
COVID-19 Anosmia: High Prevalence, Plural Neuropathogenic Mechanisms, and Scarce Neurotropism of SARS-CoV-2?



1
Healthy Longevity Translational Research Program, Department of Anatomy, Yong Loo Lin School of Medicine, National University of Singapore, Singapore 117594, Singapore
2
Infectious Diseases Translational Research Program, Department of Otolaryngology, Yong Loo Lin School of Medicine, National University of Singapore, Singapore 119228, Singapore
*
Author to whom correspondence should be addressed.
Viruses 2021, 13(11), 2225; https://doi.org/10.3390/v13112225
Submission received: 30 September 2021
/
Revised: 28 October 2021
/
Accepted: 30 October 2021
/
Published: 4 November 2021
(This article belongs to the Special Issue Early Immune-Modulator Treatment in COVID-19)
{kind=link}
{kind=link}
Abstract
:Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is the causative pathogen of coronavirus disease 2019 (COVID-19). It is known as a respiratory virus, but SARS-CoV-2 appears equally, or even more, infectious for the olfactory epithelium (OE) than for the respiratory epithelium in the nasal cavity. In light of the small area of the OE relative to the respiratory epithelium, the high prevalence of olfactory dysfunctions (ODs) in COVID-19 has been bewildering and has attracted much attention. This review aims to first examine the cytological and molecular biological characteristics of the OE, especially the microvillous apical surfaces of sustentacular cells and the abundant SARS-CoV-2 receptor molecules thereof, that may underlie the high susceptibility of this neuroepithelium to SARS-CoV-2 infection and damages. The possibility of SARS-CoV-2 neurotropism, or the lack of it, is then analyzed with regard to the expression of the receptor (angiotensin-converting enzyme 2) or priming protease (transmembrane serine protease 2), and cellular targets of infection. Neuropathology of COVID-19 in the OE, olfactory bulb, and other related neural structures are also reviewed. Toward the end, we present our perspectives regarding possible mechanisms of SARS-CoV-2 neuropathogenesis and ODs, in the absence of substantial viral infection of neurons. Plausible causes for persistent ODs in some COVID-19 convalescents are also examined.
1. Introduction
Apart from causing respiratory, cardiovascular, and systemic problems, COVID-19 is also accompanied by frequent neurological manifestations such as headache, dizziness, anosmia, ageusia, or even stroke [1,2,3,4,5]. Among others, COVID-19-related olfactory dysfunctions (ODs), as represented by anosmia or hyposmia, are highly relevant to upper respiratory infection, as these dysfunctions imply infection and pathology of the olfactory epithelium (OE) lining the superior recess of the nasal cavity. It is thus not surprising that COVID-19 related ODs have attracted much attention from both the clinical and basic medicine research communities [6,7,8,9,10].
ODs are quite common in disorders of the nose. The causes of ODs vary from nasal congestion, OE inflammation, infection or damage, or structural functional abnormalities of the olfactory nerve, olfactory bulb (OB), or other central nervous system (CNS) structures. However, the ODs in COVID-19 seem somehow special in that these deficits are unusually prevalent, sometimes appear before other symptoms, and, on occasions, might even be the only symptoms of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection without apparent nasal congestion or inflammation. The incidence of smell and/or taste impairment in COVID-19 patients varied from as low as 5% to as high as 98% in the literature, depending on areas, populations, SARS-CoV-2 variants, and methods of diagnosis, but most analyses have reported an OD rate of 20–80% [11,12,13,14,15,16,17,18,19,20,21]. Although a majority of COVID-19-related ODs disappear in a few weeks, the deficits in some patients could persist long after resolution of other COVID-19 abnormalities [22,23,24,25].
The questions then arise as to the reasons for the unusually high prevalence of ODs in COVID-19, the possibilities of SARS-CoV-2 invasion or infliction of acute and chronic damages to the peripheral or central olfactory system, and the prospects of olfaction recovery in the cases of protracted post-COVID-19 ODs. There have been many reports, experiments, or speculations with regard to COVID-19-related ODs within the short time period since the outbreak of the COVID-19 pandemic, one often contradicting the other. Here, we attempt to first discuss possible molecular and cytological substrates for high susceptibility of the OE to SARS-CoV-2 infection. SARS-CoV-2 neurotropism (or the lack of it) and COVID-19 neuropathology will then be analyzed. In view of the scarce neurotropism of the virus, plausible mechanisms of COVID-19 neuropathogenesis and ODs are explored, such as neural support deprivation, inflammation, immune reactions at the OE, anterograde degeneration or molecular trafficking along nerve fibers, and microvascular thrombosis in the OB or other CNS regions. Possible causes of protracted ODs after COVID-19 are also briefly reviewed.
2. Cytological and Molecular Basis for High Prevalence of Olfactory Dysfunctions in COVID-19
The sense of smell (olfaction) starts from the binding of airborne odor molecules (odorants) to their receptors on the surface of the OE at the superior part of the nasal cavity. Here, chemical characteristics of the odorants are encoded into electrical signals, and then transmitted monosynaptically through the olfactory nerves (cranial nerves I) to the OB. After relay and integration there, the olfactory impulses are further transmitted to higher order olfactory regions of the CNS for olfactory perception, reactions, memory, and other neural processes [26,27].
2.1. Basic Histology and Cytology of the OE
The OE lines the superior vault of the nasal cavity. Its location near the entrance of the upper respiratory tract facilitates early detection of important or potentially harmful odorants in the inhaled air, but this frontline positioning of the special sense receptor organ also renders the OE vulnerable to pathogens or damages in the upper respiratory tract [28,29]. Histologically, the OE is a layer of pseudostratified columnar epithelium, as is the respiratory epithelium (RE) lining most other parts of the nasal cavities and paranasal sinuses. At the cytological level, however, the OE and RE differ significantly from each other. Specifically, the OE is made of ciliated olfactory receptor neurons (ORNs), sustentacular supporting cells, globose and horizontal basal cells, occasional microvillar cells and ductal cells of Bowman’s glands, plus glandular cells of Bowman’s glands in the lamina propria of the olfactory mucosa [28,30,31]. The sustentacular and microvillar cell nuclei usually occupy a more apical position of the OE; ORN cell bodies are mostly located in the middle layer, whereas basal cells are found next to or close to the basement membrane. The nasal RE, however, is a ciliated pseudostratified columnar epithelium made of ciliated and non-ciliated columnar epithelial cells, secretory goblet cells, basal cells, occasional brush cells, small granule cells, and ductal cells of glands, plus glandular cells in the lamina propria [32].
The bipolar ORNs are directly exposed, at the dendritic knob and cilia, to the nasal mucus and nasal cavity environment. While the direct interaction with the inhaled air enables a high sensitivity to odorants in the immediate environment, the direct contact with nasal mucus and air subjects the ORNs to the risk of potential harm by detrimental molecules or microorganisms that are breathed in and out of the nasal cavity. Probably because of this vulnerability, the ORNs have a relatively short lifespan of only a few weeks and are constantly replaced by new receptor neurons generated from OE basal cells [28,33].
At the axonal end, the ORNs are monosynaptically connected with neurons of the olfactory bulb of the CNS [27,28]. The olfactory nerve not only conducts olfactory nerve impulses to the olfactory bulb but may also serve as a trafficking pathway for certain intrinsic or extrinsic molecules, toxins, or viruses along the axoplasm from the OE to the OB, or vice versa. As compared with trafficking through the blood stream and blood–brain barrier, the olfactory nerve represents an alternative and more direct route of CNS vulnerability to infections/toxicities of nasal origin [34,35,36,37]. The direct neural pathway and its trafficking capability are sometimes also used for delivering therapeutics or other molecules to the CNS, to bypass the blood–brain barrier [38,39,40,41,42].
2.2. Why Is the OE Especially Susceptible to SARS-CoV-2 Infection?
In terms of luminal surface area, the OE accounts for only about 5% of the total nasal epithelium in humans [7,43,44], but ODs (anosmia, hyposmia, etc.) have been reported in up to about 80% of COVID-19 patients, and ODs are sometimes the first or only clinical manifestation of the infection [11,12,13,14,15,16,17,18,19,20,21]. Sudden anosmia has been reported to be even more predictive of SARS-CoV-2 infection than any other symptoms, including fever, cough, hoarse voice, or shortness of breath [45].
The disproportionately high prevalence and specificity of ODs suggest high susceptibility of the OE to SARS-CoV-2 infection. Why is this so? There is no definitive answer to the question yet, but difference in expression of angiotensin-converting enzyme 2 (ACE2, the SARS-CoV-2 receptor) has been well noted between the OE and RE. There have been reports of more abundant ACE2 expression in the OE (up to hundreds of times more in immunofluorescence intensity, as quantified by laser scanning confocal microscopy) than in the neighboring nasal RE [46,47,48] (see below for further details concerning ACE2 expression in specific cell types of the OE, RE, and some other tissues). Besides, structurally, the OE luminal surface is mostly occupied by thin and long microvilli that are rooted from the apical surface of olfactory sustentacular cells. This coat of microvilli could effectively increase dozens-fold to hundred-fold the apical surface area of OE sustentacular cells (Figure 1). In contrast, few cells of the nasal RE bear apical microvilli. Even though the motile apical cilia of respiratory epithelial cells could also multiply the surface area, this cilia mechanism might not effectively serve the purpose for increased viral binding. Coordinated cilia motility actually propels out pathogens, particles, and cell debris to clean up the airway [49,50]. Cellular microvilli, in contrast, are well known for functional roles to increase cellular surface area for binding or absorption [51]. The possibility of OE sustentacular cell microvilli as an effective areal multiplier for binding SARS-CoV-2 is further supported by the presence here of ACE2 receptor for the virus (see below), although it awaits future experimental evidence to verify this notion specifically.
