Overexpression of Receptor Tyrosine Kinase EphB4 Triggers Tumor Growth and Hypoxia in A375 Melanoma Xenografts: Insights from Multitracer Small Animal Imaging Experiments †

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. EphB4 Expression in Transfected A375 Melanoma Cells and Corresponding Tumor Xenografts
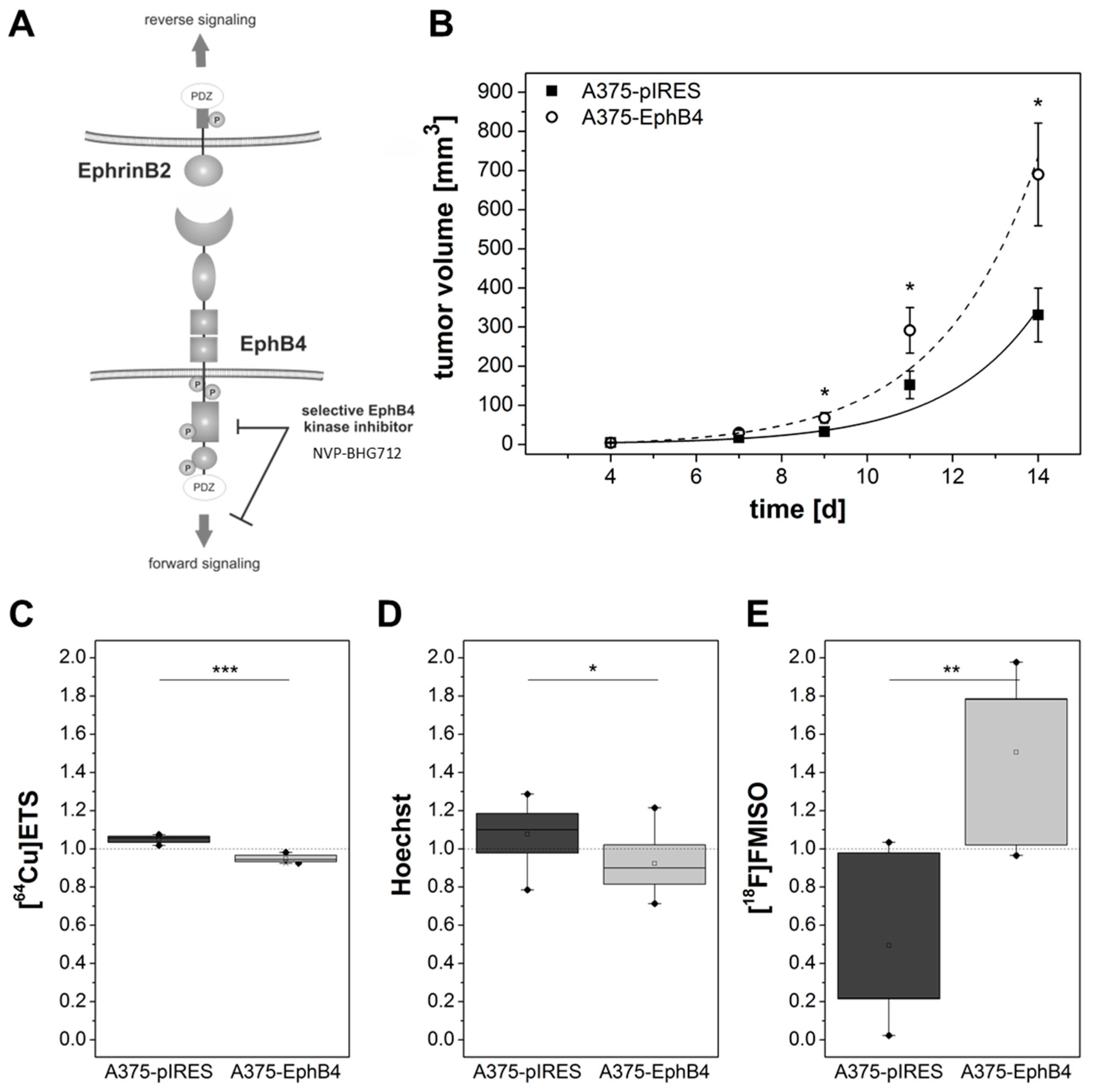
2.2. EphB4 Promotes Growth of A375 Melanoma Cells and Xenografts
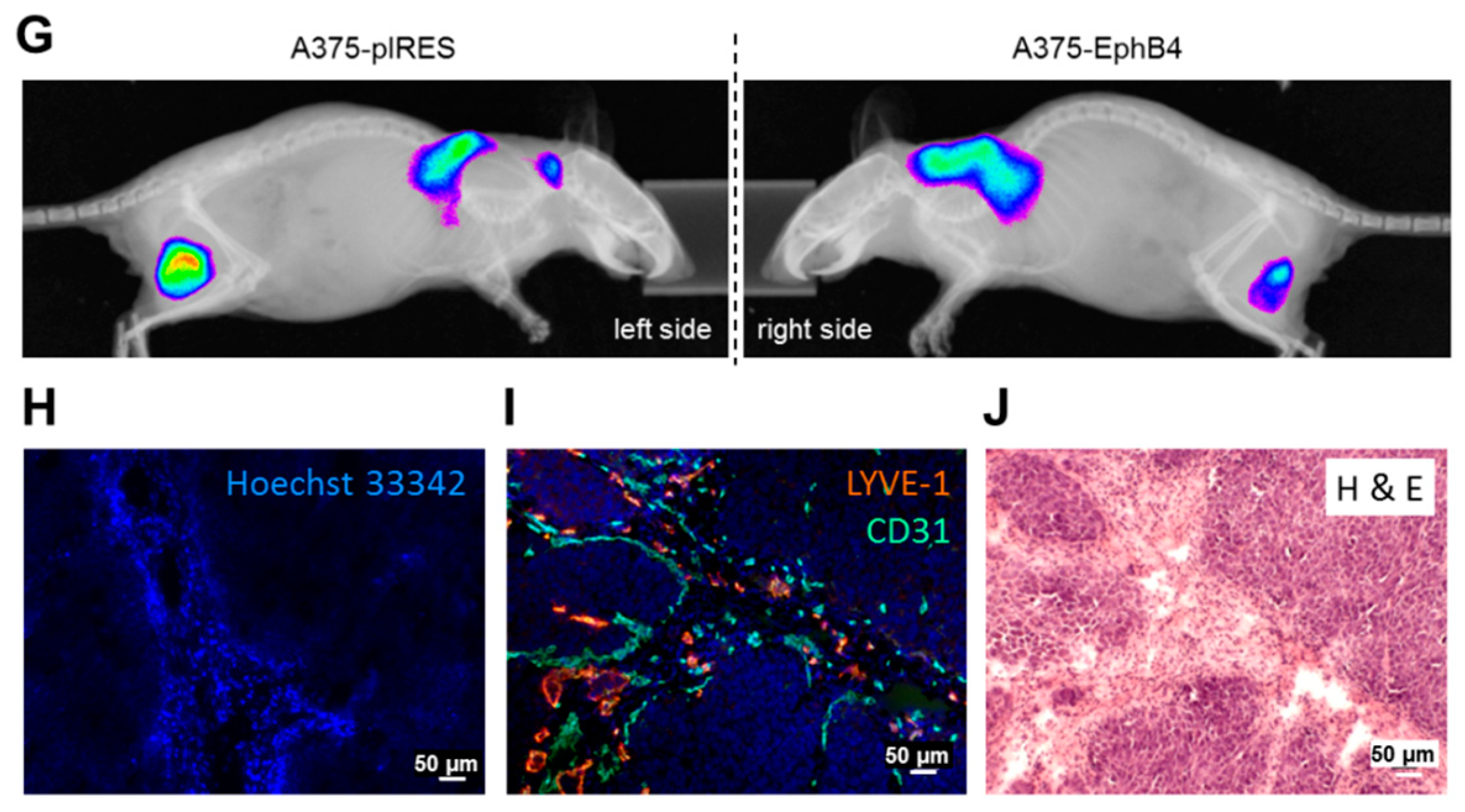
2.3. EphB4 Decreases Perfusion and Vascularization of A375 Melanoma Xenografts
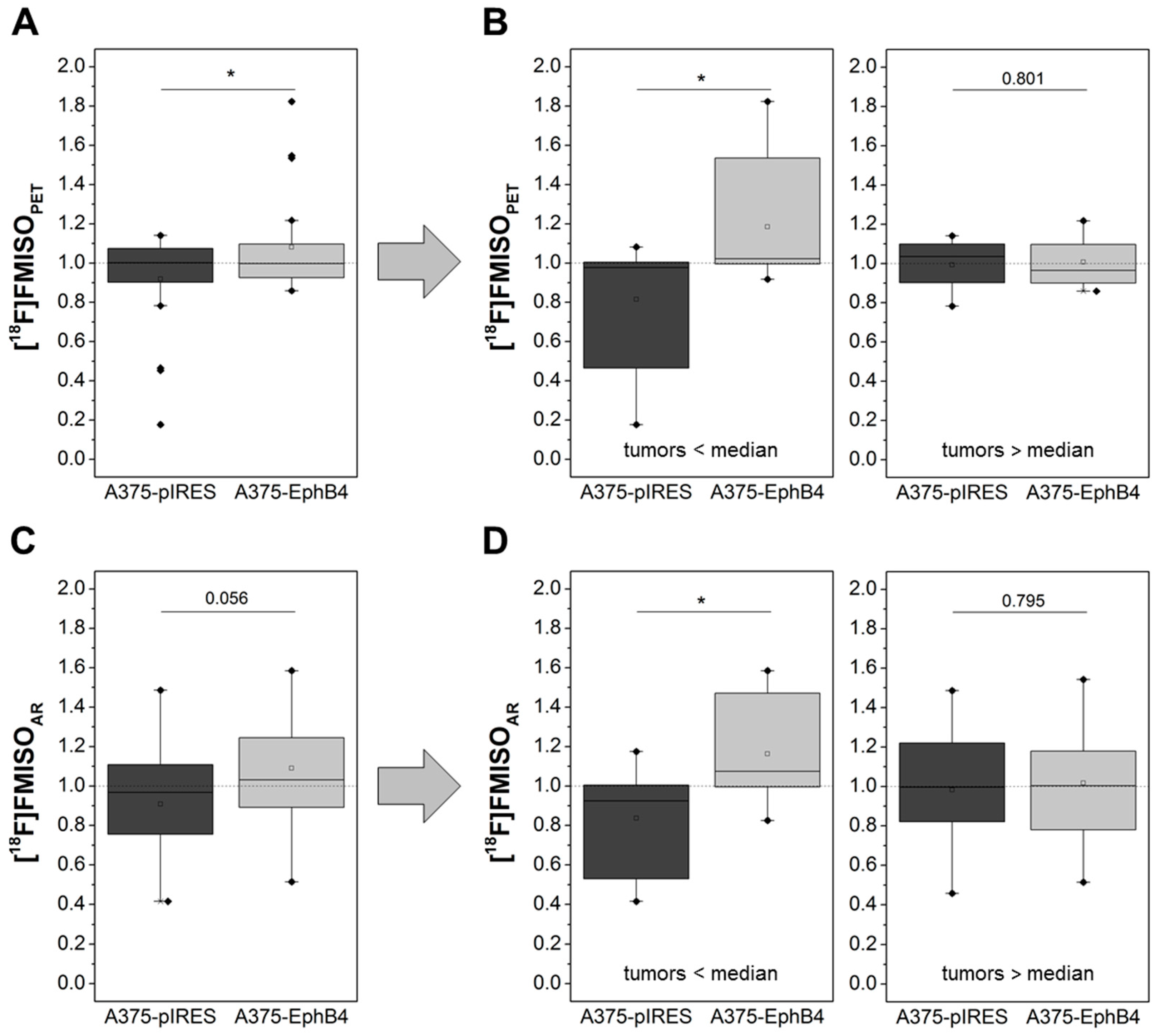
2.4. EphB4 Increases Hypoxia of A375 Melanoma Xenografts
2.5. EphB4 forward Signaling Is Not Responsible for Decreased Perfusion and Increased Hypoxia
3. Discussion
4. Materials and Methods
4.1. Generation of EphB4 Overexpressing A375 Melanoma Cells
4.2. Flow Cytometry
4.3. RNA Extraction and Quantitative Real-Time Reverse Transcription-PCR
4.4. Immunoblotting
4.5. Phospho-EphB4 ELISA
4.6. Cell Culture and Proliferation Assay
4.7. Animals and Generation of A375 Melanoma Xenografts
4.8. Blocking Experiments with EphB4 Kinase Inhibitor NVP-BHG712
4.9. Magnet Resonance Imaging
4.10. Positron Emission Tomography
4.11. In Vivo AngioSense and Ex Vivo Hoechst 33342 Fluorescence Imaging
4.12. Tumor Radioluminography
4.13. Immunohistochemistry and Fluorescence Microscopy
4.14. Quantitative Analysis of Radioluminography and Fluorescence Microscopy
4.15. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Boyle, G.M. Therapy for metastatic melanoma: an overview and update. Expert Rev. Anticancer Ther. 2011, 11, 725–737. [Google Scholar] [CrossRef] [PubMed]
- Tandler, N.; Mosch, B.; Pietzsch, J. Protein and non-protein biomarkers in melanoma: a critical update. Amino Acids 2012, 43, 2203–2230. [Google Scholar] [CrossRef] [PubMed]
- Streit, M.; Detmar, M. Angiogenesis, lymphangiogenesis, and melanoma metastasis. Oncogene 2003, 22, 3172–3179. [Google Scholar] [CrossRef] [PubMed]
- Goydos, J.S.; Gorski, D.H. Vascular endothelial growth factor C mRNA expression correlates with stage of progression in patients with melanoma. Clin. Cancer Res. 2003, 9, 5962–5967. [Google Scholar] [PubMed]
- Folkman, J. Tumor angiogenesis: therapeutic implications. N. Engl. J. Med. 1971, 285, 1182–1186. [Google Scholar] [PubMed]
- Dadras, S.S.; Paul, T.; Bertoncini, J.; Brown, L.F.; Muzikansky, A.; Jackson, D.G.; Ellwanger, U.; Garbe, C.; Mihm, M.C.; Detmar, M. Tumor lymphangiogenesis: a novel prognostic indicator for cutaneous melanoma metastasis and survival. Am. J. Pathol. 2003, 162, 1951–1960. [Google Scholar] [CrossRef]
- Dadras, S.S.; Lange-Asschenfeldt, B.; Velasco, P.; Nguyen, L.; Vora, A.; Muzikansky, A.; Jahnke, K.; Hauschild, A.; Hirakawa, S.; Mihm, M.C.; et al. Tumor lymphangiogenesis predicts melanoma metastasis to sentinel lymph nodes. Mod. Pathol. 2005, 18, 1232–1242. [Google Scholar] [CrossRef] [PubMed]
- Rinderknecht, M.; Detmar, M. Tumor lymphangiogenesis and melanoma metastasis. J. Cell. Physiol. 2008, 216, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Mumprecht, V.; Detmar, M. Lymphangiogenesis and cancer metastasis. J. Cell. Mol. Med. 2009, 13, 1405–1416. [Google Scholar] [CrossRef] [PubMed]
