369–374. [Google Scholar] [CrossRef]
- Mecocci, P.; Bladstrom, A.; Stender, K. Effects of memantine on cognition in patients with moderate to severe Alzheimer’s disease: Post-hoc analyses of ADAS-cog and SIB total and single-item scores from six randomized, double-blind, placebo-controlled studies. Int. J. Geriatr. Psychiatry 2009, 24, 532–538. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the last 25 years. J. Nat. Prod. 2007, 70, 461–477. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, L.Q.; Tang, X.C.; Zhang, H.Y. Retrospect and prospect of active principles from Chinese herbs in the treatment of dementia. Acta Pharmacol. Sin. 2010, 31, 649–664. [Google Scholar] [CrossRef]
- Liu, Z.Q.; Luo, X.Y.; Liu, G.Z.; Chen, Y.P.; Wang, Z.C.; Sun, Y.X. In vitro study of the relationship between the structure of ginsenoside and its antioxidative or prooxidative activity in free radical induced hemolysis of human erythrocytes. J. Agric. Food Chem. 2003, 51, 2555–2558. [Google Scholar] [CrossRef]
- Yun, T.K.; Lee, Y.S.; Lee, Y.H.; Kim, S.I.; Yun, H.Y. Anticarcinogenic effect of Panax ginseng C.A. Meyer and identification of active compounds. J. Korean Med. Sci. 2001, 16, S6–S18. [Google Scholar]
- Chen, F.; Eckman, E.A.; Eckman, C.B. Reductions in levels of the Alzheimer’s amyloid beta peptide after oral administration of ginsenosides. FASEB J. 2006, 20, 1269–1271. [Google Scholar] [CrossRef]
- Yang, L.; Hao, J.; Zhang, J.; Xia, W.; Dong, X.; Hu, X.; Kong, F.; Cui, X. Ginsenoside Rg3 promotes beta-amyloid peptide degradation by enhancing gene expression of neprilysin. J. Pharm. Pharmacol. 2009, 61, 375–380. [Google Scholar]
- Zhao, R.; McDaniel, W.F. Ginseng improves strategic learning by normal and brain-damaged rats. Neuroreport 1998, 9, 1619–1624. [Google Scholar] [CrossRef]
- Fang, F.; Chen, X.; Huang, T.; Luddy, J.S.; Yan, S.S. Multi-faced neuroprotective effects of Ginsenoside Rg1 in an Alzheimer mouse model. Biochim. Biophys. Acta 2012, 1822, 286–292. [Google Scholar] [CrossRef]
- Shi, Y.Q.; Huang, T.W.; Chen, L.M.; Pan, X.D.; Zhang, J.; Zhu, Y.G.; Chen, X.C. Ginsenoside Rg1 attenuates amyloid-beta content, regulates PKA/CREB activity, and improves cognitive performance in SAMP8 mice. J. Alzheimers Dis. 2010, 19, 977–989. [Google Scholar]
- Liang, W.; Ge, S.; Yang, L.; Yang, M.; Ye, Z.; Yan, M.; Du, J.; Luo, Z. Ginsenosides Rb1 and Rg1 promote proliferation and expression of neurotrophic factors in primary Schwann cell cultures. Brain Res. 2010, 1357, 19–25. [Google Scholar] [CrossRef]
- Chen, L.M.; Lin, Z.Y.; Zhu, Y.G.; Lin, N.; Zhang, J.; Pan, X.D.; Chen, X.C. Ginsenoside Rg1 attenuates beta-amyloid generation via suppressing PPARgamma-regulated BACE1 activity in N2a-APP695 cells. Eur. J. Pharmacol. 2012, 675, 15–21. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, J.; Zhang, Z.; Bi, P.; Qi, Z.; Zhang, C. Anti-neuroinflammation effect of ginsenoside Rbl in a rat model of Alzheimer disease. Neurosci. Lett. 2011, 487, 70–72. [Google Scholar] [CrossRef]
