
Abstract
Dendritic cells (DC)-derived or tumor-derived exosomes are a population of nanometer sized membrane vesicles that can induce specific anti-tumor immunity. However, the immunogenic potential and efficiency of exosomes-based tumor vaccine are not satisfactory enough to achieve a curative effect in clinical trials. In this article we investigated whether IL-18 genetic modification of tumor cells can increase the efficacy of exosomes derived from IL-18 gene-modified tumor cells. We transfected carcinoembryonic antigen (CEA)-expressing tumor cells with a recombinant adenovirus encoding human IL-18 (AdhIL-18) and prepared the exosomes, Exo/IL-18, from IL-18 gene-modified tumor cells. We found that Exo/IL-18 naturally contain CEA and bioactive IL-18. Moreover, Exo-IL-18 are potent in chemoattracting DC and T cells, enhancing the proliferation and Th1 cytokine release of PBMC, and promoting the phenotypic and functional maturation of DC. Furthermore, Exo/IL-18-pulsed DC are quite potent to induce HLA-A*0201-restricted, CEA-specific CD8+ CTL from the PBMC of HLA-A*0201 CEA+ cancer patients in vitro. In almost all of these experiments, Exo/IL-18 show more potent functions than the conventionally prepared exosomes derived from parent tumor cells without IL-18 gene modification. Our findings suggest that Exo/IL-18 has more potent capability to induce specific anti-tumor immunity, and our strategy of IL-18 modification of exosomes is a feasible approach to develop exosomes-based tumor vaccines.
Similar content being viewed by others


Introduction
Developing effective tumor vaccines is a major task of cancer immunotherapy. Ideal tumor vaccine should be able to elicit tumor-specific T cells capable of recognizing and eliminating tumor cells [1]. In recent years, the identification of a kind of potential tumor vaccine, exosomes, has brought new light for the preparation of ideal tumor vaccine. Exosomes, as a kind of cell-free vaccine, are a population of nanometer sized membrane vesicles secreted by many types of cells such as dendritic cells (DC), T and B lymphocytes and tumor cells [2, 3]. Two types of exosomes, DC-derived exosomes and tumor-derived exosomes, showed capability of inducing tumor-specific T cell responses and protective anti-tumor immunity in animal models [2, 3]. Exosome-based vaccines are even under clinical trials for cancer immunotherapy [4, 5]. However, the immunogenic potential and efficiency of exosomes-based tumor vaccine remain to be improved to obtain satisfactory curative effects. It was shown that vaccination of exosomes bearing tumor antigens in combination with immunoadjuvants (e.g. CpG) can induce the augmented anti-tumor effects [6]. Adjuvants, especially Th1-type cytokines (such as IL-2, IL-12 and IFN-γ), were proven to be helpful in tumor immunotherapy [7]. Therefore, we investigated the efficiency of tumor-derived exosomes after the incorporation of cytokines into tumor cells.
IL-18, previously identified as IFN-γ-inducing factor, is also a promising vaccine adjuvant [8]. IL-18 elicits a variety of effects on immune response, including stimulating natural killer cells (NK) and T/B lymphocytes to produce high levels of IFN-γ and IL-2 [8, 9], promoting T cell proliferation [8, 10], enhancing killing activity of cytotoxic T lymphocytes (CTL) and NK cells [11–13], facilitating DC maturation [14], exerting pro-inflammatory properties by inducing the release of IL-1β, TNF-α, chemokines [15] and chemoattracting DC, T cells and polymorphonuclear cells [16–18]. We and others have previously shown that IL-18 has potent anti-tumor immunity by inducing CD4+, cytotoxic CD8+ T cells and NK cells [12, 13, 19–23], potentiating FasL-mediated cytotoxicity and inhibiting tumor angiogenesis [12, 24]. Therefore, the pleiotropic activities of IL-18 suggest that IL-18 plays an important role in the induction of anti-tumor immunity.
In this study, we investigated the efficacy and specificity of exosomes derived from IL-18 gene-modified tumor cells. We infected LS-174T tumor cells, the well-known human colon adenocarcinoma cell line producing high levels of CEA with recombinant adenovirus expressing human IL-18 (AdhIL-18) to obtain exosomes containing bioactive IL-18 (Exo/IL-18). We demonstrated that Exo/IL-18 enhance DC functions in vitro by up-regulating the expression of co-stimulatory molecules, enhancing the production of IL-12p70 and TNF-α and promoting the immmunostimulatory capacity of DC. Exo/IL-18 also effectively chemoattract DC and T cells. Most importantly, a superior CEA-specific CTL are generated from lymphocytes derived from HLA-A*0201 CEA+ cancer patients when the lymphocytes are incubated with Exo/IL-18-pulsed DC, compared with those generated from the lymphocytes incubated with DC pulsed with other exosomes including exosomes derived from LS-174T cells (Exo) or AdLacZ genetically modified LS-174T cells (Exo/LacZ). These findings suggest that Exo/IL-18 are more potent in the induction of anti-tumor immunity than traditional exosomes, and the preparation of exosomes from tumor cells geneticaly modified with cytokine adjuvants is a simple and feasible strategy in developing exosomes-based tumor vaccines.