3. Neurotropism and Neuropathology of SARS-CoV-2?
Various molecules or viruses can preferentially bind to and enter neuronal cells, migrate along neuronal axoplasm, and are thus neurotropic. Some neurotropic molecules are of intrinsic origins from the nervous system or other parts of the body (such as NGF, BDNF, and NT-3) [37]. Other neurotropic molecules are of extrinsic origins, including bacterial or fungal toxins (such as cholera toxin, tetanus toxin, and botulinum neurotoxin) and lectins (such as wheat germ agglutinin (WGA) and phaseolus vulgaris leucoagglutinin (PhA-L)) [35,55,56,57,58]. A number of viruses, including the herpes simplex viruses, poliovirus, rabies virus, and bovine herpesvirus 5 (BHV5), are also neurotropic [34,36,59,60,61]. A majority of these neurotropic molecules/viruses have designated binding receptors identified on neuronal cells, and thus could enter neurons through receptor-mediated endocytosis. Many of them can also be transported anterogradely or retrogradely along the neuronal processes (mostly axons) or neural pathways. A few are even capable of travelling transsynaptically (transneuronally) from one neuron to other neurons or effector cells through synaptic connections. The anterograde, retrograde, and transsynaptic trafficking properties of some neurotropic molecules (such as WGA, PHA-L, cholera toxin) or viruses (such as HSV) have been exploited for tracing neuronal connections in the nervous system [58,62,63,64,65].
3.1. SARS-CoV-2 Receptor and Uptake Priming Protein in the OE and Related Structures
A few groups examined the expression of ACE2 (the SARS-CoV-2 receptor) and transmembrane serine protease 2 (TMPRSS2, the SARS-CoV-2 cell entry-priming protease) to assess possible tropism of the virus to specific tissues and cell types [66,67]. To that end, approaches such as single-cell RNA-sequencing, immunocytochemistry, immunohistochemistry, and in situ hybridization histochemistry have been employed. The results still somehow vary, but basic expression patterns in the OE, RE, and elsewhere have emerged. In both human and mouse OE, ACE2 immunoreactivity has been found mainly on the sustentacular cells, especially at the supranuclear part, apical surface, and microvilli of sustentacular cells. To a variable extent, ACE2 could also be seen on OE basal cells, ductal and glandular cells of Bowman’s glands. Minimal or no ACE2 was detected on mature ORNs. TMPRSS2 exhibited a cell-type expression pattern similar to ACE2, albeit less cell type-specific [46,47,48,68]. In human nasal RE, ACE2 has been detected in ciliated columnar respiratory epithelial cells, but not in the secretory goblet cells [69]. Another study, in contrast, found ACE2 on RE secretory cells [47].
In the CNS, including the olfactory bulb, most studies have observed significant ACE2 in brain vasculature, but no or little ACE2 in neurons or glia [68,70,71]. ACE2 was found on vascular endothelial cells and on pericytes of capillaries. TMPRSS2 transcripts were barely detected in the mouse CNS [46]. One group, however, reported expression of ACE2 and TMPRSS2 in astrocytes and microglia, in addition to brain vascular endothelial cells and pericytes [72].
3.2. SARS-CoV-2 Infection and COVID-19 Pathology in the OE and Related Structures
Many groups have examined SARS-CoV-2 infection and/or pathology in various cells, tissues, and organ systems in human autopsy or biopsy samples, and in infected model animals such as the nonhuman primates, golden Syrian hamster, ferret, and hACE2 transgenic mouse. Here, we briefly examine the data related to the nervous system, especially the OE and OB. In COVID-19 human autopsy, SARS-CoV-2 spike protein was detected in many OE sustentacular cells and RE epithelial cells, and occasionally in probable progenitor ORNs positive for Olig2 [73]. Olfactory mucosa showed the highest SARS-CoV-2 viral load as compared to the oral mucosa, cornea, conjunctiva, olfactory bulb, olfactory tubercle, trigeminal ganglion, medulla oblongata, and cerebellum [74]. Another recent study identified SARS-CoV-2 RNA in brush biopsy from the OE of COVID-19 patients. SARS-CoV-2 nucleoprotein was found mainly in sustentacular cells and immune cells. Occasional ORN infections were seen, but the number seemed insignificant compared to the uninfected control group [23]. In terms of pathology, human COVID-19 autopsies revealed focal atrophy, white blood cell infiltration in the olfactory mucosa, ORN axon damage, and inflammatory neuropathy of the olfactory nerve [74,75]. Strong cleaved caspase-3 signals indicative of cell apoptosis have also been observed in both infected and noninfected cells in the olfactory mucosa of anosmic COVID-19 patients [23]. Biopsy in a case of persistent anosmia 3 months after COVID-19 infection found massive disruption and desquamation of olfactory epithelium [22].
In golden Syrian hamster, massive OE damage and immune reactions were seen after nasal instillation of SARS-CoV-2, resulting in desquamation of the OE and infiltration of immune cells. A large number of sustentacular cells were infected. ORNs showed loss of cilia, but little or no evidence of viral infection [76,77,78,79]. In other studies, infection of some basal cells and occasional ORNs could not be ruled out [23,77]. In keeping with the findings from human autopsy and golden Syrian hamster model, SARS-CoV-2-infected ferrets also showed virus positivity in OE sustentacular cells and nasal respiratory epithelial cells [80].
Possible infection and pathology of other PNS structures in COVID-19 remain largely unexplored. The pathology of ageusia, for example, seems mostly unknown, although viral load in saliva has been found correlated with COVID-19 symptoms and loss of taste [81]. ACE2 was low or absent in taste receptor cells [7,82]. Noticeably, however, SARS-CoV-2 nucleoprotein has been detected in subsets of autopsy cranial nerves originating from the lower brainstem [83]. Diffused low ACE2 expression has been reported in some dorsal root ganglion neurons [84]. SARS-CoV-2 was rarely detected in the cerebrospinal fluid (CSF) of COVID-19 patients, including those with severe neurological manifestations or complications, but inflammatory or immunoreactive alterations seem common [9,85,86,87,88,89,90].
Regarding the CNS, brain autopsies in COVID-19 have generally revealed no SARS-CoV-2 in neurons or glia. PCR assays had found viral RNA in some brain tissue lysates, probably from virions or viral RNAs in infected blood or blood vessels of the brain [3,91,92,93]. Indeed, SARS-CoV-2 spike protein was detected in vascular endothelial cells and in microthrombi in COVID-19 brain autopsy samples including the OB, cerebellum, and brainstem [74]. Another autopsy study revealed occasional presence of viral N- or S-protein in individual cells of unknown identity in the CNS but found no direct relation of the cellular infection to major CNS pathological changes [83]. Pathological findings from COVID-19 autopsies include extensive inflammation, microglia activation, astrogliosis (especially in OB and medulla oblongata), perivascular infiltration of cytotoxic T lymphocytes or leukocytes, intravascular microthrombi [74,75,83,92], and hypoxia-associated alterations [93]. Brain imaging abnormalities, indicative of edema, injury, and microbleeding, have also been reported in the olfactory bulb of COVID-19 patients [94,95,96].
In experimental animals, irrespective of SARS-CoV-2 infection of the RE and OE, there has been no report of substantial invasion of the virus into the CNS neurons or glia (including the OB) [10,76,77,78,79,80,97,98,99], with a few exceptions (see below). SARS-CoV-2 nucleoprotein-positive myeloid cells were occasionally observed in the OB, but the exact identity (blood monocytes, macrophages, or CNS microglia) and locations (intravascular or extravascular) of these cells remained uncertain [23]. Likewise, although mostly undetectable in neurons or glia in the brain (including the OB), SARS-CoV-2 could sometimes be recovered from brain samples of infected animals, probably from infected blood or vascular endothelial cells [23,78]. Neuropathological alterations after SARS-CoV-2 infection of susceptible experimental animals ranged from absence of clear changes to inflammation, microglia activation, and infiltration of macrophages, similar to autopsy findings in human COVID-19 [76,77]. One exception is the K18-hACE2 transgenic mice that overexpress human ACE2 transgene (hACE2) under human K-18 promotor control and display unusually high sensitivity to SARS-CoV-2. Intranasal infection of K18-hACE2 transgenic mice could result in not only viral invasion of the OE, RE, and lungs, but also extensive virus spread into CNS regions such as the OB, anterior olfactory nucleus, thalamus, hypothalamus, and cerebral cortices [100,101]. In contrast, another line of transgenic mice that overexpresses hACE2 under the mouse ACE2 promotor control also suffers from SARS-CoV-2 infection and disease but did not show prominent virus spread to the CNS [102]. Even though seemingly unrepresentative, the K18-hACE2 transgenic mouse model appears suitable for therapeutic screening, as evidenced by the effectiveness of COVID-19 convalescent antisera in preventing disease or mortality by SARS-CoV-2 in these mice [101].
4. Olfactory Neuropathogenesis in COVID-19
4.1. Pathogenesis in the OE upon SARS-CoV-2 Infection
In summary, SARS-CoV-2 at the OE mainly infects the olfactory sustentacular cells (Figure 2A,B). Although OE horizontal basal cells have been shown to express moderate ACE2, these cells are normally not exposed to the nasal cavity and mucus, and thus might contract the virus only under special circumstances, such as compromise in OE integrity or secondary infection. OE desquamation, disruption, cell death, ORN cilia loss, and other pathological changes after SARS-CoV-2 infection, however, affect both ORNs and non-neuronal OE cells. Then, how are the mainly sustentacular cell infection and damages translated into ORN dysfunctions and OE pathology?
First, SARS-CoV-2-elicited sustentacular cell damages or death would compromise OE structural integrity, and significantly deprive the ORNs of the usual supports from non-neuronal, especially sustentacular, cells for structural stability, metabolism, homeostasis, and olfactory functions. The loss of supports might cause ORN injuries or even cell deaths. In case of infection and destruction of Bowman’s glands or ducts, OE mucus secretion would be adversely affected, and possible infection of OE basal cells or precursor ORNs may also hinder regeneration and functional recovery of the OE [31,103,104,105,106,107,108].
More importantly, infection of the OE would presumably mobilize immune reactions and activate inflammation as well as the release of specific cytokines or chemokines at the olfactory mucosa that could variably affect ORNs and other OE cells structurally or functionally. OE sustentacular cells are also phagocytic [105]. OE microvillar cells expressing transient receptor potential channel TRPM5 might have a role in neuroimmune detection or reactions [109]. A recent study has further demonstrated an ORN-mediated TrkA-dependent ultrarapid immune response to intranasal viral infection and OE damage in the rainbow trout [110]. Selective upregulation of interferon in the OE inhibits ORN odorant receptor protein expression and induces anosmia even without overt damage to the OE [111]. OE biopsy of COVID-19 patients showed significant increase in tumor necrosis factor alpha (TNF-α) but not IL-1β, as compared to levels in uninfected controls [112]. Transgenic overexpression of TNF-α is known to promote ORN cell death [6,113]. Interleukin 17c (IL17c) and its receptor are present in the mouse olfactory mucosa, and the former is markedly upregulated upon poly I:C intranasal instillation, mimicking viral infection [114].
