Infectious Mononucleosis Triggers Generation of IgG Auto-Antibodies against Native Myelin Oligodendrocyte Glycoprotein

Abstract
:1. Introduction
2. Materials and Methods

{kind=link}
{kind=link}
{kind=link}
| Cohort 1 | Cohort 2 | |||
|---|---|---|---|---|
| Number | 10 | 22 | 13 | 13 |
| Gender (f/m) | 6/4 | 11/11 | 9/4 | 5/8 |
| Median Age (Range) | 9.5 (3-15) | 12 (2-15) | 16 (8-18) | 16 (7-18) |
| Duration of Symptoms | NA | 7-28 days | NA | 3-21 days |
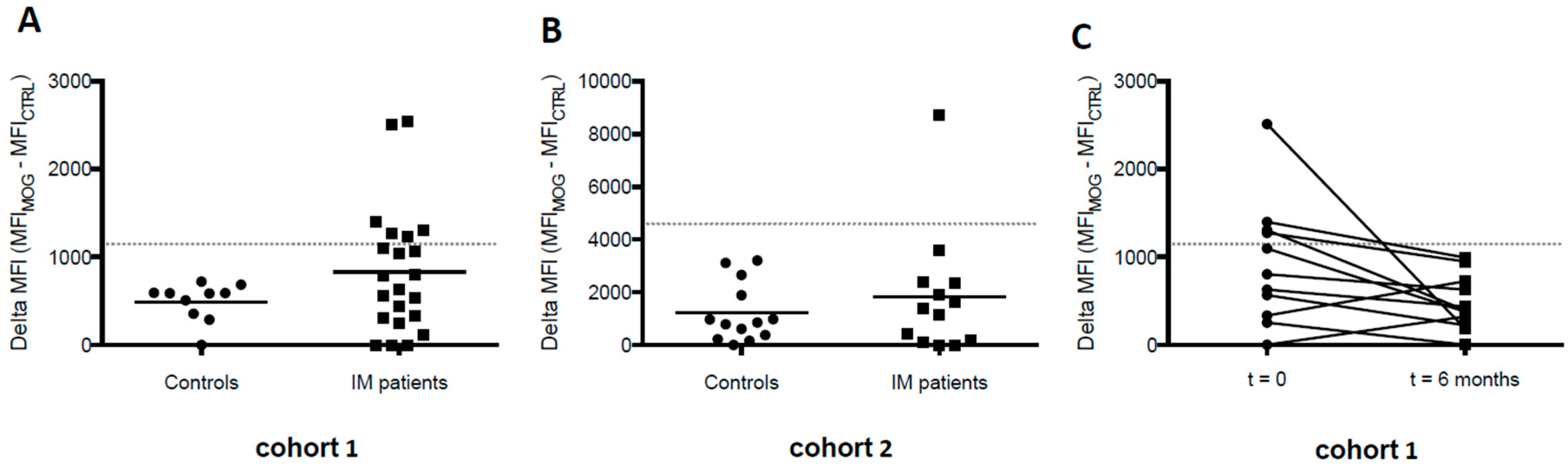
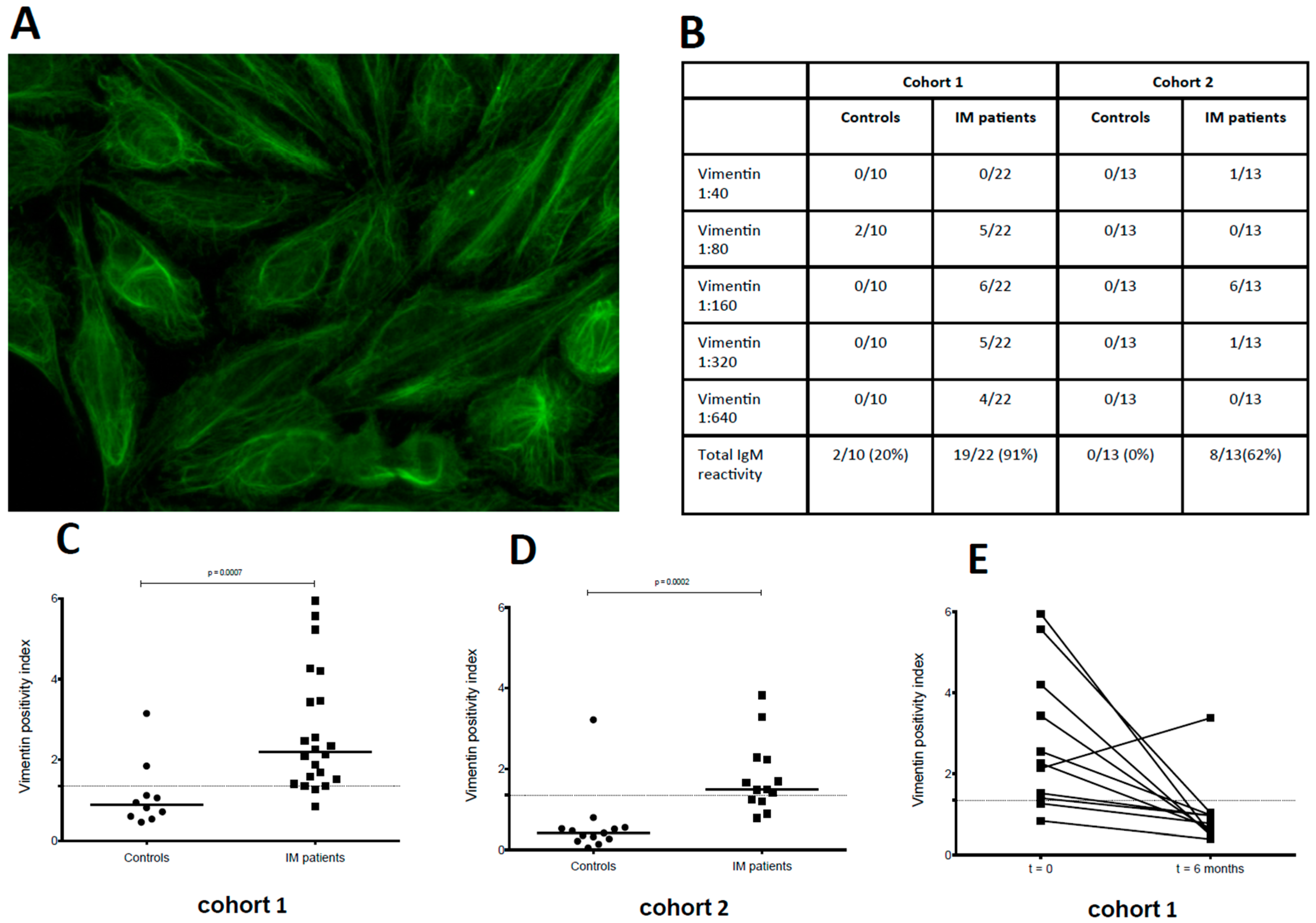
3. Results



4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Luzuriaga, K.; Sullivan, J.L. Infectious mononucleosis. N Engl. J. Med. 2010, 362, 1993–2000. [Google Scholar] [CrossRef] [PubMed]
- Balfour, H.H., Jr.; Odumade, O.A.; Schmeling, D.O.; Mullan, B.D.; Ed, J.A.; Knight, J.A.; Vezina, H.E.; Thomas, W.; Hogquist, K.A. Behavioral, virologic, and immunologic factors associated with acquisition and severity of primary Epstein-Barr virus infection in university students. J. Infect. Dis 2013, 207, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Ascherio, A.; Munger, K.L. Environmental risk factors for multiple sclerosis. Part I: The role of infection. Ann. Neurol. 2007, 61, 288–299. [Google Scholar] [CrossRef] [PubMed]
- Ascherio, A.; Munger, K.L.; Lunemann, J.D. The initiation and prevention of multiple sclerosis. Nat. Rev. Neurol. 2012, 8, 602–12. [Google Scholar] [CrossRef] [PubMed]
- James, J.A.; Neas, B.R.; Moser, K.L.; Hall, T.; Bruner, G.R.; Sestak, A.L.; Harley, J.B. Systemic lupus erythematosus in adults is associated with previous Epstein-Barr virus exposure. Arthritis Rheum. 2001, 44, 1122–1126. [Google Scholar] [CrossRef]
- Alotaibi, S.; Kennedy, J.; Tellier, R.; Stephens, D.; Banwell, B. Epstein-Barr virus in pediatric multiple sclerosis. JAMA 2004, 291, 1875–1879. [Google Scholar] [CrossRef] [PubMed]
- James, J.A.; Kaufman, K.M.; Farris, A.D.; Taylor-Albert, E.; Lehman, T.J.; Harley, J.B. An increased prevalence of Epstein-Barr virus infection in young patients suggests a possible etiology for systemic lupus erythematosus. J. Clin Investig. 1997, 100, 3019–3026. [Google Scholar] [CrossRef] [PubMed]
