Zinc and Skin Disorders


Department of Dermatology, Faculty of Medicine, University of Yamanashi, Yamanashi 409-3898, Japan
*
Author to whom correspondence should be addressed.
Nutrients 2018, 10(2), 199; https://doi.org/10.3390/nu10020199
Submission received: 7 December 2017
/
Revised: 7 February 2018
/
Accepted: 9 February 2018
/
Published: 11 February 2018
(This article belongs to the Special Issue Dietary Zn and Human Health)
{kind=link}
{kind=link}
{kind=link}
Abstract
:The skin is the third most zinc (Zn)-abundant tissue in the body. The skin consists of the epidermis, dermis, and subcutaneous tissue, and each fraction is composed of various types of cells. Firstly, we review the physiological functions of Zn and Zn transporters in these cells. Several human disorders accompanied with skin manifestations are caused by mutations or dysregulation in Zn transporters; acrodermatitis enteropathica (Zrt-, Irt-like protein (ZIP)4 in the intestinal epithelium and possibly epidermal basal keratinocytes), the spondylocheiro dysplastic form of Ehlers-Danlos syndrome (ZIP13 in the dermal fibroblasts), transient neonatal Zn deficiency (Zn transporter (ZnT)2 in the secretory vesicles of mammary glands), and epidermodysplasia verruciformis (ZnT1 in the epidermal keratinocytes). Additionally, acquired Zn deficiency is deeply involved in the development of some diseases related to nutritional deficiencies (acquired acrodermatitis enteropathica, necrolytic migratory erythema, pellagra, and biotin deficiency), alopecia, and delayed wound healing. Therefore, it is important to associate the existence of mutations or dysregulation in Zn transporters and Zn deficiency with skin manifestations.
1. Introduction
In the human body, zinc (Zn) is stably maintained in the weight of 2–3 g [1]. Skin is the third most Zn-abundant tissue in the body (skeletal muscle 60%, bones 30%, liver 5%, and skin 5%) [1]. The epidermis contains more Zn compared with the dermis [2]. In the epidermis, Zn is more abundantly distributed throughout the stratum spinosum than the other three layers of keratinocytes (KCs) [3]. In the dermis, Zn concentration in the upper dermis is higher than it in the lower dermis [2]. Zn is enriched in the granules of mast cells (MCs) [4] and MCs are more abundant in the upper dermis than in the lower dermis [5,6]. Therefore, the difference in dermal MC distributions may explain the difference of Zn distributions within the dermis.
Zn is present as a divalent ion (Zn2+) in cells and does not need a redox reaction upon crossing the cellular membrane. Thus, the tight regulation needed to maintain Zn homeostasis in cells is assumed by two solute-linked carrier (SLC) gene families: Zn transporter (ZnT; SLC30A) and Zrt-, Irt-like protein (ZIP; SLC39A) (reviewed in [7,8,9,10]). ZnTs and ZIPs are involved in Zn efflux and uptake, respectively. 10 ZnT and 14 ZIP were identified in humans so far [7]. Besides ZnTs and ZIPs, metallothioneins (MTs) also assume Zn regulation. MTs that ubiquitously distribute in the cytoplasm contain a unique cysteine-rich amino acid sequence. This allows MTs to bind to Zn, copper, and cadmium. An excess cytosolic Zn binds to MTs, whereas Zn is released from MTs in the condition of Zn deficiency or Zn required. Consequently, MTs function as a regulator of Zn homeostasis [11].
In this review, we review the relationship between Zn and the skin, including the function and distribution of Zn, ZnTs, ZIPs, and MTs in the skin with adding new recent findings in our recent review [12] by introducing the function of Zn and Zn transporters in various types of skin cells. Additionally, we expanded the description of Zn deficiency-related human disorders accompanied with skin manifestations. We started this review by describing the physiological functions of Zn and Zn transporters in various types of cells of skin (Section 2), followed by human skin disorders caused by mutations of Zn transporters (Section 3), human skin disorders caused by dysregulation of Zn transporters (Section 4), and human skin disorders associated with Zn deficiency (Section 5).
2. Physiological Functions of Zn and Zn Transporters in the Skin
Zn is a cofactor for over 1000 enzymatic reactions [13,14] and is necessary for over 2000 transcription factors [15]. Zn-finger proteins function for DNA interaction, RNA packaging, activation of transcription, regulation of apoptosis, folding and assembly of protein, and lipid binding [16,17,18]. Additionally, about 10% of human proteins binds to Zn [19]. Therefore, Zn is associated with a wide variety of organic activities such as development, differentiation, and cell growth.
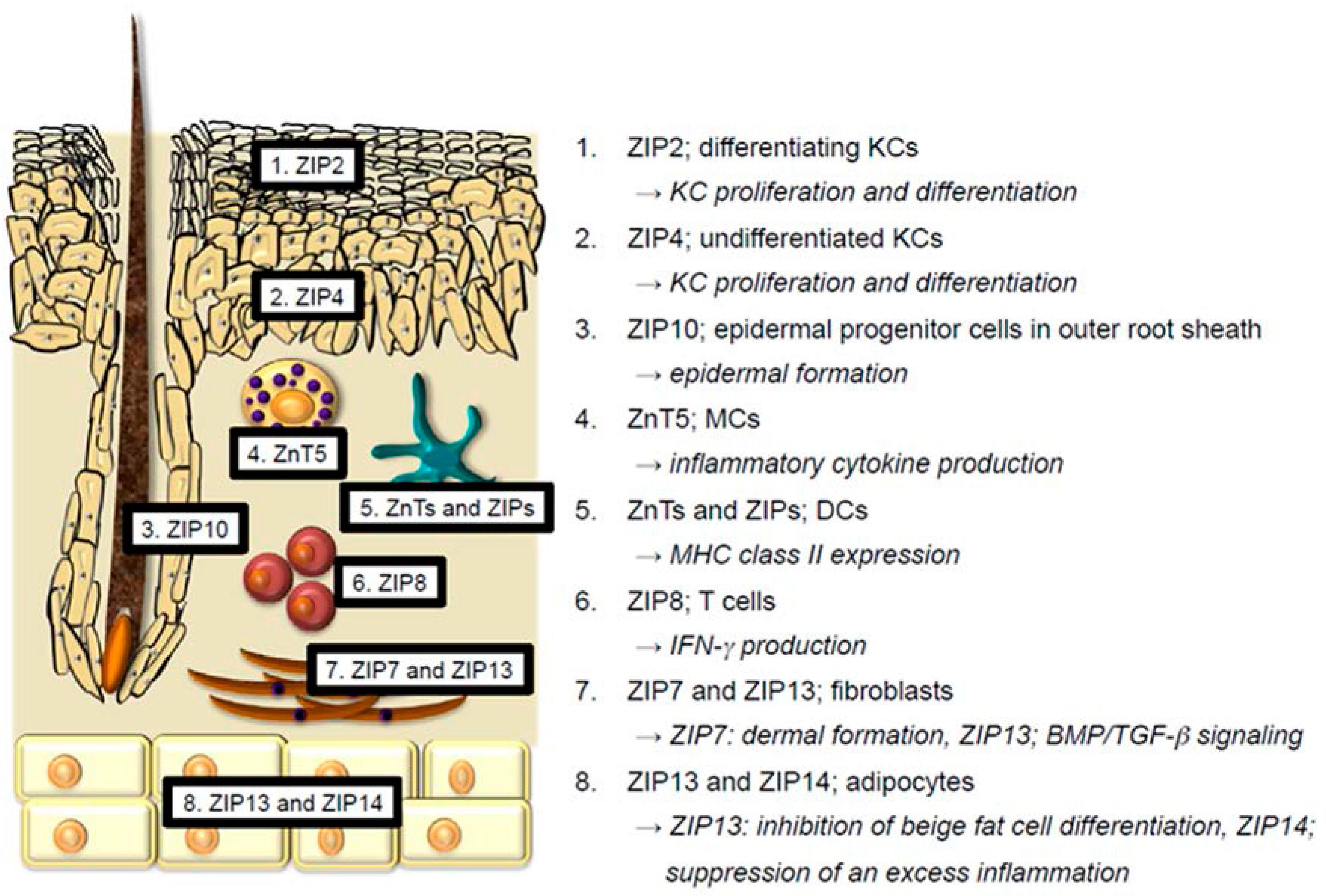
The skin consists of the epidermis, dermis, and subcutaneous tissue. Each skin region contains the following cells: epidermis; KCs, Langerhans cells (LCs), and melanocytes, dermis; antigen-presenting cells (dendritic cells (DCs), macrophages, and monocytes), T cells, MCs, fibroblasts, endothelial cells, etc., subcutaneous tissue; adipocytes. A review of the reported function of Zn and Zn transporters in these cells follows below (Figure 1).
2.1. Keratinocytes
Keratinocytes (KCs) occupy approximately 97% of epidermis and are categorized into four layers according to the degree of differentiation and keratinization. These layers include the basal layer, stratum spinosum, stratum granulosum, and stratum corneum. As described, Zn concentrations are most abundant in the stratum spinosum, more so than in the other three KC layers [3]. However, the physiological reason for this is not known. An in vitro experiment demonstrated that exposing HaCaT KCs (human immortalized KCs) to a nontoxic concentration of Zn facilitates survival and proliferation [20]. In contrast, a chelation of intracellular Zn by N,N,N′,N′-tetrakis (2-pyridylmethyl) ethylenediamine (TPEN) activates caspase-3 and DNA fragmentation, resulting in the apoptosis of KCs [21,22]. Further, a Zn-deficient diet alters the expression of keratin polypeptides in rats because of impaired keratinolytic enzyme activity [23]. In an in vitro experiment using normal human epidermal KCs, Zn suppressed IFN-γ-induced KC activation and tumor necrosis factor-α (TNF-α) production via unknown underlying mechanisms [24]. Zn also diminishes inducible nitric oxide synthase (iNOS) induction and subsequent nitric oxide (NO) production in Pam212 KCs (murine immortalized KCs) [25]. These data suggest that Zn is required for the proliferation of KCs and the suppression of inflammation in KCs (reviewed in [26]). These effects of Zn on KCs account for the clinical effects of Zn oxide ointment on skin inflammation and ulcers.
Recent research elucidated the functions of some ZIP proteins that are expressed in KCs. In mice, ZIP2 is almost exclusively expressed in the epidermis, but not in the dermis. ZIP2 is expressed in differentiating KCs. Knockdown (KD) of ZIP2 in KCs interferes with KC differentiation and proliferation. Thus, Zn uptake through ZIP2 is essential for terminal differentiation of KCs [3]. ZIP4 is associated with the development of acrodermatitis enteropathica (AE; OMIM 201100). Although ZIP4 is highly expressed in the apical side of the intestinal epithelium, human epidermal KCs also express ZIP4, particularly in undifferentitated KCs. In human epidermal KCs, ZIP4 KD reduces intracellular Zn levels up to half, interferes with normal KC differentiation, and promotes KC proliferation, thereby leading to the parakeratosis [27]. Although these histological changes by ZIP4 KD in KCs are not identical to the histological changes in AE, ZIP4, and ZIP2 are involved in the normal KC differentiation and proliferation. Zn is deeply involved in hair biology. For example, Zn deficiency (ZnD) induces telogen effluvium and abnormal hair keratinization (reviewed in [12]). Murine ZIP10 is highly expressed in the epidermal progenitor cells located in the outer root sheath of hair follicles. Thus, depletion of ZIP10 in keratin14-expressing cells results in a thin epidermis and hair follicle hypoplasia because of downregulation of the transcriptional activity of p63, a critical regulator of epidermal formation [28]. MTs are proteins that are ubiquitously expressed throughout various types of cells and predominantly distribute in the cytoplasm, and to a lesser extent in the nuclei and lysosomes [29]. MTs are comprised of four isoforms; MT-1, MT-2, MT-3, and MT-4 [30]. In the human skin, MT-1 and MT-2 are expressed in actively proliferating cells such as hair matrix, outer hair roots, and basal layer KCs [31]. These MT expressions are further upregulated in the hyperplastic KCs of inflamed skin lesions and skin cancers in conditions such as actinic keratosis, squamous cell carcinoma, and basal cell carcinoma [32]. Knock out (KO) of both MT-1 and MT-2 in mice impairs KC proliferation [33]. Thus, MT-1 and MT-2 regulate KC proliferation, likely cooperating with ZIP2 and ZIP4 in the epidermis. Similarly, MT-3 is expressed in human epidermal KCs, and its expression appears to be upregulated in skin cancers [34,35]. MT-4 expression in murine neonatal skin is reported [36]. However, the expression and function in human skin are not understood. In summary, ZIP2 and ZIP4 in KCs facilitate KC proliferation and differentiation. ZIP10 expressed in the epidermal progenitor cells in outer root sheath is crucial for the proper epidermal formation. MTs in KCs also facilitate KC proliferation.
