
Abstract
Chikungunya, a mosquito-borne viral disease caused by Chikungunya virus (CHIKV), has drawn substantial attention after its reemergence causing massive outbreaks in tropical regions of Asia and Africa. The recombinant envelope 2 (rE2) protein of CHIKV is a potential diagnostic as well as vaccine candidate. Development of cost-effective cultivation media and appropriate culture conditions are generally favorable for large-scale production of recombinant proteins in Escherichia coli. The effects of medium composition and cultivation conditions on the production of recombinant Chikungunya virus E2 (rCHIKV E2) protein were investigated in shake flask culture as well as batch cultivation of Escherichia coli. Further, the fed-batch process was also carried out for high cell density cultivation of E. coli expressing rE2 protein. Expression of rCHIKV E2 protein in E. coli was induced with 1 mM isopropyl-beta-thiogalactoside (IPTG) at ~23 g dry cell weight (DCW) per liter of culture and yielded an insoluble protein aggregating to form inclusion bodies. The final DCW after fed-batch cultivation was ~35 g/l. The inclusion bodies were isolated, solubilized in 8 M urea and purified through affinity chromatography to give a final product yield of ~190 mg/l. The reactivity of purified E2 protein was confirmed by Western blotting and enzyme-linked immunosorbent assay. These results show that rE2 protein of CHIKV may be used as a diagnostic reagent or for further prophylactic studies. This approach of producing rE2 protein in E. coli with high yield may also offer a promising method for production of other viral recombinant proteins.
Similar content being viewed by others

Introduction
Chikungunya has now emerged as an important public health problem of Asia and African countries. Chikungunya is an acute viral illness characterized by high fever, rashes and polyarthralgia (Yathi et al. 2013). CHIKV is a positive sense, single-stranded RNA virus of approximately 12 kb genome. It encodes four non-structural proteins (nsP1-4) and five structural proteins (capsid and four envelope proteins E1, E2, E3 and 6 K) (Kumar et al. 2012a, 2012b). The Chikungunya virus envelope proteins E1 and E2 are components of spikes, which composed of triplets of heterodimer of E1 and E2 glycoproteins, and cover the viral surface in the form of membrane-anchored types. The viral spike proteins facilitate attachment to cell surfaces and viral entry into the cells. The E2 envelope protein is a type I transmembrane glycoprotein and has been known to be responsible for receptor binding during the course of the alphavirus cycle (Kielian and Rey 2006; Brehin et al. 2008; Yathi et al. 2011; Yathi et al. 2013). The envelope 2 (E2) protein elicits neutralizing antibodies and plays an important role in inducing immunogenic responses in the infected host (Khan et al. 2012; Kumar et al. 2012a, 2012b). The high immunogenicity of the envelope 2 protein of Chikungunya virus has raised considerable interest both as an antigen for diagnostic methods (Brehin et al. 2008; Cho et al. 2008a, 2008b; Kumar et al. 2012a; Khan et al. 2013) and as component of subunit vaccine formulations (Kumar et al. 2012b; Khan et al. 2012; Metz et al. 2011, 2013). For this reason, the envelope 2 protein of Chikungunya virus is an important immunogen for a subunit vaccine and also a prospective diagnostic reagent for the improved clinical diagnosis of CHIKV infections.
Currently, no licensed vaccine or specific therapeutics are available for Chikungunya virus infection. Following the recent resurgence of the disease and with the availability of newer platforms for vaccine development, different approaches have been tried. So far, none of the vaccines have been licensed (Kumar et al. 2012b). Early diagnosis of disease plays an important role to forecast an early warning of epidemic and to undertake effective vector control measures. Virus isolation, RT-PCR and ELISA have been recognized as laboratory tools for the specific detection of Chikungunya virus infection in patient samples (Yathi et al. 2011; Kumar et al. 2012a). Though virus isolation and RT-PCR provides accurate diagnosis, the increased cost and sophisticated equipment requirement limit its application to referral lab. The serological test is based on the principle of ELISA in which viral antigen or virus-specific antibodies are detected. Some of the ELISA tests for the detection of anti-Chikungunya virus IgM antibodies utilize whole-virus antigen prepared from cell culture. Since risk factors are associated with the production and purification of whole antigen from isolated viruses, an alternative way of developing recombinant protein-based antigens will be essential for rapid and specific diagnosis of the diseases. Thus, there is a need to develop a detection system as well as an improved Chikungunya vaccine that may be safer, cheaper and readily available. Hence, production and purification of this protein are necessary for further studies.
The production of recombinant antigenic proteins for diagnostic use and subunit formulations is generally carried out using either prokaryotic or eukaryotic expression systems. Prokaryotic expression systems, such as E. coli, are the most commonly used host for heterologous protein production due to the low cost of production, well-characterized genetics and cultivation condition (Babaepour et al. 2010; Huang et al. 2012; Nausch et al. 2013). Hence, the objective of the study is to produce recombinant Chikungunya virus envelope 2 protein with high yield and to develop an effective diagnostic test for the detection of anti-Chikungunya virus IgM antibody. The recombinant protein expression level in E. coli depends on various factors such as medium composition, cultivation conditions, duration of induction, type of inducer, concentration of inducer and mode of cultivation, which can be optimized to achieve recombinant protein with more yield (Manderson et al. 2006; Fong and Wood 2010; Tripathi et al. 2010; Huang et al. 2012). Recombinant E. coli can be grown to high cell densities in complex media, semi-defined and defined media using batch and fed-batch cultivation modes (Manderson et al. 2006; Babaepour et al. 2010; Tripathi et al. 2012a). The media composition and cultivation strategy is crucial for enhancing protein production as well as reduction of inhibitory compound formation (Manderson et al. 2006; Huang et al. 2012). Recombinant protein production in E. coli has been extensively investigated particularly as high cell density fed-batch cultivations employing bioreactor using complex medium. High cell density can be achieved using fed-batch culture by controlling the nutrient feeding via pH, dissolved oxygen (DO) or specific growth rate (Manderson et al. 2006; Khalilzadeh et al. 2008; Bhuvanesh et al. 2010; Tripathi et al. 2012b; da Silva et al. 2013). The purification of recombinant protein is necessary with minimum possible steps to meet the required level of purity for diagnostic, therapeutic or vaccine studies. Among chromatography methods, affinity chromatography is simple, versatile and can be used for improved purification of recombinant proteins in single step (Bhuvanesh et al. 2010; Tan et al. 2010).