Materials and methods
Cell lines and synthetic peptides
LS-174T (HLA-A2+, CEA+), SW480 (HLA-A2+, CEA+), LoVo (HLA-A2−, CEA+), human TAP-deficient T2 cells and transformed HEK293 cells were obtained from the American Type Culture Collection (Manassas, VA) and maintained in the corresponding culture medium according to the supplier’s specifications. All cells are free of mycoplasma. The HLA-A*0201-restricted CEA peptide CAP-1 (YLSGANLNL) and the irrelevant HLA-A*0201-restricted coronavirus SARS-CoV spike protein peptide SSp-1 (RLNEVAKNL) were synthesized according to standard solid phase synthesis methods and were purified by HPLC. The purity (more than 95%) and identity of peptides were determined by analytical HPLC and mass spectrometry analysis, respectively.
Patients
All patients were required to give written and informed consent before participation in the study. Patients had a histologically confirmed diagnosis of stage III carcinoma of colon. Entry criteria were as follows: confirmation of HLA-A*0201 by PCR-based DNA typing (One Lambda, Inc. CA, USA), positive CEA expression (as defined by immunohistochemical analysis), no chemotherapy or radiotherapy 4 weeks before the study. In our study, four HLA-A2+-restricted CEA+ patients (two males and two females) were selected.
Construction and identification of recombinant human IL-18 adenovirus
We constructed AdhIL-18 using AdEasy™ Adenoviral Vector System (Stratagene, La Jolla, CA). Briefly, DNA fragment encoding the active form of hIL-18 with IL-2 signal peptide at the N-terminal were synthesized and inserted into the expression vector pShuttle-CMV between the Xho I and EcoR V sites, and the resulting plasmid was designated pAdhIL-18. Linearized pAdhIL-18 was co-transformed into competent Escherichia coli BJ5183 cells with supercoiled circular pAdEasy-1 by electroporation. Recombinant adenoviral vector DNA isolated from the selected positive clone was linearized and transfected into HEK293 cells to pack recombinant adenovirus AdhIL-18 by a MBS mammalian transfection kit (Stratagene) according to the manufacturer’s standard protocol. After several cycles of propagation, the recombinant adenoviruses AdhIL-18 were purified by CsCl banding, and the final yields were measured as described previously [25]. The expression of hIL-18 was confirmed by ELISA assay (Medical & Biological Laboratories Co., Ltd, Nagaoya, Japan) of the culture supernatants of AdIL-18-transducted LS-174T cells.
Bioassay of hIL-18
The biological activity of hIL-18 was determined by its ability to induce IFN-γ production of peripheral blood mononuclear cells (PBMC) in the presence of Con A as described previously [8]. PBMC were plated at 2×105 cells/well in RPMI1640 supplemented with 50 U/ml penicillin, 50 mg/L streptomycin, 2 mM l-glutamine, 1 mM sodium pyruvate, and 10% decomplemented FCS in a final volume of 200 μl together with 0.5 mg/L Con A and the samples (either supernatant or exosomes). The cultures were incubated for 48 h at 37°C in 5% CO2, and then supernatants were analyzed for human IFN-γ by ELISA (R&D systems, Minneapolis, MN).
Exosomes purification
Exosomes were purified as described previously [26–28]. 2 h after transfection of cells with or without adenovirus at 200 of MOI, cells were washed three times with PBS and cultured in complete DMEM medium. After 48 h culture, the supernatants were collected for isolation of Exo/IL-18 or Exo/LacZ or Exo. For further exosomes purification, the exosomes suspended in PBS were layered onto discontinuous density cushion (30–45% sucrose, w/v) in 20 mM Hepes/NaOH, pH7.4 (1.13–1.19 g/ml) and centrifuged at 100,000×g for 2 h. Exosomes accumulated at the interface were collected and then washed with PBS. Exosome proteins were measured by the Bradford assay (BioRad). Endotoxin contamination in exosomes was analyzed by limulus amebocyte lysate (BioWhittaker, Walkersville, MD) to be less than 0.05 EU/μg exosome protein.
Western blot assay
Western blot assay of the components in exosomes was performed as described previously by us [28]. Primary antibodies used here were: anti-CEA (clone CE05, neomarker, 47790 Westinghouse Dr., Fremont, CA, USA), anti-HSP27 (F-4), anti-HSP40 (C-20), anti-HSP70 (K-20), anti-Hsc70 (B6), anti-ICAM-1 (C-19), anti-B7.1 (N-20), anti-B7.2 (C-19), anti-transferrin receptor (TfR, K-20), anti-LAMP-3 (MX-49.129.5, Santa Cruz Biotechnology, Inc, Santa Cruz, CA) and anti-MHC-2 (HC10, Soldano Ferrone, Roswell Park Cancer Institute, NY, USA).
In vitro PBMC proliferation assay
PBMC proliferation assay was conducted as described with minor modifications [29]. Briefly, 2×105 PBMC per well were added to 96-well round-bottom culture plates and cultured with exosomes in a final volume of 200 μl RPMI1640 supplemented with 10% FCS at 37°C in 5% CO2 for 72 h in the presence of PHA (20 μg/ml). [3H] thymidine (0.5 μCi/well) was added to the cells for an additional 18 h. The cells were harvested onto filters and counted by liquid scintillation counting (Wallac 1400 DSA, USA). In parallel, after 72 h culture, the supernatants were harvested and tested for IL-2 production by ELISA (Endogen, Cambridge, MA).