- Ugurel, S.; Rappl, G.; Tilgen, W.; Reinhold, U. Increased serum concentration of angiogenic factors in malignant melanoma patients correlates with tumor progression and survival. J. Clin. Oncol. 2001, 19, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Osella-Abate, S.; Quaglino, P.; Savoia, P.; Leporati, C.; Comessatti, A.; Bernengo, M.G. VEGF-165 serum levels and tyrosinase expression in melanoma patients: correlation with the clinical course. Melanoma Res. 2002, 12, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Mouawad, R.; Spano, J.P.; Comperat, E.; Capron, F.; Khayat, D. Tumoural expression and circulating level of VEGFR-3 (Flt-4) in metastatic melanoma patients: correlation with clinical parameters and outcome. Eur. J. Cancer 2009, 45, 1407–1414. [Google Scholar] [CrossRef] [PubMed]
- Mehnert, J.M.; McCarthy, M.M.; Jitaveanu, L.; Flaherty, K.T.; Aziz, S.; Camp, R.L.; Rimm, D.L.; Kluger, H.M. Quantitative expression of VEGF, VEGF-R1, VEGF-R2, and VEGF-R3 in melanoma tissue microarrays. Hum. Pathol. 2010, 41, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Zaki, K.A.; Basu, B.; Corrie, P. The role of angiogenesis inhibitors in the management of melanoma. Curr. Top Med. Chem. 2012, 12, 32–49. [Google Scholar] [CrossRef] [PubMed]
- Grothey, A.; Ellis, L.M. Targeting angiogenesis driven by vascular endothelial growth factors using antibody-based therapies. Cancer J. 2008, 14, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N.; Hillan, K.J.; Gerber, H.P.; Novotny, W. Discovery and development of bevacizumab, an anti-VEGF antibody for treating cancer. Nat. Rev. Drug Discov. 2004, 3, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Strumberg, D. Preclinical and clinical development of the oral multikinase inhibitor sorafenib in cancer treatment. Drugs Today 2005, 41, 773–784. [Google Scholar] [CrossRef] [PubMed]
- Mendel, D.B.; Laird, A.D.; Xin, X.H.; Louie, S.G.; Christensen, J.G.; Li, G.; Schreck, R.E.; Abrams, T.J.; Ngai, T.J.; Lee, L.B.; et al. In vivo antitumor activity of SU11248, a novel tyrosine kinase inhibitor targeting vascular endothelial growth factor and platelet-derived growth factor receptors: determination of a pharmacokinetic/pharmacodynamic relationship. Clin. Cancer Res. 2003, 9, 327–337. [Google Scholar] [PubMed]
- Harris, P.A.; Boloor, A.; Cheung, M.; Kumar, R.; Crosby, R.M.; Davis-Ward, R.G.; Epperly, A.H.; Hinkle, K.W.; Hunter, R.N., 3rd; Johnson, J.H.; et al. Discovery of 5-[[4-[(2,3-dimethyl-2H-indazol-6-yl)methylamino]-2-pyrimidinyl]amino]-2-methylbenzenesulfonamide (pazopanib), a novel and potent vascular endothelial growth factor receptor inhibitor. J. Med. Chem. 2008, 51, 4632–4640. [Google Scholar] [CrossRef] [PubMed]
- Nagy, J.A.; Chang, S.H.; Dvorak, A.M.; Dvorak, H.F. Why are tumour blood vessels abnormal and why is it important to know? Brit. J. Cancer 2009, 100, 865–869. [Google Scholar] [CrossRef] [PubMed]
- Nagy, J.A.; Chang, S.H.; Shih, S.C.; Dvorak, A.M.; Dvorak, H.F. Heterogeneity of the tumor vasculature. Semin. Thromb. Hemost. 2010, 36, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Leu, A.J.; Berk, D.A.; Lymboussaki, A.; Alitalo, K.; Jain, R.K. Absence of functional lymphatics within a murine sarcoma: A molecular and functional evaluation. Cancer Res. 2000, 60, 4324–4327. [Google Scholar] [PubMed]
