
Ovatodiolide Inhibits Breast Cancer Stem/Progenitor Cells through SMURF2-Mediated Downregulation of Hsp27

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
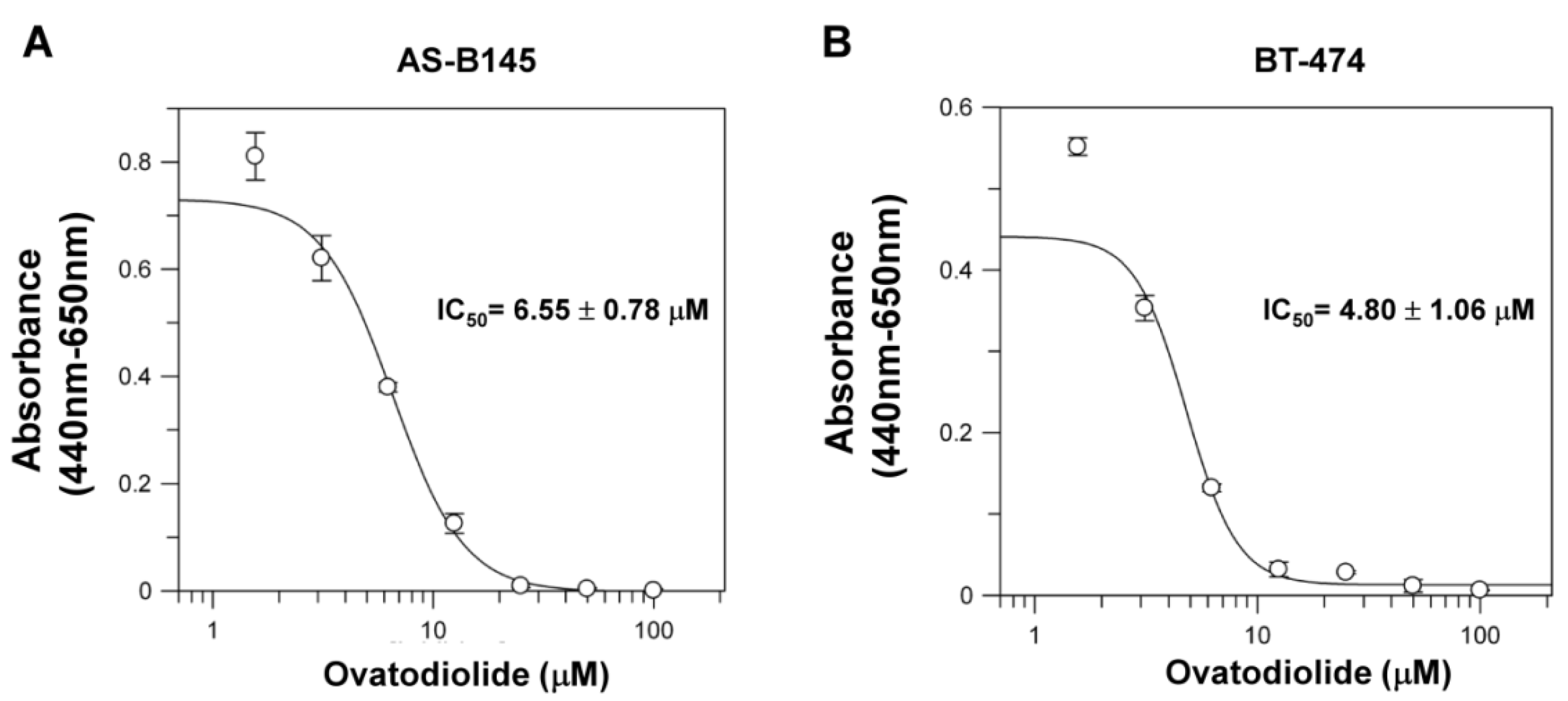
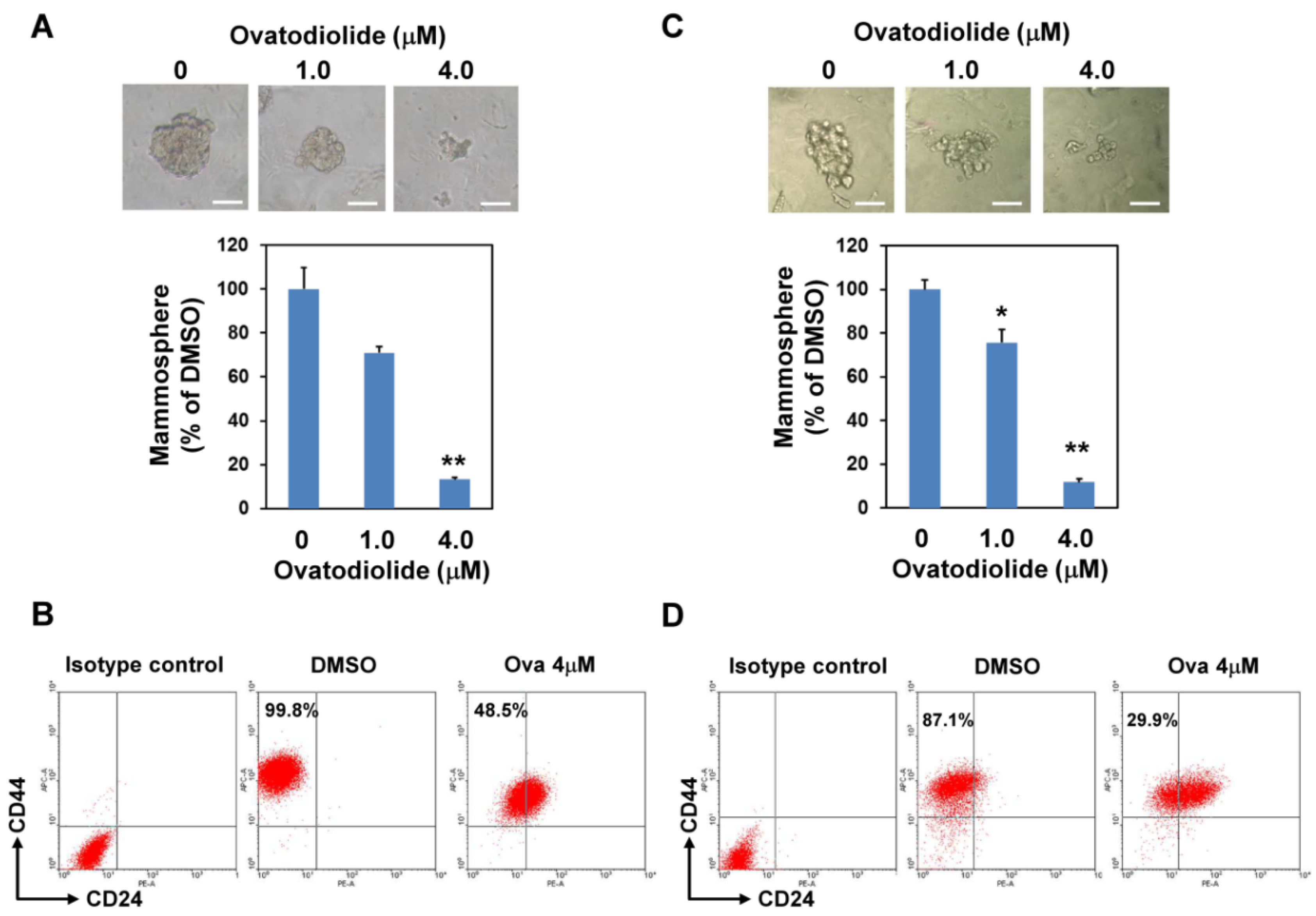
2.1. Ovatodiolide Inhibited Self-Renewal Capability of BCSCs