Induction of in vitro DC maturation by exosomes
Monocyte-derived DC from healthy individuals were generated as described [30]. TNF-α (1 ng/ml) and PGE2 (500 nM) (Cayman Chemical, Ann Arbor, MI) were used to induce DC maturation. Immature DC (iDC) on day five were cultured with 5 μg/ml exosomes or 1 μg/ml LPS or medium for an additional 48 h and then resuspended in 50 μl PBS. DC were stained with FITC-labeled anti-CD80, PE-labeled anti-CD86, PE-labeled anti-CD40, PerCP-labeled anti-HLA-DR, and analyzed on FACScan. In parallels, the supernatants were harvested and measured for IL-12p70 and TNF-α concentrations using commercially available ELISA kits (Endogen, Cambridge, MA) [31].
Mixed lymphocyte reaction (MLR)
MLR was conducted as previously described [32]. Briefly, iDC on day five were collected and incubated with 5 μg/ml exosomes, medium or 1 μg/ml LPS for 48 h, and were then used as stimulator after irradiation (5000 rad). Allogeneic T cells, isolated by anti-CD3 magnetic beads (Miltenyi Biotec, Auburn, CA), were then stimulated with the irradiated DC at different responder/stimulator ratios. After 3 days culture, [3H] thymidine (0.5 μCi/well) was added to each well, and the incorporated radioactivity was measured 18 h later.
Chemoattraction assay of DC and T cells
Chemoattraction of DC and T cells in response to exosomes was investigated in 24-well trans-well culture plates as described with minor modification [33]. Briefly, CD4+ or CD8+ T cell populations were positively purified from PBMC using the anti-CD4 or anti-CD8 magnetic beads (Miltenyi Biotec, Auburn, CA). Purity of CD4+ or CD8+ T cells was more than 90% as judged by flow cytometry analysis. 1×106 cells/ml purified CD4+ or CD8+ T cells, iDC on day five, or mature DC (mDC) on day seven in 200 μl medium were added to the upper chamber of each trans-well. The chambers were incubated for 4 h at 37°C. The number of cells that chemoattracted into the bottom well was counted by FACS. Results were expressed as the chemotactic index: the quotient of the number of chemoattracted cells in the presence of exosomes/chemoattracted cells in the presence of medium.
Induction of HLA-A*0201-restricted antigen-specific CTL from PBMC of cancer patients
PBMC from HLA-A*0201 positive cancer patients were isolated by Ficoll-Hypaque density gradient centrifugation. On day five of DC culture, 2×105 DC/well were pulsed with 10 μg exosomes in 24-well plates for 2 h in 200 μl of culture medium, and then were co-cultured with autologous T cells at a ratio of 1:10 in a final volume of 2 ml/well in the presence of 10 ng/ml recombinant human IL-7 (Peprotech Inc., Rocky Hill, NJ). CTL were induced as previously described [34]. 7 days after the final stimulation, functions of the stimulated lymphocytes were evaluated by IFN-γ release assay, cytotoxicity assay and HLA-A*0201/YLSGANLNL tetramer assay (ProImmune, Oxford BioBusiness Centre, Littlemore Park).
For IFN-γ release assay, stimulator cells (T2 cells) were pulsed with 50 μg/ml CAP-1 (T2/CAP-1) or SSp-1 (T2/SSp-1). The supernatants of 5×104 effector cells co-cultured with 1×104 T2/CAP-1 or T2/SSp-1 or native T2 were tested for the level of IFN-γ by ELISA assay (R&D systems) [35].
Cytotoxicity assays were performed using a standard 4-h 51Cr release assay as described [34]. CD8+ T cells were obtained by negative selection after four rounds of in vitro stimulation. Targets for CTL activity were SW480 and LoVo cells. All target cells were harvested, washed, counted and labeled with 100 μCi 51Cr/106 target cells (Amersham, Arlington Heights, IL). Labeled target cells (104 cells/well) were washed three times before use and cocultured with CD8+ T cells at various E:T ratios. Each assay was performed in triplicate.
Statistics
Results are expressed as the means±SEM or the means±SD. Comparisons between two groups were conducted by Student’s “t” test. Multiple comparisons were done with a one-way ANOVA followed by Fisher’s least significant difference analysis. Pairwise comparisons were done by performing nonparametric Mann–Whitney U test. Most of the experiments were repeated at least twice. P values of 0.05 or less were considered significant.
Results
Identification of the recombinant adenovirus AdhIL-18
LS-174T cells genetically modified with AdhIL-18 at a different MOI secreted different levels of hIL-18 at different times. A maximal hIL-18 production by the modified cells was observed 72 h after AdhIL-18 modification at 200 MOI (Fig. 1a), and the hIL-18 concentrations ranged from about 400 pg/ml at 100 MOI to 1000 pg/ml at 200 MOI. LS-174T cells transfected with the control adenovirus AdLacZ secreted no hIL-18 (Fig. 1b). Moreover, the supernatants from AdhIL-18 modified LS-174T cells could induce PBMC to release IFN-γ (Fig. 1c), suggesting that hIL-18 in the supernatants is functionally active. These results showed that the recombinant adenovirus AdhIL-18 was successfully constructed.

