Associations of nm23H1, VEGF-C, and VEGF-3 Receptor in Human Prostate Cancer

Abstract
:1. Introduction
2. Results and Discussion
2.1. Results



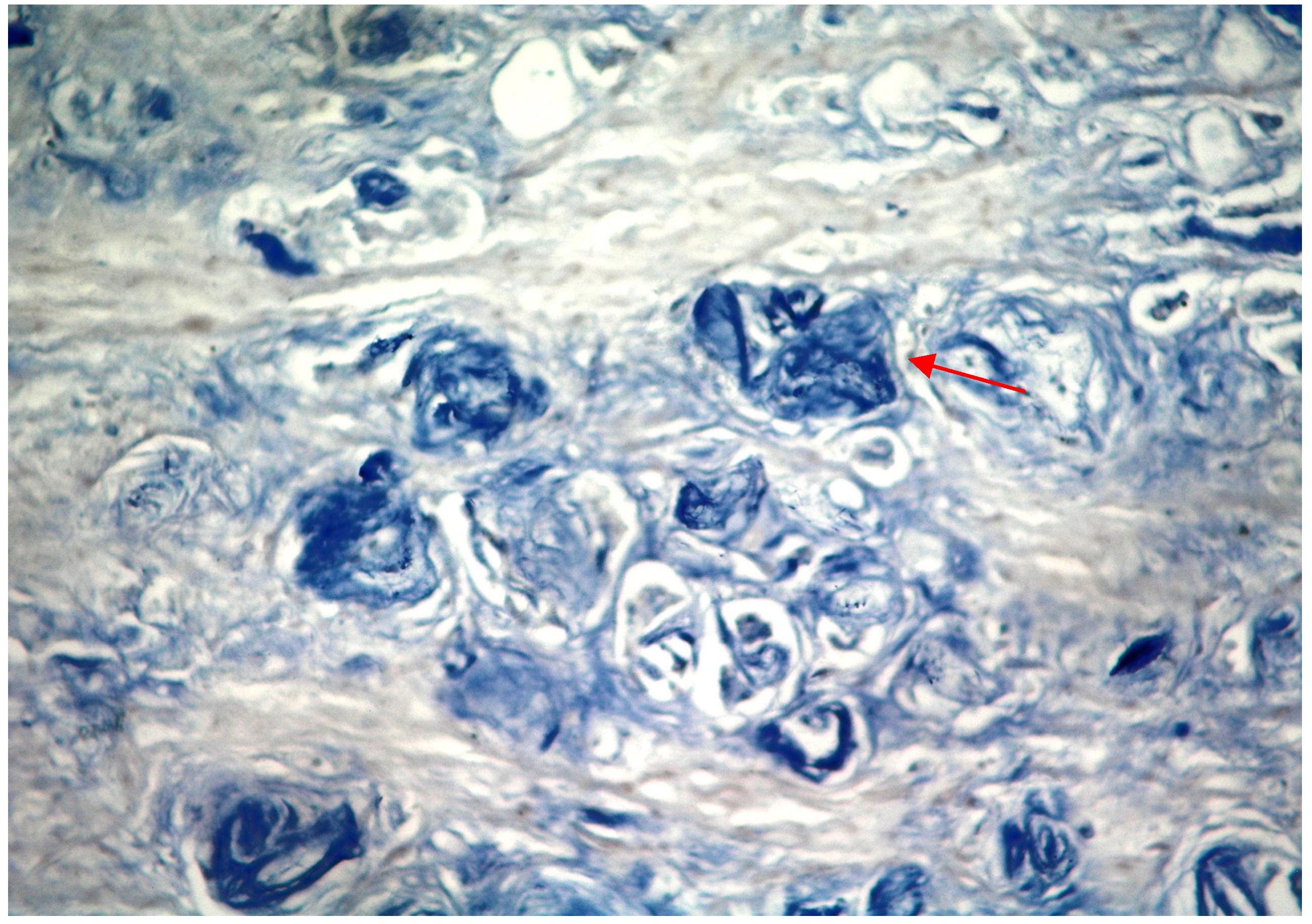{kind=link}
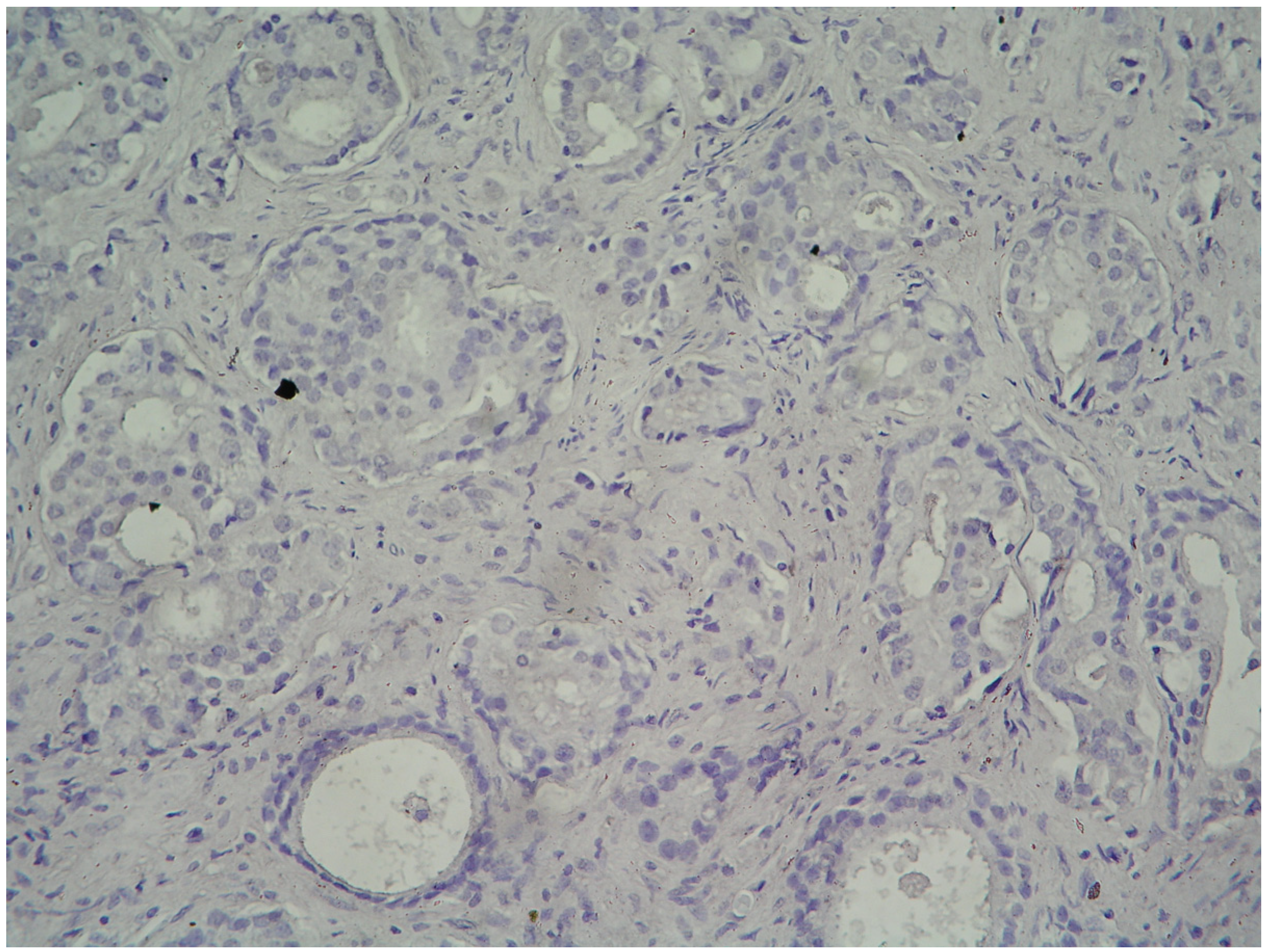{kind=link}
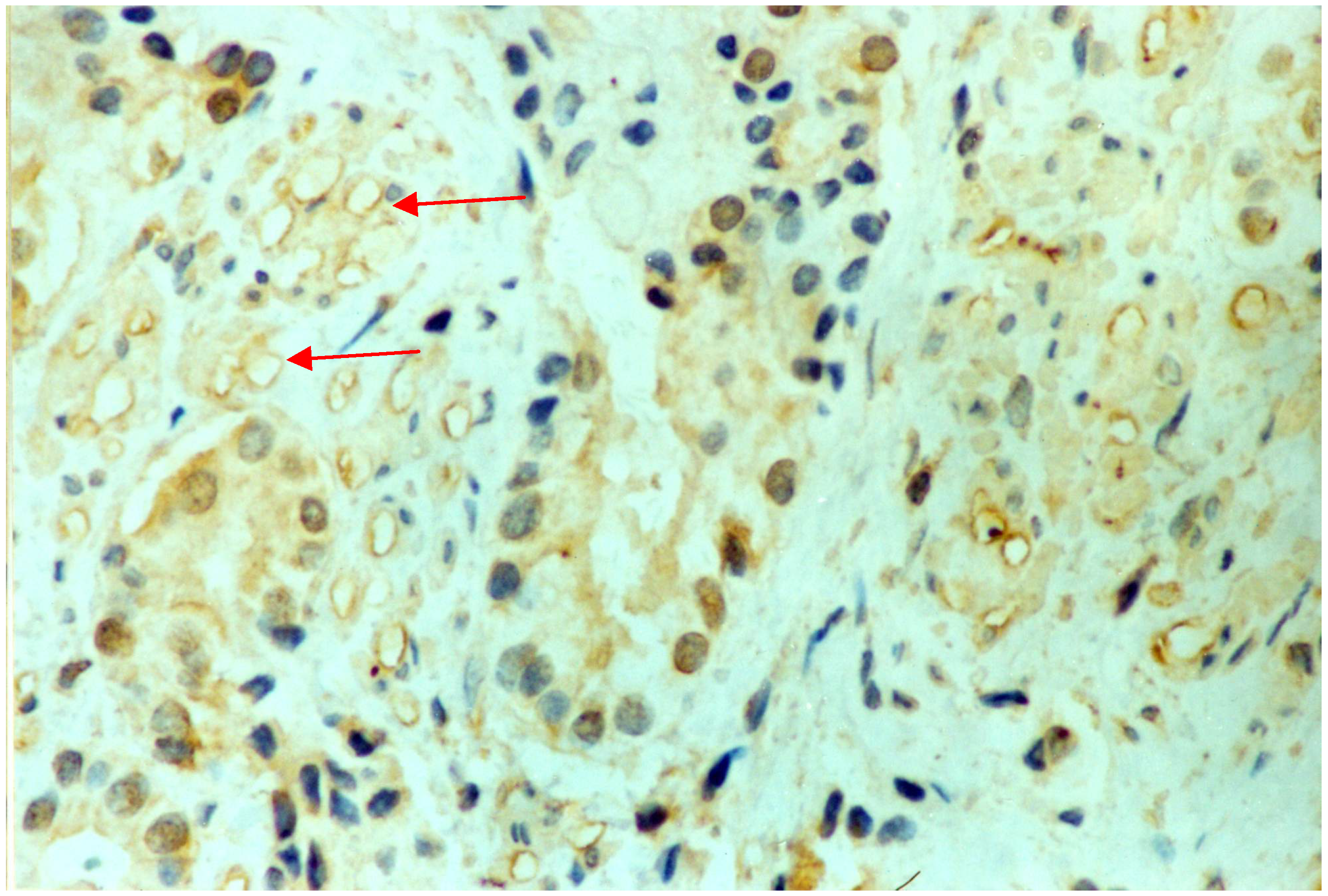{kind=link}
{kind=link}
| Characteristics | Cases (%) |
|---|---|
| Age (years) | |
| <65 | 16 (38.10%) |
| ≥65 | 26 (61.90%) |
| Lymph node metastasis | |
| N0 | |
| N1a | 31 (73.81%) |
| N1b | 11 (26.19%) |
| TNM clinical stage | |
| Stage I | 12 (28.57%) |
| Stage II | 10 (23.81%) |
| Stage III | 9 (21.43%) |
| Stage IV | 11 (26.19%) |
| Gleason score | |
| ≤4 | 7 (16.67%) |
| 5–6 | 9 (21.42%) |
| 7 | 19 (45.24%) |
| ≥8 | 7 (16.67%) |
| Pathologic grade | |
| Grade I | 12 (28.57%) |
| Grade II | 9 (21.43%) |
| Grade III | 11 (26.19%) |
| Grade IV | 10 (23.81%) |
| Clinicopathological | No. of cases | nm23H1-positive cases (%) | VEGF-C-positive cases (%) | MLC /mm2 mean ± dev |
|---|---|---|---|---|
| adjacent benign tissues | 4.51 ± 2.64 | |||
| Pathologic grade | ||||
| I and II | 21 | 6 (28.27) | 9 (42.86) | 8.61 ± 2.67 |
| III and IV | 21 | 13 (61.91) | 10 (47.62) | 7.92 ± 2.04 |
| TNM stage | ||||
| I and II | 22 | 20 (90.91) | 7 (31.82) | 7.45 ± 2.91 |
| III and IV | 20 | 9 (45.00) ** | 12 (60.00) * | 9.79 ± 2.68 * |
| Lymph node metastasis | ||||
| Negative | 31 | 25 (80.65) | 8 (25.81) | 6.93 ± 1.80 |
| Positive | 11 | 4 (36.36) ** | 11 (100) ** | 11.16 ± 1.39 * |


| No. of cases | 5-year survival ratecase (%) | 5-year death rate case (%) | |
|---|---|---|---|
| 42 | 29 | 13 | |
| MLC | 5.73 ± 2. 09 mm2 | 10.82 ± 2.51 mm2 | |
| nm23H1 | |||
| − and + | 19 | 9 (47.37) | 10 (52.63) |