There is currently a need for developing a fermentation process for high yield scalable production of recombinant Chikungunya virus envelope 2 (rCHIKV E2) protein in E. coli and development of a simple, safe and cost-effective detection system. In our previous report, envelope 2 protein of Chikungunya virus was cloned, expressed and evaluated for its immunogenicity as well as ability to induce neutralizing antibodies and cell-mediated immune response in immunized animals in combination with different adjuvants (Khan et al. 2012). The present focus of the study on Chikungunya diagnosis is being targeted to develop a simple, rapid and cheap rCHIKV E2 protein-based ELISA for early detection of Chikungunya infections. In order to produce rCHIKV E2 antigen protein in recombinant E. coli with high yield, a fed-batch fermentation process and a simple purification strategy resulting in efficient recovery of the protein from the resultant biomass are necessary. Thus, we have focused on the production of rCHIKV E2 protein by batch and fed-batch fermentations, its purification and characterization. Further, we present data demonstrating its utility as an antigen in in-house microwell plate ELISA for detection of anti-Chikungunya virus IgM antibodies in patient serum samples.
Materials and methods
Microorganism and vector system
The E. coli strain BL21 (DE3) [Novagen, USA] was used as the host for recombinant Chikungunya virus envelope 2 protein expression. The envelope 2 gene (GenBank accession No. EF210157) of CHIKV was amplified using primers (Khan et al. 2012) and cloned downstream of T7 promoter of E. coli expression vector pET28b+ bearing 6× histidine tag at C-terminal to yield plasmid pET + CHIKVE2 as previously described (Khan et al. 2012). The resultant transformants were selected on kanamycin plates. Stock cultures of the microorganisms were maintained in 30 % glycerol (v/v) at −80 °C. The source of viral RNA was DRDE-06 Chikungunya virus strain (GenBank accession No. EF210157).
Culture media and solutions
The culture media and chemicals used in this study were purchased from Becton Dickinson, USA; Sigma, USA and Qiagen, Germany. Luria Bertani broth (tryptone, 10.0 g; yeast extract, 5.0 g; NaCl per liter), terrific broth (tryptone, 12 g; yeast extract, 24 g; di-potassium phosphate, 9.4 g; mono-potassium phosphate, 2.2 g and glycerol 4 ml per liter) and modified terrific broth (TB) media (tryptone, 12 g; yeast extract, 24 g; di-potassium phosphate, 9.4 g; mono-potassium phosphate, 2.2 g; magnesium sulfate, 2.4 g; trace metal solution, 1 ml and glycerol 20 ml per liter) were used as media for shake flasks, batch and fed-batch fermentations. For shake flasks and fermentations, the antibiotic used was 50 μg/ml of kanamycin. For fed-batch fermentation, modified TB medium was used as batch medium, and the feed medium contained yeast extract, 300 g; glycerol, 300 ml; trace metal solution, 4 ml and magnesium sulfate, 10 g with an antibiotic concentration of 0.5 g kanamycin per liter. The composition of trace metal per liter was FeSO4 · 7H2O, 2.8 g; MnCl2 · 4H2O, 2 g; COCl2. 6H2O, 2.8 g; CaCl2.2H2O, 1.5 g; CuCl2 · 2H2O, 0.2 g; ZnSO4 · 7H2O, 0.3 g; H3BO3, 0.02 g and 1 M HCl.
Buffer composition
The buffers used for purification of rCHIKV E2 protein were cell wash buffer (10 mM Tris–HCl pH 8.0, 10 mM EDTA, 100 mM NaCl), cell lysis buffer (10 mM Tris–HCl pH 7.5, 5 mM EDTA, 100 mM NaCl, 100 μg/ml of lysozyme and 1 mM phenylmethylsulfonyl fluoride (PMSF)), inclusion bodies (IB) wash buffer 1 (50 mM NaH2PO4 pH 6.0, 5 mM EDTA, 200 mM NaCl, 0.5 M urea, 1 % TritonX-100), IB wash buffer 2 (50 mM NaH2PO4 pH 6.0, 1 mM EDTA and 1 M NaCl), IB solubilization buffer (10 mM Tris–HCl, pH 8.0, 100 mM NaH2PO4, 100 mM NaCl, 8 M urea), column equilibration buffer (10 mM Tris–HCl, 100 mM NaCl, 100 mM NaH2PO4, 8 M urea, pH 7.5), elution buffer (equilibration buffer at pH 4.5) and diafiltration/dialysis buffer (50 mM phosphate buffer, 250 mM NaCl, pH 5.8).
Shake flask cultures
Different media were examined for their capabilities to support cell growth and CHIKV E2 protein expression by E. coli. A single vial containing 1 ml stock culture from frozen cell bank was thawed and used to inoculate a 250-ml Erlenmeyer shake flask containing 25 ml sterile LB medium supplemented with 50 μg/ml kanamycin. The inoculum was incubated in a shaker incubator (Kuhner, Switzerland) with a shaking speed of 180 rpm at 37 °C for 8 h. The 1 ml grown culture was inoculated in 50 ml of sterile LB, TB and modified TB media supplemented with 50 μg/ml kanamycin in a 250-ml Erlenmeyer flask and incubated at 37 °C for overnight at 180 rpm. The overnight seed cultures were subsequently used to inoculate cultures. For expression, 100 ml of each sterile culture medium was inoculated using 2 ml overnight grown seed culture supplemented with 50 μg/ml kanamycin. The 100 ml culture medium contained LB, TB and modified TB media. In each case, 100 ml cultures in duplicate were grown in 500-ml Erlenmeyer flasks at 37 °C temperature and 200 rpm in an incubator shaker. When culture OD reaches about 0.6–1.0, IPTG was added to a final concentration of 1 mM, and induction was allowed to proceed for 4 h. Cells were then harvested by centrifugation at 6,000 rpm for 20 min at 4 °C, and cell pellets were recovered and frozen at −80 °C. Samples were taken for measurement of optical density at 600 nm (OD600), dry cell weight (DCW) and rCHIKV E2 protein expression. The effect of inducer concentration and duration of induction was also tested. For this purpose, cells were grown in LB medium at 37 °C and 180–200 rpm till OD of about 0.6–0.8 and induced with different IPTG concentrations (0.5, 1.0 and 1.5 mM) with fixed time (4 h) for duration of induction to test the optimum concentration of the inducer. To test the duration of induction, the cells were grown as above and induced with 1 mM IPTG for 1, 2, 3 and 4 h.
Batch and fed-batch fermentation cultures
Recombinant Chikungunya virus envelope 2 protein was produced by batch and fed-batch fermentations. To inoculate each fermentation run, a 50 ml sterile LB culture medium in 250-ml Erlenmeyer flask supplemented with 50 μg/ml kanamycin was inoculated with a 1 ml of stock culture of recombinant cells and grown at 37 °C for 8 h at 180 rpm. About 2.0 ml of this grown culture was aseptically inoculated in a 2-l Erlenmeyer flask containing 400 ml sterile modified TB medium with antibiotic and shaken at 180 rpm for overnight at 37 °C.
