Complete Nucleotide Analysis of the Structural Genome of the Infectious Bronchitis Virus Strain Md27 Reveals its Mosaic Nature

Abstract
:1. Introduction
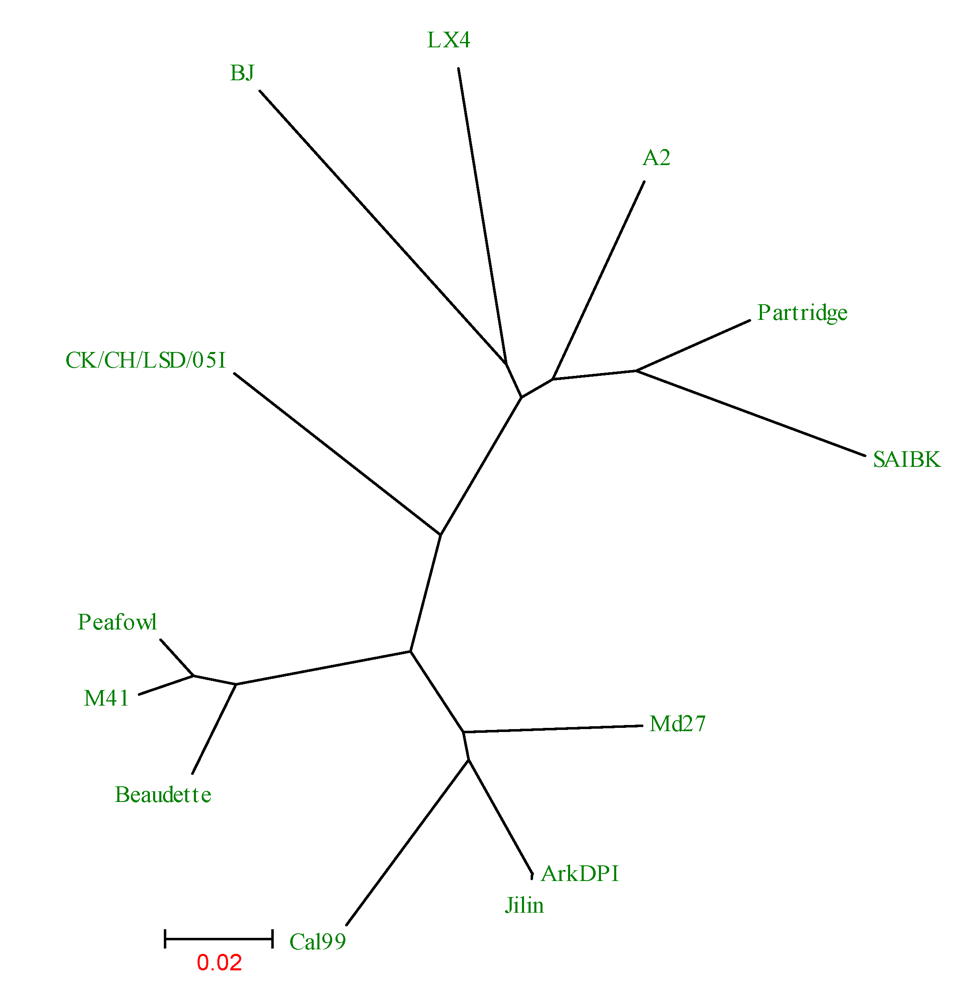
2. Results and Discussion

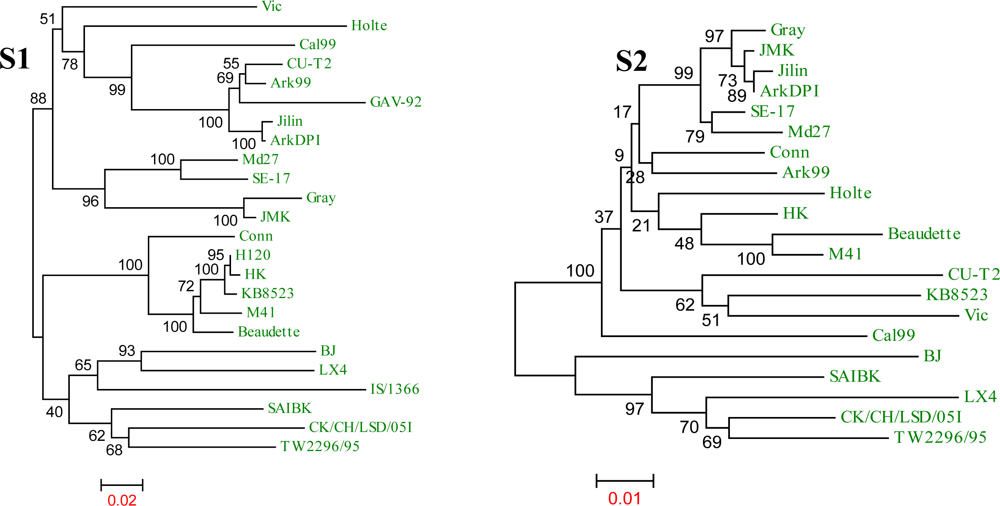{kind=link}
{kind=link}
{kind=link}
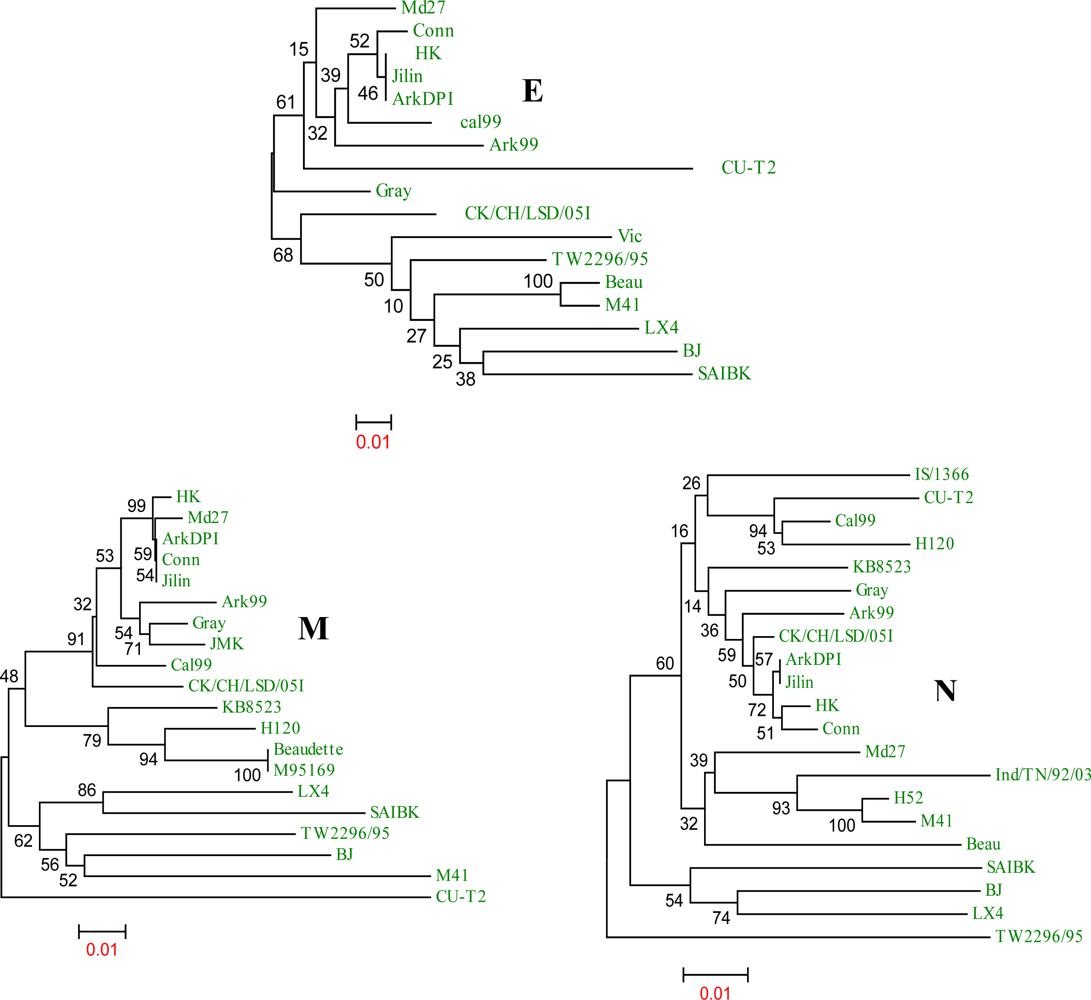
{kind=link}
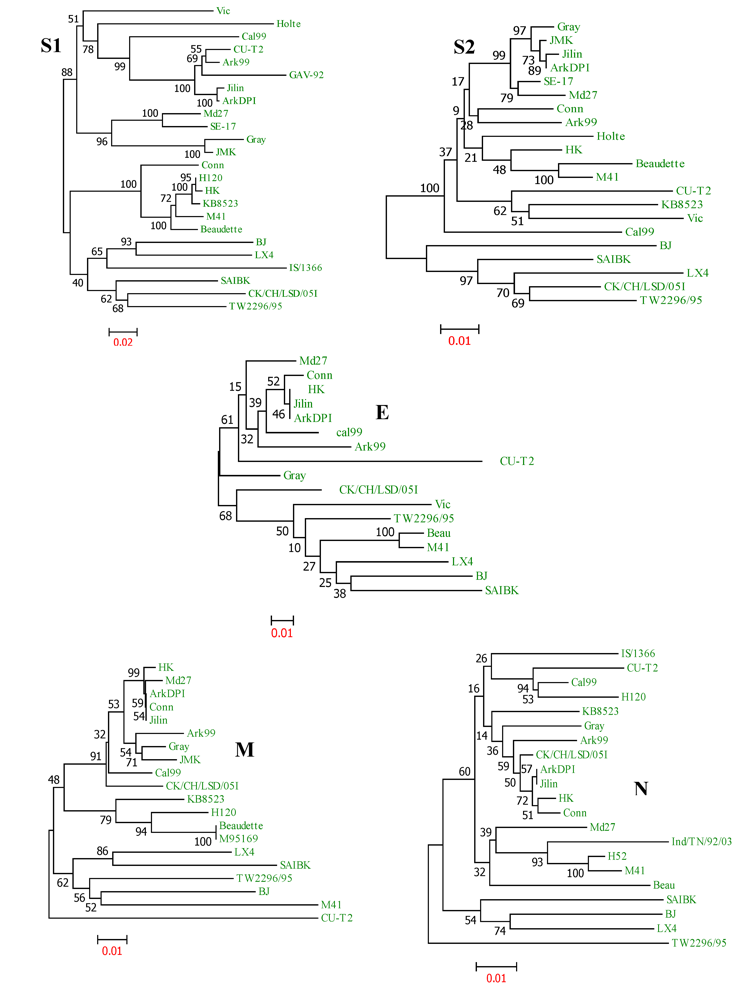{kind=link}
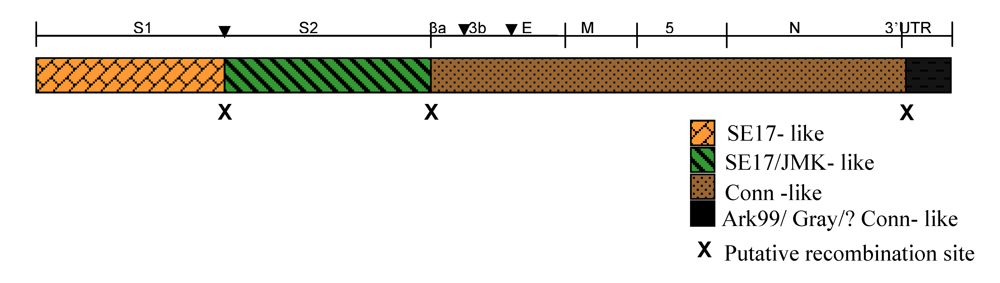{kind=link}
| Gene | Frame | Start | Stop | Size (nucleotides) | Size (amino acids) |
|---|---|---|---|---|---|
| S | +1 | 1 | 3507 | 3507 | 1168 |
| 3a | +3 | 3507 | 3680 | 174 | 57 |
| 3b | +2 | 3680 | 3874 | 195 | 64 |
| 3c(E) | +3 | 3855 | 4178 | 324 | 107 |
| M | +1 | 4156 | 4827 | 672 | 223 |
| 5a | +3 | 5178 | 5375 | 198 | 65 |
| 5b | +2 | 5372 | 5620 | 249 | 82 |
| N | +1 | 5563 | 6792 | 1230 | 409 |
| 3’ UTR | 6793 | 7276 | 483 |
| IBV Strains | S1 | S2 | 3a | 3b | 3c(E) | M | 5a | 5b | N | 3'UTR | 7kb* |
|---|---|---|---|---|---|---|---|---|---|---|---|
| ArkDPI | 86(84) | 98(98) | 91 | 98 | 98(96) | 99(98) | 99 | 98 | 96(96) | 98 | 95 |
| Ark99 | 85(84) | 96(95) | 97 | 98 | 95(91) | 97(96) | NA | NA | 96(95) | 96 | NA |
| Beaudette | 83(80) | 94(93) | 86 | 84 | 88(82) | 91(93) | 93 | 96 | 93(94) | 97 | 91 |
| BJ | 77(76) | 85(89) | 86 | 76 | 87(81) | 90(93) | 86 | 93 | 89(91) | 88 | 85 |
| Cal99 | 82(79) | 94(93) | 97 | 98 | 93(88) | 96(95) | 99 | 98 | 94(95) | 90 | 91 |
| CK/CH/LSD/05I | 78(75) | 87(90) | 85 | 84 | 92(87) | 96(96) | 99 | 99 | 96(96) | 91 | 89 |
| Connecticut | 83(80) | 95(96) | 97 | 98 | 98(97) | 99(98) | 99 | 98 | 96(95) | NA | NA |
| CU-T2 | 85(83) | 94(93) | 94 | 97 | 91(87) | 88(86) | 98 | 98 | 94(93) | 97 | NA |
| DE072 | 62(50) | 75(76) | NA | NA | NA | NA | 98 | 98 | NA | 98 | NA |
| Gray | 86(82) | 98(97) | NA | NA | 96(92) | 97(97) | 98 | 98 | 95(95) | 97 | NA |