| ++ and +++ | 23 | 20 (86.96) | 3 (13.04) |
| VEGF-CmRNA | |||
| negative | 23 | 22 (95.65) | 1 (4.35) |
| positive | 19 | 7 (36.84) | 12 (63.16) |
| Pathologic grade | |||
| I and II | 21 | 17 (80.95) | 4 (19.05) |
| III and IV | 21 | 12 (57.14) | 9 (42.86) |
2.2. Discussion
3. Experimental
3.1. Tissue Samples
3.2. In Situ Hybridization
3.3. Immunohistochemistry
3.4. Microscopic Assessment of VEGF-C mRNA, VEFGR-3, nm23H1 Expressions and MLC
3.5. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Abate-Shen, C.; Shen, M.M. Molecular genetics of prostate cancer. Genes Dev. 2000, 14, 2410–2434. [Google Scholar] [CrossRef]
- Jankevicius, F.; Miller, S.M.; Ackermann, R. Nutrition and risk of prostate cancer. Urol. Int. 2002, 68, 69–80. [Google Scholar] [CrossRef]
- Wilson, S.; Jones, L.; Couseens, C.; Hanna, K. Cancer and the Environment: Gene-Environment Interactions; The National Academies Press: Washington, DC, USA, 2002. [Google Scholar]
- Rastogi, T.; Hildesheim, A.; Sinha, R. Opportunities for cancer epidemiology in developing countries. Nat. Rev. Cancer 2004, 4, 909–917. [Google Scholar]
- Clinton, S.K. Diet, nutrition, and prostate cancer. Annu. Rev. Nutr. 1998, 18, 413–440. [Google Scholar] [CrossRef]
- Denis, L.; Morton, M.S.; Griffiths, K. Diet and its preventive role in prostatic disease. Eur. Urol. 1999, 35, 5–6. [Google Scholar]
- Weidner, N.; Carroll, P.R.; Flax, J.; Blumenfeld, W.; Folkman, J. Tumor angiogenesis correlates with metastasis in invasive prostate carcinoma. Am. J. Pathol. 1993, 143, 401–409. [Google Scholar]
- Viloria-Petit, A.; Miquerol, L.; Yu, J.L.; Gertsenstein, M.; Sheehan, C.; May, L.; Henkin, J.; Lobe, C.; Nagy, A.; Kerbel, R.S.; et al. Contrasting effects of VEGF gene disruption in embryonic stem cell-derived versus oncogene-induced tumors. EMBO J. 2003, 22, 4091–4102. [Google Scholar] [CrossRef]