Identification of the recombinant adenovirus AdhIL-18. a Time course of IL-18 production after adenovirus infection (200 MOI) of LS-174T cells. b Dose-dependent production of IL-18 after adenovirus infection of LS-174T cells. c Bioactivity of hIL-18. The supernatants from either wild type LS-174T cells (WT) or tumor cells transduced with AdhIL-18 (AdhIL-18) or with AdLac Z (AdLacZ) at 200 MOI 72 h after infection were harvested and co-cultured with PBMCs in the presence of Con A for 48 h. Results were presented as the means±SEM and were representative of three independent experiments
Characterization of exosomes isolated from AdhIL-18 transfected tumor cells
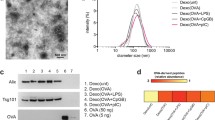
When LS-174T cells were transfected with AdhIL-18, the growth rate of these cells was not affected by the transfection (data not shown). Exo/IL-18 were isolated from the culture supernatant of AdhIL-18 transfected LS-174T cells. Exo/IL-18 were a heterogenous population of 30–90 nm vesicles in diameter when observed under electron microscopy (data not shown), which was consistent with the morphology of typical exosomes [3, 28]. Western blot analysis showed that Exo, Exo/LacZ and Exo/IL-18 all contained CEA, HSP70, Hsc70, TfR, LAMP-3 and MHC-2 while hIL-18 was only detected in Exo/IL-18 (Fig. 2). These results indicated that hIL-18 was sorted into Exo/IL-18, which contains tumor antigen (CEA) and HSPs.

Western blot assay of the protein components of exosomes. Exosomal proteins or tumor cell lysates were analyzed by Western blot using antibodies directed against IL-18, CEA, Hsc70, HSP70, HSP27, HSP40, MHC-I, B7.1, B7.2, ICAM-1, LAMP-3 and TfR
Exo/IL-18 efficiently promote PBMC proliferation and Th1 cytokine production
PBMC incubated with Exo/IL-18 significantly secreted higher levels of IFN-γ than that incubated with the control exosomes (Fig. 3a) (P<0.01), indicating that the hIL-18 in Exo/IL-18 was bioactive and could enhance IFN-γ release from PBMC in the presence of Con A. We also tested the proliferation and IL-2 secretion of Exo/IL-18-treated PBMC. We found that Exo/IL-18 augmented the proliferation (Fig. 3b, P<0.01) and IL-2 secretion of PBMC in the presence of PHA (Fig. 3c, P<0.05) while no effects were observed in the absence of PHA. Meanwhile, the control exosomes including Exo and Exo/LacZ could not demonstrate similar effects, indicating that these effects of Exo/IL-18 should be ascribed to the IL-18 sorted into Exo/IL-18 through genetic modification.

In vitro effects of exosomes on proliferation and Th1 cytokine production of PBMCs. a Exo/IL-18 induces IFN-γ release of PBMCs. PBMCs were co-cultured with indicated exosomes in the presence of Con A for 48 h to induce the release of IFN-γ. b and c Exo/IL-18 promotes the proliferation and IL-2 production of PBMCs. Cells cultured without PHA (medium) were used as negative control. In a and c, results were presented as the mean±SEM; In b, results were presented as the mean±SD of triplicate experiments. *, P<0.01; ▲, P<0.05
Exo/IL-18 potently induce phenotypic and functional maturation of DC
It was reported that exosomes derived from different origins may have different effects on DC maturation [31]. So we examined the effects of Exo/IL-18 on DC. We observed that Exo/IL-18 could efficiently upregulate the expression of HLA-DR, CD40 and CD86 on DC (Fig. 4a) and increase the cytokine production by DC (Fig. 4b, P<0.05). As a control for DC maturation, we exposed DC to LPS, and we found that Exo/IL-18-pulsed DC secreted IL-12p70 and TNF-α comparably to those induced by LPS (Fig. 4b). Our results of MLR also revealed that Exo/IL-18-treated DC were more potent in inducing T cell proliferation than DC treated with Exo or Exo/LacZ (Fig. 4c, P<0.05 at 1:5 or 1:20 of stimulator/responder ratios). These data suggested that Exo/IL-18 could induce phenotypic and functional maturation of DC, and Exo/IL-18-pulsed DC had acquired more potent capability in stimulating T cell proliferation.

Exo/IL-18 induces phenotypic and functional maturation of human DC. a Exo/IL-18 induces phenotypic maturation of DC. Positive (1 μg/ml LPS) and negative controls (medium) for DC maturation were used as indicated. After 48 h of incubation, the cells were collected and stained for HLA-DR, CD80, CD86 and CD40. b Exo/IL-18 promotes cytokine production of DC. Supernatants collected from the cells in a were measured for their content of IL-12p70 and TNF-α. c Exo/IL-18-pulsed DC exhibit potent allo-stimulatory capacity. iDC on day five were incubated with 5 μg/ml Exo, Exo/IL-18, Exo/LacZ or medium for 48 h, irradiated as stimulator cells and then cultured with allogeneic T cells at different responder/stimulator ratios as indicated. Results were presented as the mean±SD or the mean±SEM. Data were representative of three separate experiments. MFI, mean fluorescence intensity; *, P<0.01; ▲, P<0.05
Exo/IL-18 can chemoattract DC and T cells
We further tested whether Exo/IL-18 had chemotactic effects on DC and T cells. At first, we performed a dose-dependent experiment to test the chemotactic activity of Exo/IL-18 to mature DC (mDC). We found that chemotactic indexes increased with the concentration of Exo/IL-18 at the range of 5–50 μg/ml, while Exo or Exo/LacZ had no chemotactic effects even at higher concentrations (Fig. 5a). We then examined the chemoattraction of CD4+, CD8+, iDC or mDC by 50 μg/ml of Exo, Exo/LacZ or Exo/IL-18. Results showed that Exo/IL-18 had significant chemotactic effects for these immune cells to different extents (Fig. 5b), indicating that Exo/IL-18 could markedly chemoattract T cells and DC, which was consistent with the report that IL-18 has indirect and direct chemotactic effects on DC [36, 37] and T cells [18, 38]. Therefore, our data suggested that Exo/IL-18 had superior advantages over conventional exosomes derived from parent tumor cells.

