Clinical and Molecular Features of Feline Foamy Virus and Feline Leukemia Virus Co-Infection in Naturally-Infected Cats

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimen Collection
2.2. Demographic and Clinical Variable Description
2.3. Specimen Processing and Genomic DNA Extraction
2.4. Serological Screening of FIV and FeLV
2.5. Detection of FFV, FIV and FeLV Using Nested PCR
2.6. FFV Quantitative PCR (qPCR)
2.7. FeLV qPCR
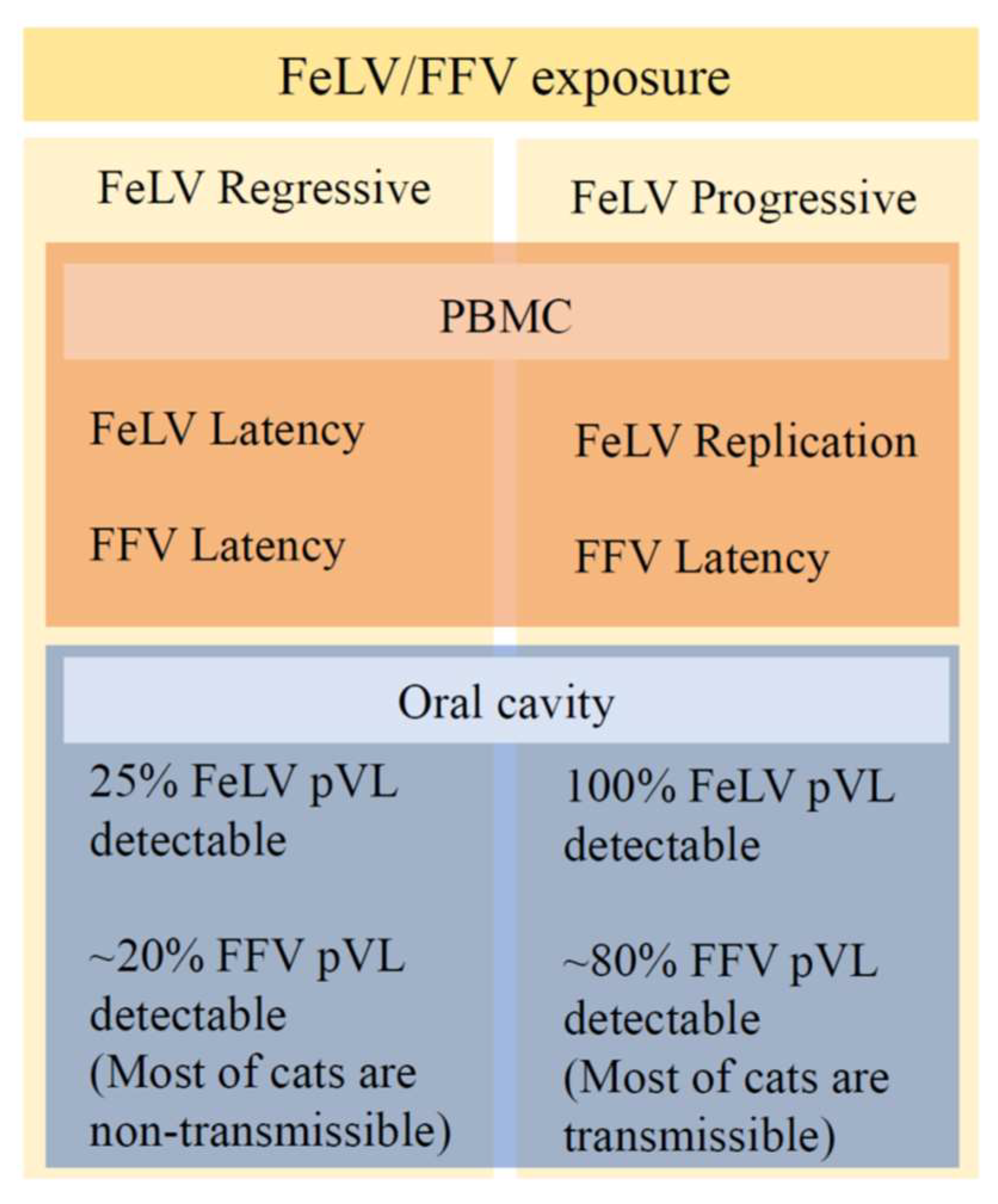
2.8. Classification of FFV and FeLV Infections
2.9. Statistical Analyses
3. Results
3.1. Population Profile
3.2. FIV and FeLV Infection of Domestic Cats
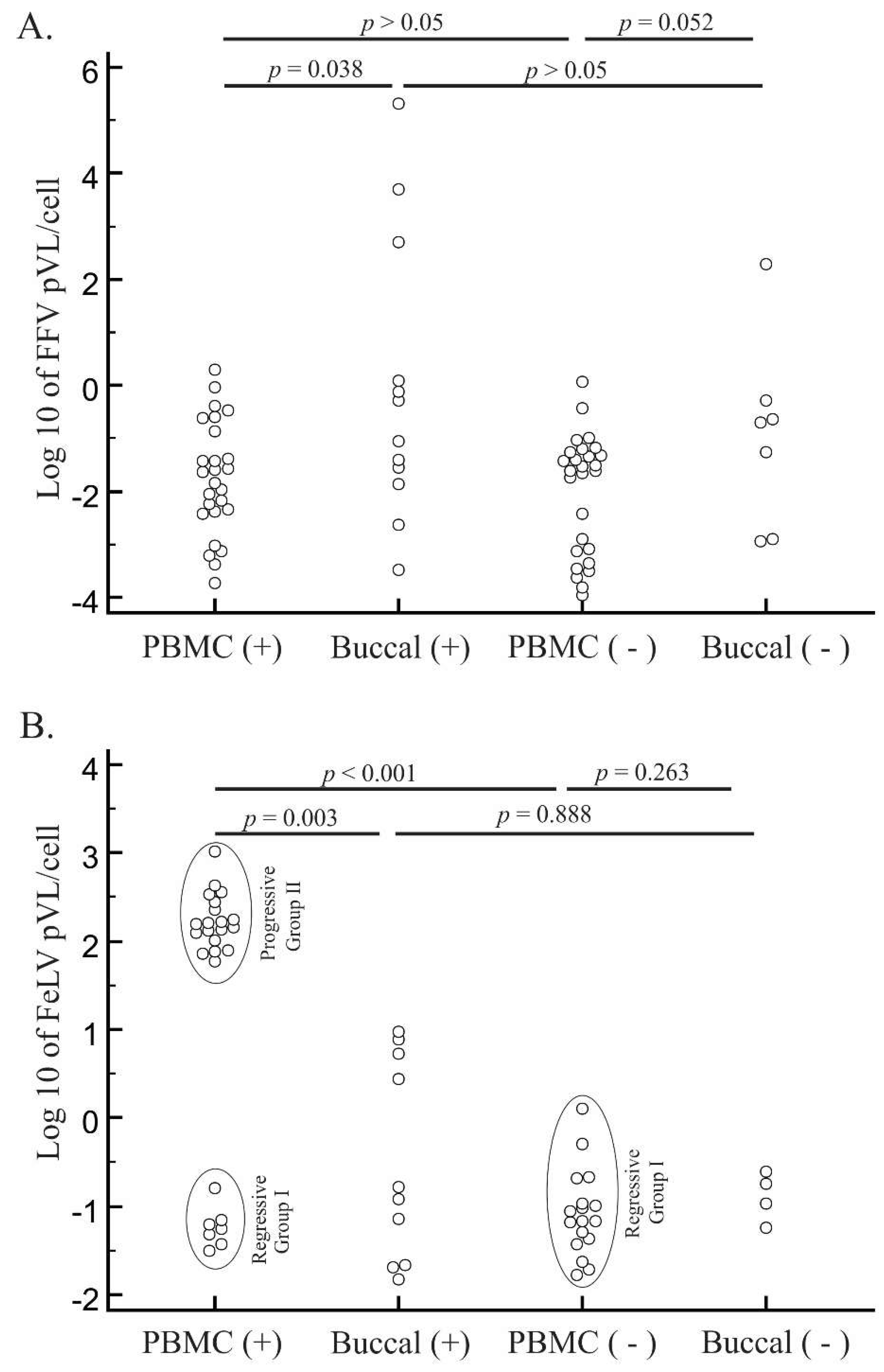
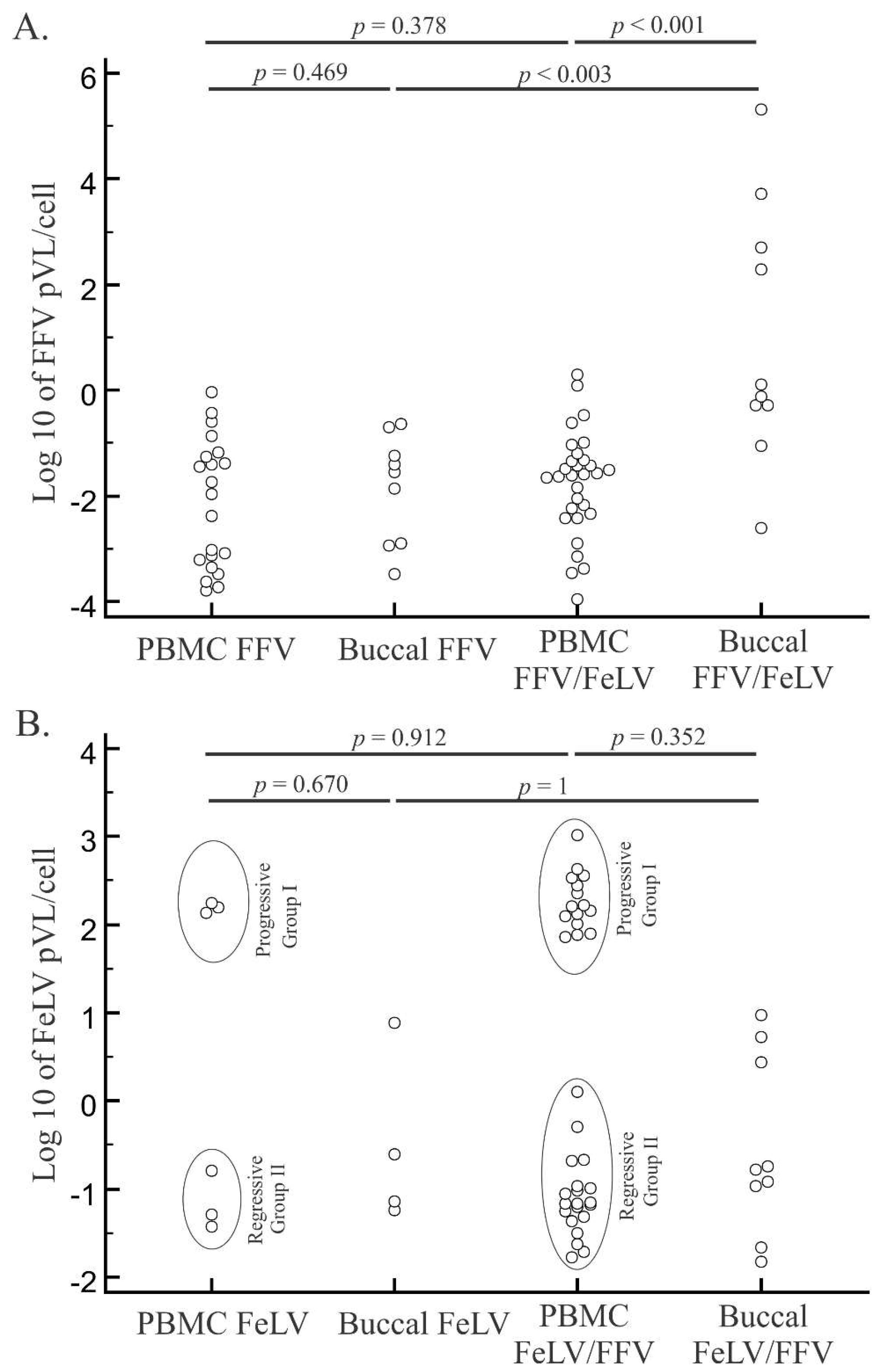
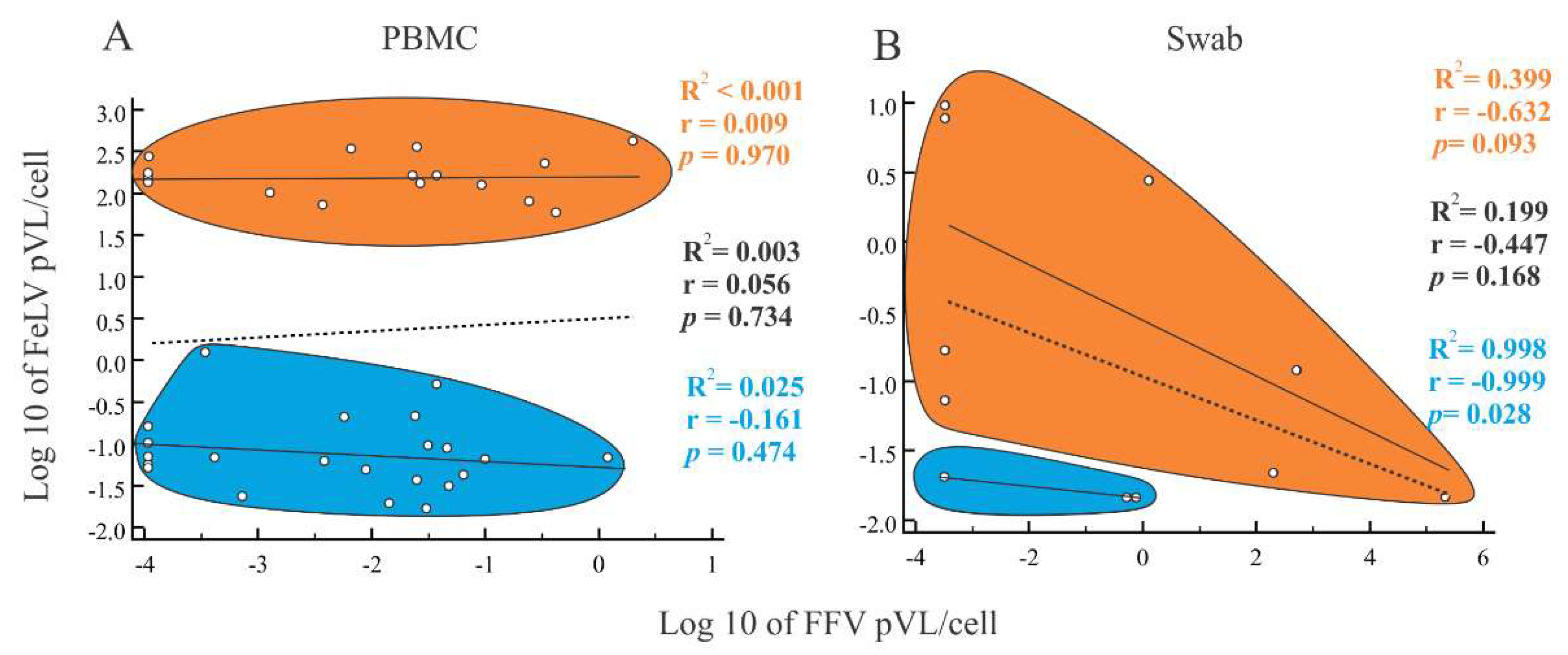
3.3. PCR Detection and Quantification of FFV
3.4. FeLV Viral Load Quantification
3.5. Characteristics of Cats with Feline Retrovirus Co-Infection
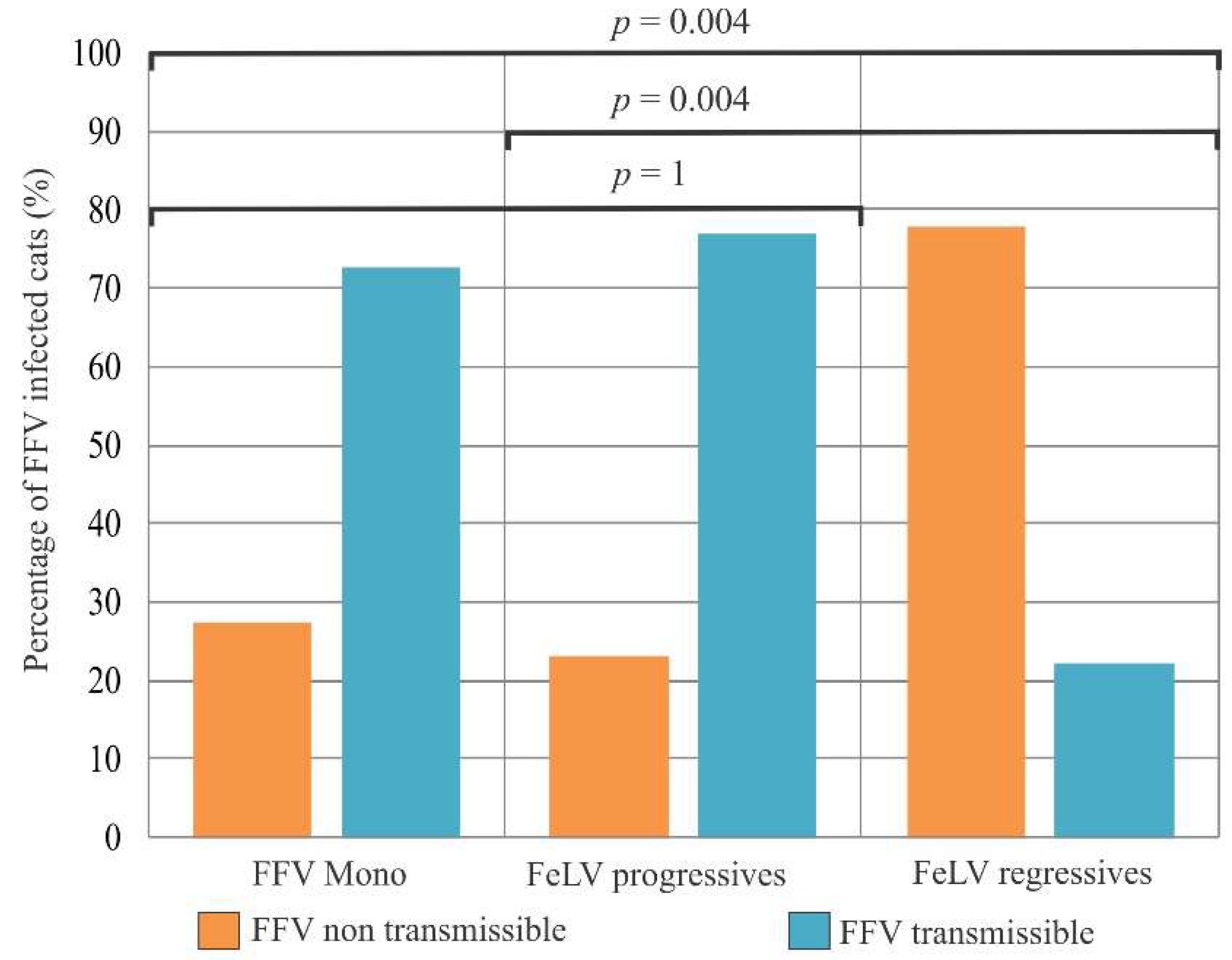
3.6. Association of FFV and FeLV Infection with Clinical Signs
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Linial, M.F.H.; Hahn, B.; Löwer, R.; Neil, J.; Quackenbusch, S.; Rethwilm, A.; Sonigo, P.; Stoye, J.; Tristem, M. Retroviridae. In Virus Taxonomy; Fauquet, C.M.M.M., Manilo, V.J., Desselberger, U., Ball, L.A., Eds.; Elsevier: San Diego, CA, USA, 2005. [Google Scholar]
- Heneine, W.; Switzer, W.M.; Sandstrom, P.; Brown, J.; Vedapuri, S.; Schable, C.A.; Khan, A.S.; Lerche, N.W.; Schweizer, M.; Neumann-Haefelin, D.; et al. Identification of a human population infected with simian foamy viruses. Nat. Med. 1998, 4, 403–407. [Google Scholar] [CrossRef] [PubMed]
- Switzer, W.M.; Bhullar, V.; Shanmugam, V.; Cong, M.E.; Parekh, B.; Lerche, N.W.; Yee, J.L.; Ely, J.J.; Boneva, R.; Chapman, L.E.; et al. Frequent simian foamy virus infection in persons occupationally exposed to nonhuman primates. J. Virol. 2004, 78, 2780–2789. [Google Scholar] [CrossRef] [PubMed]
- Hooks, J.J.; Gibbs, C.J., Jr. The foamy viruses. Bacteriol. Rev. 1975, 39, 169–185. [Google Scholar] [PubMed]
- Buseyne, F.; Betsem, E.; Montange, T.; Njouom, R.; Bilounga Ndongo, C.; Hermine, O.; Gessain, A. Clinical signs and blood test results among humans infected with zoonotic simian foamy virus: A. case-control study. J. Infect. Dis. 2018, 218, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, N.D.; Switzer, W.M.; Carr, J.K.; Bhullar, V.B.; Shanmugam, V.; Tamoufe, U.; Prosser, A.T.; Torimiro, J.N.; Wright, A.; Mpoudi-Ngole, E.; et al. Naturally acquired simian retrovirus infections in central african hunters. Lancet 2004, 363, 932–937. [Google Scholar] [CrossRef]
- Calattini, S.; Betsem, E.B.; Froment, A.; Mauclere, P.; Tortevoye, P.; Schmitt, C.; Njouom, R.; Saib, A.; Gessain, A. Simian foamy virus transmission from apes to humans, rural cameroon. Emerg. Infect. Dis. 2007, 13, 1314–1320. [Google Scholar] [CrossRef] [PubMed]