2.2. Ovatodiolide Downregulated the Expression of Stemness Genes and Hsp27 but Upregulated SMURF2 Expression
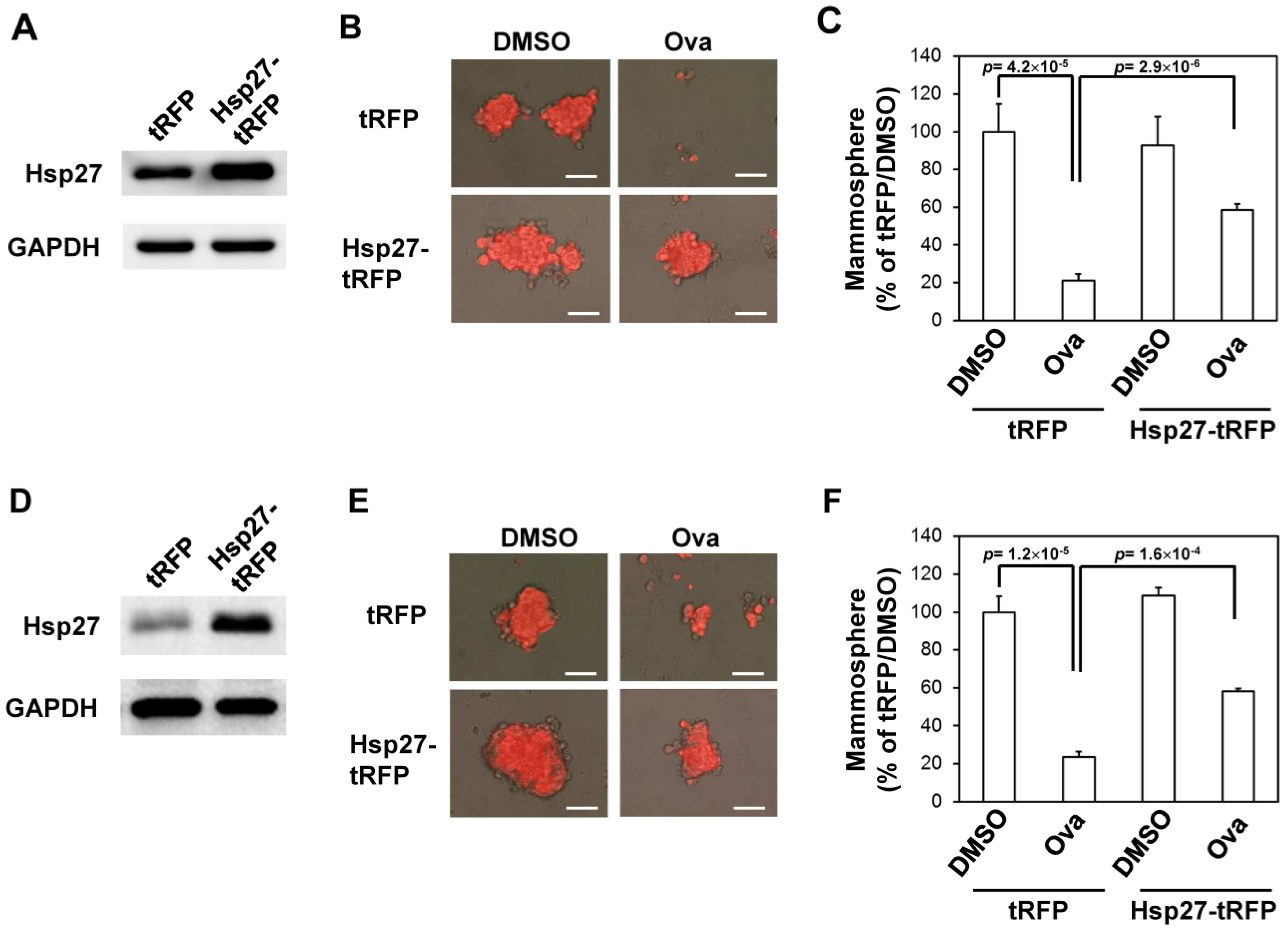
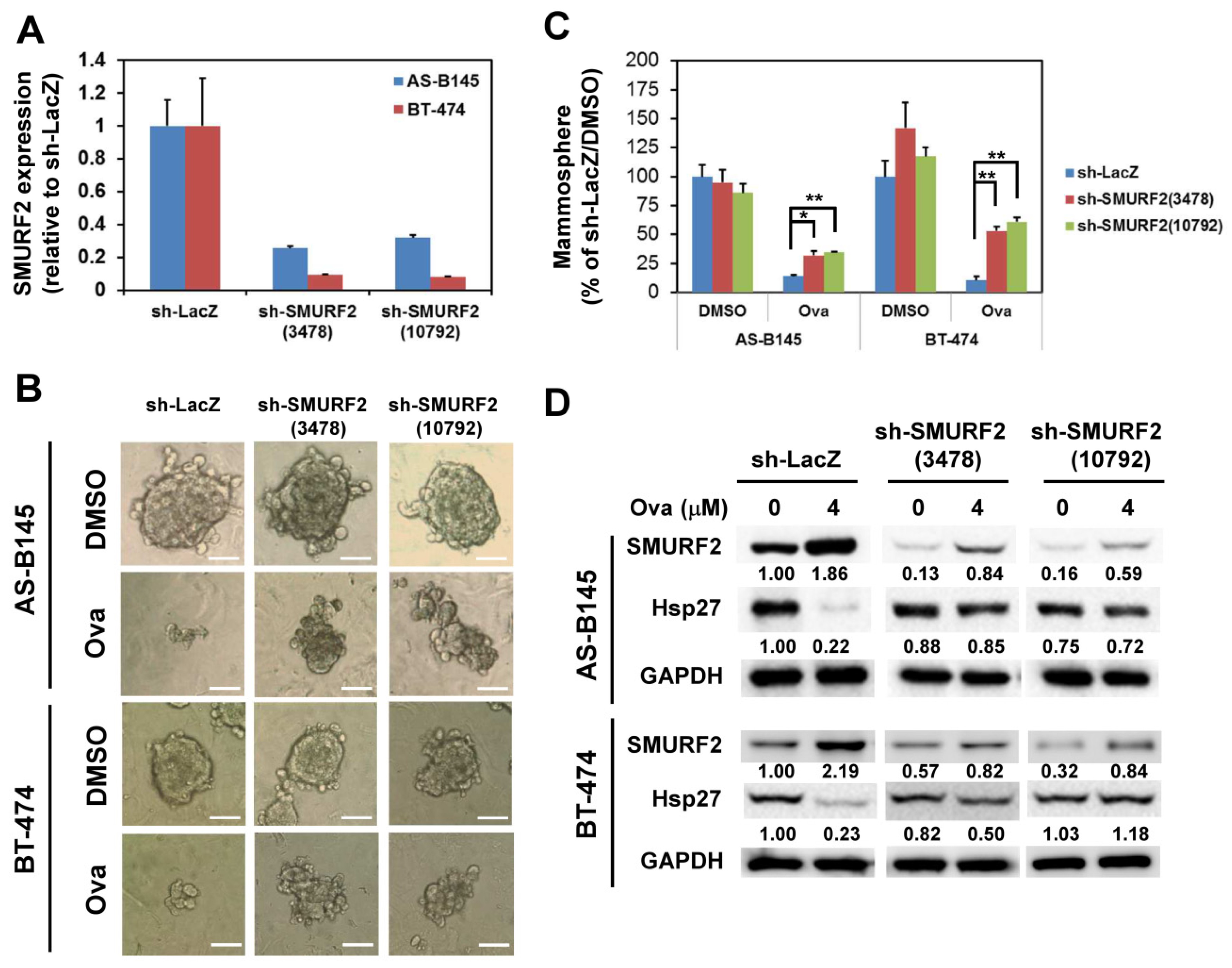
2.3. Overexpression of Hsp27 or Knockdown of SMURF2 Alleviated the Inhibitory Effect of Ovatodiolide
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Reagents and Antibodies
5.2. Cell Culture and Cytotoxicity Assay
5.3. Mammosphere Cultivation
5.4. Analysis of CD24-CD44+ Cells
5.5. Western Blot
5.6. Lentivirus Production and Transduction
- AfeI-Hsp27-F: 5’-AATAGCGCTATGACCGAGCGCCGCGTCCCC-3’
- Hsp27-EcoRI-R: 5’-CGCGAATTCTTACTTGGCGGCAGTCTCATC-3’
5.7. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
- SMURF2
- Forward: 5’-TAGCCCTGGCAGACCTCTTA-3’
- Reverse: 5’- AATACACCTGGCCTTGTTGC-3’
- MRPL19 (internal control)
- Forward: 5’- GGGATTTGCATTCAGAGATCAG-3’
- Reverse: 5’- GGAAGGGCATCTCGTAAG-3’
- qPCR data were analyzed as previous described [35].