- Padera, T.P.; Kadambi, A.; di Tomaso, E.; Carreira, C.M.; Brown, E.B.; Boucher, Y.; Choi, N.C.; Mathisen, D.; Wain, J.; Mark, E.J.; et al. Lymphatic metastasis in the absence of functional intratumor lymphatics. Science 2002, 296, 1883–1886. [Google Scholar] [CrossRef] [PubMed]
- Minchinton, A.I.; Tannock, I.F. Drug penetration in solid tumours. Nat. Rev. Cancer 2006, 6, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.K. Normalizing tumor vasculature with anti-angiogenic therapy: A new paradigm for combination therapy. Nat. Med. 2001, 7, 987–989. [Google Scholar] [CrossRef] [PubMed]
- Oku, T.; Tjuvajev, J.G.; Miyagawa, T.; Sasajima, T.; Joshi, A.; Joshi, R.; Finn, R.; Claffey, K.P.; Blasberg, R.G. Tumor growth modulation by sense and antisense vascular endothelial growth factor gene expression: Effects on angiogenesis, vascular permeability, blood volume, blood flow, fluorodeoxyglucose uptake, and proliferation of human melanoma intracerebral xenografts. Cancer Res. 1998, 58, 4185–4192. [Google Scholar] [PubMed]
- Li, Y.W.; Wang, M.N.; Li, H.L.; King, K.D.; Bassi, R.; Sun, H.; Santiago, A.; Hooper, A.T.; Bohlen, P.; Hicklin, D.J. Active immunization against the vascular endothelial growth factor receptor flk1 inhibits tumor angiogenesis and metastasis. J. Exp. Med. 2002, 195, 1575–1584. [Google Scholar] [CrossRef] [PubMed]
- Herbst, R.S.; Ansari, R.; Bustin, F.; Flynn, P.; Hart, L.; Otterson, G.A.; Vlahovic, G.; Soh, C.H.; O'Connor, P.; Hainsworth, J. Efficacy of bevacizumab plus erlotinib versus erlotinib alone in advanced non-small-cell lung cancer after failure of standard first-line chemotherapy (BeTa): A double-blind, placebo-controlled, phase 3 trial. Lancet 2011, 377, 1846–1854. [Google Scholar] [CrossRef]
- Perren, T.J.; Swart, A.M.; Pfisterer, J.; Ledermann, J.A.; Pujade-Lauraine, E.; Kristensen, G.; Carey, M.S.; Beale, P.; Cervantes, A.; Kurzeder, C.; et al. A Phase 3 Trial of Bevacizumab in Ovarian Cancer. N. Engl. J. Med. 2011, 365, 2484–2496. [Google Scholar] [CrossRef] [PubMed]
- Oza, A.M.; Cook, A.D.; Pfisterer, J.; Embleton, A.; Ledermann, J.A.; Pujade-Lauraine, E.; Kristensen, G.; Carey, M.S.; Beale, P.; Cervantes, A.; et al. Standard chemotherapy with or without bevacizumab for women with newly diagnosed ovarian cancer (ICON7): Overall survival results of a phase 3 randomised trial. Lancet Oncol. 2015, 16, 928–936. [Google Scholar] [CrossRef]
- Mosch, B.; Reissenweber, B.; Neuber, C.; Pietzsch, J. Eph receptors and ephrin ligands: important players in angiogenesis and tumor angiogenesis. J. Oncol. 2010, 2010, 135285. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.U.; Chen, Z.F.; Anderson, D.J. Molecular distinction and angiogenic interaction between embryonic arteries and veins revealed by ephrin-B2 and its receptor Eph-B4. Cell 1998, 93, 741–753. [Google Scholar] [CrossRef]
- Adams, R.H.; Wilkinson, G.A.; Weiss, C.; Diella, F.; Gale, N.W.; Deutsch, U.; Risau, W.; Klein, R. Roles of ephrinB ligands and EphB receptors in cardiovascular development: demarcation of arterial/venous domains, vascular morphogenesis, and sprouting angiogenesis. Gene Dev. 1999, 13, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Gerety, S.S.; Wang, H.U.; Chen, Z.F.; Anderson, D.J. Symmetrical mutant phenotypes of the receptor EphB4 and its specific transmembrane ligand ephrin-B2 in cardiovascular development. Mol. Cell 1999, 4, 403–414. [Google Scholar] [CrossRef]