- Filippone, C.; Betsem, E.; Tortevoye, P.; Cassar, O.; Bassot, S.; Froment, A.; Fontanet, A.; Gessain, A. A severe bite from a nonhuman primate is a major risk factor for HTLV-1 infection in hunters from central africa. Clin. Infect. Dis. 2015, 60, 1667–1676. [Google Scholar] [CrossRef]
- Muniz, C.P.; Cavalcante, L.T.F.; Jia, H.; Zheng, H.; Tang, S.; Augusto, A.M.; Pissinatti, A.; Fedullo, L.P.; Santos, A.F.; Soares, M.A.; et al. Zoonotic infection of brazilian primate workers with new world simian foamy virus. PLoS ONE 2017, 12, e0184502. [Google Scholar] [CrossRef]
- Muniz, C.P.; Troncoso, L.L.; Moreira, M.A.; Soares, E.A.; Pissinatti, A.; Bonvicino, C.R.; Seuanez, H.N.; Sharma, B.; Jia, H.; Shankar, A.; et al. Identification and characterization of highly divergent simian foamy viruses in a wide range of new world primates from brazil. PLoS ONE 2013, 8, e67568. [Google Scholar] [CrossRef]
- Huang, F.; Wang, H.; Jing, S.; Zeng, W. Simian foamy virus prevalence in macaca mulatta and zookeepers. AIDS Res. Hum. Retroviruses 2012, 28, 591–593. [Google Scholar] [CrossRef]
- Rua, R.; Gessain, A. Origin, evolution and innate immune control of simian foamy viruses in humans. Curr. Opin. Virol. 2015, 10, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Switzer, W.M.; Tang, S.; Zheng, H.; Shankar, A.; Sprinkle, P.S.; Sullivan, V.; Granade, T.C.; Heneine, W. Dual simian foamy virus/human immunodeficiency virus type 1 infections in persons from cote d’ivoire. PLoS ONE 2016, 11, e0157709. [Google Scholar] [CrossRef] [PubMed]
- Switzer, W.M.; Garcia, A.D.; Yang, C.; Wright, A.; Kalish, M.L.; Folks, T.M.; Heneine, W. Coinfection with hiv-1 and simian foamy virus in west central africans. J. Infect. Dis 2008, 197, 1389–1393. [Google Scholar] [CrossRef] [PubMed]
- Tsigrelis, C.; Berbari, E.; Temesgen, Z. Viral opportunistic infections in hiv-infected adults. J. Med. Liban. 2006, 54, 91–96. [Google Scholar] [PubMed]
- Choudhary, A.; Galvin, T.A.; Williams, D.K.; Beren, J.; Bryant, M.A.; Khan, A.S. Influence of naturally occurring simian foamy viruses (sfvs) on siv disease progression in the rhesus macaque (macaca mulatta) model. Viruses 2013, 5, 1414–1430. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Miyazawa, T.; Ikeda, Y.; Sato, E.; Nishimura, Y.; Nguyen, N.T.; Takahashi, E.; Mochizuki, M.; Mikami, T. Contrastive prevalence of feline retrovirus infections between northern and southern vietnam. J. Vet. Med. Sci. 2000, 62, 921–923. [Google Scholar] [CrossRef]
- Winkler, I.G.; Lochelt, M.; Flower, R.L. Epidemiology of feline foamy virus and feline immunodeficiency virus infections in domestic and feral cats: A seroepidemiological study. J. Clin. Microbiol. 1999, 37, 2848–2851. [Google Scholar]
- Daniels, M.J.; Golder, M.C.; Jarrett, O.; MacDonald, D.W. Feline viruses in wildcats from scotland. J. Wildl. Dis. 1999, 35, 121–124. [Google Scholar] [CrossRef]
- Glaus, T.; Hofmann-Lehmann, R.; Greene, C.; Glaus, B.; Wolfensberger, C.; Lutz, H. Seroprevalence of bartonella henselae infection and correlation with disease status in cats in switzerland. J. Clin. Microbiol. 1997, 35, 2883–2885. [Google Scholar]
