
Epitope Identification and Application for Diagnosis of Duck Tembusu Virus Infections in Ducks

Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus, E-Specific Monoclonal Antibodies, and JEV-, DENV-, and WNV-Positive Sera
2.2. Affinity Purification of Monoclonal Antibodies
2.3. Epitope Mapping
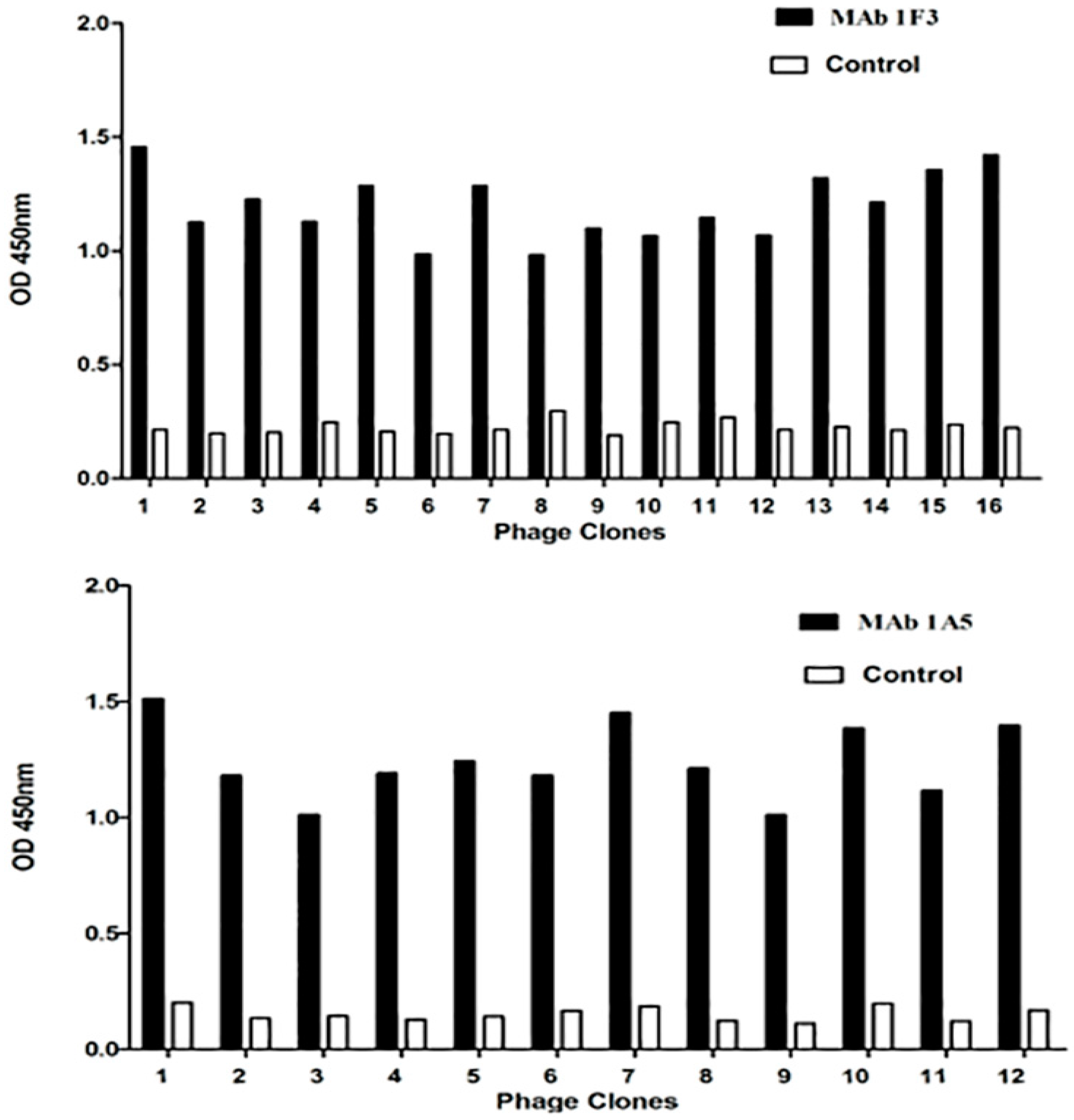
2.4. Phage Enzyme-Linked Immunosorbent Assay and Phage Clone Sequencing
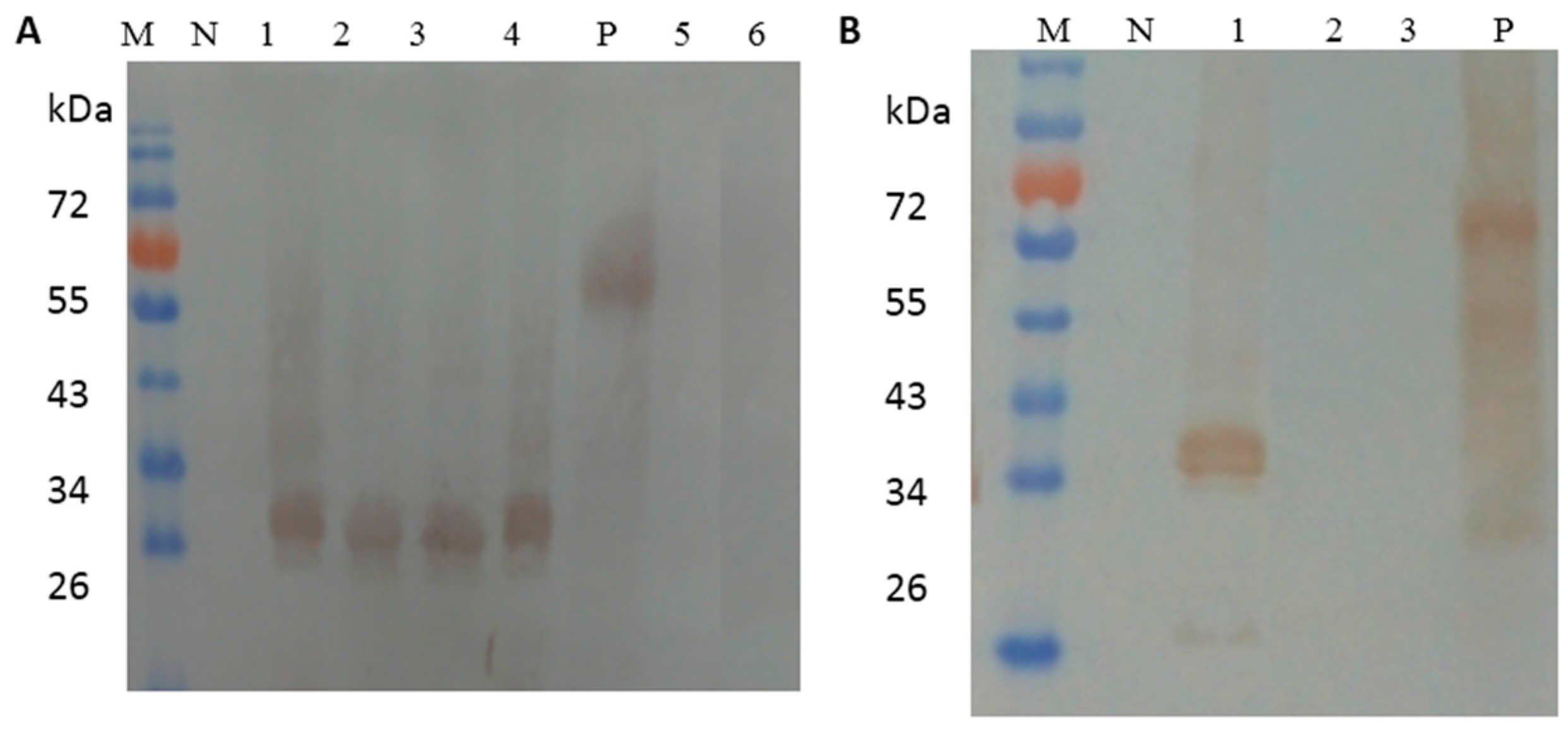
2.5. Identification of the Essential Amino Acids in the Epitopes by Dot Blotting and Western Blot Analysis
2.6. Sequence Analysis
2.7. Cross-Reactivity of the Epitopes to WNV-, JEV-, and DENV-Positive Sera
2.8. Protein E Modeling and Prediction
2.9. Competitive Inhibition Binding Assay of Monoclonal Antibody 1A5 to a Synthetic Peptide
2.10. Detection of DTMUV Infection in Duck Serum Samples
3. Results
3.1. Epitope Prediction
3.2. Mapping of the Minimal Epitopes by Dot Blotting and Western Blot
3.3. Sequence Analysis of the Identified Epitopes among the DTMUV Strains and Other Flaviviruses
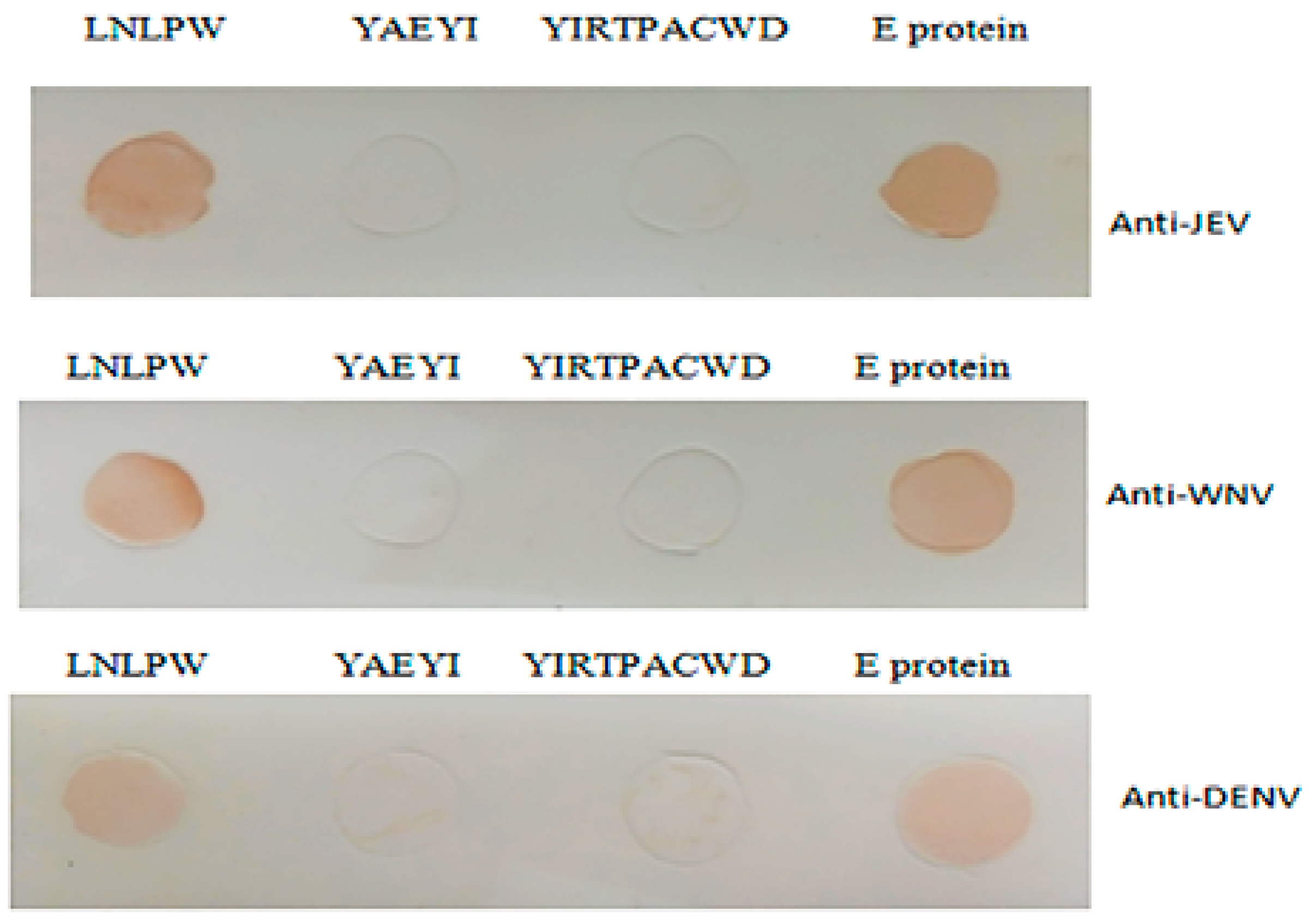
3.4. YAEYI and LNLPW Peptide Fragment Reactivity to WNV-, JEV-, and DENV-Positive Sera
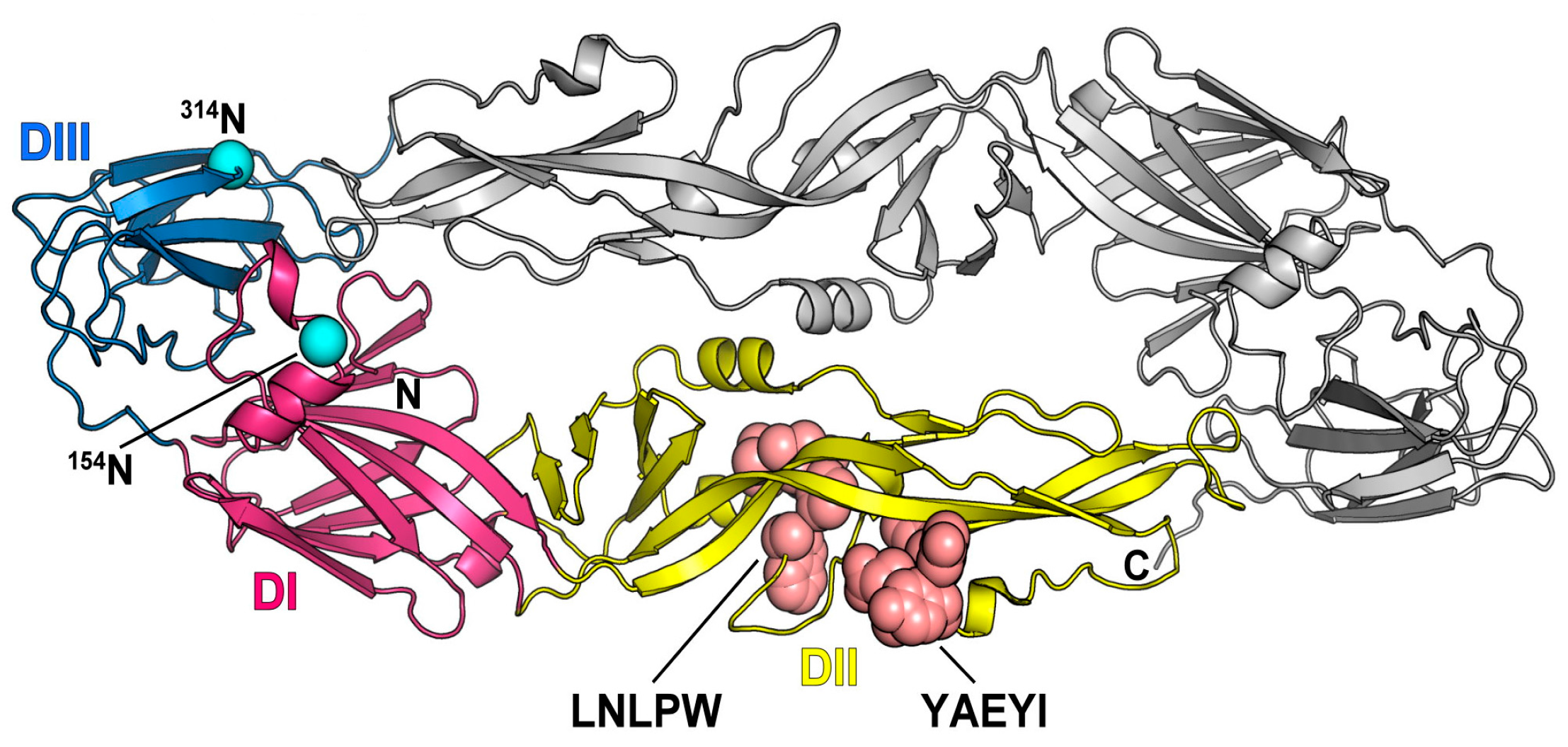
3.5. Location of Two Epitopes on the E Protein 3D Structure
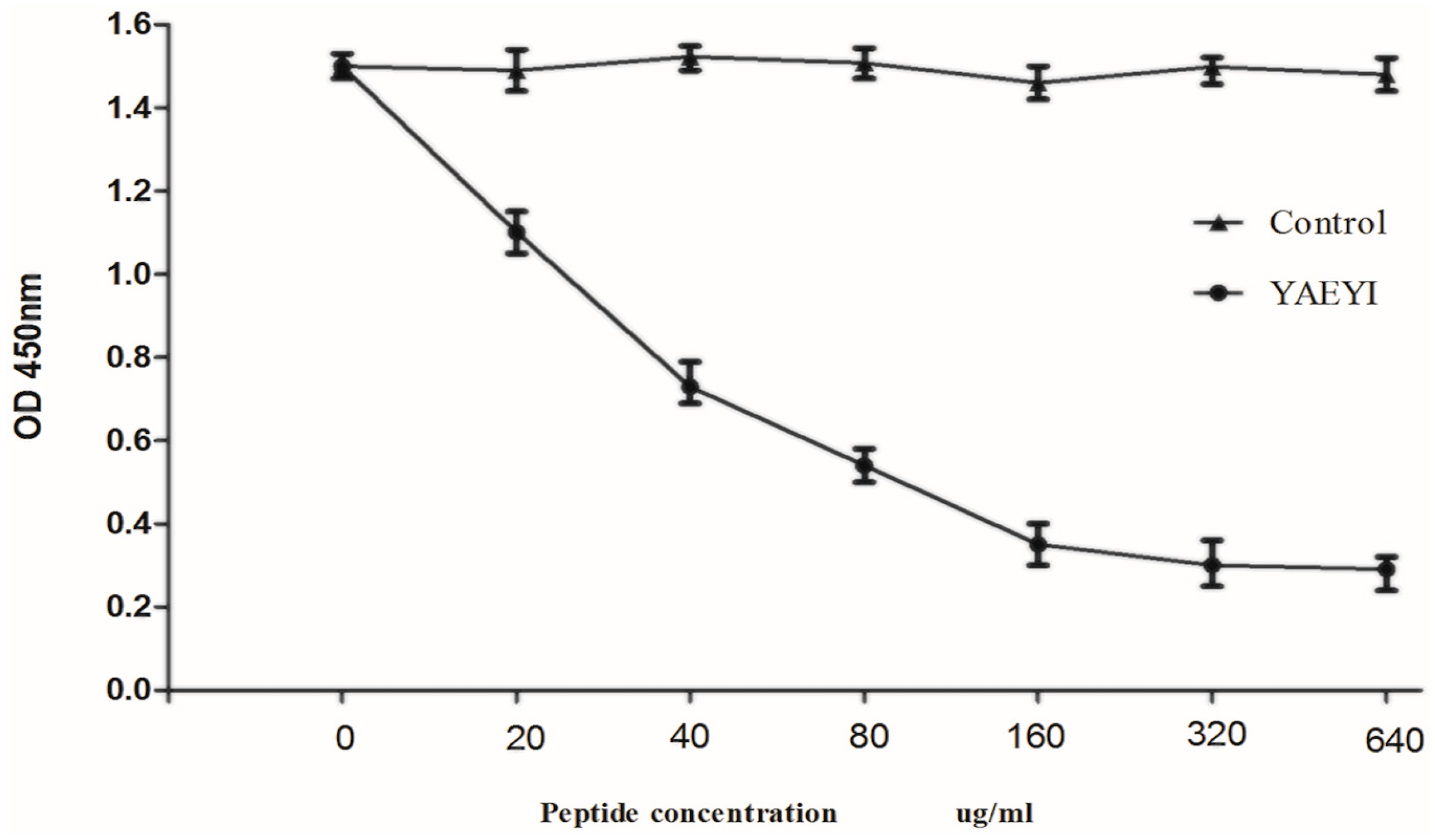
3.6. Competitive Inhibition of Synthetic Peptide YAEYI Binding to Monoclonal Antibody 1A5
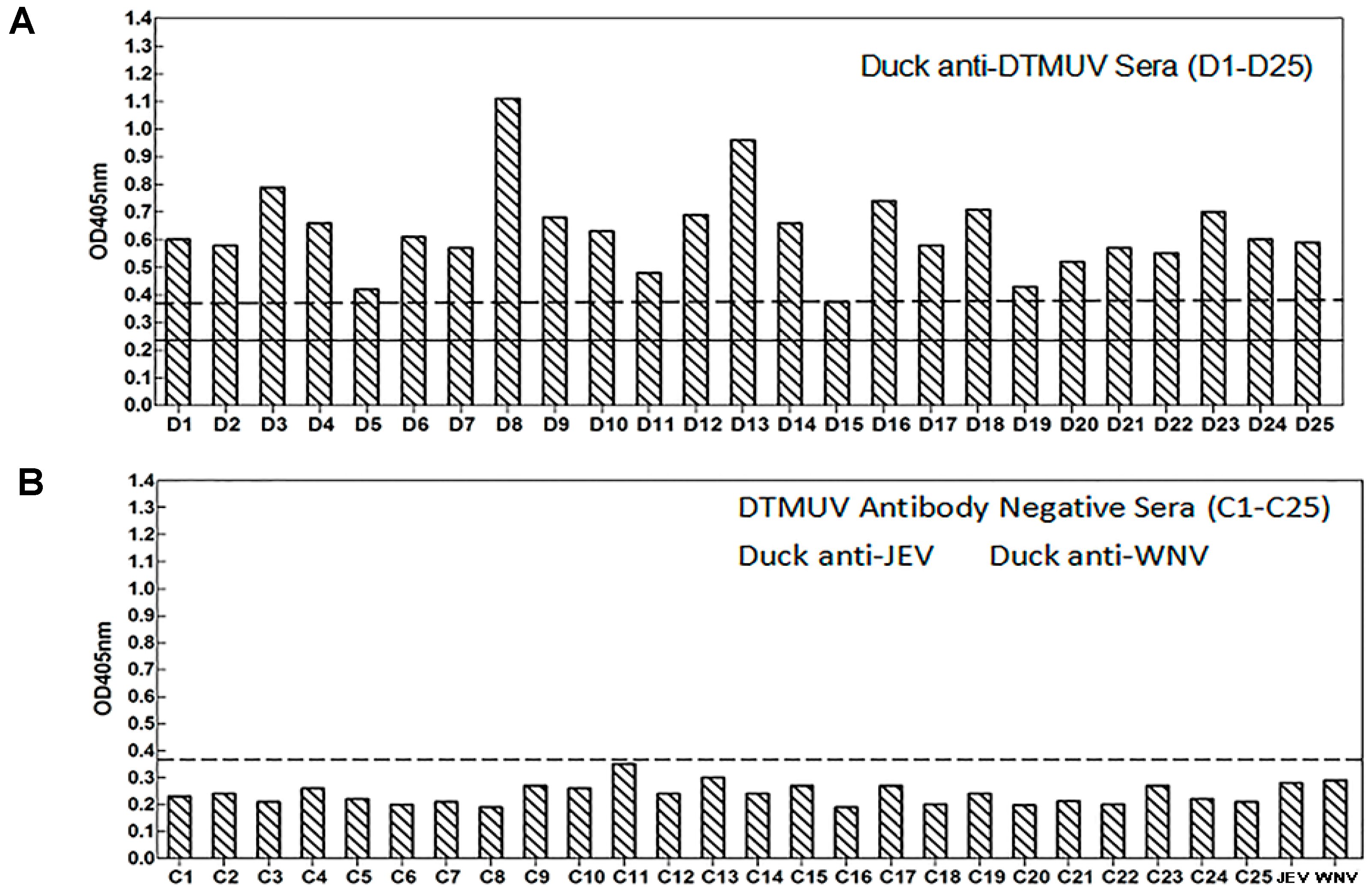
3.7. Sensitivity and Reactivity of an Epitope-Based Peptide Applied for the Diagnosis of DTMUV in Serum Samples
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lindenbach, B.D.; Thiel, H.J.; Rice, C.M. Flaviviridae: The viruses and their replication. In Fields Virology, 5th ed.; Knipe, D.M., Howley, P.M., Griffin, D.E., Lamb, R.A., Martin, M.A., Roizman, B., Straus, S.E., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; pp. 1101–1152. [Google Scholar]