- Gerety, S.S.; Anderson, D.J. Cardiovascular ephrinB2 function is essential for embryonic angiogenesis. Development 2002, 129, 1397–1410. [Google Scholar] [PubMed]
- Makinen, T.; Adams, R.H.; Bailey, J.; Lu, Q.; Ziemiecki, A.; Alitalo, K.; Klein, R.; Wilkinson, G.A. PDZ interaction site in ephrinB2 is required for the remodeling of lymphatic vasculature. Gene Dev. 2005, 19, 397–410. [Google Scholar] [CrossRef] [PubMed]
- Foo, S.S.; Turner, C.J.; Adams, S.; Compagni, A.; Aubyn, D.; Kogata, N.; Lindblom, P.; Shani, M.; Zicha, D.; Adams, R.H. Ephrin-B2 controls cell motility and adhesion during blood-vessel-wall assembly. Cell 2006, 124, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Adams, R.H.; Diella, F.; Hennig, S.; Helmbacher, F.; Deutsch, U.; Klein, R. The cytoplasmic domain of the ligand ephrinB2 is required for vascular morphogenesis but not cranial neural crest migration. Cell 2001, 104, 57–69. [Google Scholar] [CrossRef]
- Ji, R.C. Lymphatic endothelial cells, lymphedematous lymphangiogenesis, and molecular control of edema formation. Lymphat. Res. Biol. 2008, 6, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Nakayama, M.; Pitulescu, M.E.; Schmidt, T.S.; Bochenek, M.L.; Sakakibara, A.; Adams, S.; Davy, A.; Deutsch, U.; Lüthi, U.; et al. Ephrin-B2 controls VEGF-induced angiogenesis and lymphangiogenesis. Nature 2010, 465, 483–486. [Google Scholar] [CrossRef] [PubMed]
- Sawamiphak, S.; Seidel, S.; Essmann, C.L.; Wilkinson, G.A.; Pitulescu, M.E.; Acker, T.; Acker-Palmer, A. Ephrin-B2 regulates VEGFR2 function in developmental and tumour angiogenesis. Nature 2010, 465, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Noren, N.K.; Lu, M.; Freeman, A.L.; Koolpe, M.; Pasquale, E.B. Interplay between EphB4 on tumor cells and vascular ephrin-B2 regulates tumor growth. Proc. Natl. Acad. Sci. USA 2004, 101, 5583–5588. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.H.; Xia, Q.Y.; Wang, J.J.; Shen, Q.; Zhang, J.; Zhou, X. EphB4 promotes the proliferation, invasion, and angiogenesis of human colorectal cancer. Exp. Mol. Pathol. 2016, 100, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Guijarro-Munoz, I.; Sanchez, A.; Martinez-Martinez, E.; García, J.M.; Salas, C.; Provencio, M.; Alvarez-Vallina, L.; Sanz, L. Gene expression profiling identifies EPHB4 as a potential predictive biomarker in colorectal cancer patients treated with bevacizumab. Med. Oncol. 2013, 30, 572. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.R.; Singh, J.; Xia, G.B.; Krasnoperov, V.; Hassanieh, L.; Ley, E.J.; Scehnet, J.; Kumar, N.G.; Hawes, D.; Press, M.F.; et al. Receptor tyrosine kinase EphB4 is a survival factor in breast cancer. Am. J. Pathol. 2006, 169, 279–293. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, S.A.; Slomka, S.; Douglas, E.L.; Hewett, P.J.; Hardingham, J.E. Receptor protein tyrosine kinase EphB4 is up-regulated in colon cancer. BMC Mol. Biol. 2001, 2, 15. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Ahmad, S.A.; Jung, Y.D.; Reinmuth, N.; Fan, F.; Bucana, C.D.; Ellis, L.M. Coexpression of ephrin-Bs and their receptors in colon carcinoma. Cancer. 2002, 94, 934–939. [Google Scholar] [CrossRef] [PubMed]