2.2. Langerhans Cells
Langerhans cells (LCs) are one subset of antigen-presenting cells that distribute in the epidermis [37]. LCs are involved in the development of cutaneous manifestations of acrodermatitis enteropathica (AE), a rare autosomal recessive disease. AE is caused by a mutation in the SLC39A4 gene that encodes ZIP4 followed by Zn deficiency (ZnD) because of a disability of Zn absorption within the intestines (reviewed in [38] and [39]). We found that epidermal LCs were absent in AE skin lesions. Additionally, LC loss in the epidermis was reproducible using mice with Zn-deficient diets (ZD mice) [40] (Figure 2).
In patients with AE and ZD mice, transforming growth factor-β1 (TGF-β1) expression in the epidermis was impaired compared with healthy people and mice with Zn-adequate (ZA) diets (ZA mice). TGF-β1 is essential for LC homeostasis [41]. Because several Zn finger transcription factors (e.g., ZNF580) are involved in TGF-β signaling [42], ZnD might decrease TGF-β1 production in LCs and KCs by influencing the activity of those transcription factors. Therefore, impaired TGF-β1 expression in the epidermis is, at least in part, responsible for LC loss (Figure 3). Another possibility for LC loss is apoptosis. In the steady state, epidermal LCs of ZD mice exhibited an increased apoptosis rate. Murine and human LCs underwent apoptosis when cultured with TPEN, whereas the culture condition did not induce apoptosis of KCs. This suggests that LCs are prone to apoptosis in the condition of ZnD. It is worth noting that LCs were replenished in the epidermis after Zn supplementation in patients with AE and ZD mice. Considered with these findings, severe ZnD induces LC apoptosis through a synergy of its direct effect and impaired TGF-β1 expression, leading to a disappearance of epidermal LC [40] (Figure 2 and Figure 3). In summary, Zn is critical for LC homeostasis. Despite the importance of Zn in LCs, the expression and function of ZnTs, ZIPs, and MTs in LCs are poorly understood.
2.3. Melanocytes
In an in vitro experiment, exposure of human melanocytes recovered from healthy skin to exogenous and nontoxic levels of Zn increases intracellular Zn levels, particularly in lysosomes and melanosomes. This enhances proliferation of human melanocytes with upregulated expression of AKT3, extracellular signal–regulated kinase1/2 (ERK1/2), c-MYC, and CYCD and enhances mitochondrial biosynthesis, followed by the upregulated process of autophagy [43]. In summary, Zn facilitates the melanocyte proliferation and the autophagy. The expression and function of ZnTs, ZIPs, and MTs in melanocytes are poorly understood. However, MT expression in melanoma may correlate with an increased risk of its progression and metastasis [44,45].
2.4. Mast Cells and Dendritic Cells
Besides the proliferative effect of Zn on various types of cells, Zn also functions as an intracellular signaling molecule, like calcium, by transducing extracellular stimuli into intracellular signaling (reviewed in [16,46]).
In mast cells (MCs), cytosolic Zn promotes FcεRI-induced granule translocation and subsequent degranulation. ZnT5 mRNA is abundant in MCs and its expression is upregulated by FcεRI stimulation. Zn and ZnT5 cooperatively translocate protein kinase C (PKC) to the plasma membrane, and the subsequent nuclear translocation of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), thereby promoting the production of inflammatory cytokines like IL-6 and TNF-α [47,48]. In summary, Zn and ZnT5 in MCs are involved in inflammatory cytokine production.
In dendritic cells (DCs), Zn is involved in its maturation. Upon lipopolysaccharides (LPS) stimulation of murine DCs, ZIP6 and ZIP10 are downregulated, whereas ZnT1, ZnT4, and ZnT6 are upregulated. This decreases intracellular free Zn. Because Zn promotes the endocytosis of major histocompatibility complex (MHC) class II and inhibits its trafficking from the lysosome/endosome to the plasma membrane. Decreased intracellular free Zn facilitates upregulation of surface MHC class II expression [49]. In summary, Zn and Zn transporters regulate MHC class II expression in DCs.
2.5. T cells
ZnD induces atrophy of the thymus accompanied by reduced double-positive thymocytes, decreasing the number of mature single positive T cells [50,51,52]. This is mediated by elevated glucocorticoids from adrenal glands and subsequent enhancement of apoptosis rate within double-positive thymocytes [53]. This suggests that Zn is required for the normal T cell generation [26].
Although the expressions and functions of ZnTs, ZIPs, and MTs in T cells are less understood, ZIP8 is enriched in human T cells, and its expression is markedly upregulated by in vitro activation with T cell receptor (TCR) engagement [54]. Although ZIP8 is localized to both the plasma membrane and lysosomes, ZIP8 in T cells is primarily localized to lysosomes. ZIP8 KD in T cells reduces IFN-γ production, whereas overexpression of ZIP8 in T cells enhances it [54], suggesting that Zn transport from lysosomes to the cytoplasm facilitates IFN-γ production in T cells. In summary, ZIP8 in T cells is involved in IFN-γ production.
2.6. Endothelial Cells
Zn chelation leads to death of endothelial cells (ECs). On the other hand, chronic exposure to Zn accelerates senescence of ECs in an in vitro experiment [55]. An exposure to 25 μM Zn2+ impedes the migration of human ECs [56]. Similarly, Zn oxide nanoparticles inhibit angiogenesis [57]. These data suggest that Zn is involved in the cell viability of ECs and angiogenesis.
Chronic exposure to Zn upregulates the mRNA expression of ZnT1 and ZIP6, downregulates the mRNA expression of ZnT5 and ZIP10, and does not alter the mRNA expression of ZIP1, ZIP2, and ZIP3 [55]. As for MTs in ECs, human ECs express MT-1E, MT-1X, MT-2A, and MT-3. An exposure to Zn induces the expression of most MT-1 isoforms. MT-2A KD leads to the decreased proliferation and the increased migration of ECs [58]. Collectively, Zn alters the expression of ZnTs, ZIPs, and MTs in ECs. However, the function of ZnTs and ZIPs in ECs remains to be identified.
2.7. Fibroblasts
It is known that Zn promotes lipogenesis and glucose transport via its insulin-like effects on 3T3-L1 fibroblasts and adipocytes [59]. Recent research elucidated the critical roles of ZIP7 and ZIP13 in connective tissue homeostasis. Both are intracellular Zn transporters, have quite similar amino acid sequences, and share various functional characteristics [60]. However, there are some differences. (1) ZIP7 is predominantly localized in the endoplasmic reticulum (ER), whereas ZIP13 is predominantly localized in the Golgi apparatus [60]. (2) ZIP7 is distributed throughout various types of cells, whereas ZIP13 is primarily localized in connective tissues [61,62]. ZIP7 KO mice in collagen1-expressing cells exhibit thin dermis with reduced collagen1 deposition, thin subcutaneous tissues, and a reduced number of hair follicles. Additionally, ZIP7 KD in human mesenchymal stem cells inhibits proliferation and differentiation into fibroblasts, osteoblasts, and chondrocytes [63]. As described, ZIP7 is localized in ER and transports Zn from the ER lumen to the cytoplasm. ZIP7 KD increases Zn levels in the ER, resulting in Zn-dependent aggregation and inactivation of protein disulfide isomerase, which controls proper protein folding. Thus, ER stress is overwhelming in ZIP7 KD cells, and these cells undergo apoptosis [63]. These data suggest that Zn regulation by ZIP7 in the ER is critical for the proper ER function and prevention of ER stress-induced cell death. Accordingly, the third difference between ZIP7 and ZIP13 is that ZIP7 KD induces ER stress, whereas ZIP13 KD does not induce it [64]. In conclusion, ZIP7 in fibroblasts is required for the proper dermal formation. The role of ZIP13 in fibroblasts is described later.
2.8. Adipocytes
ZIP13 is also involved in adipocyte biology. Adipocytes are divided into white and brown adipocytes. The former stores energy, whereas the latter consumes it. Additionally, other brown adipocyte-like cells, named beige adipocytes, are recently found within white adipose tissue, which is developed by various stimuli such as chronic cold exposure and long-term peroxisome proliferator-activated receptor γ (PPARγ) agonist treatment [65,66,67]. CCAAT-enhancer-binding protein-β (C/EBP-β) is induced in the early phases of adipogenesis and is necessary to activate PPARγ [68]. As described, ZIP13 is localized in the Golgi apparatus and transports Zn from the Golgi lumen to the cytoplasm. This Zn inhibits C/EBP-β activation and subsequent PPARγ activation, leading to the inhibition of beige fat cell differentiation. Conversely, ZIP13 KD facilitates beige fat cell differentiation. This enhances energy consumption, leading to the inhibition of obesity [69].
Inflammatory cytokines proliferate adipocytes, expanding cell mass through both hypertrophy and hyperplasia [70]. ZIP14 is localized to the plasma membrane, and its expression is upregulated by inflammatory cytokines and/or LPS [71]. Adipocytes from ZIP14 KO mice increase cytokine production by activating the NF-κB and signal transducer and activator of transcription 3 (STAT3) pathways [72]. These data suggest that ZIP14, which is upregulated in adipocytes during inflammation, may suppress excess inflammation.
Besides the involvement of ZIP13 and ZIP14 in adipocyte biology, many Zn finger proteins, which are proteins containing the Zn finger domains, are involved in adipogenesis [73].
3. Human Skin Disorders Caused by Mutations of Zn Transporters
Some genetic disorders are caused by mutations of Zn transporter genes. Among these disorders, mutations of ZIP4, ZIP13, and ZnT2 accompany skin manifestations.
3.1. ZIP4 Mutation; Acrodermatitis Enteropathica (AE; OMIM 201100)
As described in Section 2.2, AE is caused by loss-of-function mutations in ZIP4 followed by ZnD because of a disability of Zn absorption in the intestines. Since this discovery, over 30 mutations are reported [74]. The clinical symptoms are skin manifestations, alopecia, and diarrhea. Skin manifestations that are characterized by ZnD are referred to as acrodermatitis, and occur on periorificial, anogenital, and acral regions, where frequent contact with external substances is expected. Therefore, we presumed that acrodermatitis occurs as a consequence of contact dermatitis (CD). We utilized skin specimens of patients with AE and ZD mice to examine this hypothesis [40].