- Su, J.J.; Li, S.; Hu, X.D.; Yu, X.L.; Wang, Y.Y.; Liu, P.P.; Lu, X.S.; Zhang, G.Z.; Hu, X.Y.; Liu, D.; et al. Duck egg drop syndrome caused by BYD virus, a new Tembusu related virus. PLoS ONE 2011, 6, 18106. [Google Scholar] [CrossRef] [PubMed]
- Homonnay, Z.G.; Kovacs, E.W.; Banyai, K.; Albert, M.; Feher, E.; Mato, T.; Tatar-Kis, T.; Palya, V. Tembusu-like flavivirus (Perak virus) as the cause of neurological disease outbreaks in young Pekin ducks. Avian Pathol. 2014, 43, 552–560. [Google Scholar] [CrossRef] [PubMed]
- Thontiravong, A.; Ninvilai, P.; Tunterak, W.; Nonthabenjawan, N.; Chaiyavong, S.; Angkabkingkaew, K.; Mungkundar, C.; Phuengpho, W.; Oraveerakul, K.; Amonsin, A. Tembusu-Related Flavivirus in Ducks, Thailand. Emerg. Infect. Dis. 2015, 21, 2164–2167. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Liu, C.G.; Li, G.; Li, X.J.; Yin, X.C.; Chen, Y.H.; Zhang, Y. Complete genomic sequence of duck flavivirus from China. J. Virol. 2012, 86, 3398. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Chen, S.Y.; Chen, Y.H.; Liu, C.G.; Chen, S.L.; Yin, X.C.; Li, G.; Zhang, Y. Adapted Tembusu-Like Virus in Chickens and Geese in China. J. Clin. Microbiol. 2012, 50, 2807–2809. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.Z.; Zhang, C.; Liu, Y.H.; Ye, W.C.; Han, J.W.; Ma, G.M.; Zhang, D.D.; Xu, F.; Gao, Y.; Tang, X.H.; et al. Tembusu virus in ducks, China. Emerg. Infect. Dis. 2011, 17, 1873–1875. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Liu, J.; Jiang, Y.; Zhao, Y.; Li, Q.; Wu, L.; He, X.; Chen, H. The vaccine efficacy of recombinant duck enteritis virus expressing secreted E with or without PrM proteins of duck tembusu virus. Vaccine 2014, 32, 5271–5277. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Han, K.; Zhao, D.; Liu, Y.; Zhang, J.; Niu, H.; Zhang, K.; Zhu, J.; Wu, D. Identification and molecular characterization of a novel flavivirus isolated from geese 265 in China. Res. Vet. Sci. 2013, 94, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.F.; Lv, R.; Liu, C.G.; Qiu, N.; He, Y.L.; Yin, X.C.; Li, X.J.; Liu, M.; Zhang, Y. Molecular characterization of a duck Tembusu virus from China. Virus Genes 2013, 47, 478–482. [Google Scholar] [CrossRef] [PubMed]