- Bandecchi, P.; Matteucci, D.; Baldinotti, F.; Guidi, G.; Abramo, F.; Tozzini, F.; Bendinelli, M. Prevalence of feline immunodeficiency virus and other retroviral infections in sick cats in italy. Vet. Immunol. Immunopathol. 1992, 31, 337–345. [Google Scholar] [CrossRef]
- Alke, A.; Schwantes, A.; Zemba, M.; Flugel, R.M.; Lochelt, M. Characterization of the humoral immune response and virus replication in cats experimentally infected with feline foamy virus. Virology 2000, 275, 170–176. [Google Scholar] [CrossRef] [PubMed]
- German, A.C.; Harbour, D.A.; Helps, C.R.; Gruffydd-Jones, T.J. Is feline foamy virus really apathogenic? Vet. Immunol. Immunopathol. 2008, 123, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Jarrett, W.F.; Crawford, E.M.; Martin, W.B.; Davie, F. A virus-like particle associated with leukemia (lymphosarcoma). Nature 1964, 202, 567–569. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, K. Clinical aspects of feline retroviruses: A. review. Viruses 2012, 4, 2684–2710. [Google Scholar] [CrossRef] [PubMed]
- Levy, J.; Crawford, C.; Hartmann, K.; Hofmann-Lehmann, R.; Little, S.; Sundahl, E.; Thayer, V. 2008 american association of feline practitioners’ feline retrovirus management guidelines. J. Feline Med. Surg 2008, 10, 300–316. [Google Scholar] [CrossRef] [PubMed]
- Bande, F.; Arshad, S.S.; Hassan, L.; Zakaria, Z.; Sapian, N.A.; Rahman, N.A.; Alazawy, A. Prevalence and risk factors of feline leukaemia virus and feline immunodeficiency virus in peninsular malaysia. BMC Vet. Res. 2012, 8, 33. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, N.R.; Danelli, M.G.; da Silva, L.H.; Hagiwara, M.K.; Mazur, C. Prevalence of feline leukemia virus infection in domestic cats in rio de janeiro. J. Feline Med. Surg 2012, 14, 583–586. [Google Scholar] [CrossRef] [PubMed]
- Sivagurunathan, A.; Atwa, A.M.; Lobetti, R. Prevalence of feline immunodeficiency virus and feline leukaemia virus infection in malaysia: A retrospective study. JFMS Open Rep. 2018, 4, 2055116917752587. [Google Scholar] [CrossRef]
- Francis, D.P.; Essex, M.; Hardy, W.D., Jr. Excretion of feline leukaemia virus by naturally infected pet cats. Nature 1977, 269, 252–254. [Google Scholar] [CrossRef]
- Hofmann-Lehmann, R.; Cattori, V.; Tandon, R.; Boretti, F.S.; Meli, M.L.; Riond, B.; Pepin, A.C.; Willi, B.; Ossent, P.; Lutz, H. Vaccination against the feline leukaemia virus: Outcome and response categories and long-term follow-up. Vaccine 2007, 25, 5531–5539. [Google Scholar] [CrossRef]
- Hofmann-Lehmann, R.; Cattori, V.; Tandon, R.; Boretti, F.S.; Meli, M.L.; Riond, B.; Lutz, H. How molecular methods change our views of FELV infection and vaccination. Vet. Immunol. Immunopathol. 2008, 123, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, N.C.; Ho, E.W.; Brown, M.L.; Yamamoto, J.K. Isolation of a T-lymphotropic virus from domestic cats with an immunodeficiency-like syndrome. Science 1987, 235, 790–793. [Google Scholar] [CrossRef] [PubMed]
