Choline Content of Term and Preterm Infant Formulae Compared to Expressed Breast Milk—How Do We Justify the Discrepancies?

 , , and
, , and 
Abstract
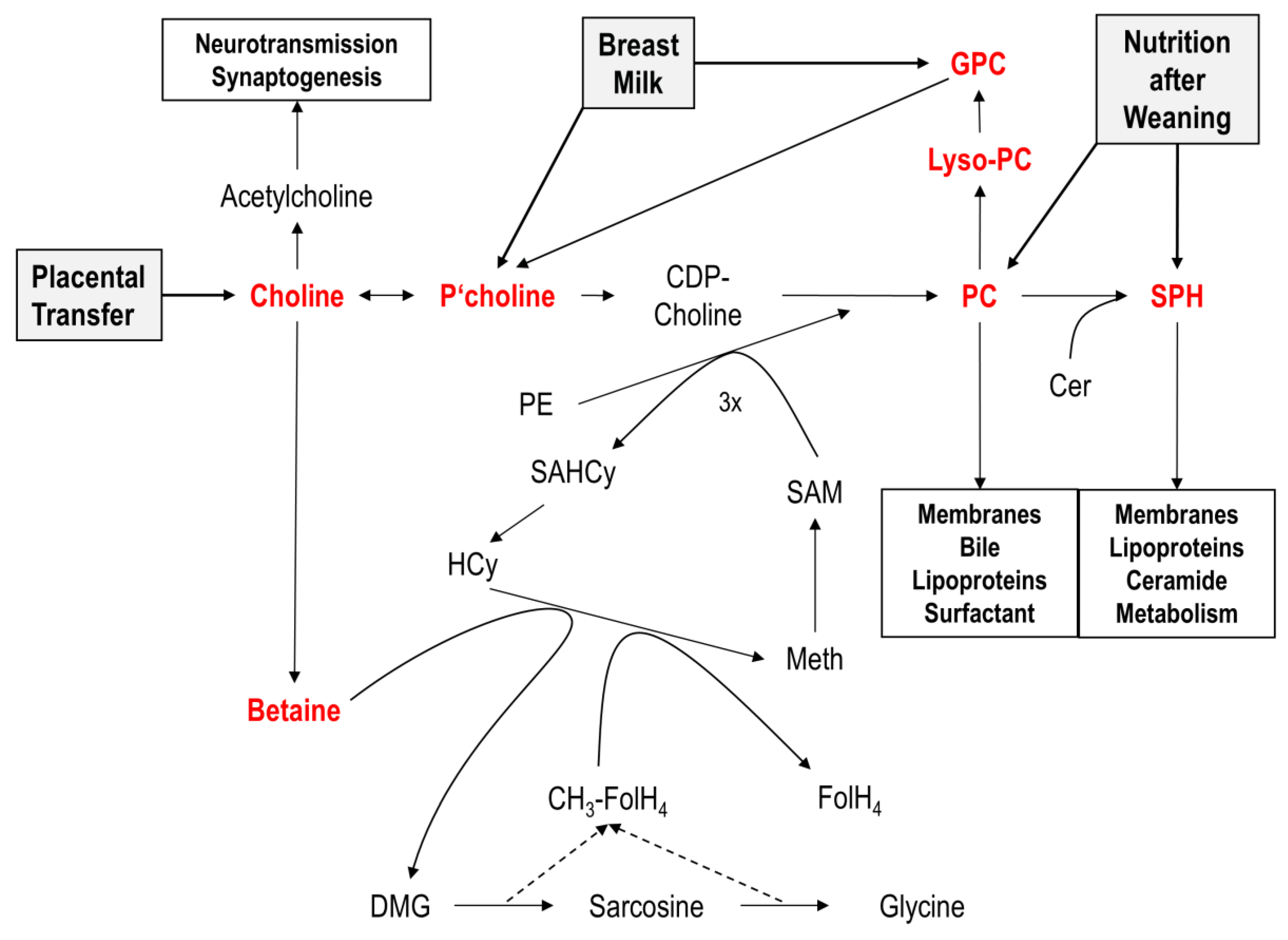
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Sample Acquisition
2.3. Sample Extraction
2.4. Analysis of Choline, Water Soluble Choline Metabolites and Phospholipids
2.5. Statistics
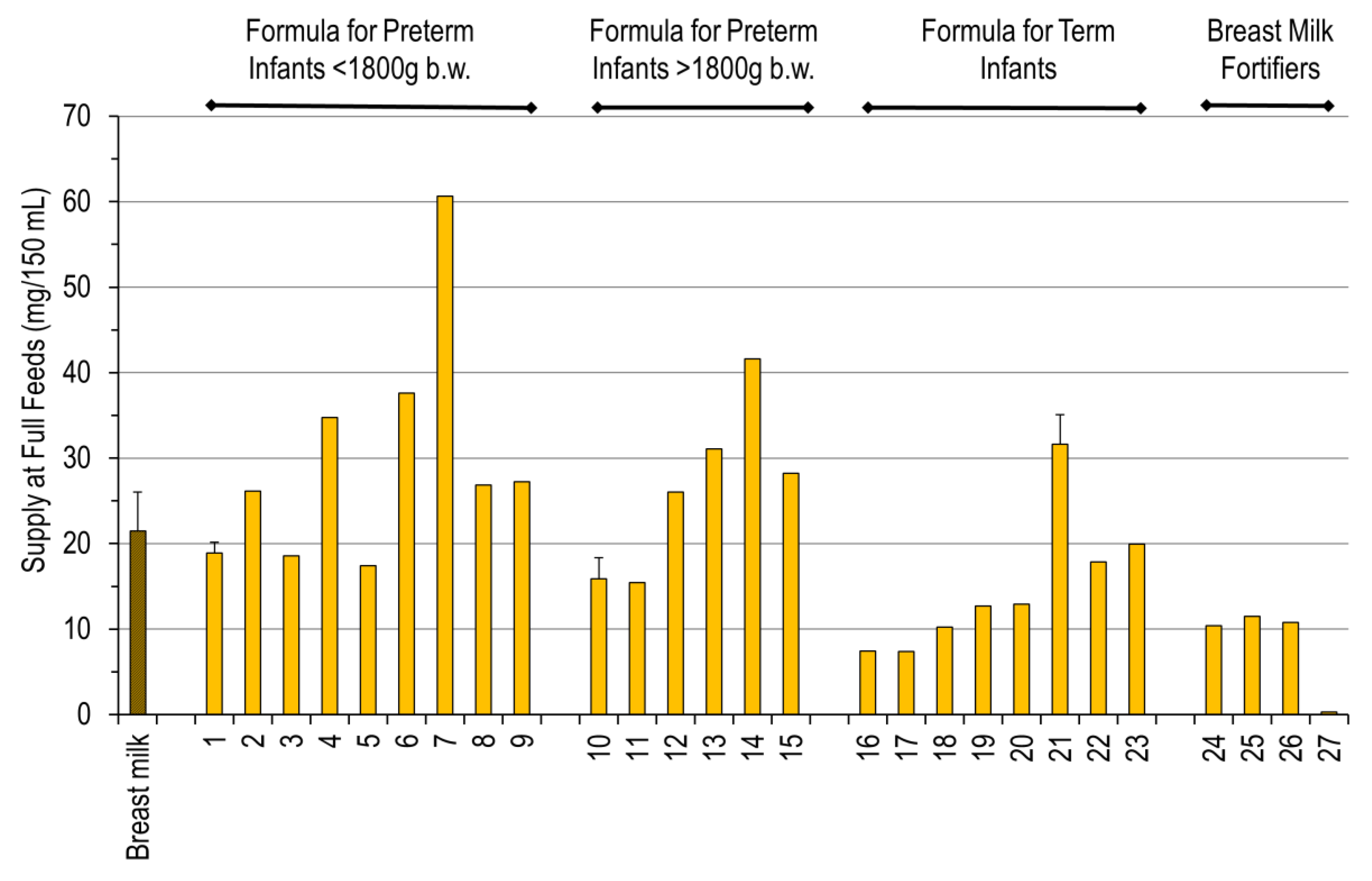
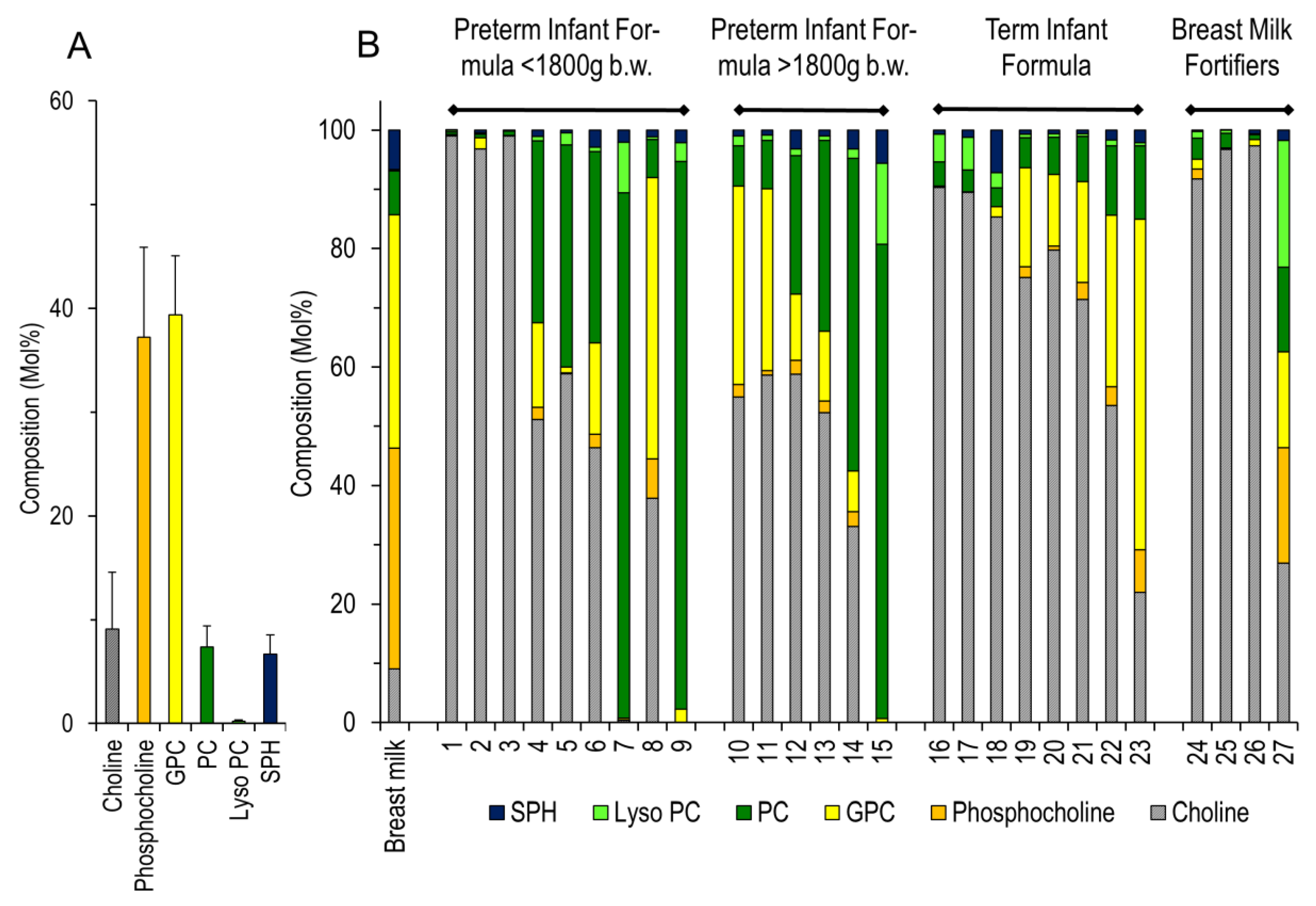
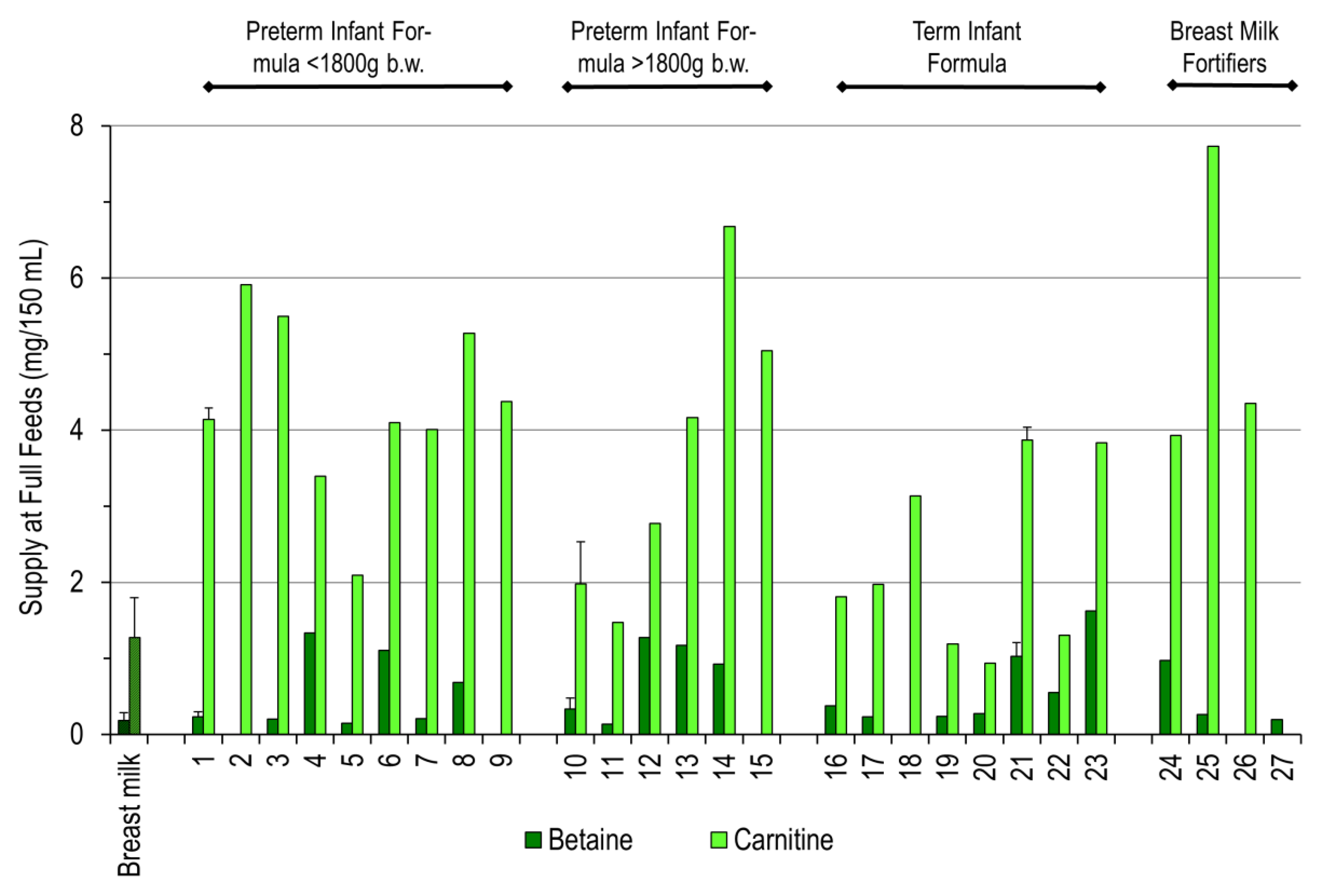
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hollenbeck, C.B. An introduction to the nutrition and metabolism of choline. Cent. Nerv. Syst. Agents Med. Chem. 2012, 12, 100–113. [Google Scholar] [CrossRef] [PubMed]
- Bernhard, W.; Poets, C.F.; Franz, A.R. Choline and choline-related nutrients in regular and preterm infant growth. Eur. J. Nutr. 2019, 58, 931–945. [Google Scholar] [CrossRef] [PubMed]
- Cohen, D.E. Hepatocellular transport and secretion of biliary phospholipids. Semin. Liver Dis. 1996, 16, 191–200. [Google Scholar] [CrossRef]
- Weaver, T.E.; Na, C.L.; Stahlman, M. Biogenesis of lamellar bodies, lysosome-related organelles involved in storage and secretion of pulmonary surfactant. Semin. Cell Dev. Biol. 2002, 13, 263–270. [Google Scholar] [CrossRef]
- Shelness, G.S.; Sellers, J.A. Very-low-density lipoprotein assembly and secretion. Curr. Opin. Lipidol. 2001, 12, 151–157. [Google Scholar] [CrossRef]
- Fujii, T.; Mashimo, M.; Moriwaki, Y.; Misawa, H.; Ono, S.; Horiguchi, K.; Kawashima, K. Expression and function of the Cholinergic system in immune cells. Front. Immunol. 2017, 8, 1085. [Google Scholar] [CrossRef] [Green Version]
