
Abstract
The TH9 subset of helper T cells was initially shown to contribute to the induction of autoimmune and allergic diseases, but subsequent evidence has suggested that these cells also exert antitumor activities. However, the molecular events that account for their effector properties are elusive. Here we found that the transcription factor IRF1 enhanced the effector function of TH9 cells and dictated their anticancer properties. Under TH9-skewing conditions, interleukin 1β (IL-1β) induced phosphorylation of the transcription factor STAT1 and subsequent expression of IRF1, which bound to the promoters of Il9 and Il21 and enhanced secretion of the cytokines IL-9 and IL-21 from TH9 cells. Furthermore, IL-1β-induced TH9 cells exerted potent anticancer functions in an IRF1- and IL-21-dependent manner. Our findings thus identify IRF1 as a target for controlling the function of TH9 cells.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout







Similar content being viewed by others

Accession codes
References
Mosmann, T.R., Cherwinski, H., Bond, M.W., Giedlin, M.A. & Coffman, R.L. Two types of murine helper T cell clone. I. Definition according to profiles of lymphokine activities and secreted proteins. J. Immunol. 136, 2348–2357 (1986).
Korn, T., Bettelli, E., Oukka, M. & Kuchroo, V.K. IL-17 and Th17 Cells. Annu. Rev. Immunol. 27, 485–517 (2009).
Crotty, S. Follicular helper CD4 T cells (TFH). Annu. Rev. Immunol. 29, 621–663 (2011).
Sakaguchi, S., Miyara, M., Costantino, C.M. & Hafler, D.A. FOXP3+ regulatory T cells in the human immune system. Nat. Rev. Immunol. 10, 490–500 (2010).
Dardalhon, V. et al. IL-4 inhibits TGF-β-induced Foxp3+ T cells and, together with TGF-β, generates IL-9+IL-10+Foxp3− effector T cells. Nat. Immunol. 9, 1347–1355 (2008).
Veldhoen, M. et al. Transforming growth factor-β 'reprograms' the differentiation of T helper 2 cells and promotes an interleukin 9-producing subset. Nat. Immunol. 9, 1341–1346 (2008).
Jäger, A., Dardalhon, V., Sobel, R.A., Bettelli, E. & Kuchroo, V.K. Th1, Th17, and Th9 effector cells induce experimental autoimmune encephalomyelitis with different pathological phenotypes. J. Immunol. 183, 7169–7177 (2009).
Staudt, V. et al. Interferon-regulatory factor 4 is essential for the developmental program of T helper 9 cells. Immunity 33, 192–202 (2010).
Chang, H.C. et al. The transcription factor PU.1 is required for the development of IL-9-producing T cells and allergic inflammation. Nat. Immunol. 11, 527–534 (2010).
Rengarajan, J. et al. Interferon regulatory factor 4 (IRF4) interacts with NFATc2 to modulate interleukin 4 gene expression. J. Exp. Med. 195, 1003–1012 (2002).
Klemsz, M.J., McKercher, S.R., Celada, A., Van Beveren, C. & Maki, R.A. The macrophage and B cell-specific transcription factor PU.1 is related to the ets oncogene. Cell 61, 113–124 (1990).
Purwar, R. et al. Robust tumor immunity to melanoma mediated by interleukin-9-producing T cells. Nat. Med. 18, 1248–1253 (2012).
Lu, Y. et al. Th9 cells promote antitumor immune responses in vivo . J. Clin. Invest. 122, 4160–4171 (2012).
Ghiringhelli, F. et al. Activation of the NLRP3 inflammasome in dendritic cells induces IL-1β-dependent adaptive immunity against tumors. Nat. Med. 15, 1170–1178 (2009).
Chung, Y. et al. Critical regulation of early Th17 cell differentiation by interleukin-1 signaling. Immunity 30, 576–587 (2009).
Horka, H. et al. The tick salivary protein sialostatin L inhibits the Th9-derived production of the asthma-promoting cytokine IL-9 and is effective in the prevention of experimental asthma. J. Immunol. 188, 2669–2676 (2012).
Taki, S. et al. Multistage regulation of Th1-type immune responses by the transcription factor IRF-1. Immunity 6, 673–679 (1997).