| GAV-92 | 83(79) | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| H120 | 83(80) | NA | NA | NA | NA | 91(94) | NA | NA | 92(94) | 82 | NA |
| H52 | 83(80) | NA | NA | NA | NA | NA | NA | NA | 95(95) | NA | NA |
| HK | 83(80) | 95(95) | 91 | 98 | 98(95) | 99(98) | 99 | 98 | 96(95) | 44 | NA |
| Holte | 83(78) | 94(94) | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| Ind/TN/92/03 | NA | NA | NA | NA | NA | NA | NA | NA | 94(94) | NA | NA |
| IS/1366 | 77(74) | NA | NA | NA | NA | NA | NA | NA | 92(94) | NA | NA |
| Jilin | 86(84) | 98(98) | 91 | 98 | 98(96) | 99(98) | 99 | 98 | 96(96) | 98 | 95 |
| JMK | 86(84) | 98(98) | NA | NA | NA | 97(96) | NA | NA | NA | NA | NA |
| KB8523 | 83(80) | 90(91) | NA | NA | NA | 93(94) | NA | NA | 93(95) | NA | NA |
| LX4 | 76(75) | 85(88) | 81 | 76 | 89(81) | 90(91) | 83 | 91 | 88(91) | NA | 26 |
| M41 | 83(79) | 94(94) | 84 | 85 | 88(82) | 91(94) | 90 | 96 | 95(94) | 97 | 91 |
| Qu16 | 84(81) | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| SAIBK | 81(79) | 87(90) | 82 | 82 | 85(81) | 89(92) | 85 | 96 | 87(92) | 90 | 86 |
| SAIBb2 | NA | NA | NA | NA | NA | NA | NA | NA | NA | 85 | NA |
| SE 17 | 96(93) | 98(98) | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| TW2296/95 | 80(78) | 86(89) | 81 | 85 | 88(83) | 90(91) | 86 | 95 | 88(89) | NA | NA |
| UK/2/91 | 77(75) | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| Vic | 82(79) | 89(91) | 84 | 88 | 88(87) | 89(94) | 88 | 94 | 90(93) | NA | NA |



| Coronavirusesb | S | E | M | N |
|---|---|---|---|---|
| BatCoV | 21 | 11 | 29 | 23 |
| BCoV | 21 | 13 | 30 | 24 |
| ECoV | 21 | 13 | 30 | 24 |
| FCoV | 26 | 16 | 23 | 22 |
| HCoV 229E | 26 | 10 | 25 | 22 |
| TGEV | 26 | 20 | 25 | 25 |
| MHV A59 | 22 | 14 | 31 | 25 |
| Peafowl, Avian | 86 | 82 | 94 | 95 |
| Partridge | 84 | 81 | 92 | 92 |
| SARS CoV | 20 | 16 | 29 | 23 |
| SW1 | 25 | 28 | 35 | 34 |
| TCoV | 33 | 91 | 97 | 94 |

3. Materials and Methods
3.1. Virus