- Innis, S.M.; Davidson, A.G.; Bay, B.N.; Slack, P.J.; Hasman, D. Plasma choline depletion is associated with decreased peripheral blood leukocyte acetylcholine in children with cystic fibrosis. Am. J. Clin. Nutr. 2011, 93, 564–568. [Google Scholar] [CrossRef] [Green Version]
- Zeisel, S.H. Choline: Critical role during fetal development and dietary requirements in adults. Annu. Rev. Nutr. 2006, 26, 229–250. [Google Scholar] [CrossRef] [Green Version]
- Oshida, K.; Shimizu, T.; Takase, M.; Tamura, Y.; Yamashiro, Y. Effects of dietary sphingomyelin on central nervous system myelination in developing rats. Pediatric Res. 2003, 53, 589–593. [Google Scholar] [CrossRef] [Green Version]
- Gault, C.R.; Obeid, L.M.; Hannun, Y.A. An overview of sphingolipid metabolism: From synthesis to breakdown. Adv. Exp. Med. Biol. 2010, 688, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Nojima, H.; Freeman, C.M.; Gulbins, E.; Lentsch, A.B. Sphingolipids in liver injury, repair and regeneration. Biol. Chem. 2015, 396, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Hendricks-Munoz, K.D.; Xu, J.; Voynow, J.A. Tracheal aspirate VEGF and sphingolipid metabolites in the preterm infant with later development of bronchopulmonary dysplasia. Pediatric Pulmonol. 2018, 53, 1046–1052. [Google Scholar] [CrossRef] [PubMed]
- Bernhard, W.; Full, A.; Arand, J.; Maas, C.; Poets, C.F.; Franz, A.R. Choline supply of preterm infants: Assessment of dietary intake and pathophysiological considerations. Eur. J. Nutr. 2013, 52, 1269–1278. [Google Scholar] [CrossRef] [PubMed]
- van Mastrigt, E.; Zweekhorst, S.; Bol, B.; Tibboel, J.; van Rosmalen, J.; Samsom, J.N.; Kroon, A.A.; de Jongste, J.C.; Reiss, I.K.M.; Post, M.; et al. Ceramides in tracheal aspirates of preterm infants: Marker for bronchopulmonary dysplasia. PLoS ONE 2018, 13, e0185969. [Google Scholar] [CrossRef] [PubMed]
