
Comparative Genomic Analysis of Classical and Variant Virulent Parental/Attenuated Strains of Porcine Epidemic Diarrhea Virus

Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Isolation and Confirmation
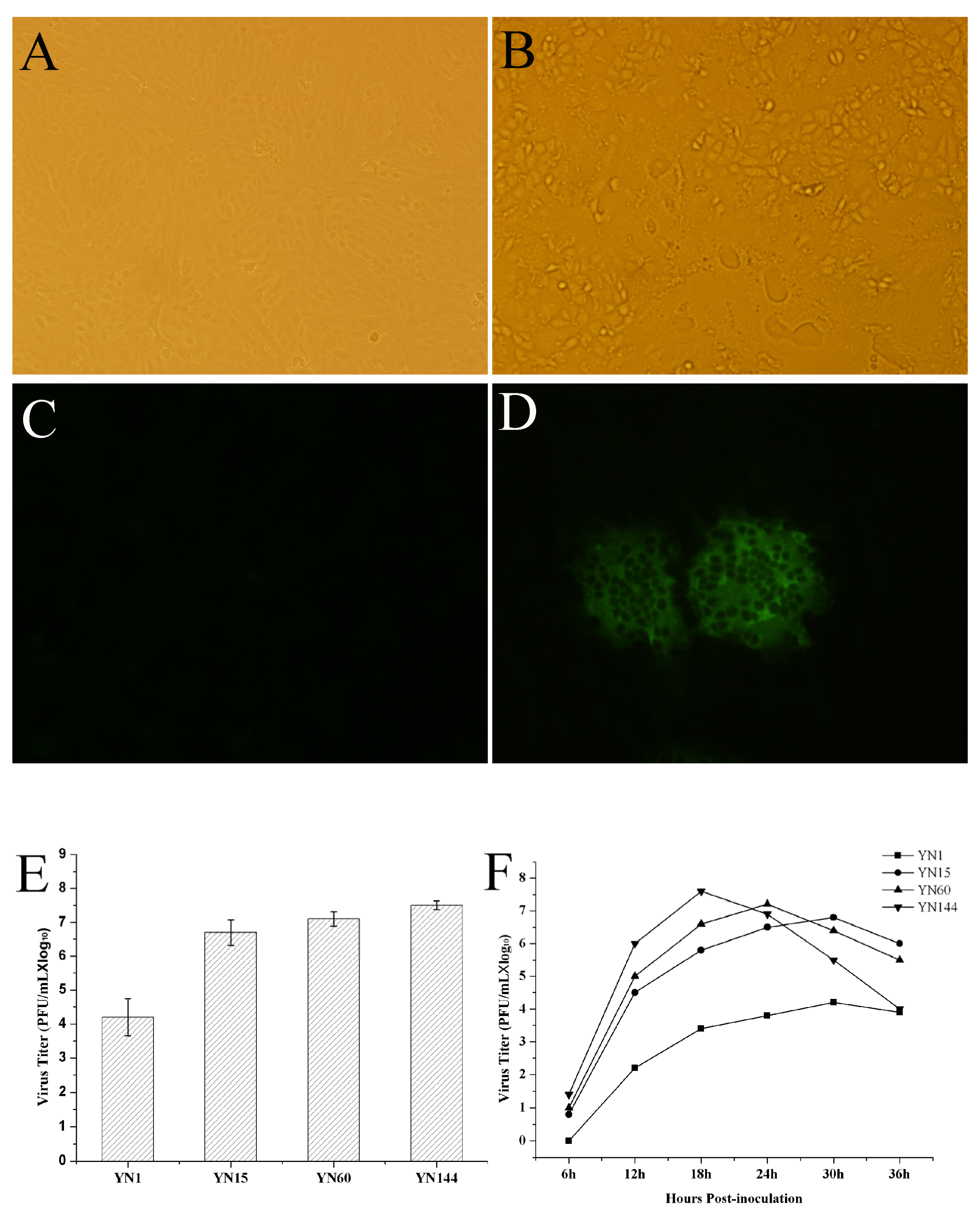
2.2. Serial Passage of PEDV YN1 Strain, Virus Titration, and Virus Growth Characteristic of YN1, YN15, YN60 and YN144
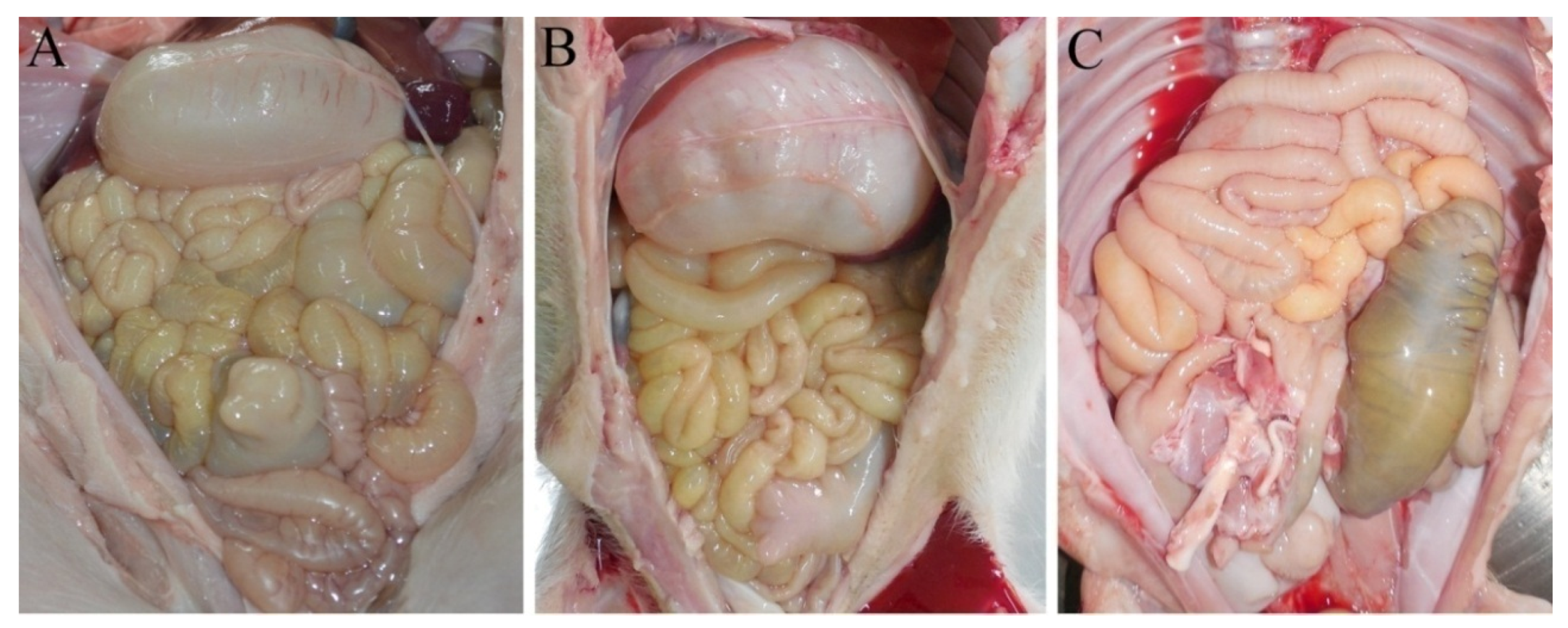
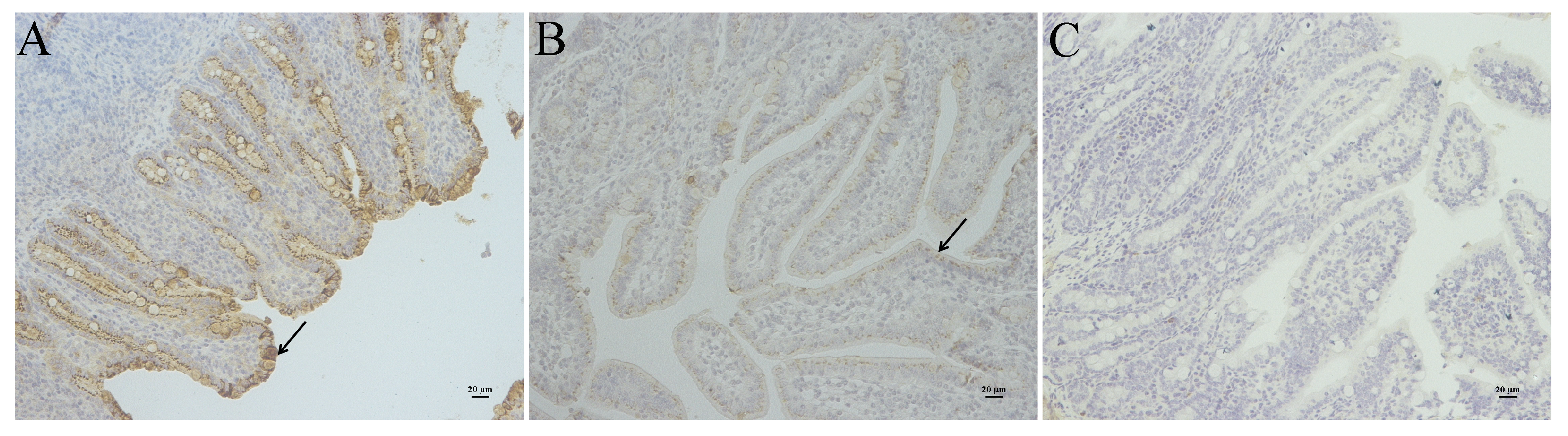
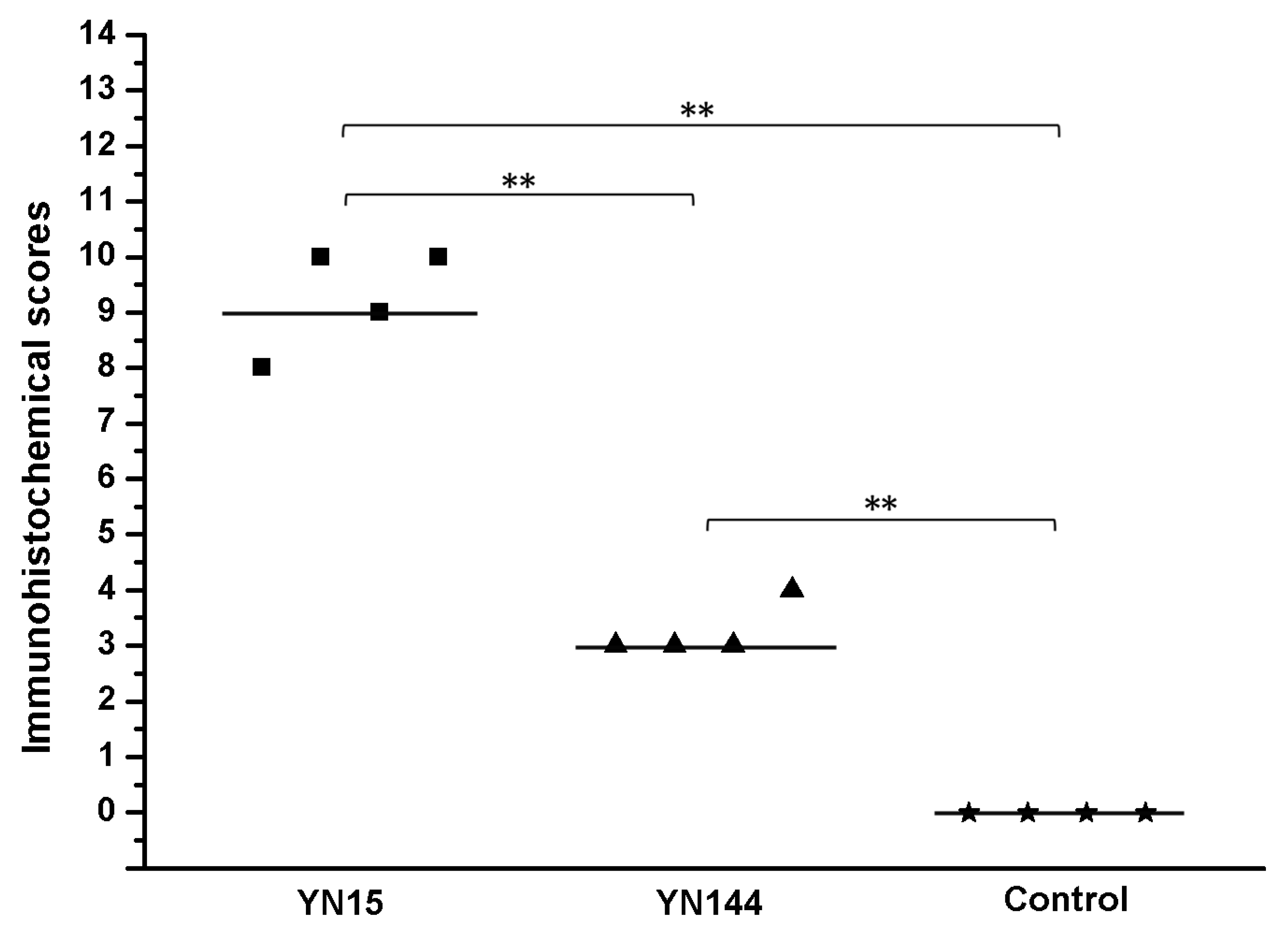
2.3. The Virulence Experiment of YN15 and YN144
2.4. Genome Sequencing
2.5. Multiple Alignment and Comparative Analysis
2.6. Statistical Analysis
3. Results
3.1. Virus Isolation, Identification and Virus Growth Characteristic of YN1, YN15, YN60 and YN144

3.2. Pathogenicity Analysis of YN15 and YN144


3.3. Amino Acid Changes during Passage


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | aa position | 1 | 15 | 30 | 60 | 90 | 144 | 200 |
|---|---|---|---|---|---|---|---|---|
| ORF1a/b | 509 | P | P | P | P | P | S | S |
| 1566 | I | I | T | T | T | T | T | |
| 2436 | L | L | L | L | L | L | F | |
| 2925 | H | H | H | H | H | Y | Y | |
| 3505 | V | V | V | V | V | E | E | |
| 4832 | V | V | V | V | F | F | F | |
| 5951 | A | A | A | A | A | A | P | |
| 6136 | L | F | F | F | F | F | F | |
| 6137 | V | S | S | S | S | S | S | |
| S | 144 | T | T | - | - | - | - | - |
| 145 | G | G | - | - | - | - | - | |
| 372 | Y | Y | Y | Y | Y | Y | H | |
| 405 | D | D | D | D | G | G | G | |
| 428 | G | G | G | A | A | A | A | |
| 490 | R | T | T | T | T | T | T | |
| 722 | S | S | S | S | S | R | R | |
| 779 | T | T | T | T | N | N | N | |
| 825 | Q | Q | H | H | H | H | H | |