- Berclaz, G.; Karamitopoulou, E.; Mazzucchelli, L.; Rohrbach, V.; Dreher, E.; Ziemiecki, A.; Andres, A.C. Activation of the receptor protein tyrosine kinase EphB4 in endometrial hyperplasia and endometrial carcinoma. Ann. Oncol. 2003, 14, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.C.; Perren, J.R.; Douglas, E.L.; Raynor, M.P.; Bartley, M.A.; Bardy, P.G.; Stephenson, S.A. Investigation of the expression of the EphB4 receptor tyrosine kinase in prostate carcinoma. BMC Cancer 2005, 5, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellano, G.; Reid, J.F.; Alberti, P.; Carcangiu, M.L.; Tomassetti, A.; Canevari, S. New potential ligand-receptor signaling loops in ovarian cancer identified in multiple gene expression studies. Cancer Res. 2006, 66, 10709–10719. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.K.; Yang, Y.D.; Tang, S.Q.; Tang, H.; Yang, G.; Xu, Q.; Wu, J. EphB4 inhibitor overcome the acquired resistance to cisplatin in melanomas xenograft model. J. Pharmacol. Sci. 2015, 129, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Muz, B.; de la Puente, P.; Azab, F.; Azab, A.K. The role of hypoxia in cancer progression, angiogenesis, metastasis, and resistance to therapy. Hypoxia 2015, 3, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Reissenweber, B.; Mosch, B.; Pietzsch, J. Experimental hypoxia does not influence gene expression and protein synthesis of Eph receptors and ephrin ligands in human melanoma cells in vitro. Melanoma Res. 2013, 23, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Mullani, N.A.; Herbst, R.S.; O'Neil, R.G.; Gould, K.L.; Barron, B.J.; Abbruzzese, J.L. Tumor blood flow measured by PET dynamic imaging of first-pass 18F-FDG uptake: A comparison with 15O-labeled water measured blood flow. J. Nucl. Med. 2008, 49, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, J.W.; Logan, T.F.; Eitel, J.A.; Mathias, C.J.; Ng, Y.; Lacy, J.L.; Hutchins, G.D.; Green, M.A. Whole-body PET/CT evaluation of tumor perfusion using generator-based 62Cu-ethylglyoxal bis(thiosemicarbazonato)copper(II): Validation by direct comparison to 15O-water in metastatic renal cell carcinoma. J. Nucl. Med. 2015, 56, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.J.; Ferdani, R. Copper-64 radiopharmaceuticals for PET imaging of cancer: Advances in preclinical and clinical research. Cancer Biother. Radiopharm. 2009, 24, 379–393. [Google Scholar] [CrossRef] [PubMed]
- Martiny-Baron, G.; Holzer, P.; Billy, E.; Schnell, C.; Brueggen, J.; Ferretti, M.; Schmiedeberg, N.; Wood, J.M.; Furet, P.; Imbach, P. The small molecule specific EphB4 kinase inhibitor NVP-BHG712 inhibits VEGF driven angiogenesis. Angiogenesis 2010, 13, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Mamat, C.; Mosch, B.; Neuber, C.; Köckerling, M.; Bergmann, R.; Pietzsch, J. Fluorine-18 radiolabeling and radiopharmacological characterization of a benzodioxolylpyrimidine-based radiotracer targeting the receptor tyrosine kinase EphB4. ChemMedChem 2012, 7, 1991–2003. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.Y.; Pasquale, E.B.; Owen, L.B.; Ethell, I.M. The EphB4 receptor-tyrosine kinase promotes the migration of melanoma cells through Rho-mediated actin cytoskeleton reorganization. J. Biol. Chem. 2006, 281, 32574–32586. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.Y.; Lopez-Bergami, P.; Goydos, J.S.; Yip, D.; Walker, A.M.; Pasquale, E.B.; Ethell, I.M. The EphB4 receptor promotes the growth of melanoma cells expressing the ephrin-B2 ligand. Pigment Cell Melanoma Res. 2010, 23, 684–687. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Yamada, Y.; Kidoya, H.; Naito, H.; Nagahama, Y.; Kong, L.; Katoh, S.Y.; Li, W.L.; Ueno, M.; Takakura, N. EphB4 overexpression in B16 melanoma cells affects arterial-venous patterning in tumor angiogenesis. Cancer Res. 2007, 67, 9800–9808. [Google Scholar] [CrossRef] [PubMed]