- Mudd, S.H.; Brosnan, J.T.; Brosnan, M.E.; Jacobs, R.L.; Stabler, S.P.; Allen, R.H.; Vance, D.E.; Wagner, C. Methyl balance and transmethylation fluxes in humans. Am. J. Clin. Nutr. 2007, 85, 19–25. [Google Scholar] [CrossRef]
- Yan, J.; Jiang, X.; West, A.A.; Perry, C.A.; Malysheva, O.V.; Devapatla, S.; Pressman, E.; Vermeylen, F.; Stabler, S.P.; Allen, R.H.; et al. Maternal choline intake modulates maternal and fetal biomarkers of choline metabolism in humans. Am. J. Clin. Nutr. 2012, 95, 1060–1071. [Google Scholar] [CrossRef] [Green Version]
- Wyss, M.; Kaddurah-Daouk, R. Creatine and creatinine metabolism. Physiol. Rev. 2000, 80, 1107–1213. [Google Scholar] [CrossRef]
- Zeisel, S. Choline, other Methyl-donors and epigenetics. Nutrients 2017, 9, 445. [Google Scholar] [CrossRef]
- Wiedeman, A.M.; Barr, S.I.; Green, T.J.; Xu, Z.; Innis, S.M.; Kitts, D.D. Dietary choline intake: Current state of knowledge across the life cycle. Nutrients 2018, 10, 1513. [Google Scholar] [CrossRef] [Green Version]
- Bernhard, W.; Lange, R.; Graepler-Mainka, U.; Engel, C.; Machann, J.; Hund, V.; Shunova, A.; Hector, A.; Riethmuller, J. Choline supplementation in cystic fibrosis—The metabolic and clinical impact. Nutrients 2019, 11, 656. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Agellon, L.B.; Vance, D.E. Choline redistribution during adaptation to choline deprivation. J. Biol. Chem. 2007, 282, 10283–10289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernhard, W.; Bockmann, K.; Maas, C.; Mathes, M.; Hovelmann, J.; Shunova, A.; Hund, V.; Schleicher, E.; Poets, C.F.; Franz, A.R. Combined choline and DHA supplementation: A randomized