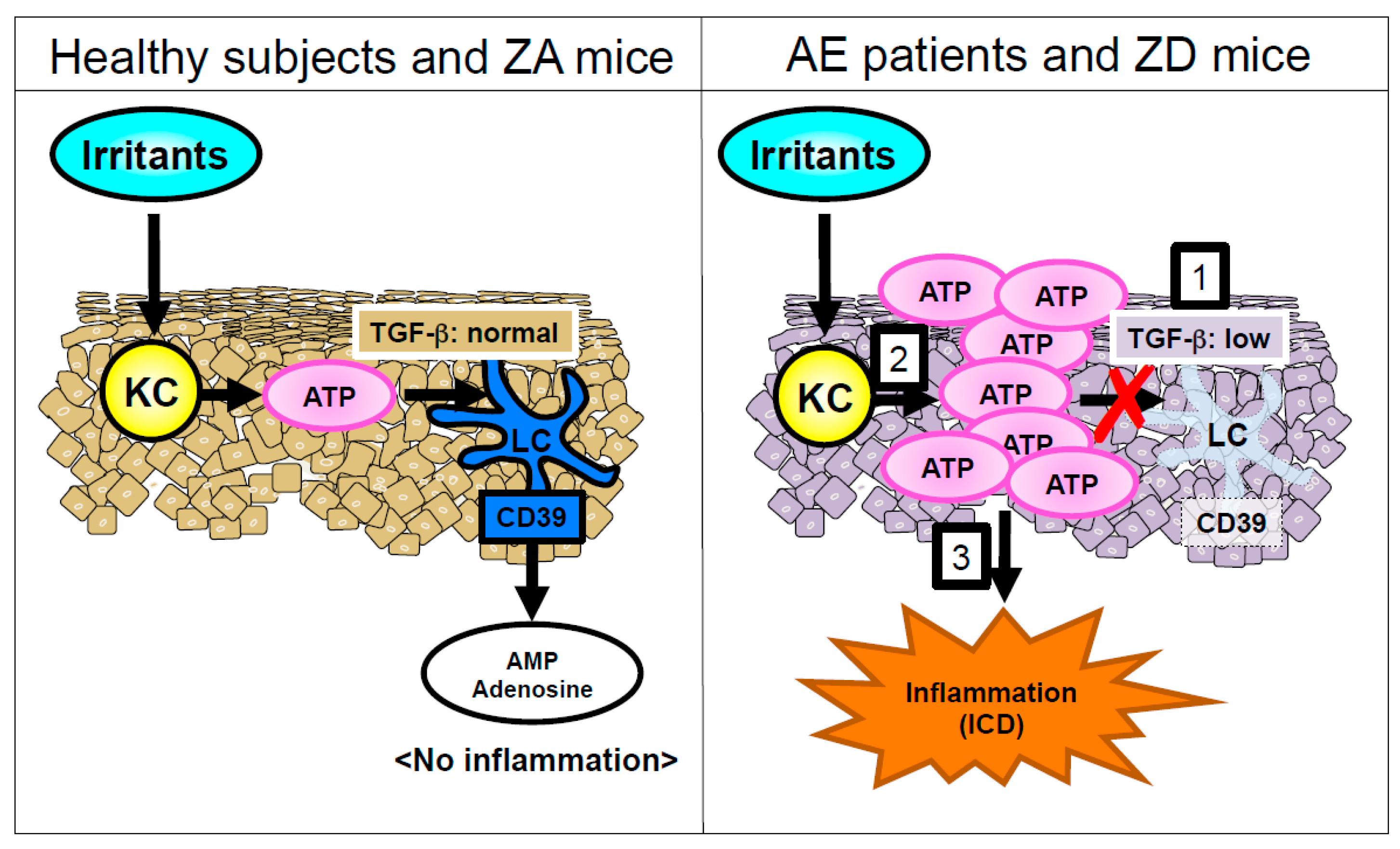
ZD mice exhibited an impaired allergic CD in response to dinitrofluorobenzene (DNFB) compared with ZA mice, because of an immunodeficiency in the ZD mice. On the other hand, ZD mice exhibited a significantly increased and prolonged irritant CD (ICD) in response to croton oil (CrO) compared with ZA mice. Adenosine triphosphate (ATP) is released from KCs in response to various environmental stimuli through lytic and non-lytic mechanisms [75,76,77]. ATP released from chemically injured mouse KCs causes ICD [77]. In an ex vivo organ culture, the released ATP amount from skin upon CrO application was much greater in the skin of ZD mice than in the skin of ZA mice. Additionally, an injection of apyrase that hydrolyzes ATP into adenosine monophosphate (AMP) restored the increased and prolonged ICD upon CrO application in ZD mice. These results suggest that the prolonged ICD response in ZD mice was mediated via the excess ATP release by KCs in response to irritants (Figure 3). As described in Section 2.2, epidermal LCs were absent in the lesioned skin of AE and ZD mice (Figure 2). LCs but not KCs express CD39 (ecto-nucleoside triphosphate diphosphohydrolase 1 (ecto-NTPDase1)) that potently hydrolyzes ATP into AMP [77,78]. Thus, the impaired ATP hydrolysis because of the disappearance of LCs leads to ATP-mediated inflammation in the epidermis, followed by the development of ICD (Figure 3).
3.2. ZIP13 Mutation; Spondylocheiro Dysplastic Form of Ehlers–Danlos Syndrome (SCD-EDS; OMIM 612350)
As described in Section 2.7, ZIP13 is localized to connective tissue cells including fibroblasts. A study with ZIP13 KO mice revealed that ZIP13 is necessary for the nuclear translocation of Smads in BMP/TGF-β signaling [61]. Thus, ZIP13 is crucial for connective tissue formation. Homozygous loss-of-function mutations of the ZIP13 gene cause SCD-EDS [61,79], characterized by hyperelastic and thin skin and hypermobility of the small joints.
3.3. ZnT2 Mutation; Transient Neonatal Zn Deficiency (TNZD; OMIM 608118)
Zn concentration in milk is maintained at higher levels than in serum, because Zn in breast milk is critical for the growth and survival of neonates [80]. In mice, the ZnT2 mutation causes severe ZnD in pups because of low Zn concentrations in breast milk. Like ZnT2-mutated mice, the breast milk from mice with a loss-of-function mutation of ZnT4 (lethal milk mutant mice; OMIM 602095) is deficient in Zn. Nursing pups of lethal milk mutant mice die before weaning [81]. Both ZnT2 and ZnT4 are localized in cytoplasmic secretory vesicles and efflux Zn from the cytoplasm to cytoplasmic secretory vesicles [7]. In humans, ZnT2 is essential for maintaining proper Zn concentrations in breast milk. Because breast milk from ZnT2-mutated mothers contains lower Zn, breast-feeding neonates show similar symptoms with AE [39,82,83,84,85]. On the other hand, there are no reports of ZnT4 involvement in Zn transport into secretory vesicles in humans.
4. Human Skin Disorders Caused by Dysregulation of Zn Transporters
Epidermodysplasia verruciformis (EV; OMIM 226400), a rare autosomal-recessive skin disease, develops non-melanoma skin cancers because of a susceptibility to oncogenic human papillomaviruses (HPVs). ZnT1 associates with the development of EV [86,87]. Although oncogenic HPVs can be detected in the skin of healthy individuals, they are asymptomatic. EV patients have mutations in either the EVER1 or EVER2 genes [88,89]. In KCs, EVER1 and 2 form a complex with ZnT1 primarily in the ER and to a lesser extent in the nuclear membrane and Golgi apparatus. The free Zn concentration in the KC nucleus is increased in patients with EV compared with healthy individuals, suggesting that the complex of ZnT1 and EVERs regulates free Zn transport in the nucleus, potentially altering cell function. A complex of ZnT1 with ‘intact’ EVERs inhibits the activator protein 1 (AP-1) activation that promotes the replication of HPV [90]. Accordingly, a complex of ZnT1 with ‘mutated’ EVERs increases free Zn transport into KC nucleus and subsequent AP-1 activity. This results in the aberrant replication of EV-related oncogenic HPVs, thereby developing skin cancers [86,87].
5. Human Skin Disorders Associated with Zn Deficiency
At present, acquired Zn deficiency (ZnD) still affects 17% of the world’s population who are in the condition of general malnutrition due to starvation, severe illness, alcohol addiction. Additionally, infants, the elderly, and pregnant women are also prone to fall into acquired ZnD [91,92,93,94]. Acquired ZnD could cause acquired AE. Additionally, ZnD is observed in diseases linked to nutritional deficiencies such as necrolytic migratory erythema (elevated serum glucagon), pellagra (niacin or tryptophan deficiency), and biotin deficiency. AE-like erythema is also observed in these diseases. Interestingly, loss or decrease of epidermal LCs that is seen in patients with AE is reported in some cases of diseases related to nutritional deficiencies. Loss of epidermal LCs and subsequent AE-like erythema are common phenomena in diseases related to nutritional deficiencies. Additionally, the nature of these abnormalities might be attributable to ZnD, because Zn supplementation restores these abnormalities. Besides these diseases related to nutritional deficiencies, ZnD is involved in other skin disorders.
5.1. Nutritional Deficiency Diseases
5.1.1. Necrolytic Migratory Erythema
Necrolytic migratory erythema (NME) has been considered as a dermadrome of pancreatic glucagonoma, because NME skin lesions could resolve if the tumor is removed [95,96,97]. Patients with glucagonoma exhibit increased serum levels of glucagon. Because glucagon is involved in the metabolism of amino acids [98], excess glucagon decreases amino acids in the serum and in the epidermis, leading to epidermal necrosis.
However, it is now clarified that NME can develop within the context of other conditions including excess inflammatory mediators, liver dysfunction, and metabolic or nutritional deficiencies, particularly of Zn, and essential amino and fatty acids [97]. Because serum glucagon levels are variable in these conditions, glucagon is not the sole causative substance of NME. Decreased serum levels of Zn are reported in patients with NME suffering from inflammatory bowel disease, celiac disease, liver dysfunction, and a malignancy other than glucagonoma [99,100,101,102,103,104,105,106,107,108,109,110,111]. Supplementation of Zn restores skin lesions of NME, implying that decreased serum levels of Zn associate with the pathogenesis of NME. As described, AE-like erythema in patients with NME and its histopathological findings—such as cytoplasmic pallor, parakeratosis, subcorneal vacuolization, and ballooning degeneration of KCs—are identical to AE. In addition, one report demonstrates that epidermal LC number is reduced in patients with NME whose serum levels of Zn are decreased [111]. This suggests that similar mechanisms may underlie the development of skin lesions in patients with AE (described in Section 2.2 and Section 3.1) and NME.
5.1.2. Pellagra
The compounds that have anti-pellagra activity are called niacin, and the major compound of niacin is nicotinamide and nicotinic acid. Nicotinamide is generated by two pathways. (1) Dietary nicotinic acid is promptly incorporated into the liver and is converted to nicotinamide; (2) Nicotinamide is synthesized from tryptophan, an essential amino acid, by the tryptophan–nicotinamide conversion pathway [112].
Deficiency of niacin and/or tryptophan contributes to pellagra. Diets rich in corn contain less niacin and tryptophan, contributing to the development of pellagra [113]. Patients with pellagra show AE-like erythema, diarrhea, and dementia [114]. Photosensitivity is a unique phenomenon in patients with pellagra and is not seen in other diseases associated with nutritional deficiencies. Niacin deficiency induces reactive oxygen species in KCs, followed by the production of prostaglandin E2 (PGE2). This PGE2 mediates photosensitivity in patients with pellagra [115]. A study investigating serum Zn levels in 81 pellagra patients showed a significant reduction of serum Zn level in patients with pellagra (69.7 ± 16.8 μg/dL) compared with that in healthy subjects (82.3 ± 34.0 μg/dL) [116]. Although the average serum Zn levels in patients with pellagra are in the range of latent ZnD, ZnD may be involved in the development of pellagra. Like AE and NME, loss or decrease of epidermal LCs is reported in the lesional skin of pellagra, but not in non-lesional skin of pellagra [117].
5.1.3. Biotin Deficiency
Biotin is a water-soluble vitamin and serves as a co-enzyme for five carboxylases in humans. Biotin-dependent carboxylases are involved in various metabolic pathways such as gluconeogenesis, fatty acid synthesis, and amino acid synthesis. Mammals cannot synthesize biotin. However, because biotin is contained in a wide range of foods and some gut microbiota produce biotin, biotin deficiency (BnD) does not occur in people who consume a mixed general diet. On the other hand, genetic deficiency of holocarboxylase synthetase and biotinidase; continuous consumption of raw egg whites; parenteral nutrition; and modified milk without biotin supplementation can lead to the development of BnD [118]. BnD causes similar symptoms as AE including skin lesions, alopecia, and diarrhea [119]. As for the underlying mechanism of development of the skin lesions in BnD, BnD causes abnormalities in fatty acid composition such as accumulation of odd-chain fatty acids and abnormal metabolism of long-chain polyunsaturated fatty acids [120,121,122,123]. Besides fatty acid abnormalities, ZnD is reported in some patients with BnD [124,125,126]. Although the serum Zn levels in patients with BnD are inconsistent among reports [127,128,129], ZnD might contribute to the development of BnD when considering their similar symptom profiles.