- Jarrett, O. Strategies of retrovirus survival in the cat. Vet. Microbiol. 1999, 69, 99–107. [Google Scholar] [CrossRef]
- Rogers, A.B.; Hoover, E.A. Maternal-fetal feline immunodeficiency virus transmission: Timing and tissue tropisms. J. Infect. Dis. 1998, 178, 960–967. [Google Scholar] [CrossRef] [PubMed]
- Medeiros Sde, O.; Martins, A.N.; Dias, C.G.; Tanuri, A.; Brindeiro Rde, M. Natural transmission of feline immunodeficiency virus from infected queen to kitten. Virol. J. 2012, 9, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohmoto, M.; Uetsuka, K.; Ikeda, Y.; Inoshima, Y.; Shimojima, M.; Sato, E.; Inada, G.; Toyosaki, T.; Miyazawa, T.; Doi, K.; et al. Eight-year observation and comparative study of specific pathogen-free cats experimentally infected with feline immunodeficiency virus (FIV) subtypes a and b: Terminal acquired immunodeficiency syndrome in a cat infected with fiv petaluma strain. J. Vet. Med. Sci. 1998, 60, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Shelton, G.H.; Grant, C.K.; Cotter, S.M.; Gardner, M.B.; Hardy, W.D., Jr.; DiGiacomo, R.F. Feline immunodeficiency virus and feline leukemia virus infections and their relationships to lymphoid malignancies in cats: A retrospective study (1968–1988). J. Acquir. Immune Defic. Syndr. 1990, 3, 623–630. [Google Scholar] [PubMed]
- Munro, H.J.; Berghuis, L.; Lang, A.S.; Rogers, L.; Whitney, H. Seroprevalence of feline immunodeficiency virus (FIV) and feline leukemia virus (FELV) in shelter cats on the island of newfoundland, Canada. Can. J. Vet. Res. 2014, 78, 140–144. [Google Scholar]
- Stavisky, J.; Dean, R.S.; Molloy, M.H. Prevalence of and risk factors for fiv and felv infection in two shelters in the United Kingdom (2011–2012). Vet. Rec. 2017, 181, 451. [Google Scholar] [CrossRef]
- Zenger, E.; Brown, W.C.; Song, W.; Wolf, A.M.; Pedersen, N.C.; Longnecker, M.; Li, J.; Collisson, E.W. Evaluation of cofactor effect of feline syncytium-forming virus on feline immunodeficiency virus infection. Am. J. Vet. Res. 1993, 54, 713–718. [Google Scholar]
- Pedersen, N.C.; Torten, M.; Rideout, B.; Sparger, E.; Tonachini, T.; Luciw, P.A.; Ackley, C.; Levy, N.; Yamamoto, J. Feline leukemia virus infection as a potentiating cofactor for the primary and secondary stages of experimentally induced feline immunodeficiency virus infection. J. Virol. 1990, 64, 598–606. [Google Scholar] [PubMed]
- Roy, J.; Rudolph, W.; Juretzek, T.; Gartner, K.; Bock, M.; Herchenroder, O.; Lindemann, D.; Heinkelein, M.; Rethwilm, A. Feline foamy virus genome and replication strategy. J. Virol. 2003, 77, 11324–11331. [Google Scholar] [CrossRef] [PubMed]
- Winkler, I.G.; Flugel, R.M.; Lochelt, M.; Flower, R.L. Detection and molecular characterisation of feline foamy virus serotypes in naturally infected cats. Virology 1998, 247, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Phung, H.T.; Ikeda, Y.; Miyazawa, T.; Nakamura, K.; Mochizuki, M.; Izumiya, Y.; Sato, E.; Nishimura, Y.; Tohya, Y.; Takahashi, E.; et al. Genetic analyses of feline foamy virus isolates from domestic and wild feline species in geographically distinct areas. Virus Res. 2001, 76, 171–181. [Google Scholar] [CrossRef]