- Mook-Jung, I.; Hong, H.S.; Boo, J.H.; Lee, K.H.; Yun, S.H.; Cheong, M.Y.; Joo, I.; Huh, K.; Jung, M.W. Ginsenoside Rb1 and Rg1 improve spatial learning and increase hippocampal synaptophysin level in mice. J. Neurosci. Res. 2001, 63, 509–515. [Google Scholar] [CrossRef]
- Kennedy, D.O.; Scholey, A.B.; Wesnes, K.A. Dose dependent changes in cognitive performance and mood following acute administration of Ginseng to healthy young volunteers. Nutr. Neurosci. 2001, 4, 295–310. [Google Scholar]
- Reay, J.L.; Scholey, A.B.; Kennedy, D.O. Panax ginseng (G115) improves aspects of working memory performance and subjective ratings of calmness in healthy young adults. Hum. Psychopharmacol. 2010, 25, 462–471. [Google Scholar] [CrossRef]
- Lee, S.T.; Chu, K.; Sim, J.Y.; Heo, J.H.; Kim, M. Panax ginseng enhances cognitive performance in Alzheimer disease. Alzheimer Dis. Assoc. Disord. 2008, 22, 222–226. [Google Scholar] [CrossRef]
- Le Bars, P.L. Magnitude of effect and special approach to Ginkgo biloba extract EGb 761® in cognitive disorders. Pharmacopsychiatry 2003, 36, 44–49. [Google Scholar] [CrossRef]
- Shi, C.; Zhao, L.; Zhu, B.; Li, Q.; Yew, D.T.; Yao, Z.; Xu, J. Protective effects of Ginkgo biloba extract (EGb761) and its constituents quercetin and ginkgolide B against beta-amyloid peptide-induced toxicity in SH-SY5Y cells. Chem. Biol. Interact. 2009, 181, 115–123. [Google Scholar] [CrossRef]
- Bate, C.; Tayebi, M.; Williams, A. Ginkgolides protect against amyloid-beta1-42-mediated synapse damage in vitro. Mol. Neurodegener. 2008, 3, 1. [Google Scholar] [CrossRef]
- Bate, C.; Salmona, M.; Williams, A. Ginkgolide B inhibits the neurotoxicity of prions or amyloid-beta1-42. J. Neuroinflammation 2004, 1, 4. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Q.; Wang, C.; Li, J.; Hou, Q.; Ma, J.; Wang, W.; Wang, Z. Ginkgolide B protects hippocampal neurons from apoptosis induced by beta-amyloid 25-35 partly via up-regulation of brain-derived neurotrophic factor. Eur. J. Pharmacol. 2010, 647, 48–54. [Google Scholar] [CrossRef]
- Hu, Y.Y.; Huang, M.; Dong, X.Q.; Xu, Q.P.; Yu, W.H.; Zhang, Z.Y. Ginkgolide B reduces neuronal cell apoptosis in the hemorrhagic rat brain: Possible involvement of Toll-like receptor 4/nuclear factor-kappa B pathway. J. Ethnopharmacol. 2011, 137, 1462–1468. [Google Scholar] [CrossRef]
- Wu, Y.; Wu, Z.; Butko, P.; Christen, Y.; Lambert, M.P.; Klein, W.L.; Link, C.D.; Luo, Y. Amyloid-beta-induced pathological behaviors are suppressed by Ginkgo biloba extract EGb 761 and ginkgolides in transgenic Caenorhabditis elegans. J. Neurosci. 2006, 26, 13102–13113. [Google Scholar] [CrossRef]
- Lee, T.F.; Chen, C.F.; Wang, L.C. Effect of ginkgolides on beta-amyloid-suppressed acetylocholine release from rat hippocampal slices. Phytother. Res. 2004, 18, 556–560. [Google Scholar] [CrossRef]
- Vitolo, O.; Gong, B.; Cao, Z.; Ishii, H.; Jaracz, S.; Nakanishi, K.; Arancio, O.; Dzyuba, S.V.; Lefort, R.; Shelanski, M. Protection against beta-amyloid induced abnormal synaptic function and cell death by Ginkgolide J. Neurobiol. Aging 2009, 30, 257–265. [Google Scholar] [CrossRef]