- Avirutnan, P.; Fuchs, A.; Hauhart, R.E.; Somnuke, P.; Youn, S.; Diamond, M.S.; Atkinson, J.P. Antagonism of the complement component C4 by flavivirus nonstructural protein NS1. J. Exp. Med. 2010, 207, 793–806. [Google Scholar] [CrossRef] [PubMed]
- Crill, W.D.; Chang, G.J. Localization and characterization of flavivirus envelope glycoprotein cross-reactive epitopes. J. Virol. 2004, 78, 13975–13986. [Google Scholar] [CrossRef] [PubMed]
- Crill, W.D.; Roehrig, J.T. Monoclonal antibodies that bind to domain III of dengue virus E glycoprotein are the most efficient blockers of virus adsorption to Vero cells. J. Virol. 2001, 75, 7769–7773. [Google Scholar] [CrossRef] [PubMed]
- Roehrig, J.T.; Diamond, M.S.; Kuhn, R.J.; Rossmann, M.G. Binding of a neutralizing antibody to dengue virus alters the arrangement of surface glycoproteins. Nat. Struct. Mol. Biol. 2008, 15, 312–317. [Google Scholar]
- Roehrig, J.T.; Bolin, R.A.; Kelly, R.G. Monoclonal antibody mapping of the envelope glycoprotein of the dengue 2 virus, Jamaica. Virology 1998, 246, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Rey, F.A.; Heinz, F.X.C.; Mandl, C.; Harrison, S.G. The envelope glycoprotein from tick-borne encephalitis virus at 2 Å resolution. Nature 1995, 375, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Roehrig, J.T. Immunochemistry of the dengue viruses. In Dengue and Dengue Hemorrhagic Fever; Gubler, D.J., Kuno, G., Eds.; CAB International: New York, NY, USA, 1997; pp. 199–219. [Google Scholar]