5.2. Alopecia
Alopecia is roughly classified into non-scarring and scarring. The former includes telogen effluvium, alopecia areata, and androgenic alopecia [130]. ZnD is related to some non-scarring alopecia.
5.2.1. Alopecia in Acrodermatitis Enteropathica
Alopecia developed in patients with acrodermatitis enteropathica (AE) shows the characteristics of telogen effluvium (TE), which is a type of non-scarring alopecia and is defined by the premature transition of anagen to telogen phase [131,132]. Patients with TE without AE also exhibit decreased serum levels of Zn compared with healthy individuals [132,133]. TE can be restored by supplementation of Zn [132]. In patients with AE, hair contains less Zn [134] and hair shafts show a characteristic irregular pattern because of impaired incorporation of cystine, a major content of hair keratin amino acids that is required for normal hair keratinization [135,136]. This is evidenced by impaired incorporation of radiolabeled cystine into hair in ZD rats [137,138,139]. Although alopecia-like hair loss is reported in dietary ZD AE models using mice [140], rats [141,142], and rabbits [143], no studies have addressed the underlying mechanisms. In summary, ZnD induces TE and abnormal hair keratinization.
5.2.2. Alopecia Areata
Alopecia areata (AA) is an autoimmune disease mediated by cytotoxic T lymphocytes [144]. IFN-γ KO mice do not develop experimental AA [145,146]. The association between AA and lower serum levels of Zn is a subject of active inquiry. The results of multiple analyses of serum Zn levels and AA are contradictory (reviewed in [12]). However, evidence indicates that serum levels of Zn are certainly decreased in patients with severe AA whose alopecia is broad and prolonged and is resistant to conventional therapies [147,148,149]. This implies that the serum levels of Zn are a useful parameter to predict the severity and that supplementation of Zn can be a promising adjuvant therapy, along with standard therapy for severe AA.
5.3. Cutaneous Wounds and Ulcers
The process of wound healing is complex and involves various Zn-related molecules, such as MTs, matrix metalloproteinases, integrins, alkaline phosphatase, and Zn finger proteins [150,151]. Oral and/or topical Zn has been long used to treat ulcers and wounds. Unexpectedly, a systematic review using data from the Cochrane Wound Group failed to conclude that oral Zn supplementation improves wound healing [152]. However, studies show the efficacy of topical Zn oxide for improving rates of wound healing, regardless of serum Zn levels of patients [153,154].
As described in Section 2.4, Zn acts as an intracellular signaling molecule. G-protein coupled receptor 39 (GPR39) has been identified as a Zn-sensing receptor [155]. GPR39 is an orphan G protein-coupled receptor that is conserved in vertebrates and transduces autocrine and paracrine Zn signals [155]. In an in vitro experiment, Zn released from injured KCs stimulates GPR39, and this signaling promotes epithelial repair [156]. Stem cells in the skin are localized in various regions including the interfollicular epidermis, hair follicles, sweat glands, and sebaceous glands (SGs) [157]. These stem cells are critical for wound repair [157]. In murine skin, GPR39 is exclusively localized to SGs and is co-localized with stem cells in SGs that express Blimp1. GPR39 KO mice show delayed wound healing [158]. These data demonstrate that Zn signaling through GPR39 plays an important role in wound healing by stimulating stem cell activity in SGs.
5.4. Other Skin Disorders Associated with Zn Deficiency
Zn deficiency (ZnD) is reported in some skin disorders including inflammatory diseases (atopic dermatitis [159,160], oral lichen planus [161], and Behcet’s disease [162,163]), autoimmune bullous diseases (pemphigus vulgaris [164] and bullous pemphigoid [165]), inherited bullous diseases (inherited epidermolysis bullosa [166,167]), and hyperpigmentation (melasma [168]). Because Zn is deeply involved in the regulation of immune systems, it is likely that ZnD leads to the development of these inflammatory and autoimmune disorders [26,169].
Accumulation of evidence is required to determine the relationship between Zn and these skin disorders.
6. Conclusions
The association of skin manifestations and ZnD is well known. AE is caused by the mutations of ZIP4 and subsequent ZnD. Substantial numbers of patients suffering from diseases related to nutritional deficiencies such as acquired AE, NME, pellagra, and BnD show low serum Zn levels. This suggests that ZnD affects any cell type. Additionally, many micronutrients are also deficient in the conditions that fall into nutritional ZnD. Patients with these disorders share common skin manifestations of acrodermatitis. Additionally, LC loss in the epidermis is reported in patients with AE, NME, and pellagra. LCs are the sole CD39 (ecto-NTPDase1)-expressing cells in the epidermis. Thus, the disappearance of LCs in the epidermis exacerbates skin inflammation after irritant exposure (Figure 2 and Figure 3). In conclusion, acrodermatitis in diseases associated with nutritional deficiencies may be caused by ICD, secondary to LC loss.
Although every cell expresses many different Zn transporters, which all contribute to homeostatic control of the cells and tissues, the role of Zn transporters in skin homeostasis is less understood. However, recent studies add some important information to this field (Figure 1). For instance, ZIP2 and ZIP4 contribute to KC proliferation and differentiation. ZIP10 is critical for skin homeostasis and epidermal formation. ZIP7 is important for proper dermal formation. ZIP13 is involved in adipocyte biology.
ZnD is a current problem in both developing and developed countries. We have to pay attention to cutaneous symptoms in order not to miss the ‘dermadrome’ of ZnD.
Author Contributions
Y.O. and T.K. conceived and constructed this manuscript with the assistance of M.K. and S.S.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jackson, M.J. Physiology of zinc: General aspects. In Zinc in Human Biology; Mills, C.F., Ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1989; pp. 1–14. [Google Scholar]
- Michaelsson, G.; Ljunghall, K.; Danielson, B.G. Zinc in epidermis and dermis in healthy subjects. Acta Derm. Venereol. 1980, 60, 295–299. [Google Scholar] [PubMed]
- Inoue, Y.; Hasegawa, S.; Ban, S.; Yamada, T.; Date, Y. ZIP2 protein, a zinc transporter, is associated with keratinocyte differentiation. J. Biol. Chem. 2014, 289, 21451–21462. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, G.T. Heavy metals in rat mast cell granules. Lab. Investig. 1967, 17, 588–598. [Google Scholar] [PubMed]
- Cowen, T.; Trigg, P.; Eady, R.A. Distribution of mast cells in human dermis: Development of a mapping technique. Br. J. Dermatol. 1979, 100, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Weber, A.; Knop, J.; Maurer, M. Pattern analysis of human cutaneous mast cell populations by total body surface mapping. Br. J. Dermatol. 2003, 148, 224–228. [Google Scholar] [CrossRef] [PubMed]
- Kambe, T.; Tsuji, T.; Hashimoto, A.; Itsumura, N. The physiological, biochemical, and molecular roles of zinc transporters in zinc homeostasis and metabolism. Physiol. Rev. 2015, 95, 749–784. [Google Scholar] [CrossRef] [PubMed]
- Hediger, M.A.; Romero, M.F.; Peng, J.B.; Rolfs, A.; Takanaga, H.; Bruford, E.A. The ABCs of solute carriers: Physiological, pathological and therapeutic implications of human membrane transport proteins. Pflugers Arch. 2004, 447, 465–468. [Google Scholar] [CrossRef] [PubMed]
- Eide, D.J. The SLC39 family of metal ion transporters. Pflugers Arch. 2004, 447, 796–800. [Google Scholar] [CrossRef] [PubMed]
- Palmiter, R.D.; Huang, L. Efflux and compartmentalization of zinc by members of the SLC30 family of solute carriers. Pflugers Arch. 2004, 447, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Tapiero, H.; Tew, K.D. Trace elements in human physiology and pathology: Zinc and metallothioneins. Biomed. Pharmacother. 2003, 57, 399–411. [Google Scholar] [CrossRef]
- Ogawa, Y.; Kawamura, T.; Shimada, S. Zinc and skin biology. Arch. Biochem. Biophys. 2016, 611, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Andreini, C.; Bertini, I.; Cavallaro, G. Minimal functional sites allow a classification of zinc sites in proteins. PLoS ONE 2011, 6, e26325. [Google Scholar] [CrossRef] [PubMed]
- Andreini, C.; Bertini, I. A bioinformatics view of zinc enzymes. J. Inorg. Biochem. 2012, 111, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Berg, J.M.; Shi, Y. The galvanization of biology: A growing appreciation for the roles of zinc. Science 1996, 271, 1081–1085. [Google Scholar] [CrossRef] [PubMed]
- Fukada, T.; Yamasaki, S.; Nishida, K.; Murakami, M.; Hirano, T. Zinc homeostasis and signaling in health and diseases: Zinc signaling. J. Biol. Inorg. Chem. 2011, 16, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Klug, A. The discovery of zinc fingers and their applications in gene regulation and genome manipulation. Annu. Rev. Biochem. 2010, 79, 213–231. [Google Scholar] [CrossRef] [PubMed]
- Laity, J.H.; Lee, B.M.; Wright, P.E. Zinc finger proteins: New insights into structural and functional diversity. Curr. Opin. Struct. Biol. 2001, 11, 39–46. [Google Scholar] [CrossRef]