For batch fermentations, 250 ml of overnight grown culture was aseptically inoculated into a 6.6-l BioFlo 3000 bioreactor (New Brunswick Scientific, USA) containing 5.0 l sterile modified TB medium with antibiotic. Biocommand Bioprocessing software (New Brunswick Scientific, USA) was used to control the fermentation and recorded all fermentation conditions. During batch fermentations, dissolved oxygen (DO) was maintained at 30 % of saturation and was controlled by a dissolved oxygen cascade of agitation up to 500 rpm and pure oxygen supply as required. The agitation, pH, temperature and air flow rate were initially set to 200 rpm, 7.0, 37 °C and 4 l/min. Culture pH was controlled between 6.8 and 7.0 using 25 % ammonium hydroxide. Foaming was prevented by use of antifoam agent during fermentation. For batch fermentations, cultures were grown for five and half hours after inoculation, after which the culture was induced with 1 mM IPTG. Cells were harvested after 4 h of induction. Sampling was performed at regular interval to measure OD600. The batch fermentation process was also carried out using TB medium, which is an 8-h (4 h for pre-induction and 4 h for post-induction) process.
For fed-batch fermentations, 200 ml of overnight grown culture was aseptically inoculated in a 6.6-l BioFlo 3000 bioreactor containing 4.0 l sterile modified TB medium with antibiotic. The cultivations were performed with the following set-up: air flow rate, 3.0–7.0 l/min; agitation, 200–800 rpm; pH, 6.8–7.0 and temperature, 37 °C. The dissolved oxygen, pH and air flow rate were recorded during the process. Dissolved oxygen was controlled at ~30 % of air saturation using agitation between 200 and 800 rpm and aeration between flow rates of 3 and 7 l/min of air as well as using pure oxygen as required at high cell densities. Cells were allowed to grow in batch mode at 37 °C until glycerol was depleted from the media, indicated by a sharp increase in dissolved oxygen. At this point (after ~ 5 h of inoculation), the base feed was turned off and feeding was initiated with feeding medium containing yeast extract, glycerol and magnesium sulfate with trace metal solutions supplemented with antibiotic. Feed solutions were added using a built-in peristaltic pump in the bioreactor console to the vessel in a linearly increasing manner from 1.0 to 2.0 ml/min over a 9-h period. If oxygen levels fell below 20 %, the feed was stopped until oxygen levels returned. A combination of pH stat and DO stat feedback mechanism regulated the feed rate by feeding in a control volume of feed in response to the increase in pH and DO of the culture. After 10 h of growth, the culture was induced with 1 mM IPTG to initiate protein expression, and cells were further grown for 4 h. Sampling was done on hourly basis to measure OD during the course of the fermentations.
Cell harvest and cell lysis
The fermentation culture was harvested by centrifugation at 6,000 rpm for 20 min at 4 °C using floor model centrifuge (Sorvall Evolution RC), and the cell pellet was recovered. The cell pellet was washed twice with cell wash buffer (1:20, w/v) by centrifugation at 6,000 rpm for 20 min at 4 °C. The washed cell pellet was stored at −80 °C for further purification processes. The washed cell pellet was resuspended in cell lysis buffer (1:20, w/v). The cell suspension was disrupted by sonication using a high-gain probe of sonicator (Sonics, USA) for a total of 10 min with 9 s on/off pulses. The cell lysate was subjected to high-speed centrifugation (10,000 rpm) for 30 min at 4 °C. The pellet containing IBs was recovered and the supernatant was discarded.
Inclusion body solubilization
The IB-containing pellet was washed by resuspending in IB wash buffer 1(1:20, w/v) and centrifuged at 10,000 rpm for 30 min at 4 °C. The IB pellet was again washed with IB wash buffer 2 (1:20, w/v) and centrifuged as above. The supernatant was carefully discarded and the IB washed pellet containing inclusion bodies was recovered for further use. Inclusion bodies were solubilized by homogenization in IB solubilization buffer (1:20, v/w) using a mechanical homogenizer (Kinematica AG, Switzerland) for about 20 min. This suspension was then centrifuged at 10,000 rpm for 40 min at 4 °C. The clear supernatant was recovered and further clarified by lab-scale tangential flow filtration using a 0.45-μm pore size membrane (Millipore, USA).
Immobilized metal affinity chromatography and diafiltration
The filtrate solution containing rCHIKV E2 protein was purified by immobilized metal affinity chromatography (Tripathi et al. 2012b). All chromatography was carried out using an Akta Explorer FPLC system (GE Healthcare, Sweden) controlled by Unicorn software. The filtrate solution was loaded at 5 ml/min onto a 20-ml HisPrep FF 16/10 column containing nickel-Sepharose fast flow resin. The column was previously equilibrated with 10 column volumes (CV) of equilibration buffer. After loading, unbound and non-specifically bound proteins were removed by washing the column with 10 CV of the equilibration buffer at a flow rate of 5 ml/min. The specifically bound protein was retrieved from the column using a pH gradient starting at pH 7.5 in equilibration buffer and ending at pH 4.5 in elution buffer. The peak fractions were pooled together and diafiltered against diafiltration buffer containing progressively decreasing urea concentration (6, 4, 2 and 0 M) using centrifugal filtration devices with a 10-kDa molecular weight cut-off (Millipore, USA).
Offline measurement and protein analysis
Cell growth was determined by taking the optical density of the culture at 600 nm (OD600) with UV visible spectrophotometer (Thermo, USA). Cultures with higher cell densities were diluted suitably to have OD600 in the range of 0.2–0.4. To determine wet and dry cell weights, 15 ml of growth culture was centrifuged at 8,000 rpm for 30 min and the supernatant was discarded. The cell pellet was weighed on a balance to determine the wet cell weight. For dry cell weight, the cell pellet was dried in a hot air oven at 105 °C till constant weight. The protein estimation was carried out by BCA kit (Pierce, USA). The SDS-PAGE and Western blot analysis were carried out as the method described elsewhere (Tripathi et al. 2012b). Purity of rCHIKV E2 protein was determined by SDS-PAGE using 12 % polyacrylamide gel stained with Coomassie Brilliant Blue. Western blot analysis was performed using polyclonal mouse anti-CHIKV antibody. Proteins resolved by SDS-PAGE were electrophoretically transferred onto a PVDF membrane for Western blot analysis. The membrane was subjected to overnight blocking in PBS containing 5 % bovine serum albumin (BSA) and then washed with PBS containing 0.05 % Tween 20. The washed membrane was then reacted with mouse anti-CHIKV antibody with 1:1,000 dilution in PBS (Khan et al. 2012) for 1 h at 37 °C. The membrane was again washed thoroughly as above and incubated with horseradish peroxidase (HRP) conjugated goat anti-mouse IgG antibodies (Sigma, USA) at a dilution of 1:4,000 in PBS for 1 h at 37 °C. Finally, the blot was developed with phosphate–citrate buffer (0.1 M citric acid and 0.2 M Na2HPO4, pH 4.5) containing 3,3-diaminobenzidine tetrahydrochloride and H2O2 for 2 to 5 min. The developed membranes were washed extensively with distilled water, and the reaction pattern was recorded. The same procedure was also carried out for negative serum sample.