| 887 | S | S | S | S | S | S | R | |
| 968 | S | A | A | A | A | A | A | |
| 1045 | H | Q | Q | Q | Q | Q | Q | |
| 1165 | D | N | N | N | N | N | N | |
| 1197 | H | - | - | - | - | - | - | |
| 1210 | F | Y | Y | Y | Y | Y | Y | |
| 1218 | S | G | G | G | G | G | G | |
| 1304 | I | I | I | I | I | L | L | |
| 1354 | C | C | C | C | C | F | F | |
| ORF3 | 138–141 | YYDG | FMTA | FMTA | FMTA | FMTA | FMTA | FMTA |
| 142–145 | KSIV | NPL * | NPL * | NPL * | NPL * | NPL * | NPL * | |
| E | 53 | I | I | I | I | V | V | V |
| 70 | P | P | P | S | S | S | S | |
| M | 187 | V | V | V | V | V | F | F |
| 214 | A | S | S | S | S | S | S | |
| 226 | V | V | V | V | V | V | F | |
| N | 112 | V | V | V | V | V | F | F |
| 379 | V | V | V | V | V | A | A | |
| 391 | S | S | R | R | R | S | S |
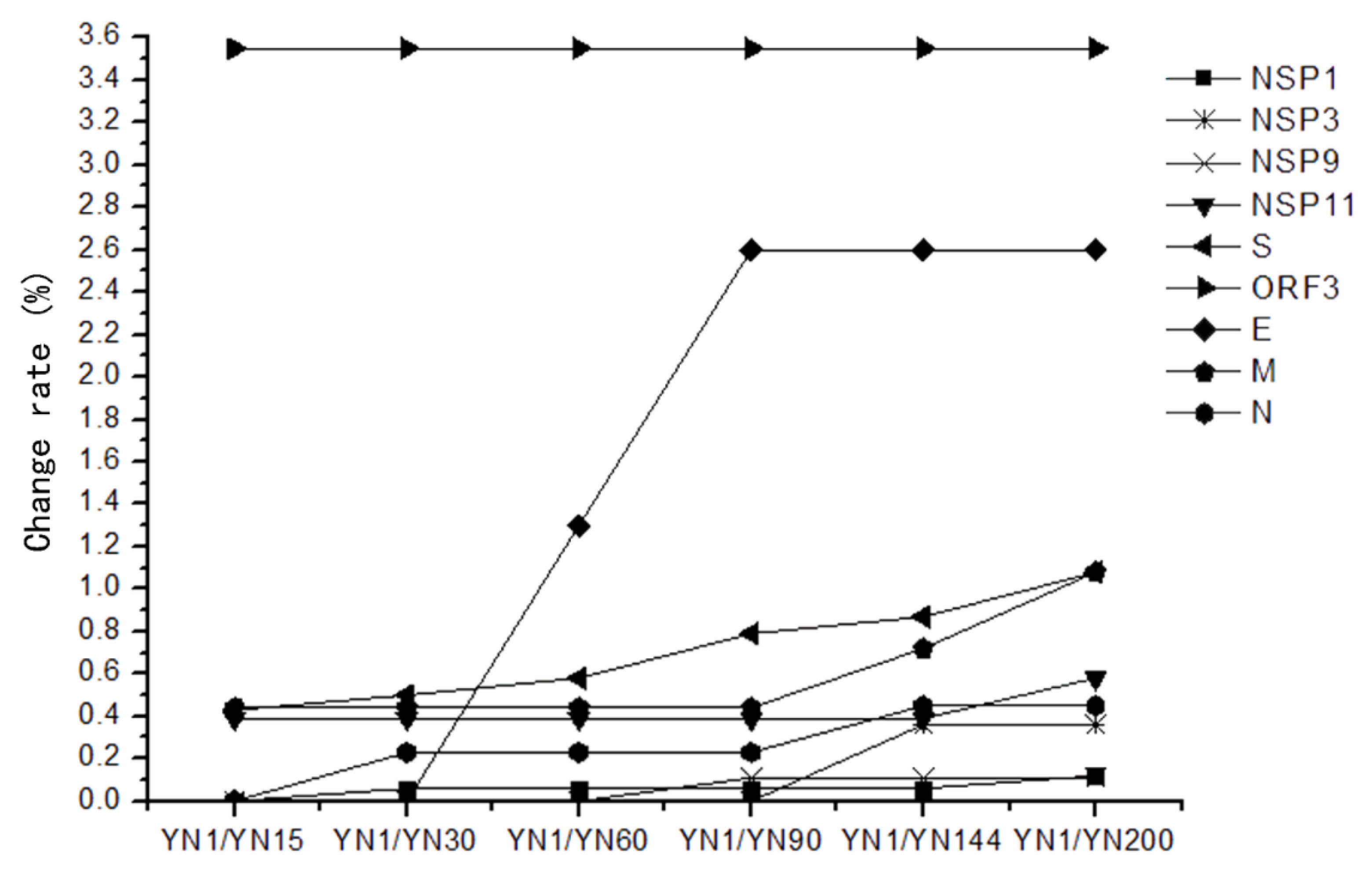
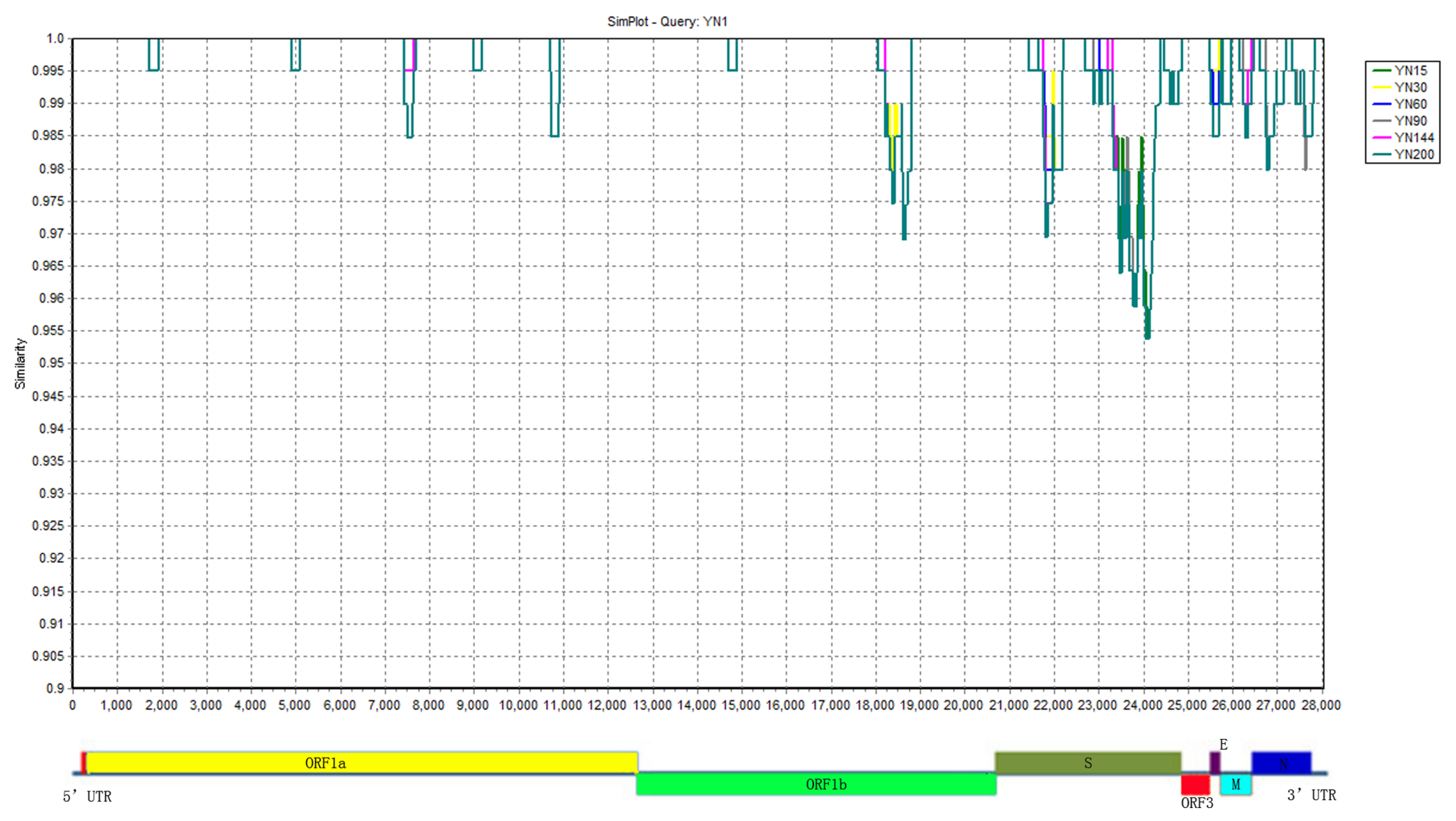
3.4. Sequence Homology and Variation Characteristic of strainsYN1, YN15, YN30, YN60, YN120, YN144 and YN200
| ORFs | Encoded Proteins | 1/15 | 1/30 | 1/60 | 1/90 | 1/144 | 1/200 |
|---|---|---|---|---|---|---|---|
| ORF1a/b | NSP1 | 0(0) * | 1(0.06) | 1(0.06) | 1(0.06) | 1(0.06) | 2(0.12) |
| NSP3 | 0(0) | 0(0) | 0(0) | 0(0) | 1(0.36) | 1(0.36) | |
| NSP9 | 0(0) | 0(0) | 0(0) | 1(0.11) | 1(0.11) | 1(0.11) | |
| NSP11 | 2(0.39) | 2(0.39) | 2(0.39) | 2(0.39) | 2(0.39) | 3(0.58) | |
| S | S protein | 6(0.43) | 7(0.50) | 8(0.58) | 11(0.79) | 12(0.87) | 15(1.08) |
| ORF3 | ORF3 | 8(3.55) | 8(3.55) | 8(3.55) | 8(3.55) | 8(3.55) | 8(3.55) |
| E | E protein | 0(0) | 0(0) | 1(1.30) | 2(2.60) | 2(2.60) | 2(2.60) |
| M | M protein | 1(0.44) | 1(0.44) | 1(0.44) | 1(0.44) | 2(0.72) | 3(1.08) |