- Kerbel, R.S. Tumor angiogenesis: Past, present and the near future. Carcinogenesis 2000, 21, 505–515. [Google Scholar] [CrossRef]
- Neufeld, G.; Cohen, T.; Gengrinovitch, S.; Poltorak, Z. Vascular endothelial growth factor (VEGF) and its receptors. FASEB J. 1999, 13, 9–22. [Google Scholar]
- Kong, D.; Li, Y.; Wang, Z.; Banerjee, S.; Sarkar, F.H. Inhibition of angiogenesis and invasion by 3,3'-Diindolylmethane is mediated by the nuclear factor-{kappa}B downstream target genes MMP-9 and uPA that regulated bioavailability of vascular endothelial growth factor in prostate cancer. Cancer Res. 2007, 67, 3310–3319. [Google Scholar] [CrossRef]
- Jackson, M.; Bentel, J.; Tilley, W. Vascular endothelial growth factor (VEGF) expression in prostate cancer and benign prostatic hyperplasia. J. Urol. 1997, 157, 2323–2328. [Google Scholar] [CrossRef]
- Ferrer, F.; Miller, L.; Andrawis, R.; Kurtzman, S.; Albertsen, P.; Laudone, V.; Kreutzer, D. Vascular endothelial growth factor (VEGF) expression in human prostate cancer: In situ and in vitro expression of VEGF by human prostate cancer cells. J. Urol. 1997, 157, 2323–2328. [Google Scholar] [CrossRef]
- Balbay, M.D.; Pettaway, C.A.; Kuniyasu, H.; Inoue, K.; Ramirez, E.; Li, E.; Fidler, I.J.; Dinney, C.P.N. Highly metastatic human prostate cancer growing within the prostate of athymic mice overexpresses vascular endothelial growth factor. Cancer Res. 1999, 4, 783–789. [Google Scholar]
- Duque, J.L.F.; Loughlin, K.R.; Adam, R.M.; Kantoff, P.W.; Zurakowski, D.; Freeman, M.R. Plasma levels of vascular endothelial growth factor are increased in patients with metastatic prostate cancer. Urology 1999, 3, 523–527. [Google Scholar]