- Shi, P.Y.; Wong, S.J. Serologic diagnosis of West Nile virus infection. Expert Rev. Mol. Diagn. 2003, 3, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Hirota, J.; Shimizu, S.; Shibahara, T.; Kobayashi, S. Cross-reactivity of chicken anti-Japanese encephalitis virus serum and anti-West Nile virus serum in serological diagnosis. J. Vet. Med. Sci. 2012, 74, 1497–1499. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.A.; Muth, D.A.; Brown, T.; Johnson, A.J.; Karabatsos, N.; Roehrig, J.T. Standardization of immunoglobulin M capture enzyme-linked immunosorbent assays for routine diagnosis of arboviral infections. J. Clin. Microbiol. 2000, 38, 1823–1826. [Google Scholar] [PubMed]
- Martin, D.A.; Biggerstaff, B.J.; Allen, B.; Johnson, A.J.; Lanciotti, R.S.; Roehrig, J.T. Use of immunoglobulin M cross-reactions in differential diagnosis of human fl aviviral encephalitis infections in the United States. Clin. Diagn. Lab. Immunol. 2002, 9, 544–549. [Google Scholar] [PubMed]
- Hall, R.A.; Broom, A.K.; Hartnett, A.C.; Howard, M.J.; Mackenzie, J.S. Immunodominant epitopes on the NS1 protein of MVE and KUN viruses serve as targets for a blocking ELISA to detect virus-specific antibodies in sentinel animal serum. J. Virol. Meth. 1995, 51, 201–210. [Google Scholar] [CrossRef]
- Bai, X.F.; Shaozhou, W.L.; Zhang, Q.S.; Li, C.X.; Qiu, N.; Meng, R.Z.; Liu, M.; Zhang, Y. Characterization of monoclonal antibodies against duck Tembusu virus E protein: An antigen-capture ELISA for the detection of Tembusu virus infection. Arch. Virol. 2015, 160, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Shi, X.M.; Zhang, J.; Zhao, Y.; Cui, H.Y. Identification of a Conserved B-cell Epitope on Reticuloendotheliosis Virus Envelope Protein by Screening a Phage-displayed Random Peptide Library. PLoS ONE 2012, 7, e49842. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.Y.; Li, X.J.; Zhang, Q.S.; Wulin, S.Z.; Bai, X.F.; Zhang, Z.Z. Identification of a Conserved B-Cell Epitope on Duck Hepatitis A Type 1 Virus VP1 Protein. PLoS ONE 2015, 10, e0118041. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.F.; Yin, X.C.; Chen, X.D.; Li, X.J.; Li, J.Z.; Liu, C.G.; Liu, M.; Zhang, Y. Antigenic analysis monoclonal antibodies against different epitopes of σB protein of Muscovy duck reovirus. Virus Res. 2012, 163, 546–551. [Google Scholar] [CrossRef] [PubMed]