| N | N protein | 0(0) | 1(0.23) | 1(0.23) | 1(0.23) | 2(0.45) | 2(0.45) |
3.5. The Conserved and Highly Mutable Proteins

3.6. The Highly Variable Regions

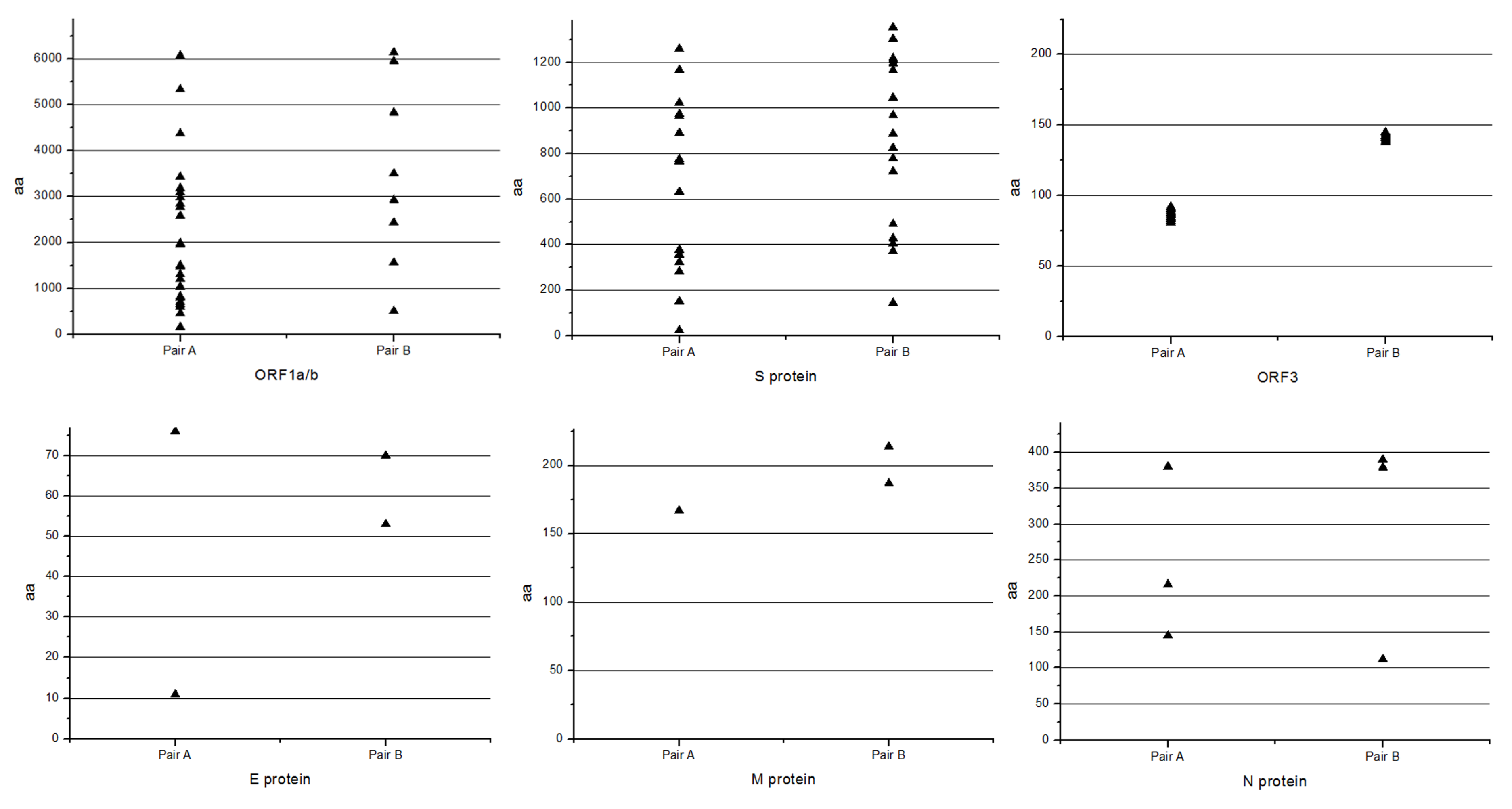
3.7. Comparison of Variation Characteristic of the Classical and Variant Virulent/Attenuated Strains
| Genes | Number of Amino Acid Change and Deletion when Compared Virulent DR13 with Attenuated DR13 | Number of Amino Acid Change and Deletion when Compared YN1 with YN144 |
|---|---|---|
| ORF1a/b | 26 | 9 |
| S | 18; One aa deletion at 151aa | 16; Three deletions at 144–145aa, and 1197aa |
| ORF3 | 12; Early termination at 92aa | 8; Early termination at 144aa |
| E | 2 | 2 |
| M | 1 | 2 |
| N | 3 | 2 |

4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Song, D.; Park, B. Porcine epidemic diarrhoea virus: A comprehensive review of molecular epidemiology, diagnosis, and vaccines. Virus Genes 2012, 44, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, H.; Liu, Y.; Pan, Y.; Deng, F.; Song, Y.; Tang, X.; He, Q. New variants of porcine epidemic diarrhea virus, China, 2011. Emerg. Infect. Dis. 2012, 18, 1350–1353. [Google Scholar] [CrossRef] [PubMed]
- Xuan, H.; Xing, D.; Wang, D.; Zhu, W.; Zhao, F.; Gong, H. Study on the culture of porcine epidemic diarrhea virus adapted to fetal porcine intestine primary cell monolayer. Chin. J. Vet. Sci. 1984, 4, 202–208. [Google Scholar]
- Chen, J.F.; Sun, D.B.; Wang, C.B.; Shi, H.Y.; Cui, X.C.; Liu, S.W.; Qiu, H.J.; Feng, L. Molecular characterization and phylogenetic analysis of membrane protein genes of porcine epidemic diarrhea virus isolates in China. Virus Genes 2008, 36, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.L.; Zhu, L.; Ma, J.Y.; Zhou, Q.F.; Song, Y.H.; Sun, B.L.; Chen, R.A.; Xie, Q.M.; Bee, Y.Z. Molecular characterization and phylogenetic analysis of porcine epidemic diarrhea virus (PEDV) field strains in south China. Virus Genes 2012, 45, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.Q.; Cai, R.J.; Chen, Y.Q.; Liang, P.S.; Chen, D.K.; Song, C.X. Outbreak of porcine epidemic diarrhea in suckling piglets, China. Emerg. Infect. Dis. 2012, 18, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Kou, Q.; Ge, X.; Zhou, L.; Guo, X.; Yang, H. Phylogenetic analysis of porcine epidemic diarrhea virus field strains prevailing recently in China. Arch. Virol. 2012, 158, 711–715. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.M.; Niu, B.B.; Yan, H.; Gao, D.S.; Yang, X.; Chen, L.; Chang, H.T.; Zhao, J.; Wang, C.Q. Genetic properties of endemic Chinese porcine epidemic diarrhea virus strains isolated since 2010. Arch. Virol. 2013, 158, 2487–2494. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.W.; Xue, C.Y.; He, L.L.; Wang, Y.; Cao, Y.C. Bioinformatics insight into the spike glycoprotein gene of field porcine epidemic diarrhea strains during 2011–2013 in Guangdong, China. Virus Genes 2014, 49, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.W.; Dickerman, A.W.; Pineyro, P.; Li, L.; Fang, L.; Kiehne, R.; Opriessnig, T.; Meng, X.J. Origin, Evolution, and Genotyping of Emergent Porcine Epidemic Diarrhea Virus Strains in the United States. mBio 2013, 4, e00737-13. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, C. Outbreak-Related Porcine Epidemic Diarrhea Virus Strains Similar to US Strains, South Korea, 2013. Emerg. Infect. Dis. 2014, 20, 1223–1226. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Chung, W.; Chang, S.; Wen, C.; Liu, H.; Chien, C.; Chiou, M.T. US-like Strain of Porcine Epidemic Diarrhea Virus Outbreaks in Taiwan, 2013–2014. J. Vet. Med. Sci. 2014, 76, 1297–1299. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, C.; Shi, H.; Qiu, H.J.; Liu, S.; Shi, D.; Zhang, X.; Feng, L. Complete genome sequence of a Chinese virulent porcine epidemic diarrhea virus strain. J. Virol. 2011, 85, 11538–11539. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Zhang, J.; Deng, X.; Ye, Y.; Liao, M.; Fan, H. Complete genome sequence of a highly prevalent isolate of porcine epidemic diarrhea virus in South China. J. Virol. 2012, 86. [Google Scholar] [CrossRef] [PubMed]
- Kweon, C.H.; Kwon, B.J.; Lee, J.G.; Kewon, G.O.; Kang, Y.B. Derivation of attenuated porcine epidemic diarrhea virus (PEDV) as vaccine candidate. Vaccine 1999, 17, 2546–2553. [Google Scholar] [CrossRef]
- Hofmann, M.; Wyler, R. Propagation of the virus of porcine epidemic diarrhea in cell culture. J. Clin. Microbiol. 1988, 26, 2235–2239. [Google Scholar] [PubMed]
- Zeng, S.L.; Zhang, H.; Ding, Z.; Luo, R.; An, K.; Liu, L.Z.; Bi, J.; Chen, H.C.; Xiao, S.B.; Fang, L.R. Proteome analysis of porcine epidemic diarrhea virus (PEDV)-infected Vero cells. Proteomics 2015, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar]
- Zhang, K.; He, Q. Establishment and Clinical Application of a Multiplex Reverse Transcription PCR for Detection of Porcine Epidemic Diarrhea Virus, Porcine Transmissible Gastroenteritis Virus and Porcine Group A Rotavirus. Chin. J. Anim. Vet. Sci. 2010, 41, 1001–1005. [Google Scholar]