- Strohmeyer, D.; Rossing, C.; Bauerfeind, A.; Kaufmann, O.; Schlechte, H.; Bartsch, G.; Loening, S. Vascular endothelial growth factor and its correlation with angiogenesis and p53 expression in prostate cancer. Prostate 2000, 45, 216–224. [Google Scholar] [CrossRef]
- Mazzucchelli, R.; Montironi, R.; Santinelli, A.; Lucarini, G.; Pugnaloni, A.; Biagini, G. Vascular endothelial growth factor expression and capillary architecture in high-grade PIN and prostate cancer in untreated and androgen-ablated patients. Prostate 2000, 45, 72–79. [Google Scholar]
- Krupski, T.; Harding, M.A.; Herce, M.E.; Gulding, K.M.; Stoler, M.H.; Theodorescu, D. The role of vascular endothelial growth factor in the tissue specific in vivo growth of prostate cancer cells. Growth Factors 2001, 18, 287–302. [Google Scholar] [CrossRef]
- Jackson, M.W.; Roberts, J.S.; Heckford, S.E.; Ricciardelli, C.; Stahl, J.; Horsfall, D.J.; Tilley, W.D. A potential autocrine role for vascular endothelial growth factor in prostate cancer. Cancer Res. 2002, 62, 854–859. [Google Scholar]
- Peyromaure, M.; Camparo, P.; Badoual, C.; Descazeaud, A.; Dinh-Xuan, A. The expression of vascular endothelial growth factor is associated with the risk of cancer progression after radical prostatectomy. BJU Int. 2007, 99, 1150–1153. [Google Scholar] [CrossRef]
- Rinaldo, F.; Li, J.; Wang, E.; Muders, M.; Datta, K. RalA regulates vascular endothelial growth factor-C(VEGF-C) synthesis in prostate cancer cells during androgen ablation. Oncogene 2007, 26, 1731–1738. [Google Scholar] [CrossRef]