- Burland, T.G. DNASTAR’s Lasergene sequence analysis software. Meth. Mol. Biol. 2000, 132, 71–91. [Google Scholar]
- Luca, V.C.; AbiMansour, J.; Nelson, C.A.; Fremont, D.H. Crystal structure of the Japanese encephalitis virus envelope protein. J. Virol. 2012, 86, 2337–2346. [Google Scholar] [CrossRef] [PubMed]
- Eswar, N.; Webb, B.; Marti-Renom, M.A.; Madhusudhan, M.S.; Eramian, D.; Shen, M.Y.; Pieper, U.; Sali, A. Comparative protein structure modeling using MODELLER. Curr. Protoc. Protein Sci. 2007. [Google Scholar] [CrossRef]
- Wiederstein, M.; Sippl, M.J. ProSA-web: Interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucleic Acids Res. 2007, 35, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, R.A.; Macarthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A program to check the stereochemical quality of protein structures. J. Appl. Cryst. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Chauhan, J.S.; Rao, A.; Raghava, G.P. In silico platform for prediction of N-, O- and C-glycosites in eukaryotic protein sequences. PLoS ONE 2013, 8, e67008. [Google Scholar] [CrossRef] [PubMed]
- Chuang, G.Y.; Boyington, J.C.; Joyce, M.G.; Zhu, J.; Nabel, G.J.; Kwong, P.D.; Georgiev, I. Computational prediction of N-linked glycosylation incorporating structural properties and patterns. Bioinformatics 2012, 28, 2249–2255. [Google Scholar] [CrossRef] [PubMed]
- DeLano, W.L. The PyMOL Molecular Graphics System; v1.5.0.4; Schrödinger, LLC: New York, NY, USA, 2004. [Google Scholar]
- Tam, J.P.; Zavala, F. Multiple antigen peptide: A novel approach to increase detection sensitivity of synthetic peptides in solid-phase immunoassays. J. Immunol. Methods 1989, 124, 53–61. [Google Scholar] [CrossRef]