- Tchantchou, F.; Lacor, P.N.; Cao, Z.; Lao, L.; Hou, Y.; Cui, C.; Klein, W.L.; Luo, Y. Stimulation of neurogenesis and synaptogenesis by bilobalide and quercetin via common final pathway in hippocampal neurons. J. Alzheimers Dis. 2009, 18, 787–798. [Google Scholar]
- Ahlemeyer, B.; Mowes, A.; Krieglstein, J. Inhibition of serum deprivation- and staurosporine-induced neuronal apoptosis by Ginkgo biloba extract and some of its constituents. Eur. J. Pharmacol. 1999, 367, 423–430. [Google Scholar] [CrossRef]
- Defeudis, F.V. Bilobalide and neuroprotection. Pharmacol. Res. 2002, 46, 565–568. [Google Scholar] [CrossRef]
- Howlett, A.C.; Barth, F.; Bonner, T.I.; Cabral, G.; Casellas, P.; Devane, W.A.; Felder, C.C.; Herkenham, M.; Mackie, K.; Martin, B.R.; et al. International Union of Pharmacology. XXVII. Classification of cannabinoid receptors. Pharmacol. Rev. 2002, 54, 161–202. [Google Scholar] [CrossRef]
- Herkenham, M.; Lynn, A.B.; Little, M.D.; Johnson, M.R.; Melvin, L.S.; de Costa, B.R.; Rice, K.C. Cannabinoid receptor localization in brain. Proc. Natl. Acad. Sci. USA 1990, 87, 1932–1936. [Google Scholar]
- Pertwee, R.G. Cannabinoid pharmacology: The first 66 years. Br. J. Pharmacol. 2006, 147, S163–S171. [Google Scholar] [CrossRef]
- Carlini, E.A. The good and the bad effects of (−) trans-delta-9-tetrahydrocannabinol (Delta 9-THC) on humans. Toxicon 2004, 44, 461–467. [Google Scholar] [CrossRef]
- Eubanks, L.M.; Rogers, C.J.; Beuscher, A.E., IV; Koob, G.F.; Olson, A.J.; Dickerson, T.J.; Janda, K.D. A molecular link between the active component of marijuana and Alzheimer’s disease pathology. Mol. Pharm. 2006, 3, 773–777. [Google Scholar] [CrossRef]
- Walther, S.; Mahlberg, R.; Eichmann, U.; Kunz, D. Delta-9-tetrahydrocannabinol for nighttime agitation in severe dementia. Psychopharmacology (Berl) 2006, 185, 524–528. [Google Scholar] [CrossRef]
- Hampson, A.J.; Grimaldi, M.; Axelrod, J.; Wink, D. Cannabidiol and (-)Delta9-tetrahydrocannabinol are neuroprotective antioxidants. Proc. Natl. Acad. Sci. USA 1998, 95, 8268–8273. [Google Scholar] [CrossRef]
- Iuvone, T.; Esposito, G.; Esposito, R.; Santamaria, R.; di Rosa, M.; Izzo, A.A. Neuroprotective effect of cannabidiol, a non-psychoactive component from Cannabis sativa, on beta-amyloid-induced toxicity in PC12 cells. J. Neurochem. 2004, 89, 134–141. [Google Scholar] [CrossRef]
- Esposito, G.; de Filippis, D.; Maiuri, M.C.; de Stefano, D.; Carnuccio, R.; Iuvone, T. Cannabidiol inhibits inducible nitric oxide synthase protein expression and nitric oxide production in beta-amyloid stimulated PC12 neurons through p38 MAP kinase and NF-kappaB involvement. Neurosci. Lett. 2006, 399, 91–95. [Google Scholar] [CrossRef]
- Esposito, G.; de Filippis, D.; Carnuccio, R.; Izzo, A.A.; Iuvone, T. The marijuana component cannabidiol inhibits beta-amyloid-induced tau protein hyperphosphorylation through Wnt/beta-catenin pathway rescue in PC12 cells. J. Mol. Med. (Berl) 2006, 84, 253–258. [Google Scholar] [CrossRef]