Exo/IL-18 can chemoattract DC and T cells in vitro. a Dose-dependent chemoattraction of immature DC (iDC) by exosomes as indicated. b Exo/IL-18 can chemoattract T cells, immature DC (iDC) and mature DC (mDC). Results were presented as the mean±SEM of chemotactic indexes, and were representative of three independent experiments
Exo/IL-18-pulsed DC significantly promote the induction of CEA-specific CD8+ CTL from PBMC of HLA-A*0201 CEA+ cancer patients
We conducted in vitro CTL induction experiments to test the possibility and efficacy of applying Exo/IL-18 in tumor immunotherapy. Because Exo/IL-18 was purified from CEA-positive LS-174T cells and contained CEA, we isolated DC and PBMC from HLA-A*0201 CEA+ cancer patients and tested the efficiency of Exo/IL-18-pulsed DC in the induction of CEA-specific CTL. CEA-specific CTL were judged by the lysis of CEA+ target cells, IFN-γ production upon CEA epitope-pulsed T2 stimulation, and CEA epitope-specific tetramer staining. Results of target cell lysis showed that CTL induced by Exo/IL-18-pulsed DC were more potent in killing SW480 cells (CEA+, HLA-A2+) (Fig. 6a, P<0.05) than that induced by Exo-pulsed or Exo/LacZ-pulsed DC, but they were less efficient in killing LoVo cells (CEA+, HLA-A2−) (Fig. 6b), which together suggested that the observed lysis was CEA-specific in a HLA-A*0201-restricted manner. Correspondingly, CTL induced by Exo/IL-18-pulsed DC produced more IFN-γ after stimulation with T2/CAP-1 than that induced by Exo-pulsed or Exo/LacZ-pulsed DC (Fig. 6c, P<0.01). In the tetramer staining assay, CAP-1 tetramer+CD8+ T cells were detected after 4 cycles of in vitro restimulation with Exo/IL-18-pulsed DC (8.67%) or Exo-pulsed DC (5.12%) or Exo/LacZ-pulsed DC (4.42%) (Fig. 6d). It should be noted that PBMC stimulated by DC loaded with Exo/IL-18 exhibited a higher percentage of CD8+ lymphocytes (up to 51%), compared to that stimulated by DC loaded with Exo (38%) or Exo/LacZ (35%) (Fig. 6d), suggesting that hIL-18 in Exo/IL-18 could promote the development of CD8+ T cells [11]. In our study, CEA-specific CD8+ CTL could be readily generated in three of four HLA-A*0201 CEA+ cancer patients.

Exo/IL-18-pulsed DC induce generation of HLA-A*0201-restricted CEA-specific CD8+ CTL from PBMC of HLA-A*0201 CEA+ cancer patients. a and b Cytotoxic assay of CTL. Targets included SW480 and LoVo cells. Data shown are representative of three independent experiments. c IFN-γ release assay. Results were presented as the mean±SEM. d FACS analysis of HLA-A*0201/CAP-1 tetramer binding lymphocytes. The effector T cell populations generated after serial restimulations with indicated exosomes-pulsed DC were stained with HLA-A*0201/CAP-1 tetramersPE or HLA-A*0201/SSp-1 tetramersPE and anti-CD8FITC mAbs. Quadrants were set based on negative controls. Results are representative of data from one of three responsive patients. *, P<0.01; ▲, P<0.05; ▼, P>0.05
Discussion
In this study, we have prepared a new kind of exosomes, namely, exosomes derived from AdhIL-18 gene-modified tumor cells (Exo/IL-18), into which bioactive IL-18 was successfully incorporated. Exo/IL-18 can promote PBMC to proliferate and secrete Th1 cytokines, chemoattract DC and T cells in vitro and enhance phenotypic and functional maturation of DC more efficiently than conventional exosomes. More importantly, Exo/IL-18-pulsed DC are quite potent to induce tumor antigen-specific CTL in vitro when co-cultured with autologous lymphocytes. Therefore, Exo/IL-18 demonstrates distinct properties to the conventionally prepared exosomes derived from parent tumor cells, and thus may represent a new approach to design more effective tumor vaccines in the future.