- Martins, A.N.; Medeiros, S.O.; Simonetti, J.P.; Schatzmayr, H.G.; Tanuri, A.; Brindeiro, R.M. Phylogenetic and genetic analysis of feline immunodeficiency virus gag, pol, and env genes from domestic cats undergoing nucleoside reverse transcriptase inhibitor treatment or treatment-naive cats in rio de janeiro, brazil. J. Virol. 2008, 82, 7863–7874. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.A.; Cunningham, M.W.; Roca, A.L.; Troyer, J.L.; Johnson, W.E.; O’Brien, S.J. Genetic characterization of feline leukemia virus from florida panthers. Emerg. Infect. Dis. 2008, 14, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Butera, S.T.; Brown, J.; Callahan, M.E.; Owen, S.M.; Matthews, A.L.; Weigner, D.D.; Chapman, L.E.; Sandstrom, P.A. Survey of veterinary conference attendees for evidence of zoonotic infection by feline retroviruses. J. Am. Vet. Med. Assoc. 2000, 217, 1475–1479. [Google Scholar] [CrossRef] [PubMed]
- Tandon, R.; Cattori, V.; Gomes-Keller, M.A.; Meli, M.L.; Golder, M.C.; Lutz, H.; Hofmann-Lehmann, R. Quantitation of feline leukaemia virus viral and proviral loads by taqman real-time polymerase chain reaction. J. Virol. Methods 2005, 130, 124–132. [Google Scholar] [CrossRef]
- Brooks, J.I.; Merks, H.W.; Fournier, J.; Boneva, R.S.; Sandstrom, P.A. Characterization of blood-borne transmission of simian foamy virus. Transfusion 2007, 47, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.M.; Picker, L.J.; Axthelm, M.K.; Hudkins, K.; Alpers, C.E.; Linial, M.L. Replication in a superficial epithelial cell niche explains the lack of pathogenicity of primate foamy virus infections. J. Virol. 2008, 82, 5981–5985. [Google Scholar] [CrossRef] [PubMed]
- Muniz, C.P.; Zheng, H.; Jia, H.; Cavalcante, L.T.F.; Augusto, A.M.; Fedullo, L.P.; Pissinatti, A.; Soares, M.A.; Switzer, W.M.; Santos, A.F. A non-invasive specimen collection method and a novel simian foamy virus (SFV) DNA quantification assay in new world primates reveal aspects of tissue tropism and improved sfv detection. PLoS ONE 2017, 12, e0184251. [Google Scholar] [CrossRef] [PubMed]
- Soliven, K.; Wang, X.; Small, C.T.; Feeroz, M.M.; Lee, E.G.; Craig, K.L.; Hasan, K.; Engel, G.A.; Jones-Engel, L.; Matsen, F.A.t.; et al. Simian foamy virus infection of rhesus macaques in bangladesh: Relationship of latent proviruses and transcriptionally active viruses. J. Virol. 2013, 87, 13628–13639. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.M.; Picker, L.J.; Axthelm, M.K.; Linial, M.L. Expanded tissue targets for foamy virus replication with simian immunodeficiency virus-induced immunosuppression. J. Virol. 2006, 80, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Willett, B.J.; Hosie, M.J. Feline leukaemia virus: Half a century since its discovery. Vet. J. 2013, 195, 16–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betsem, E.; Rua, R.; Tortevoye, P.; Froment, A.; Gessain, A. Frequent and recent human acquisition of simian foamy viruses through apes’ bites in central Africa. PLoS Pathog 2011, 7, e1002306. [Google Scholar] [CrossRef] [PubMed]