3.2. RNA extraction and amplification
3.3. DNA sequence analysis
3.4. GenBank accession numbers
4. Conclusions
Acknowledgments
References
- Cavanagh, D. Nidovirales: a new order comprising Coronaviridae and Arteriviridae. Arch. Virol. 1997, 142, 629–633. [Google Scholar] [PubMed]
- Cavanagh, D.; Naqi, S.A. Infectious bronchitis. Saif Y.M.;, Barnes, H.J.; Glisson J.R.;, Fadley, A.M.; McDougald L.R.;, Swane, D.E., Eds.; Iowa State University Press: Ames, IA, USA, 2003. [Google Scholar]
- Boursnell, M.E.G.; Brown, T.D.K.; Foulds, I.J.; Green, P.F; Tomely, F.M.; Binns, M.M. Completion of the sequence of the genome of the coronavirus avian infectious bronchitis virus. J. Gen Virol. 1987, 68, 57–77. [Google Scholar] [CrossRef] [PubMed]
- Brierley, I.; Digard, P.; Inglis, S.C. Characterization of an efficient coronavirus ribosomal frameshifting signal: Requirement for an RNA pseudoknot. Cell 1989, 57, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Sutou, S.; Sato, S.; Okabe, T.; Nakai, M.; Sasaki, N. Cloning and sequencing of genes encoding structural proteins of avian infectious bronchitis virus. Virology 1988, 165, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D. Coronaviruses in poultry and other birds. Avian. Pathol. 2005, 34, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Gelb Jr., J.Jr.; Wolff, J.B.; Morna, C.A. Variant serotypes of infectious bronchitis virus isolated from commercial layer and broiler chickens. Avian Dis. 1991, 35, 81–87. [Google Scholar]
- Gelb Jr,, J.Jr. Infectious bronchitis. 1998; pp. 169–173. [Google Scholar]
- Infectious bronchitis virus: evidence for recombination within the Massachusetts serotype. Avian Pathol. 1992, 21, 401–408. [CrossRef] [PubMed]