Recombinant cytokines such as IL-2, IFN-γ, and GM-CSF as immunoadjuvants were used in cancer treatments, but their systemic toxicities limited their use [7]. However, modifying tumor cells with cytokine genes can decrease systemic toxicities through locally secreting cytokines, and cytokine-loaded microspheres has the clinical potential of developing systemic anti-tumor immunity [39]. The strategy of natural incorporation of cytokines into exosomes may be better than that of simply mixing cytokines with exosomes for anti-tumor immune induction in that cytokine modified exosomes have combined the advantages of exosomes-based cancer immunotherapy with cytokine-mediated gene therapy. Therefore, incorporating cytokines into exosomes may be an important approach for improving the efficiency of exosomes-based vaccines. It was reported that CEA vaccination could be augmented by co-administration of GM-CSF [40, 41]. However, locally administrated cytokines are rapidly redistributed into the circulation, thus leading to many undesirable side-effects and quick clearance. Membrane-bound cytokines are proposed as appropriate adjuvants for vaccination because they are restricted to its administration sites [42]. Natural sorting of bioactive IL-18 into exosomes makes Exo/IL-18 as “IL-18-containing membrane compartments” that are supposed to slowly release IL-18. Results of our in vitro experiments have demonstrated that Exo/IL-18 containing both CEA and IL-18 have potent capability of inducing PBMC to proliferate and secrete Th1 cytokines, chemoattracting DC and T cells, promoting phenotypic and functional maturation of DC and inducing CEA-specific CD8+ CTL. Therefore, these multiple effects of Exo/IL-18 suggest that Exo/IL-18 may potentially function well to induce potent specific anti-tumor immunity in vivo, which need further extensive investigations.
Although in this study we have not directly applied Exo/IL-18 in an animal model in vivo due to species barrier, we suggest that IL-18 in Exo/IL-18 might serve as an immunoadjuvant with multiple advantages in inducing anti-tumor immunity in vivo. First, Exo/IL-18 can chemoattract DC and T cells and thus potentially accumulate these immune cells locally at the sites of administration [16, 17]. Secondly, IL-18 contained in Exo/IL-18 may induce the release of IFN-γ, leading to the final elimination of tumor cells [8–13]. Thirdly, IL-18 augments the release of IL-2 and promotes T cell proliferation [43], supporting the idea that immune molecules such as IL-18, IFN-γ and IL-2 can influence the local environment in a way that would be beneficial for the patients. Therefore, it is reasonable to predict that Exo/IL-18 may be functionally effective in the induction of anti-tumor immunity when administrated locally in vivo.
The identification of the presence of IL-18 in exosomes provides critical insights into the improvement of tumor-derived exosomes. Exosomes expressing cytokines such as IL-18 as adjuvants can thus become one of the attractive strategies in combating cancer. Using this relatively simple strategy, we can incorporate the cytokines, chemokines, adhesion molecules, co-stimulatory molecules or their combinations into exosomes, through which we can obtain cancer vaccines providing sufficient immunogenic molecules for effective cancer immunotherapy in future trials.
Abbreviations
- DC:
-
dendritic cells
- CEA:
-
carcinoembryonic antigen
- AdhIL-18:
-
recombinant adenovirus encoding human IL-18
- Exo:
-
exosomes derived from tumor cells
- Exo/IL-18:
-
exosomes derived from AdhIL-18-transfected tumor cells
- MLR:
-
mixed lymphocyte reaction
- CTL:
-
cytotoxic T lymphocytes
- MOI:
-
multiplicity of infection
- PBMC:
-
peripheral blood mononuclear cells
References
Minev B, Chavez F, Mitchell M (1998) New trends in the development of cancer vaccines. In Vivo 12:629–638
Zitvogel L, Regnault A, Lozier A, Wolfers J, Flament C, Tenza D, Ricciardi-Castagnoli P, Raposo G, Amigirena S (1998) Eradication of established murine tumors using a novel cell-free vaccine: dendritic cell-derived exosomes. Nat Med 4:594–600
Wolfers J, Lozier A, Raposo G, Regnault A, Thery C, Masurier C, Flament C, Pouzieux S, Faure F, Tursz T, Angevin E, Amigorena S, Zitvogel L (2001) Tumor-derived exosomes are a source of shared tumor rejection antigens for CTL cross-priming. Nat Med 7:297–303
Escudier B, Dorval T, Chaput N, Andre F, Caby MP, Novault S, Flament C, Leboulaire C, Borg C, Amigorena S, Boccaccio C, Bonnerot C, Dhellin O, Movassagh M, Piperno S, Robert C, Serra V, Valente N, Le Pecq JB, Spatz A, Lantz O, Tursz T, Angevin E, Zitvogel L (2005) Vaccination of metastatic melanoma patients with autologous dendritic cell (DC) derived-exosomes: results of the first phase I clinical trial. Journal of Translational Medicine 3:10