5.8. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CSCs | Cancer stem/progenitor cells |
| Hsp27 | heat shock protein 27 |
| Ova | Ovatiodiolide |
| Oct4 | POU class 5 homeobox 1 |
| Nanog | Nanog homeobox |
| SMURF2 | SMAD ubiquitin regulatory factor 2 |
References
- Ajani, J.A.; Song, S.; Hochster, H.S.; Steinberg, I.B. Cancer stem cells: The promise and the potential. Semin. Oncol. 2015, 42 (Suppl. 1), S3–S17. [Google Scholar] [CrossRef] [PubMed]
- Maccalli, C.; de Maria, R. Cancer stem cells: Perspectives for therapeutic targeting. Cancer Immunol. Immunother. CII 2015, 64, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Al-Hajj, M.; Wicha, M.S.; Benito-Hernandez, A.; Morrison, S.J.; Clarke, M.F. Prospective identification of tumorigenic breast cancer cells. Proc. Natl. Acad. Sci. USA 2003, 100, 3983–3988. [Google Scholar] [CrossRef] [PubMed]
- Ginestier, C.; Hur, M.H.; Charafe-Jauffret, E.; Monville, F.; Dutcher, J.; Brown, M.; Jacquemier, J.; Viens, P.; Kleer, C.G.; Liu, S.; et al. Aldh1 is a marker of normal and malignant human mammary stem cells and a predictor of poor clinical outcome. Cell Stem Cell 2007, 1, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Ponti, D.; Costa, A.; Zaffaroni, N.; Pratesi, G.; Petrangolini, G.; Coradini, D.; Pilotti, S.; Pierotti, M.A.; Daidone, M.G. Isolation and in vitro propagation of tumorigenic breast cancer cells with stem/progenitor cell properties. Cancer Res. 2005, 65, 5506–5511. [Google Scholar] [CrossRef]
- Hongisto, V.; Jernstrom, S.; Fey, V.; Mpindi, J.P.; Kleivi Sahlberg, K.; Kallioniemi, O.; Perala, M. High-throughput 3d screening reveals differences in drug sensitivities between culture models of JIMT1 breast cancer cells. PLoS ONE 2013, 8, e77232. [Google Scholar] [CrossRef]
- Kim, S.; Alexander, C.M. Tumorsphere assay provides more accurate prediction of in vivo responses to chemotherapeutics. Biotechnol. Lett. 2014, 36, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Yu, C.C.; Wang, B.Y.; Chang, W.W. Tumorsphere as an effective in vitro platform for screening anti-cancer stem cell drugs. Oncotarget 2016, 7, 1215–1226. [Google Scholar]
- Morrison, B.J.; Hastie, M.L.; Grewal, Y.S.; Bruce, Z.C.; Schmidt, C.; Reynolds, B.A.; Gorman, J.J.; Lopez, J.A. Proteomic comparison of mcf-7 tumoursphere and monolayer cultures. PLoS ONE 2012, 7, e52692. [Google Scholar] [CrossRef]
- Bouvard, C.; Barefield, C.; Zhu, S. Cancer stem cells as a target population for drug discovery. Future Med. Chem. 2014, 6, 1567–1585. [Google Scholar] [CrossRef]
- Powers, M.V.; Workman, P. Inhibitors of the heat shock response: Biology and pharmacology. FEBS Lett. 2007, 581, 3758–3769. [Google Scholar] [CrossRef]
- Soo, E.T.; Yip, G.W.; Lwin, Z.M.; Kumar, S.D.; Bay, B.H. Heat shock proteins as novel therapeutic targets in cancer. In Vivo 2008, 22, 311–315. [Google Scholar] [PubMed]
- Kaigorodova, E.V.; Zavyalova, M.V.; Bogatyuk, M.V.; Tarabanovskaya, N.A.; Slonimskaya, E.M.; Perelmuter, V.M. Relationship between the expression of phosphorylated heat shock protein beta-1 with lymph node metastases of breast cancer. Cancer Biomark 2015, 15, 143–150. [Google Scholar] [PubMed]
- Wei, L.; Liu, T.T.; Wang, H.H.; Hong, H.M.; Yu, A.L.; Feng, H.P.; Chang, W.W. Hsp27 participates in the maintenance of breast cancer stem cells through regulation of epithelial-mesenchymal transition and nuclear factor-kappab. Breast Cancer Res. BCR 2011, 13, R101. [Google Scholar] [CrossRef]