- Pasquale, E.B.; Noren, N.K. Paradoxes of the EphB4 receptor in cancer. Cancer Res. 2007, 67, 3994–3997. [Google Scholar]
- Rutkowski, R.; Mertens-Walker, I.; Lisle, J.E.; Herington, A.C.; Stephenson, S.A. Evidence for a dual function of EphB4 as tumor promoter and suppressor regulated by the absence or presence of the ephrin-B2 ligand. Int. J. Cancer. 2012, 131, E614–E624. [Google Scholar] [CrossRef] [PubMed]
- Martiny-Baron, G.; Korff, T.; Schaffner, F.; Esser, N.; Eggstein, S.; Marmé, D.; Augustin, H.G. Inhibition of tumor growth and angiogenesis by soluble EphB4. Neoplasia 2004, 6, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Heroult, M.; Schaffner, F.; Pfaff, D.; Prahst, C.; Kirmse, R.; Kutschera, S.; Riedel, M.; Ludwig, T.; Vajkoczy, P.; Graeser, R.; et al. EphB4 promotes site-specific metastatic tumor cell dissemination by interacting with endothelial cell-expressed ephrinB2. Mol. Cancer Res. 2010, 8, 1297–1309. [Google Scholar] [CrossRef] [PubMed]
- Vogt, T.; Stolz, W.; Welsh, J.; Jung, B.; Kerbel, R.S.; Kobayashi, H.; Landthaler, M.; McClelland, M. Overexpression of Lerk-5/Eplg5 messenger RNA: A novel marker for increased tumorigenicity and metastatic potential in human malignant melanomas. Clin. Cancer Res. 1998, 4, 791–797. [Google Scholar] [PubMed]
- Liu, W.; Jung, Y.D.; Ahmad, S.A.; McCarty, M.F.; Stoeltzing, O.; Reinmuth, N.; Fan, F.; Ellis, L.M. Effects of overexpression of ephrin-B2 on tumour growth in human colorectal cancer. Brit. J. Cancer 2004, 90, 1620–1626. [Google Scholar] [CrossRef] [PubMed]
- Green, M.A.; Mathias, C.J.; Willis, L.R.; Handa, R.K.; Lacy, J.L.; Miller, M.A.; Hutchins, G.D. Assessment of Cu-ETS as a PET radiopharmaceutical for evaluation of regional renal perfusion. Nucl. Med. Biol. 2007, 34, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Basken, N.E.; Mathias, C.J.; Lipka, A.E.; Green, M.A. Species dependence of [64Cu]Cu-Bis(thiosemicarbazone) radiopharmaceutical binding to serum albumins. Nucl. Med. Biol. 2008, 35, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Montet, X.; Figueiredo, J.L.; Alencar, H.; Ntziachristos, V.; Mahmood, U.; Weissleder, R. Tomographic fluorescence imaging of tumor vascular volume in mice. Radiology 2007, 242, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.B.; Bindokas, V.; Shen, J.K.; Fan, H.; Hoffman, R.M.; Xing, H.R. Time-course imaging of therapeutic functional tumor vascular normalization by antiangiogenic agents. Mol. Cancer Ther. 2011, 10, 1173–1184. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.A.; Hill, S.A.; Begg, A.C.; Denekamp, J. Validation of the fluorescent dye Hoechst 33342 as a vascular space marker in tumors. Brit. J. Cancer 1988, 57, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Hu, H.Q.; Guevara-Gallardo, S.; Lam, M.T.; Fong, S.Y.; Wang, R.A. Artery and vein size is balanced by Notch and ephrin B2/EphB4 during angiogenesis. Development 2008, 135, 3755–3764. [Google Scholar] [CrossRef] [PubMed]