Lohoff, M. et al. Interferon regulatory factor-1 is required for a T helper 1 immune response in vivo. Immunity 6, 681–689 (1997).
Nizamutdinova, I.T. et al. Anthocyanins from black soybean seed coats preferentially inhibit TNF-alpha-mediated induction of VCAM-1 over ICAM-1 through the regulation of GATAs and IRF-1. J. Agric. Food Chem. 57, 7324–7330 (2009).
Kara, E.E. et al. Distinct chemokine receptor axes regulate Th9 cell trafficking to allergic and autoimmune inflammatory sites. J. Immunol. 191, 1110–1117 (2013).
Adachi, O. et al. Targeted disruption of the MyD88 gene results in loss of IL-1- and IL-18-mediated function. Immunity 9, 143–150 (1998).
Burns, K. et al. MyD88, an adapter protein involved in interleukin-1 signaling. J. Biol. Chem. 273, 12203–12209 (1998).
Shuai, K., Schindler, C., Prezioso, V.R. & Darnell, J.E. Jr . Activation of transcription by IFN-γ: tyrosine phosphorylation of a 91-kD DNA binding protein. Science 258, 1808–1812 (1992).
Varinou, L. et al. Phosphorylation of the Stat1 transactivation domain is required for full-fledged IFN-gamma-dependent innate immunity. Immunity 19, 793–802 (2003).
Darnell, J.E. Jr., Kerr, I.M. & Stark, G.R. Jak-STAT pathways and transcriptional activation in response to IFNs and other extracellular signaling proteins. Science 264, 1415–1421 (1994).
Liu, A. et al. TRAF6 protein couples Toll-like receptor 4 signaling to Src family kinase activation and opening of paracellular pathway in human lung microvascular endothelia. J. Biol. Chem. 287, 16132–16145 (2012).
Kato, M. et al. Transgenic mouse model for skin malignant melanoma. Oncogene 17, 1885–1888 (1998).
Lengagne, R. et al. Spontaneous vitiligo in an animal model for human melanoma: role of tumor-specific CD8+ T cells. Cancer Res. 64, 1496–1501 (2004).
Eyles, J. et al. Tumor cells disseminate early, but immunosurveillance limits metastatic outgrowth, in a mouse model of melanoma. J. Clin. Invest. 120, 2030–2039 (2010).
Frederiksen, K.S. et al. IL-21 induces in vivo immune activation of NK cells and CD8+ T cells in patients with metastatic melanoma and renal cell carcinoma. Cancer Immunol. Immunother. 57, 1439–1449 (2008).
Spolski, R. & Leonard, W.J. Interleukin-21: basic biology and implications for cancer and autoimmunity. Annu. Rev. Immunol. 26, 57–79 (2008).
Schmitt, E. et al. IL-9 production of naive CD4+ T cells depends on IL-2, is synergistically enhanced by a combination of TGF-β and IL-4, and is inhibited by IFN-γ. J. Immunol. 153, 3989–3996 (1994).
Miyamoto, M. et al. Regulated expression of a gene encoding a nuclear factor, IRF-1, that specifically binds to IFN-β gene regulatory elements. Cell 54, 903–913 (1988).
Taniguchi, T., Ogasawara, K., Takaoka, A. & Tanaka, N. IRF family of transcription factors as regulators of host defense. Annu. Rev. Immunol. 19, 623–655 (2001).
Honda, K. & Taniguchi, T. IRFs: master regulators of signalling by Toll-like receptors and cytosolic pattern-recognition receptors. Nat. Rev. Immunol. 6, 644–658 (2006).
Elser, B. et al. IFN-γ represses IL-4 expression via IRF-1 and IRF-2. Immunity 17, 703–712 (2002).
Kano, S. et al. The contribution of transcription factor IRF1 to the interferon-γ–interleukin 12 signaling axis and TH1 versus TH-17 differentiation of CD4+ T cells. Nat. Immunol. 9, 34–41 (2008).
Jäger, A. & Kuchroo, V.K. Effector and regulatory T-cell subsets in autoimmunity and tissue inflammation. Scand. J. Immunol. 72, 173–184 (2010).
Muranski, P. et al. Th17 cells are long lived and retain a stem cell-like molecular signature. Immunity 35, 972–985 (2011).