- Cao, Y.; Linden, P.; Farnebo, J.; Cao, R.; Eriksson, A.; Kumar, V.; Qi, J.H.; Claesson-Welsh, L.; Alitalo, K. Vascular endothelial growth factor C induces angiogenesis in vivo. Proc. Natl. Acad. Sci. USA 1998, 95, 14389–14394. [Google Scholar] [CrossRef]
- Saaristo, A.; Partanen, T.A.; Arola, J.; Jussila, L.; Hytönen, M.; Mäkitie, A.; Vento, S.; Kaipainen, A.; Malmberg, H.; Alitalo, K. Vascular endothelial growth factor-C and its receptor VEGFR-3 in the nasal mucosa and in nasopharyngeal tumors. Am. J. Pathol. 2000, 157, 7–14. [Google Scholar] [CrossRef]
- Yang, J.; Wu, H.F.; Qian, L.X.; Zhang, W.; Hua, L.X.; Yu, M.L.; Wang, Z.; Xu, Z.Q.; Sui, Y.G.; Wang, X.R. Increased expressions of vascular endothelial growth factor (VEGF), VEGF-C and VEGF receptor-3 in prostate cancer tissue are associated with tumor progression. Asian J. Androl. 2006, 8, 169–175. [Google Scholar]
- Griend, D.J.V.; Berger, J.C.; Rinker-Schaeffer, C.W. Suppression of metastasis—A new function for known proteins. J. Natl. Cancer Inst. 2004, 96, 344–345. [Google Scholar] [CrossRef]
- Kauffman, E.C.; Robinson, V.; Stadler, W.M.; Sokoloff, M.H.; Rinker-Schaeffer, C.W. Metastasis suppression: The evolving role of metastasis suppressor genes for regulating cancer cell growth at the secondary site. J. Urol. 2003, 169, 1122–1133. [Google Scholar] [CrossRef]
- Jaeger, E.B.; Samant, R.S.; Rinker-Schaeffer, C.W. Metastasis suppression in prostate cancer. Cancer Metast. Rev. 2001, 20, 279–286. [Google Scholar] [CrossRef]
- Rinker-Schaeffer, C.W.; O’Keefe, J.P.; Welch, D.R.; Theodorescu, D. Metastasis suppressor proteins: Discovery, molecular mechanisms, and clinical application. Clin. Cancer Res. 2006, 12, 3882–3889. [Google Scholar] [CrossRef]