- Luo, C.Q.; Zhang, X.L.; Wang, Y.M.; Zhu, Z.H.; Deng, A.G. Effect of lipo-prostaglandin E1 on mesangial proliferative glomerulonephritis in rats. J. Huazhong Univ. Sci. Technol. Med. Sci. 2005, 25, 516–518. [Google Scholar] [PubMed]
- Pan, Y.; Tian, X.; Li, W.; Zhou, Q.; Wang, D.; Bi, Y.; Chen, F.; Song, Y. Isolation and characterization of a variant porcine epidemic diarrhea virus in China. Virol. J. 2012, 9, e195. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Lole, K.S.; Bollinger, R.C.; Paranjape, R.S.; Gadkari, D.; Kulkarni, S.S.; Novak, N.G.; Ingersoll, R.; Sheppard, H.W.; Ray, S.C. Full-Length Human Immunodeficiency Virus Type 1 Genomes from Subtype C-Infected Seroconverters in India, with Evidence of Inter-subtype Recombination. J. Virol. 1999, 73, 152–160. [Google Scholar] [PubMed]
- Krajewska, M.; Krajewski, S.; Epstein, J.I.; Shabaik, A.; Sauvageot, J.; Song, K.; Kitada, S.; Reed, J.C. Immunohistochemical analysis of bcl-2, bax, bcl-X, and mcl-1 expression in prostate cancers. Am. J. Pathol. 1996, 148, 1567–1576. [Google Scholar] [PubMed]
- Song, D.S.; Yang, J.S.; Oh, J.S.; Han, J.H.; Park, B.K. Differentiation of a Vero cell adapted porcine epidemic diarrhea virus from Korean field strains by restriction fragment length polymorphism analysis of ORF 3. Vaccine 2003, 21, 1833–1842. [Google Scholar] [CrossRef]
- Song, D.S.; Oh, J.S.; Kang, B.K.; Yang, J.S.; Moon, H.J.; Yoo, H.S.; Jang, Y.S.; Park, B.K. Oral efficacy of Vero cell attenuated porcine epidemic diarrhea virus DR13 strain. Res. Vet. Sci. 2007, 82, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Li, G.W.; Stasko, J.; Thomas, J.T.; Stensland, W.R.; Pillatzki, A.E.; Gauger, P.C.; Schwartz, K.J.; Madson, D.; Yoon, K.J.; et al. Isolation and Characterization of Porcine Epidemic Diarrhea Viruses Associated with the 2013 Disease Outbreak among Swine in the United States. J. Clin. Microbiol. 