- Broussard, S.R.; Comuzzie, A.G.; Leighton, K.L.; Leland, M.M.; Whitehead, E.M.; Allan, J.S. Characterization of new simian foamy viruses from african nonhuman primates. Virology 1997, 237, 349–359. [Google Scholar] [CrossRef]
- Hussain, A.I.; Shanmugam, V.; Bhullar, V.B.; Beer, B.E.; Vallet, D.; Gautier-Hion, A.; Wolfe, N.D.; Karesh, W.B.; Kilbourn, A.M.; Tooze, Z. Screening for simian foamy virus infection by using a combined antigen western blot assay: Evidence for a wide distribution among old world primates and identification of four new divergent viruses. Virology 2003, 309, 248–257. [Google Scholar] [CrossRef]
- Torres, A.N.; Mathiason, C.K.; Hoover, E.A. Re-examination of feline leukemia virus: Host relationships using real-time PCR. Virology 2005, 332, 272–283. [Google Scholar] [CrossRef]
- Hofmann-Lehmann, R.; Huder, J.B.; Gruber, S.; Boretti, F.; Sigrist, B.; Lutz, H. Feline leukaemia provirus load during the course of experimental infection and in naturally infected cats. J. Gen. Virol. 2001, 82, 1589–1596. [Google Scholar] [CrossRef] [Green Version]
- Nesina, S.; Katrin Helfer-Hungerbuehler, A.; Riond, B.; Boretti, F.S.; Willi, B.; Meli, M.L.; Grest, P.; Hofmann-Lehmann, R. Retroviral DNA--the silent winner: Blood transfusion containing latent feline leukemia provirus causes infection and disease in naive recipient cats. Retrovirology 2015, 12, 105. [Google Scholar] [CrossRef]
- Powers, J.A.; Chiu, E.S.; Kraberger, S.J.; Roelke-Parker, M.; Lowery, I.; Erbeck, K.; Troyer, R.; Carver, S.; VandeWoude, S. Feline leukemia virus (FELV) disease outcomes in a domestic cat breeding colony: Relationship to endogenous felv and other chronic viral infections. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed]
- Beebe, A.M.; Faith, T.G.; Sparger, E.E.; Torten, M.; Pedersen, N.C.; Dandekar, S. Evaluation of in vivo and in vitro interactions of feline immunodeficiency virus and feline leukemia virus. Aids 1994, 8, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Siekevitz, M.; Josephs, S.F.; Dukovich, M.; Peffer, N.; Wong-Staal, F.; Greene, W.C. Activation of the hiv-1 ltr by t cell mitogens and the trans-activator protein of HTLV-i. Science 1987, 238, 1575–1578. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Genomic Region | Name | Sense | Primer Sequence (5′-3′) | Reference |
|---|---|---|---|---|---|
| FFV | LTR | FFV LTR out | 1st forward | TGCACAGGAAGCTCCTTTAGGGTA | Designed herein |
| 789(rev) | 1st and 2nd reverse | TCCCCACGTGTAGAGAAACACCACTC | [43] | ||
| 788(fow) | 2nd forward | TGTACGGGAGCTCTTCTCACAGACTTGGC | [43] | ||
| glycoprotein/ | FFV_F1 | 1st forward | CCTCGGGTCCAAACACAGCC | Designed herein | |