controlled trial. Eur. J. Nutr. 2020, 59, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Goss, K.C.W.; Goss, V.M.; Townsend, J.P.; Koster, G.; Clark, H.W.; Postle, A.D. Postnatal adaptations of phosphatidylcholine metabolism in extremely preterm infants: Implications for choline and PUFA metabolism. Am. J. Clin. Nutr. 2020, 112, 1438–1447. [Google Scholar] [CrossRef] [PubMed]
- Institute of Medicine. Dietary Reference Intakes for Thiamin, Riboflavin, Niacin, Vitamin B6, Folate, Vitamin B12, Pantothenic Acid, Biotin, and Choline; The National Academic Press: Washington, DC, USA, 1998. [Google Scholar]
- EFSA. Panel on dietetic products, N.a.A.N. Dietary reference values for choline. EFSA J. 2016, 14, 4484. [Google Scholar] [CrossRef]
- Bernhard, W.; Raith, M.; Kunze, R.; Koch, V.; Heni, M.; Maas, C.; Abele, H.; Poets, C.F.; Franz, A.R. Choline concentrations are lower in postnatal plasma of preterm infants than in cord plasma. Eur. J. Nutr. 2015, 54, 733–741. [Google Scholar] [CrossRef]
- Caudill, M.A. Pre- and postnatal health: Evidence of increased choline needs. J. Am. Diet. Assoc. 2010, 110, 1198–1206. [Google Scholar] [CrossRef]
- Agostoni, C.; Buonocore, G.; Carnielli, V.P.; De Curtis, M.; Darmaun, D.; Decsi, T.; Domellof, M.; Embleton, N.D.; Fusch, C.; Genzel-Boroviczeny, O.; et al. Enteral nutrient supply for preterm infants: Commentary from the European Society of paediatric gastroenterology, hepatology and nutrition committee on nutrition. J. Pediatric Gastroenterol. Nutr. 2010, 50, 85–91. [Google Scholar] [CrossRef]
- Maas, C.; Franz, A.R.; Shunova, A.; Mathes, M.; Bleeker, C.; Poets, C.F.; Schleicher, E.; Bernhard, W. Choline and polyunsaturated fatty acids in preterm infants’ maternal milk. Eur. J. Nutr. 2017, 56, 1733–1742. [Google Scholar] [CrossRef]