Indirect enzyme-linked immunosorbent assay (ELISA) for detection of anti-CHIKV IgM antibody
Indirect ELISA was performed to show the immunoreactivity of recombinant Chikungunya virus envelope 2 protein with anti-CHIKV IgM antibodies present in serum samples similar to the protocol described earlier (Yathi et al. 2011) with minor modifications. Ninety-six-well enzyme immunoassay plates were coated with purified recombinant CHIKV E2 protein (0.3 μg/well) in coating buffer (Na2CO3 0.1 M, NaHCO3 0.2 M; pH 9.6) at 37 °C and kept for 1 h. Plates were then washed with PBS containing 0.05 % Tween 20 (PBS-T). The plates were blocked using PBS with 5 % BSA for overnight at 4 °C. The blocked plates were then washed with PBS-T and incubated with diluted serum samples (100 μl/well) with 1:100 dilution in serum diluent (PBS + 1 % BSA) for 1 h at 37 °C. The plates were again washed with PBS-T to remove non-specific unbound antibodies. The specifically bound antibody was detected using anti-human IgM antibodies HRP-conjugate (Sigma, USA) diluted (1:2,000) in PBS containing 1 % BSA to the plates (100 μl/well), which were incubated at 37 °C for 1 h. The plates were again washed as above and added with 100 μl TMB (Sigma, USA) substrate solution per well. The plates were then incubated at 37 °C for 10 min, and the reaction was stopped with 100 μl of 2 M H2SO4. The plates were read with ELISA reader (BioTek, USA) at a 450-nm wavelength. Positive and negative controls were included each time when the test was performed. Each serum sample was tested in duplicate, and the mean OD for each sample was determined. To determine the cut-off value, the mean OD of the negative serum samples was calculated and taken as twice of the calculated value. Sensitivity and specificity of recombinant CHIKV E2 protein in detecting anti-CHIKV IgM antibodies in patient serum samples were determined based on the cut-off value. All the serum samples were also tested by SD IC test as per manufacturer's protocol, and the results were compared with in-house ELISA using rCHIKV E2 antigen.
Results
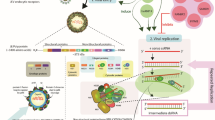
Chikungunya fever is a major public health concern of tropical countries. The epidemic spread rapidly and affected many geographical regions with an attack rate as high as 40–60 % (Yathi et al. 2011). There is neither any therapy nor any vaccine available for Chikungunya disease. Envelope proteins of Chikungunya virus are involved in binding to host cell receptors, and they also contain specific epitopes that elicit Chikungunya virus neutralizing antibodies. To study the diagnostic and vaccine potential of recombinant Chikungunya virus envelope 2 protein, large amount of pure and biologically active protein was necessary. A recombinant clone of E. coli BL21 (DE3) cells was developed using the plasmid (pET 28b + rCHIKV E2) construct to express this protein (Khan et al. 2012), and using this clone, we further developed batch and fed-batch fermentation processes to produce rCHIKV E2 protein in large quantities for use in diagnostic and vaccine studies. Figure 1 provides an overall flow diagram for the production and purification of rCHIKV E2 protein.

Procedure for preparation of recombinant CHIKV E2 protein
Recombinant Chikungunya virus envelope 2 (rCHIKV E2) protein
The envelope 2 gene (1,269 bp) of Chikungunya virus was used for the expression of rCHIKV E2 protein in E. coli. This rCHIKV E2 gene was inserted into the NcoI and XhoI sites, and the resultant NcoI–XhoI fragment was cloned downstream of the T7 promoter of E. coli expression vector pET28b to yield plasmid pET-CHIKV E2 as previously described (Khan et al. 2012). This pET28b + CHIKV E2 vector construct is predicted to encode a recombinant protein of ~47 kDa in size. The rCHIKV E2 protein was expressed as a fusion to a C-terminal 6× histidine tag to aid in detection and purification. Recombinant plasmid was transformed with E. coli BL21 (DE3) cells for protein expression. From SDS-PAGE analysis, the optimum concentration of IPTG and optimum duration of induction were 1 mM and 4 h, respectively, for expression of rCHIKV E2 protein. The level of protein expression observed after induction at log phase with 1 mM IPTG for 4 h was estimated to be 2.0 mg of rCHIKV E2 protein per liter culture (Table 1).
Improvement of rCHIKV E2 protein expression at shake flask culture
The improvement of the expression of recombinant Chikungunya virus envelope 2 protein in E. coli was tested at shake flask culture. In the shake flask culture, the final dry cell weight and protein yield were found to depend upon the media used. The highest amount of dry cell weight (about 2.51 g/l culture) was achieved in modified TB medium. Modified TB achieved about 8.75 mg of rCHIKV E2 protein per liter of culture medium, and TB achieved protein yield of 4.7 mg/l. Up to fourfold increase in the rCHIKV E2 protein expression was observed for modified TB medium in comparison to LB medium (Table 1). The rCHIKV E2 protein expression yields in LB, TB and modified TB media are shown in Table 1. This result indicates that the modified TB medium is better than the LB medium for enhanced expression of rCHIKV E2 protein by E. coli BL21 (DE3).
Batch and fed-batch fermentations
Large-scale production of rCHIKV E2 protein was carried out using batch and fed-batch cultivations. The conditions optimized at shake flask level were further scaled up to 5 l fermentor scale. For batch cultivations, cells were grown in TB and modified TB media and induced with IPTG during mid-exponential phase. During the cell growth period, the DO level was maintained more than 20 % during batch process using agitation speed (200–500 rpm) and air flow (4–7 l/min) as well as by supplying pure oxygen whenever necessary. At the time of the start of the batch process, DO level was more than 30 % and declined during the course of time as cells start growing. The dry cell weights at induction were 1.8 and 2.5 g/l, respectively, using TB and modified TB media for the batch process. The final dry cell weights were 4.5 and 6.2 g/l using TB and modified TB media after the batch fermentation process (Table 1).