- Gelb Jr,, J.Jr.; Weisman, Y.; Ladman, B.S.; Meir, R. S1 gene characteristics and efficacy of vaccination against infectious bronchitis virus field isolates from the United States and Israel (1996 to 2000). Avian Pathol. 2005, 34, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Kottier, S.A.; Cavanagh, D.; Britton, P. Experimental evidence of recombination in coronavirus infectious bronchitis virus. Virology 1995, 213, 569–580. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.B.; Marquardt, W.W. Strains of infectious bronchitis virus on the Delmarva Peninsula and in Arkansas Avian Dis. 1976, 20, 382–386. [Google Scholar] [PubMed]
- Cavanagh, D.; Davis, P.J.; Cook, J.K.A.; Li, D.; Kant, A.; Koch, G. Location of the amino acid differences in the S1 spike glycoprotein subunit of closely related serotypes of infectious bronchitis virus. Avian Pathol. 1992, 21, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Parr, R.L.; Collisson, E.W. Epitopes on the spike protein of a nephropathogenic strain of infectious bronchitis virus. Arch. Virol. 1993, 133, 369–383. [Google Scholar] [CrossRef] [PubMed]
- Callison, S.A.; Jackwood, M.W.; Hilt, D.A. Infectious bronchitis virus S2 gene sequence variability may affect S1 subunit specific antibody binding. Virus Genes 1999, 19, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.X.; Cavanagh, D.; Green, P. A polycistronic mRNA specified by the coronavirus infectious bronchitis virus. Virology 1991, 184, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Locker, J.K.G.; Griffiths, M.C.; Horzinek Rottier, P.J. O-glycosylation of the coronavirus M protein. Differential localization of sialyltransferases in N- and O-linked glycosylation. J. Biol. Chem. 1992, 267, 14094–14101. [Google Scholar] [PubMed]
- Wilbur, S.M.; Nelson, G.W.; Lai, M.M.; McMillan, M.; Stohlman, S.A. Phosphorylation of the mouse hepatitis virus nucleocapsid protein. Biochem. Biophys. Res. Commun. 1986, 141, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Zakhartchouk, A.N.; Viswanathan, S.; Mahony, J.B.; Gauldie, J.; Babiuk, L.A. Severe acute respiratory syndrome coronavirus nucleocapsid protein expressed by an adenovirus vector is phosphorylated and immunogenic in mice. J. Gen. Virol. 