- Lee, C.H.; Wu, Y.T.; Hsieh, H.C.; Yu, Y.; Yu, A.L.; Chang, W.W. Epidermal growth factor/heat shock protein 27 pathway regulates vasculogenic mimicry activity of breast cancer stem/progenitor cells. Biochimie 2014, 104, 117–126. [Google Scholar] [CrossRef]
- Arisawa, M.; Nimura, M.; Fujita, A.; Hayashi, T.; Morita, N.; Koshimura, S. Biological active macrocyclic diterpenoids from chinese drug “fang feng cao”; II. Derivatives of ovatodiolids and their cytotoxity. Planta Med. 1986, 4, 297–299. [Google Scholar] [CrossRef]
- Rao, Y.K.; Chen, Y.C.; Fang, S.H.; Lai, C.H.; Geethangili, M.; Lee, C.C.; Tzeng, Y.M. Ovatodiolide inhibits the maturation of allergen-induced bone marrow-derived dendritic cells and induction of TH2 cell differentiation. Int. Immunopharmacol. 2013, 17, 617–624. [Google Scholar] [CrossRef]
- Lien, H.M.; Wang, C.Y.; Chang, H.Y.; Huang, C.L.; Peng, M.T.; Sing, Y.T.; Chen, C.C.; Lai, C.H. Bioevaluation of anisomeles indica extracts and their inhibitory effects on helicobacter pylori-mediated inflammation. J. Ethnopharmacol. 2013, 145, 397–401. [Google Scholar] [CrossRef]
- Huang, H.C.; Lien, H.M.; Ke, H.J.; Chang, L.L.; Chen, C.C.; Chang, T.M. Antioxidative characteristics of anisomeles indica extract and inhibitory effect of ovatodiolide on melanogenesis. Int. J. Mol. Sci. 2012, 13, 6220–6235. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.F.; Rao, Y.K.; Tzeng, Y.M. Aqueous extract of anisomeles indica and its purified compound exerts anti-metastatic activity through inhibition of nf-kappab/ap-1-dependent MMP-9 activation in human breast cancer MCF-7 cells. Food Chem. Toxicol. 2012, 50, 2930–2936. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.L.; Tsai, P.C.; Hsieh, C.Y.; Chang, L.S.; Lin, S.R. Antimetastatic effect and mechanism of ovatodiolide in MDA-MB-231 human breast cancer cells. Chemico-Biol. Interact. 2011, 194, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Bamodu, O.A.; Huang, W.C.; Tzeng, D.T.; Wu, A.; Wang, L.S.; Yeh, C.T.; Chao, T.Y. Ovatodiolide sensitizes aggressive breast cancer cells to doxorubicin, eliminates their cancer stem cell-like phenotype, and reduces doxorubicin-associated toxicity. Cancer Lett. 2015, 364, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.Y.; Hsu, R.J.; Wu, C.L.; Chang, W.L.; Cha, T.L.; Yu, D.S.; Yu, C.P. Ovatodiolide targets β-catenin signaling in suppressing tumorigenesis and overcoming drug resistance in renal cell carcinoma. Evid. Based Complement. Altern. Med. 2013, 2013, 161628. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhou, M.; Fu, D.; Xu, B.; Fang, T.; Ma, Y.; Chen, J.; Zhang, J. Ubiquitination of heat shock protein 27 is mediated by its interaction with smad ubiquitination regulatory factor 2 in a549 cells. Exp. Lung Res. 2011, 37, 568–573. [Google Scholar] [CrossRef]
- Lee, C.H.; Hong, H.M.; Chang, Y.Y.; Chang, W.W. Inhibition of heat shock protein (hsp) 27 potentiates the suppressive effect of hsp90 inhibitors in targeting breast cancer stem-like cells. Biochimie 2012, 94, 1382–1389. [Google Scholar] [CrossRef]
- Hsu, H.S.; Lin, J.H.; Huang, W.C.; Hsu, T.W.; Su, K.; Chiou, S.H.; Tsai, Y.T.; Hung, S.C. Chemoresistance of lung cancer stemlike cells depends on activation of hsp27. Cancer 2011, 117, 1516–1528. [Google Scholar] [CrossRef]