High cell density fed-batch cultivations were also carried out for enhanced expression of rCHIKV E2 protein. As the pH of the bioreactor culture medium or DO level increases after the depletion of the carbon source present in batch medium (Manderson et al. 2006), we used both parameters to control the feed rate into the bioreactor. The culture at dry cell weight of 23 g/l (cell OD of 54) after elapse of 10 h of fermentation was induced with 1 mM IPTG. The final dry cell weight prior to cell lysis was 35 g/l, which was attained at ~14 h of fed-batch process. The successful production of rCHIKV E2 protein in the bioreactor using batch and fed-batch modes with a high yield was initiated to foster further applications and research with Chikungunya diagnostics and vaccines. Reproducibility of all shake flask cultures and batch fermentations as well as fed-batch fermentations were confirmed with additional experiments conducted under the above-specified optimal conditions, and the final yield was within less than 10 % of the result shown in Table 1. The fed-batch fermentation yielded approximately 0.55 kg cell paste that was frozen at −80 °C. The growth profile of the fed-batch fermentation process is shown in Fig. 2. Table 1 shows the comparative characteristics of the shake flask culture, batch and fed-batch fermentation processes.

Growth profile (OD600 vs time ) of E. coli cells during the fed-batch cultivation process. Cells were grown on batch mode till initial nutrients exhausted as indicated by DO concentration rise. Addition of feed medium started after 5 h of cultivations when DO start rising. At culture OD of about 54 after 10 h of cultivation, 1 mM IPTG was added to the bioreactor medium to induce the culture. Cells were harvested after 14 h of cultivation when the OD of the culture reaches about 84 with DCW of 35 g/l
Purification of rCHIKV E2 protein
The ultimate aim of expressing the E2 protein of the Chikungunya virus is the purification of the product using simple purification protocol by immobilized metal affinity chromatography. The affinity purified rCHIKV E2 protein was diafiltered and used in ELISA. The fed-batch process yielded 190 mg of purified protein per liter of culture, and the shake flask culture resulted to 8.75 mg of protein per liter culture using modified TB medium (Table 1). Improvements in product yield (21-fold higher) for the fed-batch process and threefold for the batch process were obtained as compared to the shake flask culture using modified TB medium (Table 1). The product yields were 4.70 and 22.60 mg/l after shake flask culture and batch cultivations, respectively, using TB medium.
Characterization of rCHIKV E2 protein
Characterization of affinity chromatography purified recombinant CHIKV E2 protein was subsequently performed. SDS-PAGE analyses demonstrated that the purified rCHIKV E2 protein has a molecular mass of approximately 47 kDa (Fig. 3a). The SDS-PAGE gel analysis showed that more than 90 % purity has been achieved using single-step affinity chromatography (Fig. 3b). The biomass recovered from 100 ml of fed-batch culture resulted to approximately 19 mg of purified rCHIKV E2 protein with 91 % purity and 45 % recovery. The estimation of crude cell lysate by densitometry analysis gave 42 mg of rCHIKV E2 protein and 380 mg total protein. The solubilized IBs were estimated to contain ~26 mg of the rCHIKV E2 protein with ~61 % recovery and ~72 % purity. The rCHIKV E2 protein was subjected to Western blot assay to confirm its antigenicity. The confirmation of antigenic authenticity of purified rCHIKV E2 protein was performed using Chikungunya virus-specific anti-sera. The immunoblot results revealed that the CHIKV anti-sera reacted with this protein (Fig. 3c) and the negative sample did not give any reaction (result not shown), thus suggesting the usefulness of this protein in diagnosis of Chikungunya infection.

a SDS–PAGE analysis of solubilized protein fraction derived from inclusion bodies. IBs were isolated from lysed cells and solubilized in solubilization buffer. Clarified solubilized supernatant was subjected to 12 % SDS-PAGE followed by Coomassie Brilliant Blue staining. Lane 1, molecular weight marker (kilodalton); lane 2, solubilized protein fraction. b Coomassie-stained SDS-PAGE of affinity chromatography purified rCHIKV E2 protein. The protein band of ~47 kDa confirmed the presence of the rCHIKV E2 protein. The protein profiles of the eluted protein in Coomassie-stained gel showed that more than 90 % purity has been achieved. Lane 1, MW marker (kilodalton); lane 2, purified rCHIKV E2 protein. c Western blot analysis of the purified rCHIKV E2 protein. Upon 12 % SDS-PAGE, the protein was electroblotted onto the PVDF membrane. The serum sample showing reaction with protein at desired size (~47 kDa) was considered positive. Lane 1, hyper-immune serum reacted with rCHIKV E2 protein
Evaluation of recombinant Chikungunya virus envelope 2 protein as an antigen for diagnosis of Chikungunya
The recombinant CHIKV envelope 2 protein was evaluated as a diagnostic reagent for the detection of anti-Chikungunya virus IgM antibodies using indirect microwell plate ELISA. Twenty-six CHIKV-positive serum samples collected earlier (Dash et al. 2007) were included for this study. Apart from positive samples, 14 serum samples suspected for CHIKV infections and ten normal sera from healthy volunteers were also included to check non-specific reactivity. All the samples were tested by SD Bioline Chikungunya IgM kit (Standard Diagnostics) before evaluating with in-house ELISA using rCHIKV E2 antigen. Among 50 sera samples, 26 samples were positive and 24 samples were negative by SD IgM assay. In-house ELISA using rCHIKV E2 antigen for each sample was done in duplicate, and the mean value was calculated. The mean absorbance (OD450) values of the CHIKV-positive and suspected negative serum samples were 0.910 and 0.330, respectively (Table 2). The mean OD value of ten normal control sera was 0.160. The cut-off value for in-house ELISA was determined based on twice the mean OD of the negative serum samples. Based on these OD values, the cut-off value was determined as 0.490. The OD values for positive and negative controls were 0.832 and 0.210, respectively. Among 50 serum samples evaluated with rCHIKV E2 protein as an antigen in in-house ELISA, 24 serum samples were positive and 26 serum samples were negative for anti-CHIKV IgM antibodies. The sensitivity of recombinant envelope 2 protein for anti-CHIKV-positive sera samples was 92 % (Table 3). The specificity of the recombinant CHIKV envelope 2 protein was 100 % (Table 3) when used with anti-CHIKV-negative serum samples which include both CHIKV-suspected and normal control serum samples. These results indicate that the recombinant CHIKV envelope 2 protein is reactive towards anti-CHIKV IgM antibodies, establishing its usefulness for detection of Chikungunya infections in patient serum samples.