| polymerase | FFV_R1 | 1st reverse | CCAAGGTACATCTCCAGG | Designed herein | |
| (gag/pol) | FUV2610s | 2nd forward | AACAGCAACACTCTGATGTTCCCG | [44] | |
| FUV3107a | 2nd reverse | ATATACATCTCCTTCCTGCGTTCC | [44] | ||
| integrase (int) | SIF5N_mod a | 1st forward | TACATGGTTATACCCCACKAAGGCTC | [10] | |
| SIR1NN | 1st and 2nd reverse | GTTTTATYTCCYTGTTTTTCCTYTCCA | [10] | ||
| SIP4N | 2nd forward | TGCATTCCGATCAAGGATCAGCATT | [10] | ||
| envelope | env-f2 | 1st forward | GCTACTTCTACTAGAATAATGTTTTGGATA | [45] | |
| env-r2 | 1st reverse | AGCCACAGTAGTAATTGCATTGGCCAGGCC | [45] | ||
| env-f3 | 2nd forward | GCTTTCAAAAATATGGACATTGTTATGTTA | [45] | ||
| env-r3 | 2nd reverse | GTTTCTCCAAAATCTGCAAGCATATGGATG | [45] | ||
| FIV | reverse | RT out F | 1st forward | GGAGTAGGAGGAGGAAAAAGAGGAAC | [46] |
| transcriptase (RT) | RT out R | 1st reverse | GCCCATCCACTTATATGGGGGC | [46] | |
| RT int F | 2nd forward | GGGCCTCAGGTAAAACAGTGGC | [46] | ||
| RT int R | 2nd reverse | GTCTTCCGGGGTTTCAAATCCCCAC | [46] | ||
| FeLV | envelope | Env Fow (1689 bp) | Forward | TCTATGTTAGGAACCTTAACCGATG | [47] |
| Env Rev (1689 bp) | Reverse | CAGAATATCTGTGGTAC AAGCCTTAA | [47] | ||
| Env Fow (437 bp) | Forward sequencing | GCYTGGTGGGTCTTAGGAA | [47] | ||
| Env Rev (437 bp) | Reverse sequencing | AACARAAGTAAAGACTGTTGG | [47] |
| Category | Number (%) | FFV Mono-Infected (%) | FeLV Mono-Infected (%) | FFV/FeLV Co-Infected (%) |
|---|---|---|---|---|
| Total | 81 (100) | 26 (32) | 8 (10) | 38 (47) |
| Gender (n = 81) | ||||
| Female | 40 (49) | 11 (28) | 3 (8) | 22 (55) |
| Male | 41 (51) | 15 (37) | 5 (12) | 16 (39) |
| Age group (n = 79) | ||||
| Kitten | 18 (23) | 8 (44) | 0 (0) | 7 (38) |
| Young | 16 (20) | 5 (31) | 0 (0) | 8 (50) |
| Adult | 45 (57) | 13 (29) | 7 (16) | 22 (49) |
| Neutered status (n = 66) | ||||
| Neutered/spayed | 35 (53) | 8 (23) | 3 (8) | 20 (57) |
| Intact | 31 (47) | 13 (42) | 3 (10) | 11 (31) |
| Household status (n = 78) | ||||
| Single | 2 (2.6) | 1 (50) | 0 (0) | 1 (50) |
| Multi-cat | 21 (27) | 2 (10) | 3 (14) | 11 (52) |
| Shelter | 12 (15) | 1 (8) | 2 (17) | 9 (75) |
| Feral | 43 (55) | 21 (49) | 2 (5) | 16 (37) |
| Outdoor Access (n = 29) | ||||
| Yes | 9 (31) | 0 (0) | 3 (33) | 5 (56) |
| No | 20 (69) | 4 (20) | 1 (5) | 12 (60) |
| Health status (n = 81) | ||||
| Healthy | 27 (34) | 8 (30) | 2 (7) | 11 (41) |
| Sick | 52 (66) | 18 (35) | 5 (10) | 26 (50) |
| Virus | Category | Household (n = 35) | Feral (n = 43) | Total |
|---|---|---|---|---|
| FeLV | Seropositive only | 3/35 (9%) | 2/43 (5%) | 5/81* (6%) |
| PCR-positive only | 4/35 (11%) | 1/43 (2%) | 5/81* (6%) | |
| PCR-negative and seropositive | 9/35 (26%) | 8/43 (19%) | 17/81* (21%) | |
| PCR-negative and seronegative | 19/35 (54%) | 32/43 (74%) | 51/81*(63%) | |
| Total FeLV infections | 16/35 (46%) | 11/43 (26%) | 27/81* (33%) | |
| FIV | PCR-positive | 0/35 | 1/43 (2%) | 1/81* (1%) |
| Seropositive | 1/35 (3%) | 1/43 (2%) | 2/81* (2%) | |
| Total FIV infections | 1/35 (3%) | 2/43 (4.7%) | 3/81* (3.7%) |