- Sun, E.C.; Zhao, J.; Yang, T.; Liu, N.H.; Geng, H.W.; Qin, Y.L.; Wang, L.F.; Bu, Z.G.; Yang, Y.H.; Lunt, R.A.; et al. Identification of a conserved JEV serocomplex B-cell epitope by screening a phage-display peptide library with a mAb generated against West Nile virus capsid protein. Virol. J. 2011, 8, 100. [Google Scholar] [CrossRef] [PubMed]
- Wahala, W.M.; Silva, A.M. The human antibody response to dengue virus infection. Viruses 2011, 3, 2374–2395. [Google Scholar] [CrossRef] [PubMed]
- Fibriansah, G.; Ng, T.S.; Kostyuchenko, V.A.; Lee, J.; Lee, S.; Wang, J.; Lok, S.M. Structural changes in dengue virus when exposed to a temperature of 37 degrees. J. Virol. 2013, 87, 7585–7592. [Google Scholar] [CrossRef] [PubMed]
- Austin, S.K.; Dowd, K.A.; Shrestha, B.; Nelson, C.A.; Edeling, M.A.; Johnson, S.; Pierson, T.C.; Diamond, M.S.; Fremont, D.H. Structural basis of differential neutralization of DENV-1 genotypes by an antibody that recognizes a cryptic epitope. PLoS Pathog. 2012, 8, e1002930. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phage Clone | Sequence | Phage Clone | Sequence | |||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A1 | E | L | N | L | P | W | Q | R | N | A | L | V | B1 | S | R | N | L | S | Y | A | E | Y | I | Q | I | |||||||||
| A2 | S | A | E | N | D | L | T | L | P | W | T | T | B2 | G | N | Y | S | E | Y | I | V | G | K | L | V | |||||||||
| A3 | M | A | N | A | E | I | D | L | P | W | T | K | B3 | S | S | Y | A | N | Y | I | Q | F | R | N | T | |||||||||
| A4 | H | P | H | D | L | N | D | L | T | S | P | F | B4 | S | S | Y | T | A | Y | I | M | A | R | G | Q | |||||||||
| A5 | E | F | W | T | A | L | S | D | P | W | Y | F | B5 | N | S | M | S | E | Y | I | N | Y | I | L | T | |||||||||
| A6 | A | H | L | H | D | P | F | T | T | L | S | P | B6 | V | D | Y | S | T | Y | I | S | R | L | T | S | |||||||||
| A7 | L | D | F | H | D | L | N | R | P | F | N | N | B7 | N | F | M | N | Y | A | E | Y | V | Q | K | K | |||||||||