Discussion
In this study, recombinant Chikungunya virus envelope 2 protein was produced in E. coli using batch and fed-batch cultivations. The rCHIKV E2 protein was purified and evaluated for its diagnostic potential. The envelope proteins (E1 or E2) of the Chikungunya virus are the major antigen used to elicit neutralizing antibody response and protective immunity in hosts, thus making efficient diagnostic and vaccine candidates against Chikungunya virus infections (Cho et al. 2008a; Cho et al. 2008b; Yathi et al. 2011; Kumar et al. 2012a; Khan et al. 2012; Kumar et al. 2012b; Metz et al. 2011; Metz et al. 2013). Expression of recombinant proteins in E. coli has been highly successful in recent past because this expression system offers high yield expression of foreign proteins (Khalilzadeh et al. 2008; Demain and Vaishnav 2009; Bhuvanesh et al. 2010; Huang et al. 2012; Nausch et al. 2013) and also allows simple and easy purification of recombinant proteins using single-step immobilized metal affinity chromatography (Bhuvanesh et al. 2010; Tan et al. 2010; Tripathi et al. 2010). Small-scale expression of recombinant proteins in E. coli is generally performed using simple growth medium such as LB broth using shake flask cultivations. However, for producing recombinant proteins at large scale using fermentor, trial with different growth media is necessary. The biomass and protein expression levels in cultures grown using TB and modified TB media were more in comparison with commonly used LB medium. Expression of recombinant proteins in E. coli is generally carried out by using either complex media or chemically defined media. Chemically defined growth media are known to produce lower growth and low protein expression levels in comparison with complex growth media which give higher expression levels and allow easier process control and simplify recovery of target proteins (Zanette et al. 1998; Lim et al. 2000; Manderson et al. 2006; Bhuvanesh et al. 2010; Volonte. 2011; Huang et al. 2012). Thus, TB and modified TB media were further used for the batch fermentation process. Large cell biomass and protein expression levels that resulted with TB and modified TB media can be explained, as these media are enriched with glycerol as carbon source, yeast extract and phosphate salts. The optimum concentration of IPTG, duration of induction and phase of growth for induction were also studied for this protein at shake flasks using LB medium. In the present study, the protein yield declined as the concentration of IPTG increased above 1 mM. This may be due to the presence of excessive IPTG that reduced the final cell density as a consequence of the growth inhibition. Maximum productivity was found to occur during induction at log phase when OD was around 0.8 for the cultures grown in LB medium. Induction at this OD for 4 h with 1 mM IPTG resulted in maximum protein yield for rCHIKV E2 protein with LB medium. This was in concurrence with previous work reported for induction (Zanette et al. 1998; Bhuvanesh et al. 2010). Cellular responses to induction depend on a number of interacting factors including the host/vector system and properties of the expressed protein. Therefore, the inducer concentration, duration of induction and phase of induction of new recombinants need to be empirically determined for each new clone (Bhuvanesh et al. 2010).
Recently, investigators have expressed the recombinant Chikungunya virus envelope 1, envelope 2 and capsid proteins in E. coli and insect cell (baculovirus) expression systems (Cho et al. 2008a, 2008b; Yathi et al. 2011; Kumar et al. 2012a; Khan et al. 2012; Yathi et al. 2013; Metz et al. 2013). Production of recombinant proteins for diagnostic, prophylactic or therapeutic use needs both increasing volumetric productivity and specific expression level of the target protein. Both volumetric product yield and specific product yield are strongly influenced by cultivation conditions (Bell et al. 2009). The other important factor for the development of any diagnostics, vaccines or therapeutics is the ability to produce the products with a process that is amenable to scale-up. Many recombinant proteins with diagnostic, prophylactic or therapeutic potential have been successfully produced using E. coli expression system employing batch and fed-batch fermentations, resulting in different cell biomass and recombinant protein production yields (Khalilzadeh et al. 2008; Fong and Wood 2010; Volonte et al. 2011; Huang et al., 20012; da Silva et al. 2013). Based on the previous works on rCHIKV E2 protein, we envisaged to produce this diagnostic and vaccine candidate using a bioreactor. In the present study, we describe the process development by batch and fed-batch cultivations, chromatographic purification and diagnostic potential of rCHIKV E2 protein. Batch fermentations were performed using TB and modified TB media. The modified TB medium again resulted in more cell biomass and product yields in comparison with TB medium (Table 1). This may be due to the presence of more glycerol in comparison with TB medium. Similar findings have already been established in case of more concentration of glycerol used with TB medium (Volonte et al. 2010). For further enhancement of rCHIKV E2 protein productivity, fed-batch fermentations were carried out using modified TB medium. The feed medium used during the fed-batch process mainly contained glycerol as carbon source, yeast extract, magnesium sulfate and trace metals. The positive attribute of complex media cultures is that they required the least amount of time for protein expression, which would decrease process utility expenses (Fong and Wood 2010). The yeast extract present in media enhances the specific cellular yield of the expressed protein, particularly during high cell density fermentation where the demand of nitrogenous source becomes very high following induction (Manderson et al. 2006; Huang et al. 2012). The composition of the growth media is crucial for enhancing product formation as well as reduction in byproduct formations. Earlier studies reported that the use of glycerol as carbon source in the medium did trigger the aerobic acetic fermentation, but the production of acetate was negligible (Korz et al. 1995; Pinsach et al. 2008; da Silva et al. 2013). Researchers also established that the fed-batch process with accumulation up to 30 g/l glycerol as well as with 0.35 per hour growth rate using complex medium resulted to faster biomass production, leading to shorter cultivations and without metabolite formation (da Silva et al. 2013). The final dry cell weight after the fed-batch process was found to be increased more than 36 times when compared with that of the shake flask culture with commonly used LB medium and about 19 times more with TB medium.
We developed a single-step affinity chromatography purification process for the rCHIKV E2 protein. Recombinant protein production in E. coli strain, IB formation is still considered as a convenient and effective way to recover target proteins (Singh and Panda 2005). The affinity chromatography purified protein was further diafiltered before used for ELISA. The SDS-PAGE analysis of purified protein resulted to the clear band of rCHIKV E2 protein with expected molecular weight of 47 kDa (Fig. 3b). The densitometry analysis of the SDS-PAGE gel revealed that more than 90 % purity was achieved. The results clearly demonstrate the successful production and purification of the recombinant Chikungunya virus envelope 2 protein. The final product concentration of recombinant CHIKV envelope 2 protein following affinity chromatography was significantly higher for the fed-batch fermentation process as compared to the batch fermentation and shake flask cultures (Table 1). About 190 mg of purified rCHIKV E2 protein was obtained from ~110 g wet cell biomass (35 g of dry cell biomass) of fed-batch culture. Improvement in rCHIKV E2 protein yield about more than 21 times was obtained for the fed-batch process as compared to shake flask culture using modified TB medium (Table 1). This is the first report on the scalable production of rCHIKV E2 proteins as well as use of bioreactor-produced protein for diagnosis of Chikungunya infection in patient serum samples. The purified rCHIKV E2 protein was analyzed by Western blot using hyper-immune sera, and the result (Fig. 3c) suggests that this protein could also be used for the purpose of diagnosis of Chikungunya virus infection.