- Patterson, K.Y.; Bhagwat, S.A.; Williams, J.R.; Howe, J.C.; Holden, J.M. USDA Database for the Choline Content of Common Foods—Release Two; Agricultural Research Service: Washington, DC, USA, 2008. [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Carlson, S.E.; Colombo, J. Docosahexaenoic acid and arachidonic acid nutrition in early development. Adv. Pediatrics 2016, 63, 453–471. [Google Scholar] [CrossRef] [Green Version]
- Hirabayashi, Y.; Furuya, S. Roles of l-serine and sphingolipid synthesis in brain development and neuronal survival. Prog. Lipid Res. 2008, 47, 188–203. [Google Scholar] [CrossRef] [PubMed]
- Grieb, P. Neuroprotective properties of citicoline: Facts, doubts and unresolved issues. CNS Drugs 2014, 28, 185–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, K.Y.; Wu, T.C.; Huang, C.F.; Lin, C.C.; Huang, I.F.; Wu, L. Profile of nucleotides and nucleosides in Taiwanese human milk. Pediatrics Neonatol. 2011, 52, 93–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leucht, K.; Fischbeck, A.; Caj, M.; Liebisch, G.; Hartlieb, E.; Benes, P.; Fried, M.; Humpf, H.U.; Rogler, G.; Hausmann, M. Sphingomyelin and phosphatidylcholine contrarily affect the induction of apoptosis in intestinal epithelial cells. Mol. Nutr. Food Res. 2014, 58, 782–798. [Google Scholar] [CrossRef]
- Hernell, O.; Timby, N.; Domellof, M.; Lonnerdal, B. Clinical benefits of milk fat globule membranes for infants and children. J. Pediatrics 2016, 173, S60–S65. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.; Holmes-McNary, M.; Mar, M.H.; Lien, E.; Zeisel, S. Bioavailability of choline and choline esters from milk in rat pups. J. Nutr. Biochem. 1996, 7, 457–464. [Google Scholar] [CrossRef]