Based on previous studies, it is also likely that pattern recognition receptors (PRRs) and related damage-associated molecular patterns (DAMPs) or pathogen-associated molecular patterns (PAMPs) play important roles in pathogenesis of SARS-CoV-2 in the OE and RE. PAMPs and DAMPs are involved in epithelial innate immunity and in pathogenesis of many acute and chronic inflammatory diseases. The single-pass transmembranous Toll-like receptors (TLRs), a type of PRRs, for example, recognize specific PAMPs, play important roles in innate immune reactions, and are expressed by neurons and glia of both the CNS and PNS [115]. TLR3, which detects double-stranded RNAs and activates NF-κB, has been shown to be preferentially expressed in mouse OE sustentacular cells [116]. Intranasal infusion of PAMPs and related mimetic molecules to activate TLRs would evoke neuroimmune or inflammatory responses [6,117,118], or protection of the OE from subsequent infection and the CNS from virus invasion [119]. It awaits future investigations to elucidate the involvement details of PRRs, PAMPs, and DAMPs in COVID-19-related olfactory dysfunctions and neuropathology.
In COVID-19 cases with obvious nasal congestion and rhinitis, obstructed airflow through the nasal cavity would also adversely affect olfaction and exacerbate ODs.
4.2. Pathogenesis in the OB and Other CNS Structures
The microglia activation, astrogliosis, inflammation, and immune reactions in the OB and related CNS regions after OE SARS-CoV-2 infection appear mostly elicited indirectly, rather than by the invasion of the virus itself [6,10]. One possible indirect pathogenesis pathway could be the anterograde and transsynaptic (transneuronal) degeneration of OB neural structures after damages or cell death of ORNs and olfactory nerve [120,121,122,123]. Moreover, even though axoplasmic transport of SARS-CoV-2 virus to the OB is rare, the situation seems different with regard to anterograde and transsynaptic transport of potentially pathogenic molecules and signals. SARS-CoV-2 spike protein cleavage peptide, for example, readily reaches the OB and other related CNS regions after intranasal instillation in the mice [73,124]. TLRs also participate in signaling between connected neurons in the olfactory system [119,125,126]. Finally, other CNS pathological changes, such as microvascular thrombosis, endothelium, and pericyte damages, microglia activation, and astrogliosis in the medulla oblongata, might have mainly or partially originated from the hematogenous route, and spread through the blood–brain barrier [74,75,83,92].
5. Persistent Anosmia, Hyposmia, or Parosmia
Most COVID-19 olfactory dysfunctions are transient, lasting for about 2–3 weeks. This is consistent with the fact that the OE undergoes regular ageing and self-replacement throughout life, and often readily repairs or regenerates itself upon damages [79,127,128,129]. In defiance of this well-known healing capability of the OE, however, a significant number of COVID-19 convalescents experience persistent ODs lasting for 12 months or longer [22,23,24,25]. The absent or exceptionally retarded recovery from COVID-19 ODs in those individuals implies a more severe or lasting damage to the OE by SARS-CoV-2. More specifically, this could result from SARS-CoV-2 infection or damage of the OE basal cells that express considerable ACE2 and TMPRSS2 [22,130]. Other possible causes of prolonged ODs after COVID-19 may pertain to persistent SARS-CoV-2 presence, chronic inflammation and immune reactions, or increased cell death in the OE. In COVID-19 convalescents with persistent anosmia, inflammation (as marked by infiltration of Iba1-positive myeloid cells), increased apoptosis (as marked by cleaved caspase 3-positive cells), and presence of SARS-CoV-2 (as marked by the viral nucleoprotein) could still be detected in the OE, but not in the RE, 6 months after the initial infection [23]. Interestingly, chronic inflammation could also modulate gene expression and switch the function of OE basal cells from neural regeneration to inflammatory signaling and immune cell proliferation [131].
6. Conclusions
SARS-CoV-2 has shown little neurotropism, apart from its high affinity to the neuron-supporting sustentacular cells of the OE. SARS-CoV-2 infection causes olfaction dysfunctions and ORN damages, most likely through indirect means such as deprivation of supports, inflammatory or immune reactions in the OE, and, to an extent, in the OB and other CNS regions (Figure 2B,C).
Apart from possible ORN anterograde degeneration and reactions affecting the OB and related CNS regions, SARS-CoV-2 infection and assaults on endothelial cells and pericytes of CNS vessels may cause microvascular thrombosis and leucocyte infiltration that, together with microglia activation, astrogliosis, inflammation, and immune responses, add to the pathogenic mechanisms for many of the COVID-19 neurological symptoms and complications (Figure 2D).
The causes and treatments of chronic SARS-CoV-2 infection of the OE and persistent post-COVID-19 ODs in a significant number of COVID-19 convalescents deserve further investigation.
Author Contributions
Conceptualization, F.L. and D.Y.W.; writing—original draft preparation, F.L.; writing—review and editing, F.L. and D.Y.W.; visualization, F.L. and D.Y.W.; funding acquisition, F.L. and D.Y.W. All authors have read and agreed to the published version of the manuscript.
Funding
The authors’ research is supported by academic research fund from the Ministry of Education, Singapore, grant number R181-000-182-114 (For FY Liang), and the Singapore National Medical Research Council, Singapore (NMRC COVID19RF2-0002) (For DY Wang).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Institutional Review Board of the National University of Singapore (protocol code: R18-0370, date of approval: 29 August 2018; protocol code: 699/04, date of approval: 31 January 2005).
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank Jennie Wong, Medical and Scientific Communication, Research Support Unit, Yong Loo Lin School of Medicine, National University Health System, for assistance in the language editing of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ellul, M.A.; Benjamin, L.; Singh, B.; Lant, S.; Michael, B.D.; Easton, A.; Kneen, R.; Defres, S.; Sejvar, J.; Solomon, T. Neurological Associations of COVID-19. Lancet Neurol. 2020, 19, 767–783. [Google Scholar] [CrossRef]
- Mao, L.; Jin, H.; Wang, M.; Hu, Y.; Chen, S.; He, Q.; Chang, J.; Hong, C.; Zhou, Y.; Wang, D.; et al. Neurologic Manifestations of Hospitalized Patients With Coronavirus Disease 2019 in Wuhan, China. JAMA Neurol. 2020, 77, 683. [Google Scholar] [CrossRef] [Green Version]
- Solomon, I.H.; Normandin, E.; Bhattacharyya, S.; Mukerji, S.S.; Keller, K.; Ali, A.S.; Adams, G.; Hornick, J.L.; Padera, R.F.; Sabeti, P. Neuropathological Features of Covid-19. N. Engl. J. Med. 2020, 383, 989–992. [Google Scholar] [CrossRef]
- Solomon, T. Neurological Infection with SARS-CoV-2—The Story so Far. Nat. Rev. Neurol. 2021, 17, 65–66. [Google Scholar] [CrossRef] [PubMed]
- Vellieux, G.; Sonneville, R.; Vledouts, S.; Jaquet, P.; Rouvel-Tallec, A.; d’Ortho, M.-P. COVID-19-Associated Neurological Manifestations: An Emerging Electroencephalographic Literature. Front. Physiol. 2021, 11, 622466. [Google Scholar] [CrossRef] [PubMed]
- Glezer, I.; Bruni-Cardoso, A.; Schechtman, D.; Malnic, B. Viral Infection and Smell Loss: The Case of COVID-19. J. Neurochem. 2021, 157, 930–943. [Google Scholar] [CrossRef] [PubMed]
- Cooper, K.W.; Brann, D.H.; Farruggia, M.C.; Bhutani, S.; Pellegrino, R.; Tsukahara, T.; Weinreb, C.; Joseph, P.V.; Larson, E.D.; Parma, V.; et al. COVID-19 and the Chemical Senses: Supporting Players Take Center Stage. Neuron 2020, 107, 219–233. [Google Scholar] [CrossRef] [PubMed]