Chikungunya virus envelope 2 protein remains as a potential marker for the detection of antigen as well as antibodies. Due to recent reemergence and spread of Chikungunya disease around the world, there is a demand for specific detection of Chikungunya virus by serodiagnosis. Several ELISA systems have been developed to detect antigens or IgM antibodies in human serum samples (Brehin et al. 2008; Cho et al. 2008a, 2008b; Shukla et al. 2009; Reddy et al. 2012; Kumar et al. 2012a; Khan et al. 2013). Most of them are based on the use of native viral antigen produced from cell culture. Therefore, there is a genuine need for a promising test system for detection of CHIKV infection. The antigenicity of the recombinant CHIKV E2 protein was evaluated by in-house indirect ELISA system for the detection of anti-CHIKV IgM antibodies. The in-house indirect IgM ELISA results indicate that the rCHIKV E2 antigen is sensitive and specific for detection of anti-CHIKV IgM antibodies in patient serum samples (Table 3). The in-house ELISA test has also shown more than 95 % agreement for detection of anti-CHIKV IgM antibodies as compared to commercial assay. Recombinant E2 protein-based ELISA is an easy, safe and affordable method for the serological diagnosis of Chikungunya infection. The rCHIKV E2 antigen-based ELISA has several distinct advantages as compared to native viral antigen (produced from inactivated Chikungunya virus) based ELISA systems. The preparation of native viral antigen from the infectious virus is associated with biohazard. The rCHIKV E2 protein obviates expensive and time-consuming cultivation of virus (for antigen preparation) and the associated biohazard risk. The importance of a rCHIKV E2 protein with antigenic potential expressed in E. coli focused to its large-scale application in the diagnosis of Chikungunya. The bacterial system is simple, easy to perform and cost-effective. The present study demonstrates that the in-house IgM ELISA test using rCHIKV E2 protein can be used as a promising test which will help authorities to undertake effective control measures and adopt management strategies against impeding Chikungunya menace. Since the envelope 2 protein of the Chikungunya virus contains neutralizing epitopes and host cell receptor recognition, it is also a useful antigen for development of subunit vaccines against Chikungunya viral infection (Kumar et al. 2012b; Metz et al. 2013). In our earlier study, we described cloning and expression of rCHIKV E2 protein as well as its immunogenicity for possible use as a vaccine candidate against Chikungunya virus (Khan et al. 2012). For use in serodiagnosis of Chikungunya infection as well as further studies in immunoprophylactic development, large quantity of purified and biologically active recombinant antigen protein was required, which could be produced using a bioreactor. The culture media optimization, batch and fed-batch fermentation strategies as mentioned in the present study established its application more appropriately. The results of in-house ELISA using purified rCHIKV E2 protein antigen with reference to commercial assay was in good correlation. The findings of the present study exhibits that the rCHIKV E2 protein can be an antigen of choice for cost-effective serodiagnosis of CHIKV infection.
In conclusion, we have developed batch and fed-batch fermentation processes for high yield production of rCHIKV E2 protein from E. coli. The cell growth and rCHIKV E2 protein yield was affected by composition of growth media and culture conditions. From economic point of view, it is essential to enhance volumetric and specific product yield by development of efficient fermentation and purification strategies. In the present study, batch and high cell density fed-batch cultivations were adopted to increase the product yield. The fed-batch fermentation strategy employed in this work is probably one of the cost-effective means to improve the cell biomass and recombinant protein production. The fed-batch fermentation process resulted to 190 mg of rCHIKV E2 protein per liter of culture. Further, characterization for its diagnostic potential to detect anti-CHIKV IgM antibodies established that its application is more attractive. The successful production of this protein, together with the one-step purification procedure, should facilitate its further investigation. Thus, the large amount of purified rCHIKV E2 protein obtained by these methods may possibly be used for Chikungunya diagnosis as well as for further vaccine development studies. Finally, this study provides us with some insight into the possible scale-up production strategies for other useful viral proteins.
References
Babaeipour V, Shojaosadati SA, Khalilzadeh R, Maghsoudi N, Farnoud AM (2010) Enhancement of human gamma-interferon production in recombinant E. coli using batch cultivation. Appl Biochem Biotechnol 160:2366–2376
Bell BA, Wood JF, Bansal R, Ragab H, Cargo J III, Washington MA, Wood CL, Ware LA, Ockenhouse CF, Yadava A (2009) Process development for the production of an E. coli produced clinical grade recombinant malaria vaccine for Plasmodium vivax. Vaccine 27:1448–1453
Bhuvanesh S, Chakkaravarthi A, Perumal K, Subramanian R (2010) Production and single-step purification of Brugia malayi abundant larval transcript (ALT-2) using hydrophobic interaction chromatography. J Ind Microbiol Biotechnol 37:1053–1059
Brehin AC, Rubrecht L, Navarro-Sanchez ME, Marechal V, Frenkiel MP, Lapalud P, Lane D, Despres P (2008) Production and characterization of mouse monoclonal antibodies reactive to Chikungunya envelope E2 glycoprotein. Virology 371:185–195
Cho B, Jeon BY, Kim J, Noh J, Kim J, Park M, Park S (2008a) Expression and evaluation of Chikungunya virus E1 and E2 envelope proteins for serodiagnosis of Chikungunya virus infection. Yonsei Med J 49:828–835
Cho B, Kim J, Cho JE, Jeon BY, Park S (2008b) Expression of the capsid protein of Chikungunya virus in a baculovirus for serodiagnosis of Chikungunya disease. J Virol Meth 154:154–159
da Silva AJ, Horta ACL, Velez AM, Iemma MRC, Sargo CR, Giordano RLC, Novo MTM, Giordano RC, Zangirolami TC (2013) Non-conventional induction strategies for production of subunit swine erysipelas vaccine antigen in E. coli fed-batch cultures. SpringerPlus 2:322