- Esposito, G.; Scuderi, C.; Savani, C.; Steardo, L., Jr.; de Filippis, D.; Cottone, P.; Iuvone, T.; Cuomo, V.; Steardo, L. Cannabidiol in vivo blunts beta-amyloid induced neuroinflammation by suppressing IL-1beta and iNOS expression. Br. J. Pharmacol. 2007, 151, 1272–1279. [Google Scholar]
- Martin-Moreno, A.M.; Reigada, D.; Ramirez, B.G.; Mechoulam, R.; Innamorato, N.; Cuadrado, A.; de Ceballos, M.L. Cannabidiol and other cannabinoids reduce microglial activation in vitro and in vivo: Relevance to Alzheimer’s disease. Mol. Pharmacol. 2011, 79, 964–973. [Google Scholar] [CrossRef]
- Ramirez, B.G.; Blazquez, C.; Gomez del Pulgar, T.; Guzman, M.; de Ceballos, M.L. Prevention of Alzheimer’s disease pathology by cannabinoids: Neuroprotection mediated by blockade of microglial activation. J. Neurosci. 2005, 25, 1904–1913. [Google Scholar] [CrossRef]
- Martin Moreno, A.M.; Brera, B.; Spuch, C.; Carro, E.; Garcia-Garcia, L.; Delgado, M.; Pozo, M.A.; Innamorato, N.G.; Cuadrado, A.; de Ceballos, M.L. Prolonged oral cannabinoid administration prevents neuroinflammation, lowers beta-amyloid levels and improves cognitive performance in Tg APP 2576 mice. J. Neuroinflamm. 2012, 9, 8. [Google Scholar] [CrossRef] [Green Version]
- Li, C.Y.; Li, L.; Li, Y.H.; Ai, H.X.; Zhang, L. Effects of extract from Cornus officinalis on nitric oxide and NF-kappaB in cortex of cerebral infarction rat model. Zhongguo Zhong Yao Za Zhi 2005, 30, 1667–1670. [Google Scholar]
- Zhao, L.H.; Ding, Y.X.; Zhang, L.; Li, L. Cornel iridoid glycoside improves memory ability and promotes neuronal survival in fimbria-fornix transected rats. Eur. J. Pharmacol. 2010, 647, 68–74. [Google Scholar] [CrossRef]
- Cho, S.O.; Ban, J.Y.; Kim, J.Y.; Jeong, H.Y.; Lee, I.S.; Song, K.S.; Bae, K.; Seong, Y.H. Aralia cordata protects against amyloid beta protein (25-35)-induced neurotoxicity in cultured neurons and has antidementia activities in mice. J. Pharmacol. Sci. 2009, 111, 22–32. [Google Scholar] [CrossRef]
- Cho, S.O.; Ban, J.Y.; Kim, J.Y.; Ju, H.S.; Lee, I.S.; Song, K.S.; Bae, K.; Seong, Y.H. Anti-ischemic activities of aralia cordata and its active component, oleanolic acid. Arch. Pharm. Res. 2009, 32, 923–932. [Google Scholar] [CrossRef]
- Jia, H.; Jiang, Y.; Ruan, Y.; Zhang, Y.; Ma, X.; Zhang, J.; Beyreuther, K.; Tu, P.; Zhang, D. Tenuigenin treatment decreases secretion of the Alzheimer’s disease amyloid beta-protein in cultured cells. Neurosci. Lett. 2004, 367, 123–128. [Google Scholar] [CrossRef]
- Lv, J.; Jia, H.; Jiang, Y.; Ruan, Y.; Liu, Z.; Yue, W.; Beyreuther, K.; Tu, P.; Zhang, D. Tenuifolin, an extract derived from tenuigenin, inhibits amyloid-beta secretion in vitro. Acta Physiol. (Oxf) 2009, 196, 419–425. [Google Scholar] [CrossRef]
- Zhang, H.; Han, T.; Zhang, L.; Yu, C.H.; Wan, D.G.; Rahman, K.; Qin, L.P.; Peng, C. Effects of tenuifolin extracted from radix polygalae on learning and memory: A behavioral and biochemical study on aged and amnesic mice. Phytomedicine 2008, 15, 587–594. [Google Scholar] [CrossRef]