- Singal, C.M.S.; Jaiswal, P.; Seth, P. SARS-CoV-2, More than a Respiratory Virus: Its Potential Role in Neuropathogenesis. ACS Chem. Neurosci. 2020, 11, 1887–1899. [Google Scholar] [CrossRef]
- Yazdanpanah, N.; Saghazadeh, A.; Rezaei, N. Anosmia: A Missing Link in the Neuroimmunology of Coronavirus Disease 2019 (COVID-19). Rev. Neurosci. 2020, 31, 691–701. [Google Scholar] [CrossRef]
- Butowt, R.; Meunier, N.; Bryche, B.; von Bartheld, C.S. The Olfactory Nerve Is Not a Likely Route to Brain Infection in COVID-19: A Critical Review of Data from Humans and Animal Models. Acta Neuropathol. 2021, 141, 809–822. [Google Scholar] [CrossRef]
- Boscolo-Rizzo, P.; Borsetto, D.; Fabbris, C.; Spinato, G.; Frezza, D.; Menegaldo, A.; Mularoni, F.; Gaudioso, P.; Cazzador, D.; Marciani, S.; et al. Evolution of Altered Sense of Smell or Taste in Patients With Mildly Symptomatic COVID-19. JAMA Otolaryngol. Neck Surg. 2020, 146, 729. [Google Scholar] [CrossRef] [PubMed]
- Lechien, J.R.; Chiesa-Estomba, C.M.; Hans, S.; Barillari, M.R.; Jouffe, L.; Saussez, S. Loss of Smell and Taste in 2013 European Patients With Mild to Moderate COVID-19. Ann. Intern. Med. 2020, 173, 672–675. [Google Scholar] [CrossRef] [PubMed]
- Mullol, J.; Alobid, I.; Mariño-Sánchez, F.; Izquierdo-Domínguez, A.; Marin, C.; Klimek, L.; Wang, D.-Y.; Liu, Z. The Loss of Smell and Taste in the COVID-19 Outbreak: A Tale of Many Countries. Curr. Allergy Asthma Rep. 2020, 20, 61. [Google Scholar] [CrossRef]
- Qiu, C.; Cui, C.; Hautefort, C.; Haehner, A.; Zhao, J.; Yao, Q.; Zeng, H.; Nisenbaum, E.J.; Liu, L.; Zhao, Y.; et al. Olfactory and Gustatory Dysfunction as an Early Identifier of COVID-19 in Adults and Children: An International Multicenter Study. Otolaryngol. Neck Surg. 2020, 163, 714–721. [Google Scholar] [CrossRef] [PubMed]
- Sedaghat, A.R.; Gengler, I.; Speth, M.M. Olfactory Dysfunction: A Highly Prevalent Symptom of COVID-19 With Public Health Significance. Otolaryngol. Neck Surg. 2020, 163, 12–15. [Google Scholar] [CrossRef]
- Tong, J.Y.; Wong, A.; Zhu, D.; Fastenberg, J.H.; Tham, T. The Prevalence of Olfactory and Gustatory Dysfunction in COVID-19 Patients: A Systematic Review and Meta-Analysis. Otolaryngol. Neck Surg. 2020, 163, 3–11. [Google Scholar] [CrossRef]
- Vaira, L.A.; Deiana, G.; Fois, A.G.; Pirina, P.; Madeddu, G.; De Vito, A.; Babudieri, S.; Petrocelli, M.; Serra, A.; Bussu, F.; et al. Objective Evaluation of Anosmia and Ageusia in COVID-19 Patients: Single-center Experience on 72 Cases. Head Neck 2020, 42, 1252–1258. [Google Scholar] [CrossRef] [PubMed]
- Wee, L.E.; Chan, Y.F.Z.; Teo, N.W.Y.; Cherng, B.P.Z.; Thien, S.Y.; Wong, H.M.; Wijaya, L.; Toh, S.T.; Tan, T.T. The Role of Self-Reported Olfactory and Gustatory Dysfunction as a Screening Criterion for Suspected COVID-19. Eur. Arch. Otorhinolaryngol. 2020, 277, 2389–2390. [Google Scholar] [CrossRef] [Green Version]
- Whitcroft, K.L.; Hummel, T. Olfactory Dysfunction in COVID-19: Diagnosis and Management. JAMA 2020, 323, 2512. [Google Scholar] [CrossRef]
- Hajare, P.S.; Harugop, A.S.; Goswami, L.; Padmavathy, O.; Aggarwal, U.; Reddy, Y.L. Prevalence of Olfactory and Gustatory Dysfunction in Coronavirus Disease (COVID-19): A Cross Sectional Study in Our Tertiary Care Hospital. Indian J. Otolaryngol. Head Neck Surg. 2021. Epub ahead of print. [Google Scholar] [CrossRef]
- Von Bartheld, C.S.; Hagen, M.M.; Butowt, R. The D614G Virus Mutation Enhances Anosmia in COVID-19 Patients: Evidence from a Systematic Review and Meta-Analysis of Studies from South Asia. ACS Chem. Neurosci. 2021, 12, 3535–3549. [Google Scholar] [CrossRef]
- Vaira, L.A.; Hopkins, C.; Sandison, A.; Manca, A.; Machouchas, N.; Turilli, D.; Lechien, J.R.; Barillari, M.R.; Salzano, G.; Cossu, A.; et al. Olfactory Epithelium Histopathological Findings in Long-Term Coronavirus Disease 2019 Related Anosmia. J. Laryngol. Otol. 2020, 134, 1123–1127. [Google Scholar] [CrossRef]
- de Melo, G.D.; Lazarini, F.; Levallois, S.; Hautefort, C.; Michel, V.; Larrous, F.; Verillaud, B.; Aparicio, C.; Wagner, S.; Gheusi, G.; et al. COVID-19–Related Anosmia Is Associated with Viral Persistence and Inflammation in Human Olfactory Epithelium and Brain Infection in Hamsters. Sci. Transl. Med. 2021, 13, eabf8396. [Google Scholar] [CrossRef]
- Boscolo-Rizzo, P.; Menegaldo, A.; Fabbris, C.; Spinato, G.; Borsetto, D.; Vaira, L.A.; Calvanese, L.; Pettorelli, A.; Sonego, M.; Frezza, D.; et al. Six-Month Psychophysical Evaluation of Olfactory Dysfunction in Patients with COVID-19. Chem. Senses 2021, 46, bjab006. [Google Scholar] [CrossRef]
- Boscolo-Rizzo, P.; Guida, F.; Polesel, J.; Marcuzzo, A.V.; Antonucci, P.; Capriotti, V.; Sacchet, E.; Cragnolini, F.; D’Alessandro, A.; Zanelli, E.; et al. Self-Reported Smell and Taste Recovery in Coronavirus Disease 2019 Patients: A One-Year Prospective Study. Eur. Arch. Otorhinolaryngol. 2021. Epub ahead of print. [Google Scholar] [CrossRef]
- Buck, L.B. Information Coding in the Vertebrate Olfactory System. Annu. Rev. Neurosci. 1996, 19, 517–544. [Google Scholar] [CrossRef]
- Buck, L.B. Unraveling the Sense of Smell (Nobel Lecture). Angew. Chem. Int. Ed. 2005, 44, 6128–6140. [Google Scholar] [CrossRef]
- Schwob, J.E. Neural Regeneration and the Peripheral Olfactory System. Anat. Rec. 2002, 269, 33–49. [Google Scholar] [CrossRef] [PubMed]
- Gray’s Anatomy: The Anatomical Basis of Clinical Practice, 40th ed.; Standring, S.; Gray, H. (Eds.) Churchill Livingstone Elsevier: Edinburgh, UK, 2009; ISBN 978-0-443-06684-9. [Google Scholar]
- Menco, B.P.M.; Morrison, E.E. Morphology of the Mammalian Olfactory Epithelium: Form, Fine Structure, Function, and Pathology. In Handbook of Olfaction and Gustation; Doty, R., Ed.; CRC Press: New York, NY, USA, 2003; pp. 17–49. ISBN 978-0-8247-0719-4. [Google Scholar]
- Liang, F. Sustentacular Cell Enwrapment of Olfactory Receptor Neuronal Dendrites: An Update. Genes 2020, 11, 493. [Google Scholar] [CrossRef] [PubMed]
- Mescher, A.L. The Respiratory System. In Junqueira’s Basic Histology, 14th ed.; McGraw-Hill Education: New York, NY, USA, 2016. [Google Scholar]
- Graziadei, P.P.C.; Graziadei, G.A.M. Neurogenesis and Neuron Regeneration in the Olfactory System of Mammals. I. Morphological Aspects of Differentiation and Structural Organization of the Olfactory Sensory Neurons. J. Neurocytol. 1979, 8, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Mori, I.; Nishiyama, Y.; Yokochi, T.; Kimura, Y. Olfactory Transmission of Neurotropic Viruses. J. Neurovirol. 2005, 11, 129–137. [Google Scholar] [CrossRef]
- Prediger, R.; Batista, L.; Medeiros, R.; Pandolfo, P.; Florio, J.; Takahashi, R. The Risk Is in the Air: Intranasal Administration of MPTP to Rats Reproducing Clinical Features of Parkinson’s Disease. Exp. Neurol. 2006, 202, 391–403. [Google Scholar] [CrossRef] [PubMed]
- Van Riel, D.; Verdijk, R.; Kuiken, T. The Olfactory Nerve: A Shortcut for Influenza and Other Viral Diseases into the Central Nervous System. J. Pathol. 2015, 235, 277–287. [Google Scholar] [CrossRef]
- Liu, H.; Lu, M.; Guthrie, K.M. Anterograde Trafficking of Neurotrophin-3 in the Adult Olfactory System in Vivo. Exp. Neurol. 2013, 241, 125–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemiale, F.; Kong, W.; Akyürek, L.M.; Ling, X.; Huang, Y.; Chakrabarti, B.K.; Eckhaus, M.; Nabel, G.J. Enhanced Mucosal Immunoglobulin A Response of Intranasal Adenoviral Vector Human Immunodeficiency Virus Vaccine and Localization in the Central Nervous System. J. Virol. 2003, 77, 10078–10087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanson, L.R.; Frey, W.H. Intranasal Delivery Bypasses the Blood-Brain Barrier to Target Therapeutic Agents to the Central Nervous System and Treat Neurodegenerative Disease. BMC Neurosci. 2008, 9, S5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Liu, Y.; Yang, N.; Wan, X.; Zuo, P. Nasal Administration of Cholera Toxin B Subunit–Nerve Growth Factor Improves the Space Learning and Memory Abilities in β-Amyloid Protein25-35-Induced Amnesic Mice. Neuroscience 2008, 155, 234–240. [Google Scholar] [CrossRef]