- Kang, S.H.; Kang, K.W.; Kim, K.H.; Kwon, B.; Kim, S.K.; Lee, H.Y.; Kong, S.Y.; Lee, E.S.; Jang, S.G.; Yoo, B.C. Upregulated hsp27 in human breast cancer cells reduces herceptin susceptibility by increasing her2 protein stability. BMC Cancer 2008, 8, 286. [Google Scholar] [CrossRef]
- Wu, R.; Kausar, H.; Johnson, P.; Montoya-Durango, D.E.; Merchant, M.; Rane, M.J. Hsp27 regulates akt activation and polymorphonuclear leukocyte apoptosis by scaffolding MK2 to akt signal complex. J. Biol. Chem. 2007, 282, 21598–21608. [Google Scholar] [CrossRef]
- Havasi, A.; Li, Z.; Wang, Z.; Martin, J.L.; Botla, V.; Ruchalski, K.; Schwartz, J.H.; Borkan, S.C. Hsp27 inhibits bax activation and apoptosis via a phosphatidylinositol 3-kinase-dependent mechanism. J. Biol. Chem. 2008, 283, 12305–12313. [Google Scholar] [CrossRef]
- Kostenko, S.; Moens, U. Heat shock protein 27 phosphorylation: Kinases, phosphatases, functions and pathology. Cell. Mol. Life Sci. CMLS 2009, 66, 3289–3307. [Google Scholar] [CrossRef]
- Wettstein, G.; Bellaye, P.S.; Kolb, M.; Hammann, A.; Crestani, B.; Soler, P.; Marchal-Somme, J.; Hazoume, A.; Gauldie, J.; Gunther, A.; et al. Inhibition of hsp27 blocks fibrosis development and emt features by promoting snail degradation. FASEB J. 2013, 27, 1549–1560. [Google Scholar] [CrossRef]
- Singh, A.; Settleman, J. Emt, cancer stem cells and drug resistance: An emerging axis of evil in the war on cancer. Oncogene 2010, 29, 4741–4751. [Google Scholar] [CrossRef]
- Fanelli, M.A.; Montt-Guevara, M.; Diblasi, A.M.; Gago, F.E.; Tello, O.; Cuello-Carrion, F.D.; Callegari, E.; Bausero, M.A.; Ciocca, D.R. P-cadherin and beta-catenin are useful prognostic markers in breast cancer patients; beta-catenin interacts with heat shock protein hsp27. Cell Stress Chaperones 2008, 13, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.W.; Lee, C.H.; Lee, P.; Lin, J.; Hsu, C.W.; Hung, J.T.; Lin, J.J.; Yu, J.C.; Shao, L.E.; Yu, J.; et al. Expression of globo H and SSEA3 in breast cancer stem cells and the involvement of fucosyl transferases 1 and 2 in globo H synthesis. Proc. Natl. Acad. Sci. USA 2008, 105, 11667–11672. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Lin, C.W.; Yu, C.C.; Wang, B.Y.; Huang, Y.H.; Hsieh, Y.C.; Kuo, Y.L.; Chang, W.W. Resveratrol suppresses myofibroblast activity of human buccal mucosal fibroblasts through the epigenetic inhibition of zeb1 expression. Oncotarget 2016. [Google Scholar] [CrossRef]





© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, K.-T.; Wang, B.-Y.; Chi, W.-Y.; Chang-Chien, J.; Yang, J.-J.; Lee, H.-T.; Tzeng, Y.-M.; Chang, W.-W. Ovatodiolide Inhibits Breast Cancer Stem/Progenitor Cells through SMURF2-Mediated Downregulation of Hsp27. Toxins 2016, 8, 127. https://doi.org/10.3390/toxins8050127
Lu K-T, Wang B-Y, Chi W-Y, Chang-Chien J, Yang J-J, Lee H-T, Tzeng Y-M, Chang W-W. Ovatodiolide Inhibits Breast Cancer Stem/Progenitor Cells through SMURF2-Mediated Downregulation of Hsp27. Toxins. 2016; 8(5):127. https://doi.org/10.3390/toxins8050127
Chicago/Turabian StyleLu, Kuan-Ta, Bing-Yen Wang, Wan-Yu Chi, Ju Chang-Chien, Jiann-Jou Yang, Hsueh-Te Lee, Yew-Min Tzeng, and Wen-Wei Chang. 2016. "Ovatodiolide Inhibits Breast Cancer Stem/Progenitor Cells through SMURF2-Mediated Downregulation of Hsp27" Toxins 8, no. 5: 127. https://doi.org/10.3390/toxins8050127