- Reindl, M.; Di Pauli, F.; Rostasy, K.; Berger, T. The spectrum of MOG autoantibody-associated demyelinating diseases. Nat. Rev. Neurol. 2013, 9, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Dale, R.C.; Tantsis, E.M.; Merheb, V.; Kumaran, R.Y.; Sinmaz, N.; Pathmanandavel, K.; Ramanathan, S.; Booth, D.R.; Wienholt, L.A.; Prelog, K.; et al. Antibodies to MOG have a demyelination phenotype and affect oligodendrocyte cytoskeleton. Neurol. Neuroimmunol. Neuroinflamm. 2014, 1, e12. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Srivastava, R.; Nessler, S.; Grummel, V.; Sommer, N.; Bruck, W.; Hartung, H.P.; Stadelmann, C.; Hemmer, B. Identification of a pathogenic antibody response to native myelin oligodendrocyte glycoprotein in multiple sclerosis. Proc. Natl. Acad. Sci. USA 2006, 103, 19057–19062. [Google Scholar] [CrossRef] [PubMed]
- Brilot, F.; Dale, R.C.; Selter, R.C.; Grummel, V.; Kalluri, S.R.; Aslam, M.; Busch, V.; Zhou, D.; Cepok, S.; Hemmer, B. Antibodies to native myelin oligodendrocyte glycoprotein in children with inflammatory demyelinating central nervous system disease. Ann. Neurol. 2009, 66, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Bahna, S.L.; Heiner, D.C.; Horwitz, C.A. Sequential changes of the five immunoglobulin classes and other responses in infectious mononucleosis. Int. Arch. Allergy Appl. Immunol. 1984, 74, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.I. Epstein-Barr virus infection. N Engl. J. Med. 2000, 343, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Sutton, R.N.; Emond, R.T.; Thomas, D.B.; Doniach, D. The occurrence of autoantibodies in infectious mononucleosis. Clin. Exp. Immunol. 1974, 17, 427–436. [Google Scholar] [PubMed]