- Belur, L.R.; Temme, A.; Podetz-Pedersen, K.M.; Riedl, M.; Vulchanova, L.; Robinson, N.; Hanson, L.R.; Kozarsky, K.F.; Orchard, P.J.; Frey, W.H.; et al. Intranasal Adeno-Associated Virus Mediated Gene Delivery and Expression of Human Iduronidase in the Central Nervous System: A Noninvasive and Effective Approach for Prevention of Neurologic Disease in Mucopolysaccharidosis Type I. Hum. Gene Ther. 2017, 28, 576–587. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, B.; Hanson, L.R.; Frey, W.H., II. Intranasal Insulin: A Treatment Strategy for Addiction. Neurotherapeutics 2020, 17, 105–115. [Google Scholar] [CrossRef]
- Besser, G.; Liu, D.T.; Renner, B.; Hummel, T.; Mueller, C.A. Reversible Obstruction of the Olfactory Cleft: Impact on Olfactory Perception and Nasal Patency. Int. Forum Allergy Rhinol. 2020, 10, 713–718. [Google Scholar] [CrossRef] [PubMed]
- Trotier, D.; Bensimon, J.L.; Herman, P.; Tran Ba Huy, P.; Doving, K.B.; Eloit, C. Inflammatory Obstruction of the Olfactory Clefts and Olfactory Loss in Humans: A New Syndrome? Chem. Senses 2007, 32, 285–292. [Google Scholar] [CrossRef] [Green Version]
- Menni, C.; Valdes, A.M.; Freidin, M.B.; Sudre, C.H.; Nguyen, L.H.; Drew, D.A.; Ganesh, S.; Varsavsky, T.; Cardoso, M.J.; El-Sayed Moustafa, J.S.; et al. Real-Time Tracking of Self-Reported Symptoms to Predict Potential COVID-19. Nat. Med. 2020, 26, 1037–1040. [Google Scholar] [CrossRef]
- Fodoulian, L.; Tuberosa, J.; Rossier, D.; Boillat, M.; Kan, C.; Pauli, V.; Egervari, K.; Lobrinus, J.A.; Landis, B.N.; Carleton, A.; et al. SARS-CoV-2 Receptors and Entry Genes Are Expressed in the Human Olfactory Neuroepithelium and Brain. iScience 2020, 23, 101839. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Shen, W.; Rowan, N.R.; Kulaga, H.; Hillel, A.; Ramanathan, M.; Lane, A.P. Elevated ACE-2 Expression in the Olfactory Neuroepithelium: Implications for Anosmia and Upper Respiratory SARS-CoV-2 Entry and Replication. Eur. Respir. J. 2020, 56, 2001948. [Google Scholar] [CrossRef] [PubMed]
- Bilinska, K.; Jakubowska, P.; Von Bartheld, C.S.; Butowt, R. Expression of the SARS-CoV-2 Entry Proteins, ACE2 and TMPRSS2, in Cells of the Olfactory Epithelium: Identification of Cell Types and Trends with Age. ACS Chem. Neurosci. 2020, 11, 1555–1562. [Google Scholar] [CrossRef] [PubMed]
- Hermens, W.A.J.J.; Merkus, F.W.H.M. The Influence of Drugs on Nasal Ciliary Movement. Pharm. Res. 1987, 4, 445–449. [Google Scholar] [CrossRef]
- Yaghi, A.; Dolovich, M. Airway Epithelial Cell Cilia and Obstructive Lung Disease. Cells 2016, 5, 40. [Google Scholar] [CrossRef]
- Sauvanet, C.; Wayt, J.; Pelaseyed, T.; Bretscher, A. Structure, Regulation, and Functional Diversity of Microvilli on the Apical Domain of Epithelial Cells. Annu. Rev. Cell Dev. Biol. 2015, 31, 593–621. [Google Scholar] [CrossRef]
- Morrison, E.E.; Costanzo, R.M. Morphology of the Human Olfactory Epithelium. J. Comp. Neurol. 1990, 297, 1–13. [Google Scholar] [CrossRef]
- Morrison, E.E.; Costanzo, R.M. Morphology of Olfactory Epithelium in Humans and Other Vertebrates. Microsc. Res. Tech. 1992, 23, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Jafek, B.W. Biopsies of Human Olfactory Epithelium. Chem. Senses 2002, 27, 623–628. [Google Scholar] [CrossRef] [Green Version]
- Flink, R.; Westman, J. Different Neuron Populations in the Feline Lateral Cervical Nucleus: A Light and Electron Microscopic Study with the Retrograde Axonal Transport Technique. J. Comp. Neurol. 1986, 250, 265–281. [Google Scholar] [CrossRef] [PubMed]
- Braz, J.M.; Rico, B.; Basbaum, A.I. Transneuronal Tracing of Diverse CNS Circuits by Cre-Mediated Induction of Wheat Germ Agglutinin in Transgenic Mice. Proc. Natl. Acad. Sci. USA 2002, 99, 15148–15153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, K.-H.; Yao, G.; Jin, R. Diverse Binding Modes, Same Goal: The Receptor Recognition Mechanism of Botulinum Neurotoxin. Prog. Biophys. Mol. Biol. 2015, 117, 225–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerfen, C.R.; Sawchenko, P.E. An Anterograde Neuroanatomical Tracing Method That Shows the Detailed Morphology of Neurons, Their Axons and Terminals: Immunohistochemical Localization of an Axonally Transported Plant Lectin, Phaseolus Vulgaris Leucoagglutinin (PHA-L). Brain Res. 1984, 290, 219–238. [Google Scholar] [CrossRef] [Green Version]
- Desforges, M.; Le Coupanec, A.; Brison, É.; Meessen-Pinard, M.; Talbot, P.J. Neuroinvasive and Neurotropic Human Respiratory Coronaviruses: Potential Neurovirulent Agents in Humans. In Infectious Diseases and Nanomedicine I; Adhikari, R., Thapa, S., Eds.; Advances in Experimental Medicine and Biology; Springer: New Delhi, India, 2014; Volume 807, pp. 75–96. ISBN 978-81-322-1776-3. [Google Scholar]
- Schnell, M.J.; McGettigan, J.P.; Wirblich, C.; Papaneri, A. The Cell Biology of Rabies Virus: Using Stealth to Reach the Brain. Nat. Rev. Microbiol. 2010, 8, 51–61. [Google Scholar] [CrossRef]
- Richards, A.; Berth, S.H.; Brady, S.; Morfini, G. Engagement of Neurotropic Viruses in Fast Axonal Transport: Mechanisms, Potential Role of Host Kinases and Implications for Neuronal Dysfunction. Front. Cell. Neurosci. 2021, 15, 684762. [Google Scholar] [CrossRef] [PubMed]
- Trojanowski, J.Q.; Gonatas, J.O.; Gonatas, N.K. Conjugates of Horseradish Peroxidase (HRP) with Cholera Toxin and Wheat Germ Agglutinin Are Superior to Free HRP as Orthogradely Transported Markers. Brain Res. 1981, 223, 381–385. [Google Scholar] [CrossRef]
- Sun, N.; Cassell, M.D.; Perlman, S. Anterograde, Transneuronal Transport of Herpes Simplex Virus Type 1 Strain H129 in the Murine Visual System. J. Virol. 1996, 70, 5405–5413. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, N. Adenovirus-Mediated WGA Gene Delivery for Transsynaptic Labeling of Mouse Olfactory Pathways. Chem. Senses 2002, 27, 215–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ugolini, G. Rabies Virus as a Transneuronal Tracer of Neuronal Connections. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 2011; Volume 79, pp. 165–202. ISBN 978-0-12-387040-7. [Google Scholar]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, C.G.K.; Allon, S.J.; Nyquist, S.K.; Mbano, I.M.; Miao, V.N.; Tzouanas, C.N.; Cao, Y.; Yousif, A.S.; Bals, J.; Hauser, B.M.; et al. SARS-CoV-2 Receptor ACE2 Is an Interferon-Stimulated Gene in Human Airway Epithelial Cells and Is Detected in Specific Cell Subsets across Tissues. Cell 2020, 181, 1016–1035.e19. [Google Scholar] [CrossRef]
- Brann, D.H.; Tsukahara, T.; Weinreb, C.; Lipovsek, M.; Van den Berge, K.; Gong, B.; Chance, R.; Macaulay, I.C.; Chou, H.-J.; Fletcher, R.B.; et al. Non-Neuronal Expression of SARS-CoV-2 Entry Genes in the Olfactory System Suggests Mechanisms Underlying COVID-19-Associated Anosmia. Sci. Adv. 2020, 6, eabc5801. [Google Scholar] [CrossRef]
- Lee, I.T.; Nakayama, T.; Wu, C.-T.; Goltsev, Y.; Jiang, S.; Gall, P.A.; Liao, C.-K.; Shih, L.-C.; Schürch, C.M.; McIlwain, D.R.; et al. ACE2 Localizes to the Respiratory Cilia and Is Not Increased by ACE Inhibitors or ARBs. Nat. Commun. 2020, 11, 5453. [Google Scholar] [CrossRef]
- Hamming, I.; Timens, W.; Bulthuis, M.; Lely, A.; Navis, G.; van Goor, H. Tissue Distribution of ACE2 Protein, the Functional Receptor for SARS Coronavirus. A First Step in Understanding SARS Pathogenesis. J. Pathol. 2004, 203, 631–637. [Google Scholar] [CrossRef]