- Koh, W.J.; Rasey, J.S.; Evans, M.L.; Grierson, J.R.; Lewellen, T.K.; Graham, M.M.; Krohn, K.A.; Griffin, T.W. Imaging of hypoxia in human tumors with [F-18]fluoromisonidazole. Int. J. Radiat. Oncol. Biol. Phys. 1992, 22, 199–212. [Google Scholar] [CrossRef]
- Jentsch, C.; Beuthien-Baumann, B.; Troost, E.G.C.; Shakirin, G. Validation of functional imaging as a biomarker for radiation treatment response. Brit. J. Radiol. 2015, 88, 20150014. [Google Scholar] [CrossRef] [PubMed]
- Bechmann, N.; Hauser, S.; Hofheinz, F.; Kniess, T.; Pietzsch, J. Nitric oxide-releasing selective cyclooxygenase-2 inhibitors as promising radiosensitizers in melanoma cells in vitro. Ann. Radiat. Ther. Oncol. 2017, 1, 1010. [Google Scholar]
- Rockwell, S.; Dobrucki, I.T.; Kim, E.Y.; Marrison, S.T.; Vu, V.T. Hypoxia and radiation therapy: Past history, ongoing research, and future promise. Curr. Mol. Med. 2009, 9, 442–458. [Google Scholar] [CrossRef] [PubMed]
- Colliez, F.; Gallez, B.; Jordan, B.F. Assessing tumor oxygenation for predicting outcome in radiation oncology: A review of studies correlating tumor hypoxic status and outcome in the preclinical and clinical settings. Front. Oncol. 2017, 7, 10. [Google Scholar] [CrossRef] [PubMed]
- Banerji, S.; Ni, J.; Wang, S.X.; Clasper, S.; Su, J.; Tammi, R.; Jones, M.; Jackson, D.G. LYVE-1, a new homologue of the CD44 glycoprotein, is a lymph-specific receptor for hyaluronan. J. Cell Biol. 1999, 144, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Jackson, D.G.; Prevo, R.; Clasper, S.; Banerji, S. LYVE-1, the lymphatic system and tumor lymphangiogenesis. Trends Immunol. 2001, 22, 317–321. [Google Scholar] [CrossRef]
- Germain, S.; Eichmann, A. VEGF and ephrin-B2: a bloody duo. Nat. Med. 2010, 16, 752–754. [Google Scholar] [CrossRef] [PubMed]
- Krasnoperov, V.; Kumar, S.R.; Ley, E.; Li, X.; Scehnet, J.; Liu, R.; Zozulya, S.; Gill, P.S. Novel EphB4 monoclonal antibodies modulate angiogenesis and inhibit tumor growth. Am. J. Pathol. 2010, 176, 2029–2038. [Google Scholar] [CrossRef] [PubMed]
- Abengozar, M.A.; de Frutos, S.; Ferreiro, S.; Soriano, J.; Perez-Martinez, M.; Olmeda, D.; Marenchino, M.; Cañamero, M.; Ortega, S.; Megias, D.; et al. Blocking ephrinB2 with highly specific antibodies inhibits angiogenesis, lymphangiogenesis, and tumor growth. Blood 2012, 119, 4565–4576. [Google Scholar] [CrossRef] [PubMed]
- Mosch, B.; Pietzsch, D.; Pietzsch, J. Irradiation affects cellular properties and Eph receptor expression in human melanoma cells. Cell Adhes. Migr. 2012, 6, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Tondera, C.; Hauser, S.; Kruger-Genge, A.; Jung, F.; Neffe, A.T.; Lendlein, A.; Klopfleisch, R.; Steinbach, J.; Neuber, C.; Pietzsch, J. Gelatin-based Hydrogel Degradation and Tissue Interaction in vivo: Insights from Multimodal Preclinical Imaging in Immunocompetent Nude Mice. Theranostics 2016, 6, 2114–2128. [Google Scholar] [CrossRef] [PubMed]
- Tondera, C.; Wieduwild, R.; Roeder, E.; Werner, C.; Zhang, Y.; Pietzsch, J. In vivo examination of an injectable hydrogel system crosslinked by peptide-oligosaccharide interaction in immunocompetent nude mice. Adv. Funct. Mater. 2017, 27, 1605189. [Google Scholar] [CrossRef]