Dash PK, Parida MM, Santhosh SR, Verma SK, Tripathi NK, Ambuj S, Saxena P, Gupta N, Chaudhary M, Babu JP, Lakshmi V, Mamidi N, Rao PVL, Sekhar K (2007) East Central South African genotype as the causative agent in reemergence of Chikungunya outbreak in India. Vector Borne Zoonotic Dis 7:519–528
Demain AL, Vaishnav P (2009) Production of recombinant proteins by microbes and higher organisms. Biotechnol Adv 27:297–306
Fong BA, Wood DW (2010) Expression and purification of ELP-intein-tagged target proteins in high cell density E. coli fermentation. Microb Cell Fact 9:77
Huang CJ, Lin H, Yang X (2012) Industrial production of recombinant therapeutics in E. coli and its recent advancements. J Ind Microbiol Biotechnol 39:383–399
Khalilzadeh R, Mohammadian MJ, Bahrami A, Tabbar AN, Khalili MAN, Amouheidari A (2008) Process development for production of human granulocyte-colony stimulating factor by high cell density cultivation of recombinant Escherichia coli. J Ind Microbiol Biotechnol 35:1643–1650
Khan M, Dhanwani R, Rao PV, Parida M (2012) Subunit vaccine formulations based on recombinant envelope proteins of Chikungunya virus elicit balanced Th1/Th2 response and virus-neutralizing antibodies in mice. Virus Res 167:236–246
Khan M, Dhanwani R, Kumar JS, Rao PVL, Parida M (2013) Comparative evaluation of the diagnostic potential of recombinant envelope proteins and native cell culture purified viral antigens of Chikungunya virus. J Med Virol doi:10.1002/jmv.23752
Kielian M, Rey FA (2006) Virus membrane-fusion proteins: more than one way to make a hairpin. Nat Rev Microbiol 4:67–76
Korz DJ, Rinas U, Hellmuth K, Sanders EA, Deckwer WD (1995) Simple fed-batch technique for high cell density cultivation of E. coli. J Biotechnol 39:59–65
Kumar J, Khan M, Gupta G, Bhoopati M, Lakshmana Rao PV, Parida M (2012a) Production, characterization, and application of monoclonal antibodies specific to recombinant (E2) structural protein in antigen-capture ELISA for clinical diagnosis of Chikungunya virus. Viral Immunol 25:153–160
Kumar M, Sudeep AB, Arankalle VA (2012b) Evaluation of recombinant E2 protein-based and whole-virus inactivated candidate vaccines against Chikungunya virus. Vaccine 30:6142–6149
Lim HK, Jung KH, Park DH, Chung SI (2000) Production characteristics of interferon-α using an l-arabinose promoter system in a high-cell-density culture. Appl Microbiol Biotechnol 53:201–208
Manderson D, Dempster R, Chisti Y (2006) Recombinant vaccine against hydatidosis: production of the antigen in E. coli. J Ind Microbiol Biotechnol 33:173–182
Metz SW, Geertsema C, Martina BE, Andrade P, Heldens JG, van Oers MM, Goldbach RW, Vlak JM, Pijlman GP (2011) Functional processing and secretion of Chikungunya virus E1 and E2 glycoproteins in insect cells. Virol J 8:353
Metz SW, van den Doel P, Geertsema C, Osterhaus AD, Vlak JM, Martina BE, Pijlman GP (2013) Chikungunya virus-like particles are more immunogenic in a lethal AG129 mouse model compared to glycoprotein E1 or E2 subunits. Vaccine 31:6092–6096
Nausch H, Huckauf J, Koslowski R, Meyer U, Broer I, Mikschofsky H (2013) Recombinant production of human interleukin 6 in Escherichia coli. PLoS One 8:e54933
Pinsach J, de Mas C, Lopez-Santin J (2008) Induction strategies in fed-batch cultures for recombinant protein production in Escherichia coli: application to rhamnulose 1-phosphate aldolase. Biochemical Engg J 41:181–187
Reddy V, Ravi V, Desai A, Parida M, Powers AM, Johnson BW (2012) Utility of IgM ELISA, TaqMan real‐time PCR, reverse transcription PCR, and RT‐LAMP assay for the diagnosis of Chikungunya fever. J Med Virol 84:1771–1778
Shukla J, Khan M, Tiwari M, Sannarangaiah S, Sharma S, Rao PVL, Parida M (2009) Development and evaluation of antigen capture ELISA for early clinical diagnosis of Chikungunya. Diagn Microbiol Infect Dis 65:142–149
Singh SM, Panda AK (2005) Solubilization and refolding of bacterial inclusion body proteins. J Biosci Bioeng 99:303–310
Tan LCM, Chua AJS, Goh LSL, Pua SM, Cheong YK, Ng ML (2010) Rapid purification of recombinant dengue and West Nile virus envelope domain III proteins by metal affinity membrane chromatography. Protein Expr Purif 74:129–139
Tripathi NK, Shukla J, Biswal KC, Rao PVL (2010) Development of a simple fed-batch process for high yield production of Japanese encephalitis virus protein. Appl Microbiol Biotechnol 86:1795–1803
Tripathi NK, Shrivastava A, Biswal KC, Rao PV (2012a) Development of a pilot-scale production process and characterization of a recombinant Japanese encephalitis virus envelope domain III protein expressed in Escherichia coli. Appl Microbiol Biotechnol 95:1179–1189
Tripathi NK, Kumar JS, Biswal KC, Rao PV (2012b) Production of recombinant nonstructural 1 protein in Escherichia coli for early detection of Japanese encephalitis virus infection. Microbial Biotechnol 5:599–606
Volonte F, Pollegioni L, Molla G, Frattini L, Marinelli F, Piubelli L (2010) Production of recombinant cholesterol oxidase containing covalently bound FAD in Escherichia coli. BMC Biotechnol 10:33
Volonte F, Piubelli L, Pollegioni L (2011) Optimizing HIV-1 protease production in Escherichia coli as fusion protein. Microb Cell Fact 10:53
Yathi KK, Joseph JM, Bhasker S, Kumar R, Chinnamma M (2011) Recombinant CHIK virus E1 coat protein of 11 kDa with antigenic domains for the detection of Chikungunya. J Immunol Methods 372:171–176
Yathi KK, Bhasker S, Chinnamma M (2013) Determination of B cell epitopes and evaluation of antigen capture ELISA for the earlier diagnosis of CHIK virus using anti-rCHIK E1 rabbit antibodies. J Immunol Meth 393:45–52
Zanette D, Dundon W, Soffientini A, Sottani C, Marinelli F, Akeson A, Sarubbi E (1998) Human IL-1 receptor antagonist from Escherichia coli: large-scale microbial growth and protein purification. J Biotechnol 64:187–196
Acknowledgments
The authors are thankful to Prof. M. P. Kaushik, Director, DRDE, Gwalior for his keen interest, constant support and for providing necessary facilities for this study.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Tripathi, N.K., Priya, R. & Shrivastava, A. Production of recombinant Chikungunya virus envelope 2 protein in Escherichia coli . Appl Microbiol Biotechnol 98, 2461–2471 (2014). https://doi.org/10.1007/s00253-013-5426-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-013-5426-4