| A8 | T | H | D | P | L | D | S | P | W | N | F | S | B8 | V | D | Y | S | T | Y | I | S | R | L | T | S | |||||||||
| A9 | F | N | D | L | D | L | P | F | G | K | R | A | B9 | T | V | H | S | Y | E | E | Y | T | A | R | R | |||||||||
| A10 | S | Y | D | L | D | L | P | W | I | A | R | K | B10 | V | S | P | Y | A | E | Y | W | L | S | Q | M | |||||||||
| A11 | S | F | L | E | L | D | P | P | W | T | T | N | B11 | W | D | Y | N | L | Y | I | K | Y | V | A | R | |||||||||
| A12 | Q | H | S | F | L | D | L | P | W | H | L | T | B12 | V | D | Y | A | T | Y | I | S | R | L | T | S | |||||||||
| A13 | H | P | H | D | L | N | L | P | T | S | P | F | ||||||||||||||||||||||
| A14 | H | P | H | D | L | N | L | P | T | S | P | F | ||||||||||||||||||||||
| A15 | M | A | N | A | D | L | N | L | P | W | T | K | ||||||||||||||||||||||
| A16 | T | S | H | S | W | D | L | N | L | P | S | G | ||||||||||||||||||||||
| Consensus | D | L | D/N | L | P | W | T | Y | A | E | Y | I | ||||||||||||||||||||||
| Virus TA | 219 | H | D | L | N | L | P | W | T | 226 | 84 | K | A | T | Y | A | E | Y | I | C | K | K | D | 97 | ||||||||||
| Primers | Sequence | Truncated Peptide |
|---|---|---|
| 1F3-1-F | 5′-aattcgatctcaacttaccatggacac-3′ | GST-DLNLPWT |
| 1F3-1-R | 5′-tcgagtgtccatggtaagttgagatcg-3′ | |
| 1F3-2-F | 5′-aattcgatctcgacttaccatggacac-3′ | GST-DLDLPWT |
| 1F3-2-R | 5′-tcgagtgtccatggtaagtcgagatcg-3′ | |
| 1F3-3-F | 5′-aattcgatctcaacttaccatggc-3′ | GST-DLNLPW |
| 1F3-3-R | 5′-tcgagccatggtaagttgagatcg-3′ | |
| 1F3-4-F | 5′-aattcctcaacttaccatggc-3′ | GST-LNLPW |
| 1F3-4-R | 5′-tcgagccatggtaagttgagg-3′ | |
| 1F3-5-F | 5′-aattcctcaacttaccac-3′ | GST-LNLP |
| 1F3-5-R | 5′-tcgagtggtaagttgagg-3′ | |
| 1F3-6-F | 5′-aattcaacttaccatggc-3′ | GST-NLPW |
| 1F3-6-R | 5′-tcgagccatggtaagttg-3′ | |
| 1A5-1-F | 5′-aattctacgctgaatacatac-3′ | GST-YAEYI |
| 1A5-1-R | 5′-tcgagtatgtattcagcgtag-3′ | |