- al-Waiz, M.; Mikov, M.; Mitchell, S.C.; Smith, R.L. The exogenous origin of trimethylamine in the mouse. Metab. Clin. Exp. 1992, 41, 135–136. [Google Scholar] [CrossRef]
- Cho, C.E.; Aardema, N.D.J.; Bunnell, M.L.; Larson, D.P.; Aguilar, S.S.; Bergeson, J.R.; Malysheva, O.V.; Caudill, M.A.; Lefevre, M. Effect of choline forms and gut microbiota composition on Trimethylamine-N-oxide response in healthy men. Nutrients 2020, 12, 2220. [Google Scholar] [CrossRef]
- Bernhard, W. Choline in cystic fibrosis: Relations to pancreas insufficiency, enterohepatic cycle, PEMT and intestinal microbiota. Eur. J. Nutr. 2020. Online ahead of print. [Google Scholar] [CrossRef]
- Nilsson, A.; Duan, R.D. Pancreatic and mucosal enzymes in choline phospholipid digestion. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 316, G425–G445. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Full Formula for Preterm Infants below 1800 g Birth Weight | |
| 1 | BEBA Frühgeborenennahrung Stufe 1-2011–2012 | Ready to use |
| 2 | BEBA Frühgeborenennahrung Stufe 1-2020 | Ready to use |
| 3 | BEBA Frühgeborenennahrung Stufe 1 PRO HA-2015 | Ready to use |
| 4 | Aptamil PREMATIL-2020 | Ready to use |
| 5 | Aptamil PREMATIL HA-2020 | Ready to use |
| 6 | Aptamil Prematil HA-2015 | Ready to use |
| 7 | Humana 0-VLB-2015 | Ready to use |
| 8 | Humana 0 medical-2015 | Ready to use |
| 9 | Humana 0-VLB-2020 | Ready to use |
| Full Formula for Preterm Infants above 1800 g Birth Weight | ||
| 10 | BEBA Frühgeborenennahrung Stufe 2-2011–2012 | Ready to use |
| 11 | BEBA Frühgeborenennahrung Stufe 2 PRO HA-2015 | Ready to use |
| 12 | Aptamil PDF-2016 | Ready to use |
| 13 | Aptamil PDF-2020 | Ready to use |