2014, 52, 234–243. [Google Scholar] [CrossRef] [PubMed]
- An, T.Q.; Tian, Z.J.; Zhou, Y.J.; Xiao, Y.; Peng, J.M.; Chen, J.; Jiang, Y.F.; Hao, X.F.; Tong, G.Z. Comparative genomic analysis of five pairs of virulent parental/attenuated vaccine strains of PRRSV. Vet. Microbiol. 2011, 149, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Moon, H.J.; Yang, J.S.; Lee, C.S.; Song, D.S.; Kang, B.K.; Park, B.K. Cloning and further sequence analysis of the spike gene of attenuated porcine epidemic diarrhea virus DR13. Virus Genes 2007, 35, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Puranaveja, S.; Poolperm, P.; Lertwatcharasarakul, P.; Kesdaengsakonwut, S.; Boonsoongnern, A.; Urairong, K.; Kitikoon, P.; Choojai, P.; Kedkovid, R.; Teankum, K.; et al. Chinese-like strain of porcine epidemic diarrhea virus, Thailand. Emerg. Infect. Dis. 2009, 15, 1112–1115. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Takeyama, N.; Katsumata, A.; Tuchiya, K.; Kodama, T.; Kusanagi, K. Mutations in the spike gene of porcine epidemic diarrhea virus associated with growth adaptation in vitro and attenuation of virulence in vivo. Virus Genes 2011, 43, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.P.; Hogue, B.G. Protein interactions during coronavirus assembly. J. Virol. 1997, 71, 9278–9284. [Google Scholar] [PubMed]