- Andreini, C.; Banci, L.; Bertini, I.; Rosato, A. Counting the zinc-proteins encoded in the human genome. J. Proteome Res. 2006, 5, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Emri, E.; Miko, E.; Bai, P.; Boros, G.; Nagy, G.; Rózsa, D.; Juhász, T.; Hegedűs, C.; Horkay, I.; Remenyik, É.; et al. Effects of non-toxic zinc exposure on human epidermal keratinocytes. Metallomics 2015, 7, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Chai, F.; Truong-Tran, A.Q.; Evdokiou, A. Intracellular zinc depletion induces caspase activation and p21Waf1/Cip1 cleavage in human epithelial cell lines. J. Infect. Dis. 2000, 182 (Suppl. 1), S85–S92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, D.; Varigos, G.; Ackland, M.L. Apoptosis may underlie the pathology of zinc-deficient skin. Immunol. Cell Biol. 2006, 84, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Hsu, D.J.; Daniel, J.C.; Gerson, S.J. Effect of zinc deficiency on keratins in buccal epithelium of rats. Arch. Oral Biol. 1991, 36, 759–763. [Google Scholar] [CrossRef]
- Gueniche, A.; Viac, J.; Lizard, G.; Charveron, M.; Schmitt, D. Protective effect of zinc on keratinocyte activation markers induced by interferon or nickel. Acta Derm. Venereol. 1995, 75, 19–23. [Google Scholar] [PubMed]
- Yamaoka, J.; Kume, T.; Akaike, A.; Miyachi, Y. Suppressive effect of zinc ion on iNOS expression induced by interferon-γ or tumor necrosis factor-α in murine keratinocytes. J. Dermatol. Sci. 2000, 23, 27–35. [Google Scholar] [CrossRef]
- Wessels, I.; Maywald, M.; Rink, L. Zinc as a gatekeeper of immune function. Nutrients 2017, 9, 1286. [Google Scholar] [CrossRef] [PubMed]
- Bin, B.H.; Bhin, J.; Kim, N.H.; Lee, S.-H.; Jung, H.-S.; Seo, J.; Kim, D.-K.; Hwang, D.; Fukada, T.; Lee, A.-Y.; et al. An acrodermatitis enteropathica-associated Zn transporter, ZIP4, Regulates human epidermal homeostasis. J. Investig. Dermatol. 2017, 137, 874–883. [Google Scholar] [CrossRef] [PubMed]
- Bin, B.H.; Bhin, J.; Takaishi, M.; Toyoshima, K.-E.; Kawamata, S.; Ito, K.; Hara, T.; Watanabe, T.; Irié, T.; Takagishi, T.; et al. Requirement of zinc transporter ZIP10 for epidermal development: Implication of the ZIP10-p63 axis in epithelial homeostasis. Proc. Natl. Acad. Sci. USA 2017, 114, 12243–12248. [Google Scholar] [CrossRef] [PubMed]
- Nartey, N.O.; Banerjee, D.; Cherian, M.G. Immunohistochemical localization of metallothionein in cell nucleus and cytoplasm of fetal human liver and kidney and its changes during development. Pathology 1987, 19, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Thirumoorthy, N.; Shyam Sunder, A.; Manisenthil Kumar, K.; Senthil kumar, M.; Ganesh, G.N.K.; Chatterjee, M. A review of metallothionein isoforms and their role in pathophysiology. World J. Surg. Oncol. 2011, 9, 54. [Google Scholar] [CrossRef] [PubMed]
- Van den Oord, J.J.; De Ley, M. Distribution of metallothionein in normal and pathological human skin. Arch. Dermatol. Res. 1994, 286, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Zamirska, A.; Matusiak, L.; Dziegiel, P.; Szybejko-Machaj, G.; Szepietowski, J.C. Expression of metallothioneins in cutaneous squamous cell carcinoma and actinic keratosis. Pathol. Oncol. Res. 2012, 18, 849–855. [Google Scholar] [CrossRef] [PubMed]
- Hanada, K.; Sawamura, D.; Hashimoto, I.; Kida, K.; Naganuma, A. Epidermal proliferation of the skin in metallothionein-null mice. J. Investig. Dermatol. 1998, 110, 259–262. [Google Scholar] [CrossRef] [PubMed]
- Slusser, A.; Zheng, Y.; Zhou, X.D.; Somji, S.; Sens, D.A.; Sens, M.A.; Garrett, S.H. Metallothionein isoform 3 expression in human skin, related cancers and human skin derived cell cultures. Toxicol. Lett. 2015, 232, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Pula, B.; Tazbierski, T.; Zamirska, A.; Werynska, B.; Bieniek, A.; Szepietowski, J.; Rys, J.; Dziegiel, P.; Podhorska-Okolow, M. Metallothionein 3 expression in normal skin and malignant skin lesions. Pathol. Oncol. Res. 2015, 21, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Quaife, C.J.; Findley, S.D.; Erickson, J.C.; Froelick, G.J.; Kelly, E.J.; Zambrowicz, B.P.; Palmiter, R.D. Induction of a new metallothionein isoform (MT-IV) occurs during differentiation of stratified squamous epithelia. Biochemistry 1994, 33, 7250–7259. [Google Scholar] [CrossRef] [PubMed]
- Merad, M.; Ginhoux, F.; Collin, M. Origin, homeostasis and function of Langerhans cells and other langerin-expressing dendritic cells. Nat. Rev. Immunol. 2008, 8, 935–947. [Google Scholar] [CrossRef] [PubMed]
- Maverakis, E.; Fung, M.A.; Lynch, P.J.; Draznin, M.; Michael, D.J.; Ruben, B.; Fazel, N. Acrodermatitis enteropathica and an overview of zinc metabolism. J. Am. Acad. Dermatol. 2007, 56, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Kasana, S.; Din, J.; Maret, W. Genetic causes and gene-nutrient interactions in mammalian zinc deficiencies: Acrodermatitis enteropathica and transient neonatal zinc deficiency as examples. J. Trace Elem. Med. Biol. 2015, 29, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, T.; Ogawa, Y.; Nakamura, Y.; Nakamizo, S.; Ohta, Y.; Nakano, H.; Kabashima, K.; Katayama, I.; Koizumi, S.; Kodama, T.; et al. Severe dermatitis with loss of epidermal Langerhans cells in human and mouse zinc deficiency. J. Clin. Investig. 2012, 122, 722–732. [Google Scholar] [CrossRef] [PubMed]
- Borkowski, T.A.; Letterio, J.J.; Farr, A.G.; Udey, M.C. A role for endogenous transforming growth factor β1 in Langerhans cell biology: The skin of transforming growth factor β1 null mice is devoid of epidermal Langerhans cells. J. Exp. Med. 1996, 184, 2417–2422. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Hu, W.; Xu, R.; Hou, B.; Zhang, L.; Zhang, W. ZNF580, a novel C2H2 zinc-finger transcription factor, interacts with the TGF-β signal molecule Smad2. Cell Biol. Int. 2011, 35, 1153–1157. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, E.; Rudolf, K. Increases in intracellular zinc enhance proliferative signaling as well as mitochondrial and endolysosomal activity in human melanocytes. Cell. Physiol. Biochem. 2017, 43, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Weinlich, G.; Bitterlich, W.; Mayr, V.; Fritsch, P.O.; Zelger, B. Metallothionein-overexpression as a prognostic factor for progression and survival in melanoma. A prospective study on 520 patients. Br. J. Dermatol. 2003, 149, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Emri, E.; Egervari, K.; Varvolgyi, T.; Rozsa, D.; Miko, E.; Dezso, B.; Veres, I.; Mehes, G.; Emri, G.; Remenyik, E. Correlation among metallothionein expression, intratumoural macrophage infiltration and the risk of metastasis in human cutaneous malignant melanoma. J. Eur. Acad. Dermatol. Venereol. 2013, 27, e320–e327. [Google Scholar] [CrossRef] [PubMed]
- Hojyo, S.; Fukada, T. Roles of zinc signaling in the immune system. J. Immunol. Res. 2016, 2016, 6762343. [Google Scholar] [CrossRef] [PubMed]
- Nishida, K.; Hasegawa, A.; Nakae, S.; Oboki, K.; Saito, H.; Yamasaki, S.; Hirano, T. Zinc transporter Znt5/Slc30a5 is required for the mast cell-mediated delayed-type allergic reaction but not the immediate-type reaction. J. Exp. Med. 2009, 206, 1351–1364. [Google Scholar] [CrossRef] [PubMed]
- Kabu, K.; Yamasaki, S.; Kamimura, D.; Ito, Y.; Hasegawa, A.; Sato, E.; Kitamura, H.; Nishida, K.; Hirano, T. Zinc is required for FcεRI-mediated mast cell activation. J. Immunol. 2006, 177, 1296–1305. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, H.; Morikawa, H.; Kamon, H.; Iguchi, M.; Hojyo, S.; Fukada, T.; Yamashita, S.; Kaisho, T.; Akira, S.; Murakami, M.; et al. Toll-like receptor-mediated regulation of zinc homeostasis influences dendritic cell function. Nat. Immunol. 2006, 7, 971–977. [Google Scholar] [CrossRef] [PubMed]
- Dowd, P.S.; Kelleher, J.; Guillou, P.J. T-lymphocyte subsets and interleukin-2 production in zinc-deficient rats. Br. J. Nutr. 1986, 55, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, G.; Nair, M.; Onoe, K.; Tanaka, T.; Floyd, R.; Good, R.A. Impairment of cell-mediated immunity functions by dietary zinc deficiency in mice. Proc. Natl. Acad. Sci. USA 1979, 76, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Golden, M.H.; Jackson, A.A.; Golden, B.E. Effect of zinc on thymus of recently malnourished children. Lancet 1977, 2, 1057–1059. [Google Scholar] [CrossRef]
- DePasquale-Jardieu, P.; Fraker, P.J. The role of corticosterone in the loss in immune function in the zinc-deficient A/J mouse. J. Nutr. 1979, 109, 1847–1855. [Google Scholar] [CrossRef] [PubMed]
- Aydemir, T.B.; Liuzzi, J.P.; McClellan, S.; Cousins, R.J. Zinc transporter ZIP8 (SLC39A8) and zinc influence IFN-γ expression in activated human T cells. J. Leukoc. Biol. 2009, 86, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Malavolta, M.; Costarelli, L.; Giacconi, R.; Basso, A.; Piacenza, F.; Pierpaoli, E.; Provinciali, M.; Ogo, O.A.; Ford, D. Changes in Zn homeostasis during long term culture of primary endothelial cells and effects of Zn on endothelial cell senescence. Exp. Gerontol. 2017, 99, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Shearier, E.R.; Bowen, P.K.; He, W.; Drelich, A.; Drelich, J.; Goldman, J.; Zhao, F. In vitro cytotoxicity, adhesion, and proliferation of human vascular cells exposed to zinc. ACS Biomater. Sci. Eng. 2016, 2, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Tada-Oikawa, S.; Ichihara, G.; Suzuki, Y.; Izuoka, K.; Wu, W.; Yamada, Y.; Mishima, T.; Ichihara, S. Zn(II) released from zinc oxide nano/micro particles suppresses vasculogenesis in human endothelial colony-forming cells. Toxicol. Rep. 2015, 2, 692–701. [Google Scholar] [CrossRef] [PubMed]
- Schulkens, I.A.; Castricum, K.C.; Weijers, E.M.; Koolwijk, P.; Griffioen, A.W.; Thijssen, V.L. Expression, regulation and function of human metallothioneins in endothelial cells. J. Vasc. Res. 2014, 51, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Shay, N.F. Zinc has an insulin-like effect on glucose transport mediated by phosphoinositol-3-kinase and Akt in 3T3-L1 fibroblasts and adipocytes. J. Nutr. 2001, 131, 1414–1420. [Google Scholar] [CrossRef] [PubMed]