- Kehoe, P.G.; Wong, S.; AL Mulhim, N.; Palmer, L.E.; Miners, J.S. Angiotensin-Converting Enzyme 2 Is Reduced in Alzheimer’s Disease in Association with Increasing Amyloid-β and Tau Pathology. Alzheimers Res. Ther. 2016, 8, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torices, S.; Cabrera, R.; Stangis, M.; Naranjo, O.; Fattakhov, N.; Teglas, T.; Adesse, D.; Toborek, M. Expression of SARS-CoV-2-Related Receptors in Cells of the Neurovascular Unit: Implications for HIV-1 Infection. J. Neuroinflamm. 2021, 18, 167. [Google Scholar] [CrossRef]
- Cantuti-Castelvetri, L.; Ojha, R.; Pedro, L.D.; Djannatian, M.; Franz, J.; Kuivanen, S.; van der Meer, F.; Kallio, K.; Kaya, T.; Anastasina, M.; et al. Neuropilin-1 Facilitates SARS-CoV-2 Cell Entry and Infectivity. Science 2020, 370, 856–860. [Google Scholar] [CrossRef] [PubMed]
- Meinhardt, J.; Radke, J.; Dittmayer, C.; Franz, J.; Thomas, C.; Mothes, R.; Laue, M.; Schneider, J.; Brünink, S.; Greuel, S.; et al. Olfactory Transmucosal SARS-CoV-2 Invasion as a Port of Central Nervous System Entry in Individuals with COVID-19. Nat. Neurosci. 2021, 24, 168–175. [Google Scholar] [CrossRef]
- Kirschenbaum, D.; Imbach, L.L.; Ulrich, S.; Rushing, E.J.; Keller, E.; Reimann, R.R.; Frauenknecht, K.B.M.; Lichtblau, M.; Witt, M.; Hummel, T.; et al. Inflammatory Olfactory Neuropathy in Two Patients with COVID-19. Lancet 2020, 396, 166. [Google Scholar] [CrossRef]
- Bryche, B.; St Albin, A.; Murri, S.; Lacôte, S.; Pulido, C.; Ar Gouilh, M.; Lesellier, S.; Servat, A.; Wasniewski, M.; Picard-Meyer, E.; et al. Massive Transient Damage of the Olfactory Epithelium Associated with Infection of Sustentacular Cells by SARS-CoV-2 in Golden Syrian Hamsters. Brain. Behav. Immun. 2020, 89, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Sia, S.F.; Yan, L.-M.; Chin, A.W.H.; Fung, K.; Choy, K.-T.; Wong, A.Y.L.; Kaewpreedee, P.; Perera, R.A.P.M.; Poon, L.L.M.; Nicholls, J.M.; et al. Pathogenesis and Transmission of SARS-CoV-2 in Golden Hamsters. Nature 2020, 583, 834–838. [Google Scholar] [CrossRef]
- Imai, M.; Iwatsuki-Horimoto, K.; Hatta, M.; Loeber, S.; Halfmann, P.J.; Nakajima, N.; Watanabe, T.; Ujie, M.; Takahashi, K.; Ito, M.; et al. Syrian Hamsters as a Small Animal Model for SARS-CoV-2 Infection and Countermeasure Development. Proc. Natl. Acad. Sci. USA 2020, 117, 16587–16595. [Google Scholar] [CrossRef]
- Urata, S.; Maruyama, J.; Kishimoto-Urata, M.; Sattler, R.A.; Cook, R.; Lin, N.; Yamasoba, T.; Makishima, T.; Paessler, S. Regeneration Profiles of Olfactory Epithelium after SARS-CoV-2 Infection in Golden Syrian Hamsters. ACS Chem. Neurosci. 2021, 12, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Ryan, K.A.; Bewley, K.R.; Fotheringham, S.A.; Slack, G.S.; Brown, P.; Hall, Y.; Wand, N.I.; Marriott, A.C.; Cavell, B.E.; Tree, J.A.; et al. Dose-Dependent Response to Infection with SARS-CoV-2 in the Ferret Model and Evidence of Protective Immunity. Nat. Commun. 2021, 12, 81. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.; Pérez, P.; Kato, T.; Mikami, Y.; Okuda, K.; Gilmore, R.C.; Conde, C.D.; Gasmi, B.; Stein, S.; Beach, M.; et al. SARS-CoV-2 Infection of the Oral Cavity and Saliva. Nat. Med. 2021, 27, 892–903. [Google Scholar] [CrossRef]
- Wang, Z.; Zhou, J.; Marshall, B.; Rekaya, R.; Ye, K.; Liu, H.-X. SARS-CoV-2 Receptor ACE2 Is Enriched in a Subpopulation of Mouse Tongue Epithelial Cells in Nongustatory Papillae but Not in Taste Buds or Embryonic Oral Epithelium. ACS Pharmacol. Transl. Sci. 2020, 3, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Matschke, J.; Lütgehetmann, M.; Hagel, C.; Sperhake, J.P.; Schröder, A.S.; Edler, C.; Mushumba, H.; Fitzek, A.; Allweiss, L.; Dandri, M.; et al. Neuropathology of Patients with COVID-19 in Germany: A Post-Mortem Case Series. Lancet Neurol. 2020, 19, 919–929. [Google Scholar] [CrossRef]
- McFarland, A.J.; Yousuf, M.S.; Shiers, S.; Price, T.J. Neurobiology of SARS-CoV-2 Interactions with the Peripheral Nervous System: Implications for COVID-19 and Pain. PAIN Rep. 2021, 6, e885. [Google Scholar] [CrossRef]
- Helms, J.; Kremer, S.; Merdji, H.; Clere-Jehl, R.; Schenck, M.; Kummerlen, C.; Collange, O.; Boulay, C.; Fafi-Kremer, S.; Ohana, M.; et al. Neurologic Features in Severe SARS-CoV-2 Infection. N. Engl. J. Med. 2020, 382, 2268–2270. [Google Scholar] [CrossRef]
- Toscano, G.; Palmerini, F.; Ravaglia, S.; Ruiz, L.; Invernizzi, P.; Cuzzoni, M.G.; Franciotta, D.; Baldanti, F.; Daturi, R.; Postorino, P.; et al. Guillain–Barré Syndrome Associated with SARS-CoV-2. N. Engl. J. Med. 2020, 382, 2574–2576. [Google Scholar] [CrossRef]
- Ye, M.; Ren, Y.; Lv, T. Encephalitis as a Clinical Manifestation of COVID-19. Brain. Behav. Immun. 2020, 88, 945–946. [Google Scholar] [CrossRef]
- Lucchese, G. Cerebrospinal Fluid Findings in COVID-19 Indicate Autoimmunity. Lancet Microbe 2020, 1, e242. [Google Scholar] [CrossRef]
- Edén, A.; Kanberg, N.; Gostner, J.; Fuchs, D.; Hagberg, L.; Andersson, L.-M.; Lindh, M.; Price, R.W.; Zetterberg, H.; Gisslén, M. CSF Biomarkers in Patients with COVID-19 and Neurological Symptoms: A Case Series. Neurology 2020, 96, e294–e300. [Google Scholar] [CrossRef]
- Lewis, A.; Frontera, J.; Placantonakis, D.G.; Lighter, J.; Galetta, S.; Balcer, L.; Melmed, K.R. Cerebrospinal Fluid in COVID-19: A Systematic Review of the Literature. J. Neurol. Sci. 2021, 421, 117316. [Google Scholar] [CrossRef] [PubMed]
- Puelles, V.G.; Lütgehetmann, M.; Lindenmeyer, M.T.; Sperhake, J.P.; Wong, M.N.; Allweiss, L.; Chilla, S.; Heinemann, A.; Wanner, N.; Liu, S.; et al. Multiorgan and Renal Tropism of SARS-CoV-2. N. Engl. J. Med. 2020, 383, 590–592. [Google Scholar] [CrossRef] [PubMed]
- Schurink, B.; Roos, E.; Radonic, T.; Barbe, E.; Bouman, C.S.C.; de Boer, H.H.; de Bree, G.J.; Bulle, E.B.; Aronica, E.M.; Florquin, S.; et al. Viral Presence and Immunopathology in Patients with Lethal COVID-19: A Prospective Autopsy Cohort Study. Lancet Microbe 2020, 1, e290–e299. [Google Scholar] [CrossRef]
- Kantonen, J.; Mahzabin, S.; Mäyränpää, M.I.; Tynninen, O.; Paetau, A.; Andersson, N.; Sajantila, A.; Vapalahti, O.; Carpén, O.; Kekäläinen, E.; et al. Neuropathologic Features of Four Autopsied COVID-19 Patients. Brain Pathol. 2020, 30, 1012–1016. [Google Scholar] [CrossRef] [PubMed]
- Aragão, M.F.V.V.; Leal, M.C.; Cartaxo Filho, O.Q.; Fonseca, T.M.; Valença, M.M. Anosmia in COVID-19 Associated with Injury to the Olfactory Bulbs Evident on MRI. Am. J. Neuroradiol. 2020, 41, 1703–1706. [Google Scholar] [CrossRef]
- Laurendon, T.; Radulesco, T.; Mugnier, J.; Gérault, M.; Chagnaud, C.; El Ahmadi, A.-A.; Varoquaux, A. Bilateral Transient Olfactory Bulb Edema during COVID-19–Related Anosmia. Neurology 2020, 95, 224–225. [Google Scholar] [CrossRef]
- Kandemirli, S.G.; Altundag, A.; Yildirim, D.; Tekcan Sanli, D.E.; Saatci, O. Olfactory Bulb MRI and Paranasal Sinus CT Findings in Persistent COVID-19 Anosmia. Acad. Radiol. 2021, 28, 28–35. [Google Scholar] [CrossRef]
- Sit, T.H.C.; Brackman, C.J.; Ip, S.M.; Tam, K.W.S.; Law, P.Y.T.; To, E.M.W.; Yu, V.Y.T.; Sims, L.D.; Tsang, D.N.C.; Chu, D.K.W.; et al. Infection of Dogs with SARS-CoV-2. Nature 2020, 586, 776–778. [Google Scholar] [CrossRef] [PubMed]
- Munster, V.J.; Feldmann, F.; Williamson, B.N.; van Doremalen, N.; Pérez-Pérez, L.; Schulz, J.; Meade-White, K.; Okumura, A.; Callison, J.; Brumbaugh, B.; et al. Respiratory Disease in Rhesus Macaques Inoculated with SARS-CoV-2. Nature 2020, 585, 268–272. [Google Scholar] [CrossRef]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z.; et al. Susceptibility of Ferrets, Cats, Dogs, and Other Domesticated Animals to SARS–Coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumari, P.; Rothan, H.A.; Natekar, J.P.; Stone, S.; Pathak, H.; Strate, P.G.; Arora, K.; Brinton, M.A.; Kumar, M. Neuroinvasion and Encephalitis Following Intranasal Inoculation of SARS-CoV-2 in K18-HACE2 Mice. Viruses 2021, 13, 132. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wong, L.-Y.R.; Li, K.; Verma, A.K.; Ortiz, M.E.; Wohlford-Lenane, C.; Leidinger, M.R.; Knudson, C.M.; Meyerholz, D.K.; McCray, P.B.; et al. COVID-19 Treatments and Pathogenesis Including Anosmia in K18-HACE2 Mice. Nature 2021, 589, 603–607. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Deng, W.; Huang, B.; Gao, H.; Liu, J.; Ren, L.; Wei, Q.; Yu, P.; Xu, Y.; Qi, F.; et al. The Pathogenicity of SARS-CoV-2 in HACE2 Transgenic Mice. Nature 2020, 583, 830–833. [Google Scholar] [CrossRef]