- Rhodes, G.; Rumpold, H.; Kurki, P.; Patrick, K.M.; Carson, D.A.; Vaughan, J.H. Autoantibodies in infectious mononucleosis have specificity for the glycine-alanine repeating region of the Epstein-Barr virus nuclear antigen. J. Exp. Med. 1987, 165, 1026–1040. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, J.H.; Valbracht, J.R.; Nguyen, M.D.; Handley, H.H.; Smith, R.S.; Patrick, K.; Rhodes, G.H. Epstein-Barr virus-induced autoimmune responses. I. Immunoglobulin M autoantibodies to proteins mimicking and not mimicking Epstein-Barr virus nuclear antigen-1. J. Clin. Investig. 1995, 95, 1306–1315. [Google Scholar] [CrossRef] [PubMed]
- Liebowitz, D.; Kopan, R.; Fuchs, E.; Sample, J.; Kieff, E. An Epstein-Barr virus transforming protein associates with vimentin in lymphocytes. Mol. Cell. Biol 1987, 7, 2299–2308. [Google Scholar] [CrossRef] [PubMed]
- Meckes, D.G., Jr.; Menaker, N.F.; Raab-Traub, N. Epstein-Barr virus LMP1 modulates lipid raft microdomains and the vimentin cytoskeleton for signal transduction and transformation. J. Virol 2013, 87, 1301–1311. [Google Scholar] [CrossRef] [PubMed]
- Bretherton, L.; Toh, B.H. IgM autoantibody to intermediate filaments in infectious mononucleosis. J. Clin. Lab. Immunol. 1981, 5, 7–10. [Google Scholar] [PubMed]
- Kataaha, P.K.; Mortazavi-Milani, S.M.; Russell, G.; Holborow, E.J. Anti-intermediate filament antibodies, antikeratin antibody, and antiperinuclear factor in rheumatoid arthritis and infectious mononucleosis. Ann. Rheum. Dis. 1985, 44, 446–449. [Google Scholar] [CrossRef] [PubMed]
- Sairenji, T.; Nguyen, Q.V.; Woda, B.; Humphreys, R.E. Immune response to intermediate filament-associated, Epstein-Barr virus-induced early antigen. J. Immunol. 1987, 138, 2645–2652. [Google Scholar] [PubMed]
- Thorley-Lawson, D.A. Epstein-Barr virus: exploiting the immune system. Nat. Rev. Immunol 2001, 1, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Tracy, S.I.; Kakalacheva, K.; Lunemann, J.D.; Luzuriaga, K.; Middeldorp, J.; Thorley-Lawson, D.A. Persistence of Epstein-Barr virus in self-reactive memory B cells. J. Virol. 2012, 8, 12330–12340. [Google Scholar] [CrossRef] [PubMed]
- Bernasconi, N.L.; Traggiai, E.; Lanzavecchia, A. Maintenance of serological memory by polyclonal activation of human memory B cells. Science 2002, 298, 2199–22202. [Google Scholar] [CrossRef] [PubMed]