- Bin, B.H.; Fukada, T.; Hosaka, T.; Yamasaki, S.; Ohashi, W.; Hojyo, S.; Miyai, T.; Nishida, K.; Yokoyama, S.; Hirano, T. Biochemical characterization of human ZIP13 protein: A homo-dimerized zinc transporter involved in the spondylocheiro dysplastic Ehlers-Danlos syndrome. J. Biol. Chem. 2011, 286, 40255–40265. [Google Scholar] [CrossRef] [PubMed]
- Fukada, T.; Civic, N.; Furuichi, T.; Shimoda, S.; Mishima, K.; Higashiyama, H.; Idaira, Y.; Asada, Y.; Kitamura, H.; Yamasaki, S.; et al. The zinc transporter SLC39A13/ZIP13 is required for connective tissue development; its involvement in BMP/TGF-β signaling pathways. PLoS ONE 2008, 3, e3642. [Google Scholar] [CrossRef]
- Taylor, K.M.; Morgan, H.E.; Johnson, A.; Nicholson, R.I. Structure-function analysis of HKE4, A member of the new LIV-1 subfamily of zinc transporters. Biochem. J. 2004, 377, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Bin, B.H.; Bhin, J.; Seo, J.; Kim, S.Y.; Lee, E.; Park, K.; Choi, D.H.; Takagishi, T.; Hara, T.; Hwang, D.; et al. Requirement of zinc transporter SLC39A7/ZIP7 for dermal development to fine-tune endoplasmic reticulum function by regulating protein disulfide isomerase. J. Investig. Dermatol. 2017, 137, 1682–1691. [Google Scholar] [CrossRef] [PubMed]
- Bin, B.H.; Hojyo, S.; Hosaka, T.; Shimoda, S.; Mishima, K.; Higashiyama, H.; Idaira, Y.; Asada, Y.; Kitamura, H.; Yamasaki, S.; et al. Molecular pathogenesis of spondylocheirodysplastic Ehlers-Danlos syndrome caused by mutant ZIP13 proteins. EMBO Mol. Med. 2014, 6, 1028–1042. [Google Scholar] [CrossRef] [PubMed]
- Kajimura, S.; Spiegelman, B.M.; Seale, P. Brown and beige fat: Physiological roles beyond heat generation. Cell Metab. 2015, 22, 546–559. [Google Scholar] [CrossRef] [PubMed]
- Harms, M.; Seale, P. Brown and beige fat: Development, function and therapeutic potential. Nat. Med. 2013, 19, 1252–1263. [Google Scholar] [CrossRef] [PubMed]
- Ohno, H.; Shinoda, K.; Spiegelman, B.M.; Kajimura, S. PPARγ agonists induce a white-to-brown fat conversion through stabilization of PRDM16 protein. Cell Metab. 2012, 15, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.Q.; Lane, M.D. Adipogenesis: From stem cell to adipocyte. Annu. Rev. Biochem. 2012, 81, 715–736. [Google Scholar] [CrossRef] [PubMed]
- Fukunaka, A.; Fukada, T.; Bhin, J.; Suzuki, L.; Tsuzuki, T.; Takamine, Y.; Bin, B.H.; Yoshihara, T.; Ichinoseki-Sekine, N.; Naito, H.; et al. Zinc transporter ZIP13 suppresses beige adipocyte biogenesis and energy expenditure by regulating C/EBP-β expression. PLoS Genet. 2017, 13, e1006950. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.; Lapoint, K.; Martinez, K.; Kennedy, A.; Boysen Sandberg, M.; McIntosh, M.K. Preadipocytes mediate lipopolysaccharide-induced inflammation and insulin resistance in primary cultures of newly differentiated human adipocytes. Endocrinology 2006, 147, 5340–5351. [Google Scholar] [CrossRef] [PubMed]
- Liuzzi, J.P.; Lichten, L.A.; Rivera, S.; Blanchard, R.K.; Aydemir, T.B.; Knutson, M.D.; Ganz, T.; Cousins, R.J. Interleukin-6 regulates the zinc transporter Zip14 in liver and contributes to the hypozincemia of the acute-phase response. Proc. Natl. Acad. Sci. USA 2005, 102, 6843–6848. [Google Scholar] [CrossRef] [PubMed]
- Troche, C.; Aydemir, T.B.; Cousins, R.J. Zinc transporter Slc39a14 regulates inflammatory signaling associated with hypertrophic adiposity. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E258–E268. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Zhang, L.; Zhou, X.; Du, M.; Jiang, Z.; Hausman, G.J.; Bergen, W.G.; Zan, L.; Dodson, M.V. Emerging roles of zinc finger proteins in regulating adipogenesis. Cell. Mol. Life Sci. 2013, 70, 4569–4584. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, S.; Kury, S.; Giraud, M.; Dreno, B.; Kharfi, M.; Bezieau, S. An update on mutations of the SLC39A4 gene in acrodermatitis enteropathica. Hum. Mutat. 2009, 30, 926–933. [Google Scholar] [CrossRef] [PubMed]
- Lazarowski, E.R.; Boucher, R.C.; Harden, T.K. Mechanisms of release of nucleotides and integration of their action as P2X- and P2Y-receptor activating molecules. Mol. Pharmacol. 2003, 64, 785–795. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, S.; Fujishita, K.; Inoue, K.; Shigemoto-Mogami, Y.; Tsuda, M.; Inoue, K. Ca2+ waves in keratinocytes are transmitted to sensory neurons: The involvement of extracellular ATP and P2Y2 receptor activation. Biochem. J. 2004, 380, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Mizumoto, N.; Kumamoto, T.; Robson, S.C.; Sevigny, J.; Matsue, H.; Enjyoji, K.; Takashima, A. CD39 is the dominant Langerhans cell-associated ecto-NTPDase: Modulatory roles in inflammation and immune responsiveness. Nat. Med. 2002, 8, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.L.; Yang, C.Y.; Lin, W.J.; Lin, C.H. Ecto-nucleoside triphosphate diphosphohydrolase 2 modulates local ATP-induced calcium signaling in human HaCaT keratinocytes. PLoS ONE 2013, 8, e57666. [Google Scholar] [CrossRef] [PubMed]
- Giunta, C.; Elcioglu, N.H.; Albrecht, B.; Eich, G.; Chambaz, C.; Janecke, A.R.; Yeowell, H.; Weis, M.; Eyre, D.R.; Kraenzlin, M.; et al. Spondylocheiro dysplastic form of the Ehlers-Danlos syndrome—An autosomal-recessive entity caused by mutations in the zinc transporter gene SLC39A13. Am. J. Hum. Genet. 2008, 82, 1290–1305. [Google Scholar] [CrossRef] [PubMed]
- Yamawaki, N.; Yamada, M.; Kan-no, T.; Kojima, T.; Kaneko, T.; Yonekubo, A. Macronutrient, mineral and trace element composition of breast milk from Japanese women. J. Trace Elem. Med. Biol. 2005, 19, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Gitschier, J. A novel gene involved in zinc transport is deficient in the lethal milk mouse. Nat. Genet. 1997, 17, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Chowanadisai, W.; Lonnerdal, B.; Kelleher, S.L. Identification of a mutation in SLC30A2 (ZnT-2) in women with low milk zinc concentration that results in transient neonatal zinc deficiency. J. Biol. Chem. 2006, 281, 39699–39707. [Google Scholar] [CrossRef] [PubMed]
- Lasry, I.; Seo, Y.A.; Ityel, H.; Shalva, N.; Pode-Shakked, B.; Glaser, F.; Berman, B.; Berezovsky, I.; Goncearenco, A.; Klar, A.; et al. A dominant negative heterozygous G87R mutation in the zinc transporter, ZnT-2 (SLC30A2), results in transient neonatal zinc deficiency. J. Biol. Chem. 2012, 287, 29348–29361. [Google Scholar] [CrossRef] [PubMed]
- Itsumura, N.; Inamo, Y.; Okazaki, F.; Teranishi, F.; Narita, H.; Kambe, T.; Kodama, H. Compound heterozygous mutations in SLC30A2/ZnT2 results in low milk zinc concentrations: A novel mechanism for zinc deficiency in a breast-fed infant. PLoS ONE 2013, 8, e64045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miletta, M.C.; Bieri, A.; Kernland, K.; Schoni, M.H.; Petkovic, V.; Fluck, C.E.; Eble, A.; Mullis, P.E. Transient neonatal zinc deficiency caused by a heterozygous G87R mutation in the Zinc transporter ZnT-2 (SLC30A2) Gene in the mother highlighting the importance of Zn2+ for normal growth and development. Int. J. Endocrinol. 2013, 2013, 259189. [Google Scholar] [CrossRef] [PubMed]
- Lazarczyk, M.; Pons, C.; Mendoza, J.A.; Cassonnet, P.; Jacob, Y.; Favre, M. Regulation of cellular zinc balance as a potential mechanism of EVER-mediated protection against pathogenesis by cutaneous oncogenic human papillomaviruses. J. Exp. Med. 2008, 205, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Orth, G. Host defenses against human papillomaviruses: Lessons from epidermodysplasia verruciformis. Curr. Top. Microbiol. Immunol. 2008, 321, 59–83. [Google Scholar] [PubMed]
- Ramoz, N.; Taieb, A.; Rueda, L.A.; Montoya, L.S.; Bouadjar, B.; Favre, M.; Orth, G. Evidence for a nonallelic heterogeneity of epidermodysplasia verruciformis with two susceptibility loci mapped to chromosome regions 2p21-p24 and 17q25. J. Investig. Dermatol. 2000, 114, 1148–1153. [Google Scholar] [CrossRef] [PubMed]
- Ramoz, N.; Rueda, L.A.; Bouadjar, B.; Montoya, L.S.; Orth, G.; Favre, M. Mutations in two adjacent novel genes are associated with epidermodysplasia verruciformis. Nat. Genet. 2002, 32, 579–581. [Google Scholar] [CrossRef] [PubMed]
- Offord, E.A.; Beard, P. A member of the activator protein 1 family found in keratinocytes but not in fibroblasts required for transcription from a human papillomavirus type 18 promoter. J. Virol. 1990, 64, 4792–4798. [Google Scholar] [PubMed]
- Stammers, A.L.; Lowe, N.M.; Medina, M.W.; Patel, S.; Dykes, F.; Perez-Rodrigo, C.; Serra-Majam, L.; Nissensohn, M.; Moran, V.H. The relationship between zinc intake and growth in children aged 1–8 years: A systematic review and meta-analysis. Eur. J. Clin. Nutr. 2015, 69, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Krebs, N.F.; Miller, L.V.; Hambidge, K.M. Zinc deficiency in infants and children: A review of its complex and synergistic interactions. Paediatr. Int. Child Health 2014, 34, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Penny, M.E. Zinc supplementation in public health. Ann. Nutr. Metab. 2013, 62 (Suppl. 1), 31–42. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hu, Y.F.; Hao, J.H.; Chen, Y.H.; Su, P.Y.; Wang, Y.; Yu, Z.; Fu, L.; Xu, Y.Y.; Zhang, C.; et al. Maternal zinc deficiency during pregnancy elevates the risks of fetal growth restriction: A population-based birth cohort study. Sci. Rep. 2015, 5, 11262. [Google Scholar] [CrossRef] [PubMed]
- Mullans, E.A.; Cohen, P.R. Iatrogenic necrolytic migratory erythema: A case report and review of nonglucagonoma-associated necrolytic migratory erythema. J. Am. Acad. Dermatol. 1998, 38, 866–873. [Google Scholar] [CrossRef]
- Alexander, E.K.; Robinson, M.; Staniec, M.; Dluhy, R.G. Peripheral amino acid and fatty acid infusion for the treatment of necrolytic migratory erythema in the glucagonoma syndrome. Clin. Endocrinol. 2002, 57, 827–831. [Google Scholar] [CrossRef]
- Tierney, E.P.; Badger, J. Etiology and pathogenesis of necrolytic migratory erythema: Review of the literature. MedGenMed 2004, 6, 4. [Google Scholar] [PubMed]