- Okano, M.; Takagi, S.F. Secretion and Electrogenesis of the Supporting Cell in the Olfactory Epithelium. J. Physiol. 1974, 242, 353–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Getchell, M.L.; Ding, X.; Getchell, T.V. Immunolocalization of Two Cytochrome P450 Isozymes in Rat Nasal Chemosensory Tissue. NeuroReport 1992, 3, 749–752. [Google Scholar] [CrossRef]
- Suzuki, Y.; Takeda, M.; Farbman, A.I. Supporting Cells as Phagocytes in the Olfactory Epithelium after Bulbectomy. J. Comp. Neurol. 1996, 376, 509–517. [Google Scholar] [CrossRef]
- Suzuki, Y.; Takeda, M.; Obara, N.; Suzuki, N.; Takeichi, N. Olfactory Epithelium Consisting of Supporting Cells and Horizontal Basal Cells in the Posterior Nasal Cavity of Mice. Cell Tissue Res. 2000, 299, 313–325. [Google Scholar] [CrossRef]
- Hegg, C.C.; Irwin, M.; Lucero, M.T. Calcium Store-Mediated Signaling in Sustentacular Cells of the Mouse Olfactory Epithelium. Glia 2009, 57, 634–644. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Tang, J.; Ling, E.-A.; Wu, Y.; Liang, F. Juxtanodin in the Rat Olfactory Epithelium: Specific Expression in Sustentacular Cells and Preferential Subcellular Positioning at the Apical Junctional Belt. Neuroscience 2009, 161, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Genovese, F.; Tizzano, M. Microvillous Cells in the Olfactory Epithelium Express Elements of the Solitary Chemosensory Cell Transduction Signaling Cascade. PLoS ONE 2018, 13, e0202754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sepahi, A.; Kraus, A.; Casadei, E.; Johnston, C.A.; Galindo-Villegas, J.; Kelly, C.; García-Moreno, D.; Muñoz, P.; Mulero, V.; Huertas, M.; et al. Olfactory Sensory Neurons Mediate Ultrarapid Antiviral Immune Responses in a TrkA-Dependent Manner. Proc. Natl. Acad. Sci. USA 2019, 116, 12428–12436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, S.; Cao, L.; Rickenbacher, G.T.; Benz, E.G.; Magdamo, C.; Gomez, L.R.; Holbrook, E.H.; Albers, A.D.; Gallagher, R.; Westover, M.B.; et al. Innate Immune Signaling in the Olfactory Epithelium Reduces Odorant Receptor Levels: Modeling Transient Smell Loss in COVID-19 Patients. medRxiv 2020. [Google Scholar] [CrossRef]
- Torabi, A.; Mohammadbagheri, E.; Akbari Dilmaghani, N.; Bayat, A.-H.; Fathi, M.; Vakili, K.; Alizadeh, R.; Rezaeimirghaed, O.; Hajiesmaeili, M.; Ramezani, M.; et al. Proinflammatory Cytokines in the Olfactory Mucosa Result in COVID-19 Induced Anosmia. ACS Chem. Neurosci. 2020, 11, 1909–1913. [Google Scholar] [CrossRef]
- Lane, A.P.; Turner, J.; May, L.; Reed, R. A Genetic Model of Chronic Rhinosinusitis-Associated Olfactory Inflammation Reveals Reversible Functional Impairment and Dramatic Neuroepithelial Reorganization. J. Neurosci. 2010, 30, 2324–2329. [Google Scholar] [CrossRef]
- Bryche, B.; Dewaele, A.; Saint-Albin, A.; Le Poupon Schlegel, C.; Congar, P.; Meunier, N. IL-17c Is Involved in Olfactory Mucosa Responses to Poly(I:C) Mimicking Virus Presence. Brain. Behav. Immun. 2019, 79, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Rietdijk, C.D.; Van Wezel, R.J.A.; Garssen, J.; Kraneveld, A.D. Neuronal Toll-like Receptors and Neuro-Immunity in Parkinson’s Disease, Alzheimer’s Disease and Stroke. Neuroimmunol. Neuroinflamm. 2016, 3, 27. [Google Scholar] [CrossRef] [Green Version]
- Kanaya, K.; Kondo, K.; Suzukawa, K.; Sakamoto, T.; Kikuta, S.; Okada, K.; Yamasoba, T. Innate Immune Responses and Neuroepithelial Degeneration and Regeneration in the Mouse Olfactory Mucosa Induced by Intranasal Administration of Poly(I:C). Cell Tissue Res. 2014, 357, 279–299. [Google Scholar] [CrossRef] [Green Version]
- Crisafulli, U.; Xavier, A.M.; dos Santos, F.B.; Cambiaghi, T.D.; Chang, S.Y.; Porcionatto, M.; Castilho, B.A.; Malnic, B.; Glezer, I. Topical Dexamethasone Administration Impairs Protein Synthesis and Neuronal Regeneration in the Olfactory Epithelium. Front. Mol. Neurosci. 2018, 11, 50. [Google Scholar] [CrossRef] [Green Version]
- Imamura, F.; Hasegawa-Ishii, S. Environmental Toxicants-Induced Immune Responses in the Olfactory Mucosa. Front. Immunol. 2016, 7, 475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boivin, N.; Sergerie, Y.; Rivest, S.; Boivin, G. Effect of Pretreatment with Toll-like Receptor Agonists in a Mouse Model of Herpes Simplex Virus Type 1 Encephalitis. J. Infect. Dis. 2008, 198, 664–672. [Google Scholar] [CrossRef] [PubMed]
- Okuyama-Yamamoto, A.; Yamamoto, T.; Miki, A.; Terashima, T. Changes in Reelin Expression in the Mouse Olfactory Bulb after Chemical Lesion to the Olfactory Epithelium. Eur. J. Neurosci. 2005, 21, 2586–2592. [Google Scholar] [CrossRef]
- Maor-Nof, M.; Romi, E.; Sar Shalom, H.; Ulisse, V.; Raanan, C.; Nof, A.; Leshkowitz, D.; Lang, R.; Yaron, A. Axonal Degeneration Is Regulated by a Transcriptional Program That Coordinates Expression of Pro- and Anti-Degenerative Factors. Neuron 2016, 92, 991–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.; Chitranshi, N.; Wall, R.V.; Basavarajappa, D.; Gupta, V.; Mirzaei, M.; Graham, S.L.; Klistorner, A.; You, Y. Trans-Synaptic Degeneration in the Visual Pathway: Neural Connectivity, Pathophysiology, and Clinical Implications in Neurodegenerative Disorders. Surv. Ophthalmol. 2021, 66, S0039625721001387. [Google Scholar] [CrossRef]
- Simon, D.J.; Belsky, D.M.; Bowen, M.E.; Ohn, C.Y.J.; O’Rourke, M.K.; Shen, R.; Kim, G.; Pitts, J.; Attardi, L.D.; Tessier-Lavigne, M. An Anterograde Pathway for Sensory Axon Degeneration Gated by a Cytoplasmic Action of the Transcriptional Regulator P53. Dev. Cell 2021, 56, 976–984.e3. [Google Scholar] [CrossRef]
- Teesalu, T.; Sugahara, K.N.; Kotamraju, V.R.; Ruoslahti, E. C-End Rule Peptides Mediate Neuropilin-1-Dependent Cell, Vascular, and Tissue Penetration. Proc. Natl. Acad. Sci. USA 2009, 106, 16157–16162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, A.; Hong, W.; Favaloro, V.; Luo, L. Toll Receptors Instruct Axon and Dendrite Targeting and Participate in Synaptic Partner Matching in a Drosophila Olfactory Circuit. Neuron 2015, 85, 1013–1028. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa-Ishii, S.; Shimada, A.; Imamura, F. Lipopolysaccharide-Initiated Persistent Rhinitis Causes Gliosis and Synaptic Loss in the Olfactory Bulb. Sci. Rep. 2017, 7, 11605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graziadei, G.A.M.; Graziadei, P.P.C. Neurogenesis and Neuron Regeneration in the Olfactory System of Mammals. II. Degeneration and Reconstitution of the Olfactory Sensory Neurons after Axotomy. J. Neurocytol. 1979, 8, 197–213. [Google Scholar] [CrossRef] [PubMed]
- Herzog, C.; Otto, T. Regeneration of Olfactory Receptor Neurons Following Chemical Lesion: Time Course and Enhancement with Growth Factor Administration. Brain Res. 1999, 849, 155–161. [Google Scholar] [CrossRef]
- Brann, J.H.; Firestein, S.J. A Lifetime of Neurogenesis in the Olfactory System. Front. Neurosci. 2014, 8. [Google Scholar] [CrossRef]
- Gallo, O.; Locatello, L.G.; Mazzoni, A.; Novelli, L.; Annunziato, F. The Central Role of the Nasal Microenvironment in the Transmission, Modulation, and Clinical Progression of SARS-CoV-2 Infection. Mucosal Immunol. 2021, 14, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Reed, R.R.; Lane, A.P. Chronic Inflammation Directs an Olfactory Stem Cell Functional Switch from Neuroregeneration to Immune Defense. Cell Stem Cell 2019, 25, 501–513.e5. [Google Scholar] [CrossRef]
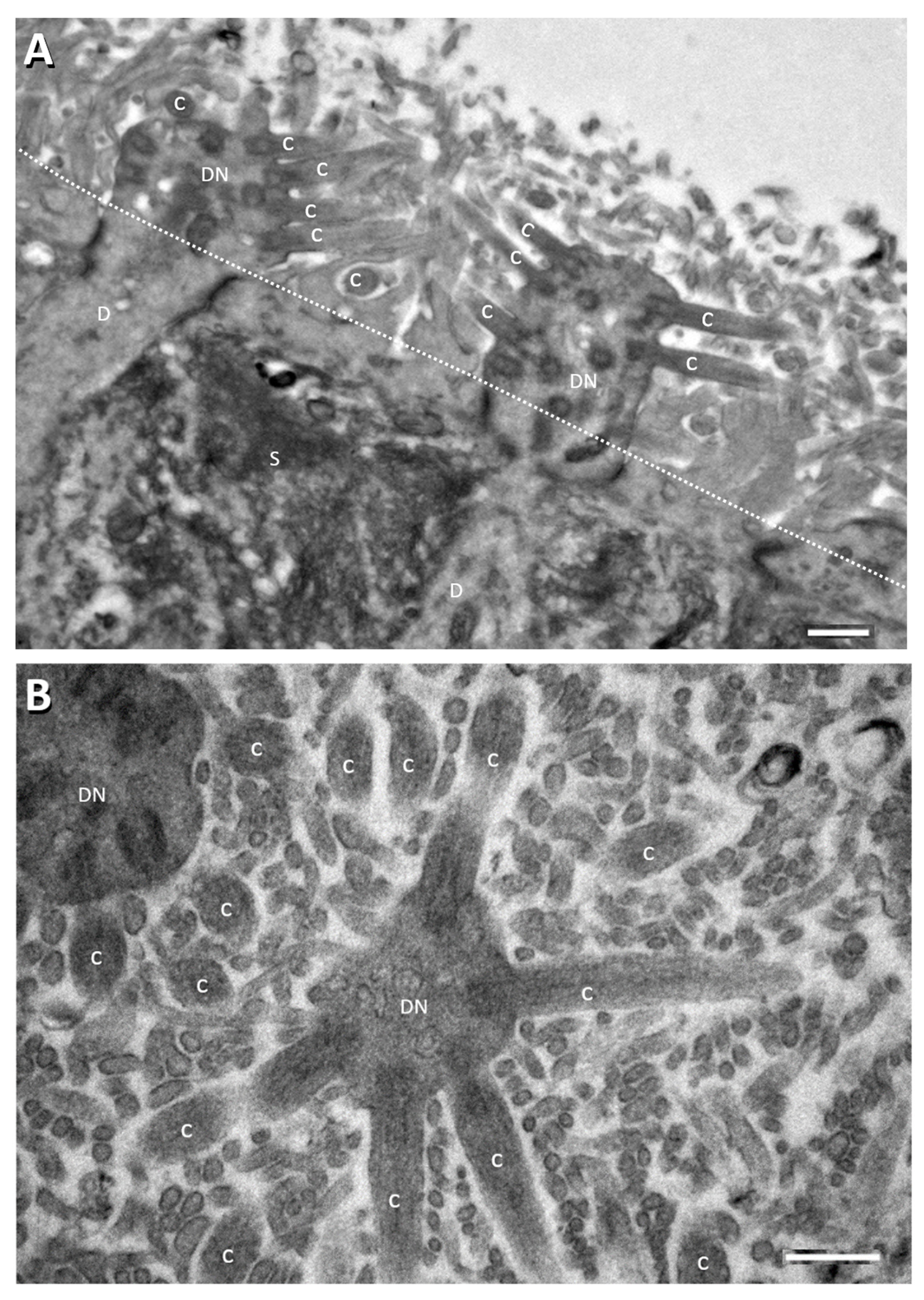
Figure 1.