- Saintigny, P.; Kambouchner, M.; Ly, M.; Gomes, N.; Sainte-Catherine, O.; Vassy, R.; Czernichow, S.; Letoumelin, P.; Breau, J.; Bernaudin, J.; et al. Vascular endothelial growth factor-C and its receptor VEGFR-3 in non-small-cell lung cancer: Concurrent expression in cancer cells from primary tumour and metastatic lymph node. Lung Cancer 2007, 58, 205–213. [Google Scholar] [CrossRef]
- Stacker, S.A.; Achen, M.G.; Jussila, L.; Baldwin, M.E.; Alitalo, K. Lymphangiogenesis and cancer metastases. Nat. Rev. Cancer 2002, 2, 573–583. [Google Scholar] [CrossRef]
- He, Y.; Rajantie, I.; Pajusola, K.; Jeltsch, M.; Holopainen, T.; Yla-Herttuala, S.; Harding, T.; Jooss, K.; Takahashi, T.; Alitalo, K. Vascular endothelial cell growth factor receptor 3-mediated activation of lymphatic endothelium is crucial for tumor cell entry and spread via lymphatic vessels. Cancer Res. 2005, 65, 4739–4746. [Google Scholar] [CrossRef]
- Salven, P.; Lymboussaki, A.; Heikkila, P.; Saari, H.J.; Enholm, B.; Aase, K.; von-Euler, G.; Eriksson, U.; Alitalo, K.; Joensuu, H. Vascular endothelial growth factors VEGF-B and VEGF-C are expressed in human tumors. Am. J. Pathol. 1998, 153, 103–108. [Google Scholar] [CrossRef]
- Valtola, R.; Salven, P.; Heikkila, P.; Taipale, J.; Joensuu, H.; Rehn, M.; Pihlajaniemi, T.; Weich, H.; de Waal, R.; Alitalo, K. VEGFR-3 and its ligand VEGF-C are associated with angiogenesis in breast cancer. Am. J. Pathol. 1999, 154, 1381–1390. [Google Scholar] [CrossRef]
- Steeg, P.S.; Bevilacqua, G.; Kopper, L.; Thorgeirsson, U.P.; Talmadge, J.E.; Liotta, L.A.; Sobel, M.E. Evidence for a novel gene associated with low tumor metastatic potential. J. Natl. Cancer Inst. 1988, 80, 200–204. [Google Scholar] [CrossRef]