- Van Beek, A.P.; de Haas, E.R.; van Vloten, W.A.; Lips, C.J.; Roijers, J.F.; Canninga-van Dijk, M.R. The glucagonoma syndrome and necrolytic migratory erythema: A clinical review. Eur. J. Endocrinol. 2004, 151, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Walker, N.P. Atypical necrolytic migratory erythema in association with a jejunal adenocarcinoma. J. R. Soc. Med. 1982, 75, 134–135. [Google Scholar] [PubMed]
- Kelly, C.P.; Johnston, C.F.; Nolan, N.; Keeling, P.W.; Weir, D.G. Necrolytic migratory erythema with elevated plasma enteroglucagon in celiac disease. Gastroenterology 1989, 96, 1350–1353. [Google Scholar] [CrossRef]
- Blackford, S.; Wright, S.; Roberts, D.L. Necrolytic migratory erythema without glucagonoma: The role of dietary essential fatty acids. Br. J. Dermatol. 1991, 125, 460–462. [Google Scholar] [CrossRef] [PubMed]
- Kerleau, J.M.; Levesque, H.; Cailleux, N.; Gancel, A.; Boullie, M.C.; Courtois, H. Isolated zinc deficiency and necrolytic migratory erythema. Apropos of a case. Rev. Med. Interne 1993, 14, 784–787. [Google Scholar] [CrossRef]
- Thorisdottir, K.; Camisa, C.; Tomecki, K.J.; Bergfeld, W.F. Necrolytic migratory erythema: A report of three cases. J. Am. Acad. Dermatol. 1994, 30, 324–329. [Google Scholar] [CrossRef]
- Marinkovich, M.P.; Botella, R.; Datloff, J.; Sangueza, O.P. Necrolytic migratory erythema without glucagonoma in patients with liver disease. J. Am. Acad. Dermatol. 1995, 32, 604–609. [Google Scholar] [CrossRef]
- Delaporte, E.; Catteau, B.; Piette, F. Necrolytic migratory erythema-like eruption in zinc deficiency associated with alcoholic liver disease. Br. J. Dermatol. 1997, 137, 1027–1028. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, S.A.; Reynolds, N.J. Necrolytic migratory erythema and zinc deficiency. Br. J. Dermatol. 1997, 136, 783–785. [Google Scholar] [CrossRef] [PubMed]
- Al-Rikabi, A.C.; Al-Homsi, H.I. Propionic acidemia and zinc deficiency presenting as necrolytic migratory erythema. Saudi Med. J. 2004, 25, 660–662. [Google Scholar] [PubMed]
- Topham, E.J.; Child, F.J. Exfoliative erythema of malnutrition with zinc and essential amino acid deficiency. Clin. Exp. Dermatol. 2005, 30, 235–237. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, H.; Komine, M.; Sasaki, K.; Mitsui, H.; Fujimoto, M.; Ihn, H.; Asahina, A.; Kikuchi, K.; Tamaki, K. Necrolytic migratory erythema without glucagonoma in a patient with short bowel syndrome. J. Dermatol. 2006, 33, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Healy, E.; Scanlain, N.I.; Barnes, L. Necrolytic migratory erythema due to zinc deficiency. Br. J. Dermatol. 1992, 127, 57–58. [Google Scholar] [CrossRef]
- Rokunohe, D.; Nakano, H.; Ikenaga, S.; Umegaki, N.; Kaneko, T.; Matsuhashi, Y.; Tando, Y.; Toyoki, Y.; Hakamada, K.; Kusumi, T.; et al. Reduction in epidermal Langerhans cells in patients with necrolytic migratory erythema. J. Dermatol. Sci. 2008, 50, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Bogan, K.L.; Brenner, C. Nicotinic acid, nicotinamide, and nicotinamide riboside: A molecular evaluation of NAD+ precursor vitamins in human nutrition. Annu. Rev. Nutr. 2008, 28, 115–130. [Google Scholar] [CrossRef] [PubMed]
- Rajakumar, K. Pellagra in the United States: A historical perspective. South. Med. J. 2000, 93, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Chick, H. The aetiology of pellagra: A review of current theories. J. Trop. Med. Hyg. 1951, 54, 207–213. [Google Scholar] [CrossRef]
- Sugita, K.; Ikenouchi-Sugita, A.; Nakayama, Y.; Yoshioka, H.; Nomura, T.; Sakabe, J.; Nakahigashi, K.; Kuroda, E.; Uematsu, S.; Nakamura, J.; et al. Prostaglandin E2 is critical for the development of niacin-deficiency-induced photosensitivity via ROS production. Sci. Rep. 2013, 3, 2973. [Google Scholar] [CrossRef] [PubMed]
- Vannucchi, H.; Favaro, R.M.; Cunha, D.F.; Marchini, J.S. Assessment of zinc nutritional status of pellagra patients. Alcohol Alcohol. 1995, 30, 297–302. [Google Scholar] [PubMed]
- Yamaguchi, S.; Miyagi, T.; Sogabe, Y.; Yasuda, M.; Kanazawa, N.; Utani, A.; Izaki, S.; Uezato, H.; Takahashi, K. Depletion of Epidermal Langerhans Cells in the Skin Lesions of Pellagra Patients. Am. J. Dermatopathol. 2017, 39, 428–432. [Google Scholar] [CrossRef] [PubMed]
- Zempleni, J.; Wijeratne, S.S.; Hassan, Y.I. Biotin. Biofactors 2009, 35, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Mock, D.M. Skin manifestations of biotin deficiency. Semin. Dermatol. 1991, 10, 296–302. [Google Scholar] [PubMed]
- Mock, D.M.; Johnson, S.B.; Holman, R.T. Effects of biotin deficiency on serum fatty acid composition: Evidence for abnormalities in humans. J. Nutr. 1988, 118, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Mock, D.M.; Mock, N.I.; Johnson, S.B.; Holman, R.T. Effects of biotin deficiency on plasma and tissue fatty acid composition: Evidence for abnormalities in rats. Pediatr. Res. 1988, 24, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Kramer, T.R.; Briske-Anderson, M.; Johnson, S.B.; Holman, R.T. Effects of biotin deficiency on polyunsaturated fatty acid metabolism in rats. J. Nutr. 1984, 114, 2047–2052. [Google Scholar] [CrossRef] [PubMed]
- Suchy, S.F.; Rizzo, W.B.; Wolf, B. Effect of biotin deficiency and supplementation on lipid metabolism in rats: Saturated fatty acids. Am. J. Clin. Nutr. 1986, 44, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Matsusue, S.; Kashihara, S.; Takeda, H.; Koizumi, S. Biotin deficiency during total parenteral nutrition: Its clinical manifestation and plasma nonesterified fatty acid level. JPEN J. Parenter. Enter. Nutr. 1985, 9, 760–763. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, R.; Mizukoshi, M.; Koyama, H.; Kitano, N.; Koike, M. Intractable diaper dermatitis as an early sign of biotin deficiency. Acta Paediatr. 1998, 87, 228–229. [Google Scholar] [CrossRef] [PubMed]
- Lagier, P.; Bimar, P.; Seriat-Gautier, S.; Dejode, J.M.; Brun, T.; Bimar, J. Zinc and biotin deficiency during prolonged parenteral nutrition in the infant. Presse Med. 1987, 16, 1795–1797. [Google Scholar] [PubMed]
- Khalidi, N.; Wesley, J.R.; Thoene, J.G.; Whitehouse, W.M., Jr.; Baker, W.L. Biotin deficiency in a patient with short bowel syndrome during home parenteral nutrition. JPEN J. Parenter. Enter. Nutr. 1984, 8, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, R.; Noda, E.; Koyama, Y.; Shirai, T.; Horino, A.; Juri, T.; Koike, M. Biotin deficiency in an infant fed with amino acid formula and hypoallergenic rice. Acta Paediatr. 1996, 85, 872–874. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, W.; Inaoki, M.; Fukui, T.; Inoue, Y.; Kuhara, T. Biotin deficiency in an infant fed with amino acid formula. J. Dermatol. 2005, 32, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, J.R.; Kossard, S. Acquired scalp alopecia. Part I: A review. Australas. J. Dermatol. 1998, 39, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Finner, A.M. Nutrition and hair: Deficiencies and supplements. Dermatol. Clin. 2013, 31, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Karashima, T.; Tsuruta, D.; Hamada, T.; Ono, F.; Ishii, N.; Abe, T.; Ohyama, B.; Nakama, T.; Dainichi, T.; Hashimoto, T. Oral zinc therapy for zinc deficiency-related telogen effluvium. Dermatol. Ther. 2012, 25, 210–213. [Google Scholar] [CrossRef] [PubMed]
- Kil, M.S.; Kim, C.W.; Kim, S.S. Analysis of serum zinc and copper concentrations in hair loss. Ann. Dermatol. 2013, 25, 405–409. [Google Scholar] [CrossRef] [PubMed]
- Amador, M.; Pena, M.; Garcia-Miranda, A.; Gonzalez, A.; Hermelo, M. Letter: Low hair-zinc concentrations in acrodermatitis enteropathica. Lancet 1975, 1, 1379. [Google Scholar] [CrossRef]
- Traupe, H.; Happle, R.; Grobe, H.; Bertram, H.P. Polarization microscopy of hair in acrodermatitis enteropathica. Pediatr. Dermatol. 1986, 3, 300–303. [Google Scholar] [CrossRef] [PubMed]
- Dupre, A.; Bonafe, J.L.; Carriere, J.P. The hair in acrodermatitis interopathica—A disease indicator? Acta Derm. Venereol. 1979, 59, 177–178. [Google Scholar] [PubMed]
- Follis, R.H.; Day, H.G.; McCollum, E.V. Histological studies of the tissues of rats fed a diet extremely low in zinc. J. Nutr. 1941, 22, 223–237. [Google Scholar] [CrossRef]
- Hsu, J.N. Zinc as Related to Cystine Metabolism; Academic Press: London, UK, 1976. [Google Scholar]
- Wilson, R.H.; Lewis, H.B. The cystine content of hair and other epidermal tissues. J. Biol. Chem. 1927, 73, 543–553. [Google Scholar]
- Day, H.G.; Skidmore, B.E. Some effects of dietary zinc deficiency in the mouse. J. Nutr. 1946, 33, 27–38. [Google Scholar] [CrossRef]
- Swenerton, H.; Hurley, L.S. Severe zinc deficiency in male and female rats. J. Nutr. 1968, 95, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Todd, W.R.; Elvehjem, C.A.; Hart, E.B. Zinc in the nutrition of the rat. Am. J. Physiol. 1933, 107, 146–156. [Google Scholar] [CrossRef]
- Shaw, N.A.; Dickey, H.C.; Brugman, H.H.; Blamberg, D.L.; Witter, J.F. Zinc deficiency in female rabbits. Lab. Anim. 1974, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Dai, Z.; Jabbari, A.; Cerise, J.E.; Higgins, C.A.; Gong, W.; de Jong, A.; Harel, S.; DeStefano, G.M.; Rothman, L.; et al. Alopecia areata is driven by cytotoxic T lymphocytes and is reversed by JAK inhibition. Nat. Med. 2014, 20, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Petukhova, L.; Duvic, M.; Hordinsky, M.; Norris, D.; Price, V.; Shimomura, Y.; Kim, H.; Singh, P.; Lee, A.; Chen, W.V.; et al. Genome-wide association study in alopecia areata implicates both innate and adaptive immunity. Nature 2010, 466, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Freyschmidt-Paul, P.; McElwee, K.J.; Hoffmann, R.; Sundberg, J.P.; Vitacolonna, M.; Kissling, S.; Zoller, M. Interferon-gamma-deficient mice are resistant to the development of alopecia areata. Br. J. Dermatol. 2006, 155, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Kim, C.W.; Kim, S.S.; Park, C.W. The therapeutic effect and the changed serum zinc level after zinc supplementation in alopecia areata patients who had a low serum zinc level. Ann. Dermatol. 2009, 21, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Bhat, Y.J.; Manzoor, S.; Khan, A.R.; Qayoom, S. Trace element levels in alopecia areata. Indian J. Dermatol. Venereol. Leprol. 2009, 75, 29–31. [Google Scholar] [CrossRef] [PubMed]