Morse MA, Garst J, Osada T, Khan S, Hobeika A, Clay TM, Valente N, Shreeniwas R, Sutton MA, Delcayre A, Hsu DH, Le Pecq JB, Lyerly HK (2005) A phase I study of dexosome immunotherapy in patients with advanced non-small cell lung cancer. Journal of Translational Medicine 2005 3:9
Chaput N, Schartz N, Andre F, Taieb J, Novault S, Bonnaventure P, Aubert N, Bernard J, Lemonnier F, Merad M, Adema G, Adams M, Ferrantini M, Carpentier A, Escudier B, Tursz T, Angevin E, Zitvogel L (2004) Exosomes as potent cell-free peptide-based vaccine. II. Exosomes in CpG adjuvants efficiently prime naive Tc1 lymphocytes leading to tumor rejection. J Immunol 172:2137–2146
Dredge K, Marriott J, Todryk S, Dalgleish A (2002) Adjuvants and the promotion of Th1-type cytokines in tumour immunotherapy. Cancer Immunol Immunother 51:521–531
Okamura H, Tsutsi H, Komatsu T, Yutsudo M, Hakura A, Tanimoto T, Torigoe K, Okura T, Nukada Y, Hattori K, Akita K, Namba M, Tanabe F, Konishi K, Fukuda S, Kurimoto M (1995) Cloning of a new cytokine that induces IFN-γ production by T cells. Nature 378:88–91
Okamura H, Nagata K, Komatsu T, Tanimoto T, Nukata Y, Tanabe F, Akita K, Torigoe K, Okura T, Fukuda S, Kurimoto M (1995) A novel costimulatory factor for g interferon induction found in the livers of mice causes endotoxic shock. Infect Immun 63:3966–3972
Micallef M, Ohtsuki T, Kohno K, Tanabe F, Ushio S, Namba M, Tanimoto T, Torigoe K, Fujii M, Ikeda M, Fukuda S, Kurimoto M (1996) Interferon-gamma-inducing factor enhances T helper 1 cytokine production by stimulated human T cells: synergism with interleukin-12 for interferon-gamma production. Eur J Immunol 26:1647–1651
Okamoto I, Kohno K, Tanimoto T, Ikegami H, Kurimoto M (1999) Development of CD8+ effector T cells is differentially regulated by IL-18 and IL-12. J Immunol 162:3202–3211
Dao T, Mehal W, Crispe I (1998) IL-18 augments perforin-dependent cytotoxicity of liver NK-T cells. J Immunol 161:2217–2222
Hashimoto W, Osaki T, Okamura H, Robbins P, Kurimoto M, Nagata S, Lotze M, Tahara H (1999) Differential anti-tumor effects of administration of rIL-18 or rIL-12 are mediated primarily by Fas–Fas ligand- and perforin-induced tumor apoptosis, respectively. J Immunol 163:583–589
Li J, Lamine Mbow M, Sun L, Li L, Yang G, Griswold D, Schantz A, Shealy D, Goletz T, Wan J, Peritt D (2004) Induction of dendritic cell maturation by IL-18. Cell Immunol 227:103–108
Puren A, Razeghi P, Fantuzzi G, Dinarello C (1998) Interleukin-18 enhances lipopolysaccharide-induced interferon-gamma production in human whole blood cultures. J Infect Dis 178:1830–1834
Komai-Koma M, Gracie J, Wei X, Xu D, Thomson N, McInnes I, Liew F (2003) Chemoattraction of human T cells by IL-18. J Immunol 170:1084–1090
Gutzmer R, Langer K, Mommert S, Wittmann M, Kapp A, Werfel T (2003) Human dendritic cells express the IL-18R and are chemoattracted to IL-18. J Immunol 171:6363–6371
Puren A, Fantuzzi G, Gu Y, Su M, Dinarello C (1998) Interleukin-18 (IFNγ-inducing factor) induces IL-8 and IL-1β via TNFα production from non-CD14+ human blood mononuclear cells. J Clin Invest 101:711–721
Yoshida Y, Tasaki K, Kimurai M, Takenaga K, Yamamoto H, Yamaguchi T, Saisho H, Sakiyama S, Tagawa M (1998) Antitumor effect of human pancreatic cancer cells transduced with cytokine genes which activate Th1 helper T cells. Anticancer Res 18:333–335
Heuer J, Tucker-McClung C, Hock R (1999) Neuroblastoma cells expressing mature IL-18, but not proIL-18, induce a strong and immediate antitumor immune response. J Immunother 22:324–335
Ju D, Tao Q, Lou G, Bai M, He L, Yang Y, Cao X (2001) Interleukin 18 transfection enhances anti-tumor immunity induced by dendritic cell–tumor cell conjugates. Cancer Res 61:3735–3740
Wang Q, Yu H, Ju D, He L, Pan J, Xia D, Zhang L, Cao X (2001) Intratumoral IL-18 gene transfer improves therapeutic efficacy of antibody-targeted superantigen in established murine melanoma. Gene Ther 8:542–550
Xia D, Zheng S, Zhang W, He L, Wang Q, Pan J, Zhang L, Wang J, Cao X (2003) Effective induction of therapeutic anti-tumor immunity by dendritic cells coexpressing interleukin-18 and tumor antigen. J Mol Med 81:585–596
Coughlin C, Salhany K, Wysocka M, Aruga E, Kurzawa H, Chang A, Hunter C, Fox J, Trinchieri G, Lee W (1998) Interleukin-12 and interleukin-18 synergistically induce murine tumor regression which involves inhibition of angiogenesis. J Clin Invest 101:1441–1452
He T, Zhou S, da Costa L, Yu J, Kinzler K, Vogelstein B (1998) A simplified system for generating recombinant adenoviruses. Proc Natl Acad Sci USA 95:2509–2514
Théry C, Regnault A, Garin J, Wolfers J, Zitvogel L, Ricciardi-Castagnoli P, Raposo G, Amigorena S (1999) Molecular characterization of dendritic cell-derived exosomes: selective accumulation of the heat shock protein hsc73. J Cell Biol 147:599–610