| 1A5-2-F | 5′-aattcgctgaatacatac-3′ | GST-AEYI |
| 1A5-2-R | 5′-tcgagtatgtattcagcg-3′ | |
| 1A5-3-F | 5′-aattctacgctgaatacc-3′ | GST-YAEY |
| 1A5-3-R | 5′-tcgaggtattcagcgtag-3′ |
| Species | Strain | GenBank No. | Location/Year of Isolation | |
|---|---|---|---|---|
| DTMUV | TA | JQ289550.1 | China | 2010 |
| DTMUV | DK/TH/CU-1 | KR061333.1 | Thailand | 2013 |
| DTMUV | GDHD2014-3 | KT159713.1 | China | 2014 |
| DTMUV | WFZ-2012 | KC990545.1 | China | 2012 |
| DTMUV | SH001 | KP742476.1 | China | 2015 |
| DTMUV | AH2011 | KJ958533.1 | China | 2012 |
| DTMUV | GDLH01 | KT824876.1 | China | 2015 |
| DTMUV | WZDu | AB917089.1 | China | 2012 |
| DTMUV | ZJ GH-2 | JQ314465.1 | China | 2010 |
| DTMUV | SDLC | KJ740747.1 | China | 2013 |
| DENV-1 | HNRG24827 | KC692511.1 | Argentina | 2010 |
| DENV-2 | DENV-2/Pk/Swat-02 | KJ701507.1 | Pakistan | 2013 |
| DENV-3 | D83-144 | KJ737430.1 | Thailand | 1983 |
| DENV-4 | EHI310A129SY10 | JX024758.1 | Singapore | 2010 |
| WNV | USA | AY646354.1 | USA | 2002 |
| WNV | FtC-3699 | KR868734.1 | USA | 2012 |
| JEV | HEN0701 | FJ495189.1 | China | 2007 |
| JEV | CC27-S6 | AY303797.1 | Taiwan | 2003 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Liu, J.; Shaozhou, W.; Bai, X.; Zhang, Q.; Hua, R.; Liu, J.-H.; Liu, M.; Zhang, Y. Epitope Identification and Application for Diagnosis of Duck Tembusu Virus Infections in Ducks. Viruses 2016, 8, 306. https://doi.org/10.3390/v8110306
Li C, Liu J, Shaozhou W, Bai X, Zhang Q, Hua R, Liu J-H, Liu M, Zhang Y. Epitope Identification and Application for Diagnosis of Duck Tembusu Virus Infections in Ducks. Viruses. 2016; 8(11):306. https://doi.org/10.3390/v8110306
Chicago/Turabian StyleLi, Chenxi, Junyan Liu, Wulin Shaozhou, Xiaofei Bai, Qingshan Zhang, Ronghong Hua, Jyung-Hurng Liu, Ming Liu, and Yun Zhang. 2016. "Epitope Identification and Application for Diagnosis of Duck Tembusu Virus Infections in Ducks" Viruses 8, no. 11: 306. https://doi.org/10.3390/v8110306