2005, 86, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Spaan, W.; Cavanagh, D.; Horzinek, M.C. Coronaviruses: structure and genome expression. J. Gen. Virol. 1988, 69, 2939–2952. [Google Scholar] [CrossRef] [PubMed]
- Collisson, E.W.; Williams, A.K.; Chung, S.I. Interactions between the IBV nucleocapsid protein and RNA sequences specific for the 3' end of the genome. Adv. Exp. Med. Biol. 1995, 380, 523–528. [Google Scholar] [PubMed]
- Williams, A.K.; Wang, L.; Sneed, L.W.; Collisson, E.W. Analysis of a hypervariable region in the 3’ non-coding end of the infectious bronchitis virus genome. Virus Res. 1993, 28, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D.; Mawditt, K.; Adzhar, A.; Gough, R.E.; Picault, J.P.; Naylor, C.J.; Haydon, D.; Shaw, K.; Britton, P. Does IBV change slowly despite the capacity of the spike protein to vary greatly? Adv. Exp. Med. Biol. 1998, 440, 729–734. [Google Scholar] [PubMed]
- Koch, G.; Hartog, L.; Kant, A.; van Roozelaar, D.J. Antigenic domains on the peplomer protein of avian infectious bronchitis virus: correlation with biological functions. J. Gen. Virol. 1990, 71, 1929–1935. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Karaca, K.; Parrish, C.R.; Naqi, S.A. A novel variant of avian infectious bronchitis virus resulting from recombination among three different strains. Arch. Virol. 1995, 140, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.K.; Wang, L.; Sneed, L.W; Collisson, E W. Comparative analyses of the nucleocapsid genes of several strains of infectious bronchitis virus and other coronaviruses. Virus Res. 1992, 25, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Gelb Jr,, J.; Leary, J.H.; Rosenberger, J.K. Prevalence of Arkansas-type infectious bronchitis virus in Delmarva