Raposo G, Nijman H, Stoorvogel W, Leijendekker R, Harding C, Melief C, Geuze H (1996) B lymphocytes secrete antigen-presenting vesicles. J Exp Med 183:1–12
Dai S, Wan T, Wang B, Zhou X, Xiu F, Chen T, Wu Y, Cao X (2005) More efficient induction of HLA-A*0201-restricted and CEA-specific CTL response by immunization with exosomes prepared from heat-stressed CEA-positive tumour cells. Clin Cancer Res 11:7554–7563
Vazirabadi G, Geiger T, Coffin W, Martin J (2003) Epstein–Barr virus latent membrane protein-1 (LMP-1) and lytic LMP-1 localization in plasma membrane-derived extracellular vesicles and intracellular virions. J Gen Virol 84:1997–2008
Liu S, Yu Y, Zhang M, Wang W, Cao X (2001) The involvement of TNF-alpha-related apoptosis-inducing ligand in the enhanced cytotoxicity of IFN-beta-stimulated human dendritic cells to tumor cells. J Immunol 166:5407–5415
Skokos D, Botros HG, Demeure C, Morin J, Peronet R, Birkenmeier G, Boudaly S, Me´cheri S (2003) Mast cell-derived exosomes induce phenotypic and functional maturation of dendritic cells and elicit specific immune responses In vivo. J Immunol 170:3037–3045
Wan T, Zhou X, Chen G, An H, Chen T, Zhang W, Liu S, Jiang Y, Yang F, Wu Y, Cao X (2004) Novel heat shock protein Hsp70L1 activates dendritic cells and acts as a Th1 polarizing adjuvant. Blood 103:1747–1754
Chantry D, Romagnani P, Raport C, Wood C, Epp A, Romagnani S, Gray P (1999) Macrophage-derived chemokine is localized to thymic medullary epithelial cells and is a chemoattractant for CD3+, CD4+, CD8low thymocytes. Blood 94:1890–1898
Wang B, Chen H, Jiang X, Zhang M, Wan T, Li N, Zhou X, Yang F, Wu Y, Yu Y, Wang X, Yang R, Cao X (2004) Identification of an HLA-A*0201-restricted CD8+ T-cell epitope SSp-1 of SARS-CoV spike Protein. Blood 104:200–206
Kim C, Todoroki T, Matsumura M, Ohno T (2000) Eligibility of antigenic-peptide-pre-loaded and fixed adhesive peripheral blood cells for induction of cytotoxic T lymphocytes from cancer patients with elevated serum levels of carcinoembryonic antigen. J Cancer Res Clin Onco 126:383–390
Morel J, Park C, Kumar P, Koch A (2001) Interleukin-18 induces rheumatoid arthritis synovial fibroblast CXC chemokine production through NF-κB activation. Lab Invest 81:1371–1383
Gutzmer R, Langer K, Mommert S, Wittmann M, Kapp A, Werfel T (2003) Human dendritic cells express the IL-18R and are chemoattracted to IL-18. J Immunol 171:6363–6371
Komai-Koma M, Gracie J, Wei X, Xu D, Thomson N, McInnes I, Liew F (2003) Chemoattraction of human T cells by IL-18. J Immunol 170:1084–1090
Rosenberg S, Yang J, Schwartzentruber D, Marincola H, Topalian F, Restifo S, Dudley N, Schwarz M, Spiess S, Wunderlich P, Parkhurst J, Kawakami M, Seipp Y, Einhorn C, White D (1998) Immunologic and therapeutic evaluation of a synthetic peptide vaccine for the treatment of patients with metastatic melanoma. Nat Med 4:321–327
Ullenhag G, Frodin J, Jeddi-Tehrani M, Strigard K, Eriksson E, Samanci A, Choudhury A, Nilsson B, Rossmann E, Mosolits S, Mellstedt H (2004) Durable carcinoembryonic antigen (CEA)-specific humoral and cellular immune responses in colorectal carcinoma patients vaccinated with recombinant CEA and granulocyte/macrophage colony-stimulating factor. Clin Cancer Res 10:3273–3281
von Mehren M, Arlen P, Gulley J, Rogatko A, Cooper H, Meropol N, Alpaugh R, Davey M, McLaughlin S, Beard M, Tsang K, Schlom J, Weiner L (2001) The influence of granulocyte macrophage colony-stimulating factor and prior chemotherapy on the immunological response to a vaccine (ALVAC-CEA B7.1) in patients with metastatic carcinoma. Clin Cancer Res 7:1181–1191
Chakrabarti R, Chang Y, Song K, Prud’homme G (2004) Plasmids encoding membrane-bound IL-4 or IL-12 strongly costimulate DNA vaccination against carcinoembryonic antigen (CEA). Vaccine 22:1199–1205
Tartour E, Fridman W (1998) Cytokines and cancer. Int Rev Immunol 16:683–704
Acknowledgments
This work was supported by Grants from the National Key Basic Research Program of China (2004CB518807, 2001CB510002) and the National Natural Science Foundation of China (30328011, 30490240, 30121002). We appreciate the technical assistance of Ms. Jun Li, Ms. Linhong Sun and Mr. Xiaohui Huang. We thank Dr. Guisheng Li for obtaining the blood samples of the cancer patients.
Author information
Authors and Affiliations
Corresponding author
Additional information
S. Dai and X. Zhou contributed equally to this study.
Rights and permissions
About this article
Cite this article
Dai, S., Zhou, X., Wang, B. et al. Enhanced induction of dendritic cell maturation and HLA-A*0201-restricted CEA-specific CD8+ CTL response by exosomes derived from IL-18 gene-modified CEA-positive tumor cells. J Mol Med 84, 1067–1076 (2006). https://doi.org/10.1007/s00109-006-0102-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00109-006-0102-0