- Kniess, T.; Laube, M.; Bergmann, R.; Sehn, F.; Graf, F.; Steinbach, J.; Wuest, F.; Pietzsch, J. Radiosynthesis of a ¹⁸F-labeled 2,3-diarylsubstituted indole via McMurry coupling for functional characterization of cyclooxygenase-2 (COX-2) in vitro and in vivo. Bioorgan. Med. Chem. 2012, 20, 3410–3421. [Google Scholar] [CrossRef] [PubMed]
- Mullani, N.; Herbst, R.; Abbruzzese, J.; Barron, B.; Lamki, L.; Charnsangavej, C.; Kim, E.; Tran, H.; Jiwani, A.; Gould, K. First pass FDG measured blood flow in tumors: A comparison with O-15 labeled water measured blood flow. Clin. Positron. Imaging. 2000, 3, 153. [Google Scholar] [CrossRef]
- Patlak, C.S.; Blasberg, R.G. Graphical evaluation of blood-to-brain transfer constants from multiple-time uptake data. Gen. J. Cereb. Blood Flow Metab. 1985, 5, 584–590. [Google Scholar] [CrossRef] [PubMed]
- Casciari, J.J.; Graham, M.M.; Rasey, J.S. A modeling approach for quantifying tumor hypoxia with [F-18]fluoromisonidazole PET time-activity data. Med. Phys. 1995, 22, 1127–1139. [Google Scholar] [CrossRef] [PubMed]
- Kelada, O.J.; Rockwell, S.; Zheng, M.Q.; Huang, Y.; Liu, Y.; Booth, C.J.; Decker, R.H.; Oelfke, U.; Carson, R.E.; Carlson, D.J. Quantification of tumor hypoxic fractions using positron emission tomography with [18F]fluoromisonidazole ([18F]FMISO) kinetic analysis and invasive oxygen measurements. Mol. Imaging Biol. 2017, 19, 893–902. [Google Scholar] [CrossRef] [PubMed]
- RCoreTeam. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Ebert, K.; Wiemer, J.; Caballero, J.; Köckerling, M.; Steinbach, J.; Pietzsch, J.; Mamat, C. Development of indazolylpyrimidine derivatives as high-affine EphB4 receptor ligands and potential PET radiotracers. Biorg. Med. Chem. 2015, 23, 6025–6035. [Google Scholar] [CrossRef] [PubMed]
- Wiemer, J.; Steinbach, J.; Pietzsch, J.; Mamat, C. Preparation of a novel radiotracer targeting the EphB4 receptor via radiofluorination using spiro azetidinium salts as precursor. J. Label. Compd. Radiopharm. 2017, 60, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Belter, B.; Haase-Kohn, C.; Pietzsch, J. Biomarkers in malignant melanoma: recent trends and critical perspective. In Cutaneous Melanoma: Etiology and Therapy; Ward, W.H., Farma, J.M., Eds.; Codon Publications: Brisbane, Australia, 2017; pp. 39–56. [Google Scholar]
Sample Availability: Not available. |










© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neuber, C.; Belter, B.; Meister, S.; Hofheinz, F.; Bergmann, R.; Pietzsch, H.-J.; Pietzsch, J. Overexpression of Receptor Tyrosine Kinase EphB4 Triggers Tumor Growth and Hypoxia in A375 Melanoma Xenografts: Insights from Multitracer Small Animal Imaging Experiments. Molecules 2018, 23, 444. https://doi.org/10.3390/molecules23020444
Neuber C, Belter B, Meister S, Hofheinz F, Bergmann R, Pietzsch H-J, Pietzsch J. Overexpression of Receptor Tyrosine Kinase EphB4 Triggers Tumor Growth and Hypoxia in A375 Melanoma Xenografts: Insights from Multitracer Small Animal Imaging Experiments. Molecules. 2018; 23(2):444. https://doi.org/10.3390/molecules23020444
Chicago/Turabian StyleNeuber, Christin, Birgit Belter, Sebastian Meister, Frank Hofheinz, Ralf Bergmann, Hans-Jürgen Pietzsch, and Jens Pietzsch. 2018. "Overexpression of Receptor Tyrosine Kinase EphB4 Triggers Tumor Growth and Hypoxia in A375 Melanoma Xenografts: Insights from Multitracer Small Animal Imaging Experiments" Molecules 23, no. 2: 444. https://doi.org/10.3390/molecules23020444