Peninsula chickens. Avian Dis. 1983, 27, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Ammayappan, A.; Upadhyay, C.; Gelb Jr,, J.Jr.; Vakharia, V.N. Complete genomic sequence analysis of infectious bronchitis virus Ark DPI strain and its evolution by recombination. Virol. J. 2008, 5, 157. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D.; Picault, J.P.; Gough, R.; Hess, M.; Mawditt, K.; Britton, P. Variation in the spike protein of the 793/B type of infectious bronchitis virus, in the field and during alternate passage in chickens and embryonated eggs. Avian Pathol. 2005, 34, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.M.C.; Cavanagh, D. The molecular biology of coronaviruses. Adv. Virus Res. 1997, 48, 1–100. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yuan, X.; Collisson, E.W. Experimental confirmation of recombination upstream of the S1 hypervariable region of infectious bronchitis virus. Virus Res. 1997, 49, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, K.B.; Nicholas, H.B.J.; Deerfield, D.W. GeneDoc: Analysis and visualization of genetic variation. EMBNEW. NEWS 1997, 4, 14. [Google Scholar]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Share and Cite
Ammayappan, A.; Vakharia, V.N. Complete Nucleotide Analysis of the Structural Genome of the Infectious Bronchitis Virus Strain Md27 Reveals its Mosaic Nature. Viruses 2009, 1, 1166-1177. https://doi.org/10.3390/v1031166
Ammayappan A, Vakharia VN. Complete Nucleotide Analysis of the Structural Genome of the Infectious Bronchitis Virus Strain Md27 Reveals its Mosaic Nature. Viruses. 2009; 1(3):1166-1177. https://doi.org/10.3390/v1031166
Chicago/Turabian StyleAmmayappan, Arun, and Vikram N. Vakharia. 2009. "Complete Nucleotide Analysis of the Structural Genome of the Infectious Bronchitis Virus Strain Md27 Reveals its Mosaic Nature" Viruses 1, no. 3: 1166-1177. https://doi.org/10.3390/v1031166