- Kim, S.Y.; Moon, T.C.; Chang, H.W.; Son, K.H.; Kang, S.S.; Kim, H.P. Effects of tanshinone I isolated from Salvia miltiorrhiza bunge on arachidonic acid metabolism and in vivo inflammatory responses. Phytother. Res. 2002, 16, 616–620. [Google Scholar] [CrossRef]
- Ng, T.B.; Liu, F.; Wang, Z.T. Antioxidative activity of natural products from plants. Life Sci. 2000, 66, 709–723. [Google Scholar] [CrossRef]
- Park, E.J.; Zhao, Y.Z.; Kim, Y.C.; Sohn, D.H. PF2401-SF, standardized fraction of Salvia miltiorrhiza and its constituents, tanshinone I, tanshinone IIA, and cryptotanshinone, protect primary cultured rat hepatocytes from bile acid-induced apoptosis by inhibiting JNK phosphorylation. Food Chem. Toxicol. 2007, 45, 1891–1898. [Google Scholar] [CrossRef]
- Kim, D.H.; Jeon, S.J.; Jung, J.W.; Lee, S.; Yoon, B.H.; Shin, B.Y.; Son, K.H.; Cheong, J.H.; Kim, Y.S.; Kang, S.S.; et al. Tanshinone congeners improve memory impairments induced by scopolamine on passive avoidance tasks in mice. Eur. J. Pharmacol. 2007, 574, 140–147. [Google Scholar] [CrossRef]
- Mei, Z.; Zhang, F.; Tao, L.; Zheng, W.; Cao, Y.; Wang, Z.; Tang, S.; Le, K.; Chen, S.; Pi, R.; et al. Cryptotanshinone, a compound from Salvia miltiorrhiza modulates amyloid precursor protein metabolism and attenuates beta-amyloid deposition through upregulating alpha-secretase in vivo and in vitro. Neurosci. Lett. 2009, 452, 90–95. [Google Scholar] [CrossRef]
- Mei, Z.; Situ, B.; Tan, X.; Zheng, S.; Zhang, F.; Yan, P.; Liu, P. Cryptotanshinione upregulates alpha-secretase by activation PI3K pathway in cortical neurons. Brain Res. 2010, 1348, 165–173. [Google Scholar] [CrossRef]
- Mei, Z.; Yan, P.; Situ, B.; Mou, Y.; Liu, P. Cryptotanshinione inhibits beta-amyloid aggregation and protects damage from beta-amyloid in SH-SY5Y cells. Neurochem. Res. 2012, 37, 622–628. [Google Scholar] [CrossRef]
- Chung, Y.K.; Heo, H.J.; Kim, E.K.; Kim, H.K.; Huh, T.L.; Lim, Y.; Kim, S.K.; Shin, D.H. Inhibitory effect of ursolic acid purified from Origanum majorana L on the acetylcholinesterase. Mol. Cells 2001, 11, 137–143. [Google Scholar]
- Heo, H.J.; Cho, H.Y.; Hong, B.; Kim, H.K.; Heo, T.R.; Kim, E.K.; Kim, S.K.; Kim, C.J.; Shin, D.H. Ursolic acid of Origanum majorana L. reduces Abeta-induced oxidative injury. Mol. Cells 2002, 13, 5–11. [Google Scholar]
- Wilkinson, K.; Boyd, J.D.; Glicksman, M.; Moore, K.J.; El Khoury, J. A high content drug screen identifies ursolic acid as an inhibitor of amyloid beta protein interactions with its receptor CD36. J. Biol. Chem. 2011, 286, 34914–34922. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Yoo, K.-Y.; Park, S.-Y. Terpenoids as Potential Anti-Alzheimer’s Disease Therapeutics. Molecules 2012, 17, 3524-3538. https://doi.org/10.3390/molecules17033524
AMA Style
Yoo K-Y, Park S-Y. Terpenoids as Potential Anti-Alzheimer’s Disease Therapeutics. Molecules. 2012; 17(3):3524-3538. https://doi.org/10.3390/molecules17033524
Chicago/Turabian StyleYoo, Ki-Yeol, and So-Young Park. 2012. "Terpenoids as Potential Anti-Alzheimer’s Disease Therapeutics" Molecules 17, no. 3: 3524-3538. https://doi.org/10.3390/molecules17033524