- Wang, K.; Lu, W.; Chen, J.; Xie, S.; Shi, H.; Hsu, H.; Yu, W.; Xu, K.; Bian, C.; Fischer, W.B.; et al. PEDV ORF3 encodes an ion channel protein and regulates virus production. FEBS Lett. 2012, 586, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Woods, R.D. Efficacy of a transmissible gastroenteritis coronavirus with an altered ORF 3 gene. Can. J. Vet. Res. 2001, 65, 28–32. [Google Scholar] [PubMed]
- Park, J.H.; Han, J.H.; Kwon, H.M. Sequence analysis of the ORF 7 region of transmissible gastroenteritis viruses isolated in Korea. Virus Genes 2008, 36, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, L.L.; Woodworth, J.C.; Zhang, J.Q.; Gauger, P.C.; Chen, Q.; Welch, M.; Salzebrenner, H.; Thomas, J.; Main, R.; Dirtz, S.S.; et al. Determining the minimum infectious dose of porcine epidemic diarrhea virus in a feed matrix. In Proceedings of the Midwest Meeting, Des Moines, IA, USA, 16–18 March 2015.
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, F.; Zhu, Y.; Wu, M.; Ku, X.; Ye, S.; Li, Z.; Guo, X.; He, Q. Comparative Genomic Analysis of Classical and Variant Virulent Parental/Attenuated Strains of Porcine Epidemic Diarrhea Virus. Viruses 2015, 7, 5525-5538. https://doi.org/10.3390/v7102891
Chen F, Zhu Y, Wu M, Ku X, Ye S, Li Z, Guo X, He Q. Comparative Genomic Analysis of Classical and Variant Virulent Parental/Attenuated Strains of Porcine Epidemic Diarrhea Virus. Viruses. 2015; 7(10):5525-5538. https://doi.org/10.3390/v7102891
Chicago/Turabian StyleChen, Fangzhou, Yinxing Zhu, Meizhou Wu, Xugang Ku, Shiyi Ye, Zhonghua Li, Xiaozhen Guo, and Qigai He. 2015. "Comparative Genomic Analysis of Classical and Variant Virulent Parental/Attenuated Strains of Porcine Epidemic Diarrhea Virus" Viruses 7, no. 10: 5525-5538. https://doi.org/10.3390/v7102891