- Abdel Fattah, N.S.; Atef, M.M.; Al-Qaradaghi, S.M. Evaluation of serum zinc level in patients with newly diagnosed and resistant alopecia areata. Int. J. Dermatol. 2016, 55, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Lansdown, A.B.; Mirastschijski, U.; Stubbs, N.; Scanlon, E.; Agren, M.S. Zinc in wound healing: Theoretical, experimental, and clinical aspects. Wound Repair Regen. 2007, 15, 2–16. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, J.R.; Marsh, R.G.; Draelos, Z.D. Zinc and skin health: Overview of physiology and pharmacology. Dermatol. Surg. 2005, 31, 837–847. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, E.A.; Hawke, C.I. Does oral zinc aid the healing of chronic leg ulcers? A systematic literature review. Arch. Dermatol. 1998, 134, 1556–1560. [Google Scholar] [CrossRef] [PubMed]
- Agren, M.S.; Stromberg, H.E. Topical treatment of pressure ulcers. A randomized comparative trial of Varidase and zinc oxide. Scand. J. Plast. Reconstr. Surg. 1985, 19, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Agren, M.S. Studies on zinc in wound healing. Acta Derm. Venereol. Suppl. 1990, 154, 1–36. [Google Scholar]
- Popovics, P.; Stewart, A.J. GPR39, a Zn2+-activated G protein-coupled receptor that regulates pancreatic, gastrointestinal and neuronal functions. Cell. Mol. Life Sci. 2011, 68, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Sharir, H.; Zinger, A.; Nevo, A.; Sekler, I.; Hershfinkel, M. Zinc released from injured cells is acting via the Zn2+-sensing receptor, ZnR, to trigger signaling leading to epithelial repair. J. Biol. Chem. 2010, 285, 26097–26106. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, K.A.U.; Fuchs, E. Skin and its regenerative powers: An alliance between stem cells and their niche. Dev. Cell 2017, 43, 387–401. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Qiao, J.; Zhang, S.; Zhang, H.; Lei, X.; Wang, X.; Deng, Z.; Ning, L.; Cao, Y.; Guo, Y.; et al. GPR39 marks specific cells within the sebaceous gland and contributes to skin wound healing. Sci. Rep. 2015, 5, 7913. [Google Scholar] [CrossRef] [PubMed]
- David, T.J.; Wells, F.E.; Sharpe, T.C.; Gibbs, A.C. Low serum zinc in children with atopic eczema. Br. J. Dermatol. 1984, 111, 597–601. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Yoo, S.R.; Jeong, M.G.; Ko, J.Y.; Ro, Y.S. Hair zinc levels and the efficacy of oral zinc supplementation in patients with atopic dermatitis. Acta Derm. Venereol. 2014, 94, 558–562. [Google Scholar] [CrossRef] [PubMed]
- Gholizadeh, N.; Mehdipour, M.; Najafi, S.; Bahramian, A.; Garjani, S.; Khoeini Poorfar, H. Evaluation of the serum zinc level in erosive and non-erosive oral lichen planus. J. Dent. 2014, 15, 52–56. [Google Scholar]
- Dogan, P.; Dogan, M.; Klockenkamper, R. Determination of trace elements in blood serum of patients with Behcet disease by total reflection X-ray fluorescence analysis. Clin. Chem. 1993, 39, 1037–1041. [Google Scholar] [PubMed]
- Saglam, K.; Serce, A.F.; Yilmaz, M.I.; Bulucu, F.; Aydin, A.; Akay, C.; Sayal, A. Trace elements and antioxidant enzymes in Behcet’s disease. Rheumatol. Int. 2002, 22, 93–96. [Google Scholar] [PubMed]
- Yazdanpanah, M.J.; Ghayour-Mobarhan, M.; Taji, A.; Javidi, Z.; Pezeshkpoor, F.; Tavallaie, S.; Momenzadeh, A.; Esmaili, H.; Shojaie-Noori, S.; Khoddami, M.; et al. Serum zinc and copper status in Iranian patients with pemphigus vulgaris. Int. J. Dermatol. 2011, 50, 1343–1346. [Google Scholar] [CrossRef] [PubMed]
- Tasaki, M.; Hanada, K.; Hashimoto, I. Analyses of serum copper and zinc levels and copper/zinc ratios in skin diseases. J. Dermatol. 1993, 20, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Ingen-Housz-Oro, S.; Blanchet-Bardon, C.; Vrillat, M.; Dubertret, L. Vitamin and trace metal levels in recessive dystrophic epidermolysis bullosa. J. Eur. Acad. Dermatol. Venereol. 2004, 18, 649–653. [Google Scholar] [CrossRef] [PubMed]
- Fine, J.D.; Tamura, T.; Johnson, L. Blood vitamin and trace metal levels in epidermolysis bullosa. Arch. Dermatol. 1989, 125, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Rostami, M.M.; Safavi, A.N.; Iranparvar, A.M.; Maleki, N.; Aghabalaei, D.M. Evaluation of the serum zinc level in adult patients with melasma: Is there a relationship with serum zinc deficiency and melasma? J. Cosmet. Dermatol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Fraker, P.J.; King, L.E. Reprogramming of the immune system during zinc deficiency. Annu. Rev. Nutr. 2004, 24, 277–298. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Reported distribution and function of Zn transporters in the skin. Every skin cell expresses many different Zn transporters. However, the expression and role of Zn transporters in skin cells are less understood. This figure shows the reported distribution and representative function of Zn transporters in the skin. 1 and 2. ZIP2 and ZIP4 in KCs facilitate KC proliferation and differentiation. 3. ZIP10 expressed in the epidermal progenitor cells in outer root sheath is crucial for the proper epidermal formation. 4. ZnT5 in MCs is involved in inflammatory cytokine production. 5. Many ZnTs and ZIPs regulate MHC class II expression in DCs. 6. ZIP8 in T cells is involved in interferon-γ (IFN-γ) production. 7. ZIP7 and ZIP13 in fibroblasts are required for the dermal formation and bone morphogenetic protein/transforming growth factor-β (BMP/TGF-β) signaling, respectively. 8. ZIP13 in adipocytes inhibits beige fat cell differentiation. ZIP14 in adipocytes suppresses an excess inflammation.
Figure 1.
Reported distribution and function of Zn transporters in the skin. Every skin cell expresses many different Zn transporters. However, the expression and role of Zn transporters in skin cells are less understood. This figure shows the reported distribution and representative function of Zn transporters in the skin. 1 and 2. ZIP2 and ZIP4 in KCs facilitate KC proliferation and differentiation. 3. ZIP10 expressed in the epidermal progenitor cells in outer root sheath is crucial for the proper epidermal formation. 4. ZnT5 in MCs is involved in inflammatory cytokine production. 5. Many ZnTs and ZIPs regulate MHC class II expression in DCs. 6. ZIP8 in T cells is involved in interferon-γ (IFN-γ) production. 7. ZIP7 and ZIP13 in fibroblasts are required for the dermal formation and bone morphogenetic protein/transforming growth factor-β (BMP/TGF-β) signaling, respectively. 8. ZIP13 in adipocytes inhibits beige fat cell differentiation. ZIP14 in adipocytes suppresses an excess inflammation.

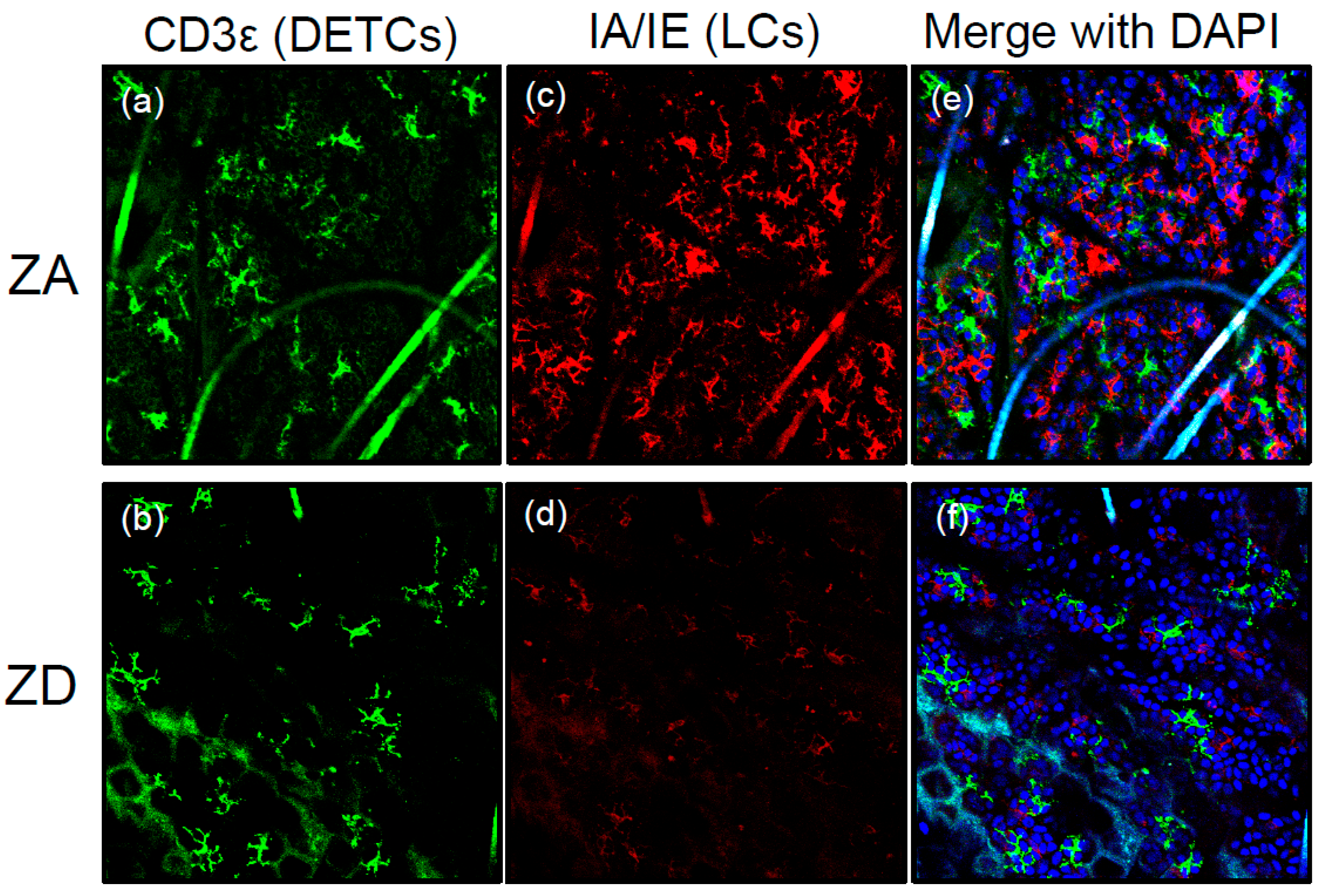
Figure 2.
LC loss in ZD ear epidermis. Immunofluorescence of epidermal whole mounts stained for IA/IE (red) and CD3ε (green) from mice fed ZA or ZD diets for seven weeks. Original magnification, ×100. IA/IE-positive LCs are absent in ZD ear epidermis (c) and (d); whereas the distribution of CD3ε-positive dendritic epidermal T cells are unaffected (a) and (b). (a) and (c) merge with DAPI is (e); (b) and (d) merge with DAPI is (f).
Figure 2.
LC loss in ZD ear epidermis. Immunofluorescence of epidermal whole mounts stained for IA/IE (red) and CD3ε (green) from mice fed ZA or ZD diets for seven weeks. Original magnification, ×100. IA/IE-positive LCs are absent in ZD ear epidermis (c) and (d); whereas the distribution of CD3ε-positive dendritic epidermal T cells are unaffected (a) and (b). (a) and (c) merge with DAPI is (e); (b) and (d) merge with DAPI is (f).

Figure 3.
An underlying mechanism of development of acrodermatitis. Right panel: 1. In patients with AE and ZD mice, TGF-β1 expression in the epidermis is impaired compared with healthy subjects and ZA mice. Subsequently, LCs disappear from the epidermis. LCs are the sole CD39 (ecto-NTPDase1)-expressing cells in the epidermis. 2. Ear epidermis from ZD mice produces much more ATP upon the exposure of irritants than it from ZA mice. Additionally, this ATP is not hydrolyzed because of the absence of CD39-expressing LCs. 3. ATP in the epidermis elicits ICD, followed by the formation of acrodermatitis in AE.
Figure 3.
An underlying mechanism of development of acrodermatitis. Right panel: 1. In patients with AE and ZD mice, TGF-β1 expression in the epidermis is impaired compared with healthy subjects and ZA mice. Subsequently, LCs disappear from the epidermis. LCs are the sole CD39 (ecto-NTPDase1)-expressing cells in the epidermis. 2. Ear epidermis from ZD mice produces much more ATP upon the exposure of irritants than it from ZA mice. Additionally, this ATP is not hydrolyzed because of the absence of CD39-expressing LCs. 3. ATP in the epidermis elicits ICD, followed by the formation of acrodermatitis in AE.

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ogawa, Y.; Kinoshita, M.; Shimada, S.; Kawamura, T. Zinc and Skin Disorders. Nutrients 2018, 10, 199. https://doi.org/10.3390/nu10020199
AMA Style
Ogawa Y, Kinoshita M, Shimada S, Kawamura T. Zinc and Skin Disorders. Nutrients. 2018; 10(2):199. https://doi.org/10.3390/nu10020199
Chicago/Turabian StyleOgawa, Youichi, Manao Kinoshita, Shinji Shimada, and Tatsuyoshi Kawamura. 2018. "Zinc and Skin Disorders" Nutrients 10, no. 2: 199. https://doi.org/10.3390/nu10020199
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.