- Huggins, J.; Pellegrin, T.; Felgar, R.E.; Wei, C.; Brown, M.; Zheng, B.; Milner, E.C.; Bernstein, S.H.; Sanz, I.; Zand, M.S. CpG DNA activation and plasma-cell differentiation of CD27-naive human B cells. Blood 2007, 109, 1611–1619. [Google Scholar] [CrossRef] [PubMed]
- Dorner, M.; Brandt, S.; Tinguely, M.; Zucol, F.; Bourquin, J.P.; Zauner, L.; Berger, C.; Bernasconi, M.; Speck, R.F.; Nadal, D. Plasma cell toll-like receptor (TLR) expression differs from that of B cells, and plasma cell TLR triggering enhances immunoglobulin production. Immunology 2009, 128, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Lim, W.H.; Kireta, S.; Russ, G.R.; Coates, P.T. Human plasmacytoid dendritic cells regulate immune responses to Epstein-Barr virus (EBV) infection and delay EBV-related mortality in humanized NOD-SCID mice. Blood 2007, 109, 1043–1050. [Google Scholar] [CrossRef] [PubMed]
- Fiola, S.; Gosselin, D.; Takada, K.; Gosselin, J. TLR9 contributes to the recognition of EBV by primary monocytes and plasmacytoid dendritic cells. J. Immunol. 2010, 185, 3620–3631. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kakalacheva, K.; Regenass, S.; Wiesmayr, S.; Azzi, T.; Berger, C.; Dale, R.C.; Brilot, F.; Münz, C.; Rostasy, K.; Nadal, D.; et al. Infectious Mononucleosis Triggers Generation of IgG Auto-Antibodies against Native Myelin Oligodendrocyte Glycoprotein. Viruses 2016, 8, 51. https://doi.org/10.3390/v8020051
Kakalacheva K, Regenass S, Wiesmayr S, Azzi T, Berger C, Dale RC, Brilot F, Münz C, Rostasy K, Nadal D, et al. Infectious Mononucleosis Triggers Generation of IgG Auto-Antibodies against Native Myelin Oligodendrocyte Glycoprotein. Viruses. 2016; 8(2):51. https://doi.org/10.3390/v8020051
Chicago/Turabian StyleKakalacheva, Kristina, Stephan Regenass, Silke Wiesmayr, Tarik Azzi, Christoph Berger, Russell C. Dale, Fabienne Brilot, Christian Münz, Kevin Rostasy, David Nadal, and et al. 2016. "Infectious Mononucleosis Triggers Generation of IgG Auto-Antibodies against Native Myelin Oligodendrocyte Glycoprotein" Viruses 8, no. 2: 51. https://doi.org/10.3390/v8020051