| Fragment | PBMC (%) | Buccal (%) |
|---|---|---|
| long terminal repeat (LTR) | 15/79 b (19) | 0/67 b (0) |
| gag/polymerase (pol) | 27/81 (33) | 13/68 (19) |
| Integrase | 14/81 (17) | 9/68 (13) |
| Envelope | 19/81 (23) | 1/43 b (2) |
| Nested FFV PCR-positive cats a | 33/81 (41) | 19/68 (28) |
| FFV qPCR-positive cats | 53/73 (73) | 19/64 (30) |
| Total FFV-positive cats | 62/81 (77) | 31/68 (46) |
| Factor | Category | FeLV Co-Infection Prevalence | Odds Ratio | 95%CI | p Value |
|---|---|---|---|---|---|
| Sex | Female | 0.67 (22/33) | 1.87 | 0.683–5.148 | 0.223 |
| Male | 0.52 (16/31) | Reference | - | - | |
| Age | Adult | 0.63 (22/35) | 1.47 | 0.534–4.03 | 0.458 |
| Non-adult | 0.53 (15/28) | Reference | - | - | |
| Young | 0.61 (8/13) | 1.1586 | 0.332–4.046 | 0.817 | |
| Non-young | 0.58 (29/50) | Reference | - | - | |
| Kitten | 0.47 (7/15) | 0.5250 | 0.1628–1.6927 | 0.281 | |
| Non-kitten | 0.62 (30/48) | Reference | - | - | |
| Neuter Status | Neutered | 0.71 (20/28) | 2.954 | 0.9378–9.3088 | 0.064 |
| Intact | 0.46 (11/24) | Reference | |||
| Household | Multicat | 0.73 (11/15) | 2.221 | 0.6171–7.9947 | 0.222 |
| Non-multicat | 0.55 (26/47) | Reference | - | - | |
| Shelter | 1 (9/9) | ND a | ND | ND | |
| Non-shelter | 0.47 (28/53) | ND | ND | ND | |
| Street | 0.43 (16/37) | Reference | - | - | |
| Non-street | 0.84 (21/25) | 6.89 | 1.971–24.088 | 0.0025 | |
| Single | 1 (1/1) | ND | ND | ND | |
| Non-single | 0.59 (36/61) | ND | ND | ND |
| Infection Status (n) | Respiratory Tract Disease (%) | Progressive Weight Loss (%) | Ocular Secretion (%) | Anemia (%) |
|---|---|---|---|---|
| FFV-monoinfected (n = 26) | 6 (23.1%) | 4 (15.4%) | 5 (19.2%) | 0/1 (0%) |
| FeLV regressives (n = 23) | 11 (47.8%) | 6 (26.1%) | 2 (8.7%) | 0/2 (0%) |
| FeLV progressives (n = 19) | 6 (31.6%) | 5 (26.3%) | 0 (0%) | 6/8 (75%) |
| FFV negative/FeLV negative (n = 5) | 1 (20%) | 0 (0%) | 0 (0%) | N/A a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavalcante, L.T.F.; Muniz, C.P.; Jia, H.; Augusto, A.M.; Troccoli, F.; Medeiros, S.d.O.; Dias, C.G.A.; Switzer, W.M.; Soares, M.A.; Santos, A.F. Clinical and Molecular Features of Feline Foamy Virus and Feline Leukemia Virus Co-Infection in Naturally-Infected Cats. Viruses 2018, 10, 702. https://doi.org/10.3390/v10120702
Cavalcante LTF, Muniz CP, Jia H, Augusto AM, Troccoli F, Medeiros SdO, Dias CGA, Switzer WM, Soares MA, Santos AF. Clinical and Molecular Features of Feline Foamy Virus and Feline Leukemia Virus Co-Infection in Naturally-Infected Cats. Viruses. 2018; 10(12):702. https://doi.org/10.3390/v10120702
Chicago/Turabian StyleCavalcante, Liliane T. F., Cláudia P. Muniz, Hongwei Jia, Anderson M. Augusto, Fernando Troccoli, Sheila de O. Medeiros, Carlos G. A. Dias, William M. Switzer, Marcelo A. Soares, and André F. Santos. 2018. "Clinical and Molecular Features of Feline Foamy Virus and Feline Leukemia Virus Co-Infection in Naturally-Infected Cats" Viruses 10, no. 12: 702. https://doi.org/10.3390/v10120702