| 14 | Humana 0-2020 | Ready to use |
| 15 | Humana 0-HA-2020 | Ready to use |
| Full Formula for Term Infants above 2500 g Birth Weight | ||
| 16 | BEBA HA Start PRE-2011 | Ready to use |
| 17 | BEBA HA PRE PRO HA-2015 | Ready to use |
| 18 | BEBA SUPREME PRE-2020 | Ready to use |
| 19 | Aptamil PRE-2011 | Ready to use |
| 20 | Aptamil PRE-2015 | Ready to use |
| 21 | Aptamil PRE-2020 | Ready to use |
| 22 | Aptamil GOS/FOS HA Pre-2015 | Ready to use |
| 23 | Humana Anfangsmilch PRE-2019 | Ready to use |
| Breast Milk Fortifiers for Preterm Infants | ||
| 24 | BEBA FM 85-2016 | 5.0% dilution in water |
| 25 | Nestlé Study B.M.S. | 5.0% dilution in water |
| 26 | BEBA FM 85-2019 | 4.0% dilution in water |
| 27 | Aptamil FMS-2016 | 4.4% dilution in water |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shunova, A.; Böckmann, K.A.; Minarski, M.; Franz, A.R.; Wiechers, C.; Poets, C.F.; Bernhard, W. Choline Content of Term and Preterm Infant Formulae Compared to Expressed Breast Milk—How Do We Justify the Discrepancies? Nutrients 2020, 12, 3815. https://doi.org/10.3390/nu12123815
Shunova A, Böckmann KA, Minarski M, Franz AR, Wiechers C, Poets CF, Bernhard W. Choline Content of Term and Preterm Infant Formulae Compared to Expressed Breast Milk—How Do We Justify the Discrepancies? Nutrients. 2020; 12(12):3815. https://doi.org/10.3390/nu12123815
Chicago/Turabian StyleShunova, Anna, Katrin A. Böckmann, Michaela Minarski, Axel R. Franz, Cornelia Wiechers, Christian F. Poets, and Wolfgang Bernhard. 2020. "Choline Content of Term and Preterm Infant Formulae Compared to Expressed Breast Milk—How Do We Justify the Discrepancies?" Nutrients 12, no. 12: 3815. https://doi.org/10.3390/nu12123815