- Garinis, G.A.; Manolis, E.N.; Spanakis, N.E.; Patrinos, G.P.; Peros, G.; Menounos, P.G. High frequency of concomitant nm23-H1 and E-cadherin transcriptional inactivation in primary non-inheriting colorectal carcinomas. J. Mol. Med. 2003, 81, 256–263. [Google Scholar]
- Guetz, G.D.; Uzzan, B.; Nicolas, P.; Cucherat, M.; Morere, J.F.; Benamouzig, R.; Breau, J.-L.; Perret, G.-Y. Microvessel density and VEGF expression are prognostic factors in colorectal cancer. Meta-analysis of the literature. Br. J. Cancer 2006, 94, 1823–1832. [Google Scholar] [CrossRef]
- Akagi, K.; Ikeda, Y.; Miyazaki, M.; Abe, T.; Kinoshita, J.; Maehara, Y.; Sugimachi, K. Vascular endothelial growth factor-C (VEGF-C) expression in human colorectal cancer tissues. Br. J. Cancer 2000, 83, 887–891. [Google Scholar] [CrossRef]
- Weidner, N.; Semple, J.P.; Welch, W.R.; Folkman, J. Tumor angiogenesis and metastasis—Correlation in invasive breast carcinoma. N. Engl. J. Med. 1991, 324, 1–8. [Google Scholar] [CrossRef]
- Sample Availability: Contact the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yang, Z.-S.; Xu, Y.-F.; Huang, F.-F.; Ding, G.-F. Associations of nm23H1, VEGF-C, and VEGF-3 Receptor in Human Prostate Cancer. Molecules 2014, 19, 6851-6862. https://doi.org/10.3390/molecules19056851
Yang Z-S, Xu Y-F, Huang F-F, Ding G-F. Associations of nm23H1, VEGF-C, and VEGF-3 Receptor in Human Prostate Cancer. Molecules. 2014; 19(5):6851-6862. https://doi.org/10.3390/molecules19056851
Chicago/Turabian StyleYang, Zui-Su, Yin-Feng Xu, Fang-Fang Huang, and Guo-Fang Ding. 2014. "Associations of nm23H1, VEGF-C, and VEGF-3 Receptor in Human Prostate Cancer" Molecules 19, no. 5: 6851-6862. https://doi.org/10.3390/molecules19056851