Electron micrographs showing perpendicular (A) and tangential/oblique section (B) of the apical part of the rat OE. Dotted line in panel A denotes sustentacular cell (S) apical surface from which the long thin sustentacular-cell microvilli protrude into the nasal cavity for about 2–3 µm. ORN dendritic knobs (DN) and cilia (C) at apical ends of ORN dendrites (D) are mostly found among the sustentacular microvilli (most of the unlabeled small profile structures in (B) and in area above the dotted line in (A). Human OE is similarly organized [52,53,54]. Scale bars = 0.5 µm.
Figure 1.
Electron micrographs showing perpendicular (A) and tangential/oblique section (B) of the apical part of the rat OE. Dotted line in panel A denotes sustentacular cell (S) apical surface from which the long thin sustentacular-cell microvilli protrude into the nasal cavity for about 2–3 µm. ORN dendritic knobs (DN) and cilia (C) at apical ends of ORN dendrites (D) are mostly found among the sustentacular microvilli (most of the unlabeled small profile structures in (B) and in area above the dotted line in (A). Human OE is similarly organized [52,53,54]. Scale bars = 0.5 µm.

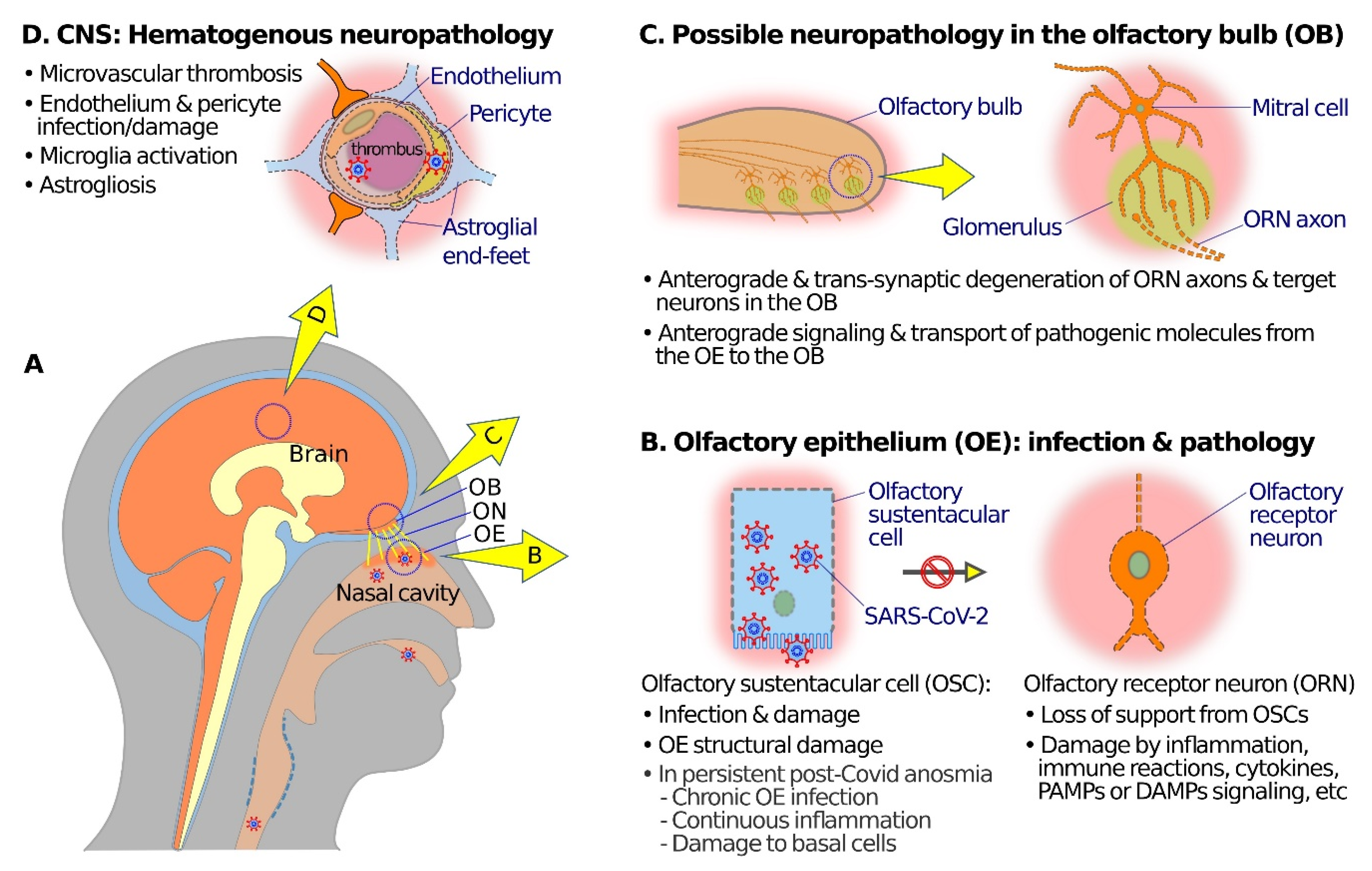
Figure 2.
Schematic diagrams showing possible mechanisms of olfactory neuropathogenesis in COVID-19. (A) A schematic overview to illustrate relations among nasal cavity, olfactory epithelium (OE), olfactory nerve (ON), olfactory bulb (OB), and the brain. (B) At the OE, SARS-CoV-2 mainly infects olfactory sustentacular cells (OSCs) that express high levels of SARS-CoV-2 receptor ACE2 on the luminal surface. Sustentacular cell infection and damage may lead to inflammation, immune reactions, release of cytokines, and signaling through pathogen-associated molecular patterns (PAMPs), damage-associated molecular patterns (DAMPs), and pattern recognition receptors (PRRs) which in turn may cause dysfunctions (such as anosmia or hyposmia) and damage and/or anterograde degeneration of olfactory receptor neuronal cells (ORNs). In the case of post-COVID-19 persistent olfactory dysfunctions, pathogenic mechanisms may include damage of basal cells, continuous inflammation, or chronic SARS-CoV-2 infection in the OE. (C) Anterograde degeneration, signaling, and transport of pathogenic molecules from the OE to the OB along ORN axons may result in dysfunction and transsynaptic degeneration of neural structures in the OB. (D) SARS-CoV-2 infection of endothelial cells or pericytes, and microthrombi in capillary blood vessels, may compromise the blood–brain barrier, and give rise to hematogenous neuropathology and dysfunctions in various brain regions, including the OB.
Figure 2.
Schematic diagrams showing possible mechanisms of olfactory neuropathogenesis in COVID-19. (A) A schematic overview to illustrate relations among nasal cavity, olfactory epithelium (OE), olfactory nerve (ON), olfactory bulb (OB), and the brain. (B) At the OE, SARS-CoV-2 mainly infects olfactory sustentacular cells (OSCs) that express high levels of SARS-CoV-2 receptor ACE2 on the luminal surface. Sustentacular cell infection and damage may lead to inflammation, immune reactions, release of cytokines, and signaling through pathogen-associated molecular patterns (PAMPs), damage-associated molecular patterns (DAMPs), and pattern recognition receptors (PRRs) which in turn may cause dysfunctions (such as anosmia or hyposmia) and damage and/or anterograde degeneration of olfactory receptor neuronal cells (ORNs). In the case of post-COVID-19 persistent olfactory dysfunctions, pathogenic mechanisms may include damage of basal cells, continuous inflammation, or chronic SARS-CoV-2 infection in the OE. (C) Anterograde degeneration, signaling, and transport of pathogenic molecules from the OE to the OB along ORN axons may result in dysfunction and transsynaptic degeneration of neural structures in the OB. (D) SARS-CoV-2 infection of endothelial cells or pericytes, and microthrombi in capillary blood vessels, may compromise the blood–brain barrier, and give rise to hematogenous neuropathology and dysfunctions in various brain regions, including the OB.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liang, F.; Wang, D.Y. COVID-19 Anosmia: High Prevalence, Plural Neuropathogenic Mechanisms, and Scarce Neurotropism of SARS-CoV-2? Viruses 2021, 13, 2225. https://doi.org/10.3390/v13112225
AMA Style
Liang F, Wang DY. COVID-19 Anosmia: High Prevalence, Plural Neuropathogenic Mechanisms, and Scarce Neurotropism of SARS-CoV-2? Viruses. 2021; 13(11):2225. https://doi.org/10.3390/v13112225
Chicago/Turabian StyleLiang, Fengyi, and De Yun Wang. 2021. "COVID-19 Anosmia: High Prevalence, Plural Neuropathogenic Mechanisms, and Scarce Neurotropism of SARS-CoV-2?" Viruses 13, no. 11: 2225. https://doi.org/10.3390/v13112225
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.