Okamoto, M. et al. Constitutively active inflammasome in human melanoma cells mediating autoinflammation via caspase-1 processing and secretion of interleukin-1beta. J. Biol. Chem. 285, 6477–6488 (2010).
Zeng, R. et al. Synergy of IL-21 and IL-15 in regulating CD8+ T cell expansion and function. J. Exp. Med. 201, 139–148 (2005).
Petrella, T.M. et al. Interleukin-21 has activity in patients with metastatic melanoma: a Phase II study. J. Clin. Oncol. 30, 3396–3401 (2012).
Dudley, M.E. et al. Cancer regression and autoimmunity in patients after clonal repopulation with antitumor lymphocytes. Science 298, 850–854 (2002).
Muranski, P. & Restifo, N.P. Adoptive immunotherapy of cancer using CD4+ T cells. Curr. Opin. Immunol. 21, 200–208 (2009).
Hunder, N.N. et al. Treatment of metastatic melanoma with autologous CD4+ T cells against NY-ESO-1. N. Engl. J. Med. 358, 2698–2703 (2008).
Steenwinckel, V. et al. IL-13 mediates in vivo IL-9 activities on lung epithelial cells but not on hematopoietic cells. J. Immunol. 178, 3244–3251 (2007).
Taylor, J., Schenck, I., Blankenberg, D. & Nekrutenko, A. in Current Protocols in Bioinformatics (ed. Minor, W.) Ch. 10, Unit 10 15 (Wiley Online Library, 2007).
Li, H. & Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 25, 1754–1760 (2009).
Trapnell, C., Pachter, L. & Salzberg, S.L. TopHat: discovering splice junctions with RNA-Seq. Bioinformatics 25, 1105–1111 (2009).
Trapnell, C. et al. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 28, 511–515 (2010).
Acknowledgements
We thank J.C. Renauld (Université catholique de Louvain) and V. Kuchroo (Brigham and Women's Hospital) for Il9r−/− mice and Il21r−/− mice; B. Ryffel (Université d'Orléans) for Ticam1−/−, Myd88−/−, Il1r1−/− and Ifng−/− mice; A. Prévost-Blondel (Cochin Institute) for MT-ret mice; J.D. Rosenblatt (University of Miami Miller School of Medicine) for B16-OVA cells; and R. Vile (Mayo Clinic) for LLC-OVA cells. Supported by the French National Research Agency (“Investissements d'Avenir” program; ANR-11-LABX-0021), the Ligue nationale contre le cancer (F.G. and F.V.), the Fondation de France (L.A.), the Institut National du Cancer (F.G.), the Association pour la recherche sur le cancer (L.A., F.G., G.M. and A.P.-B.), the Conseil Régional de Bourgogne/INSERM (H.B.), the Centre National de la Recherche Scientifique, Fonds Européen de Développement Économique et Régional, Le Studium, Orléans and Fondation pour la Recherche Médicale (F.G. and B.R.), the French National Research Agency (ANR-13-JSV3-0001 to L.A.), the Fondation pour l'aide à la Recherche sur la Sclérose en Plaques (L.A. and M.D.), the Ligue Régionale contre le cancer Comité Grand-Est (L.A.) and the European Commission (PCIG10-GA-2011-303719 to L.A.).
Author information
Authors and Affiliations
Contributions
F.V. and H.B. did in vitro and in vivo experiments; G.M. and A.P.-B. did in vivo experiments; R.B., M.B., M.D., F.C., C.R. and V.D. did in vitro experiments; B.R., M.K. and A.P.-B. provided materials and suggestions for the study; and F.G. and L.A. supervised the study and wrote the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Integrated supplementary information
Supplementary Figure 1 IL-1β enhances but does not skew TH9 cell polarization.
a) Cell-sorted naive CD4+CD62Lhi CD44lo T cells were isolated from mice and differentiated in the absence of any differentiating cytokines (TH0 cells) or with TGF-β and IL-4 (TH9 cells) without (CT: control) or with IL-1α, IL-1β, IL-18 or IL-33 in the presence of anti-CD3 and anti-CD28 for 72 hours and secretion of IL-9, IL-10 and IL-21 was assessed. b) Naive CD4 T cells were differentiated as in (a) into TH9 cells with or without IL-1β and IL-9 and IL-21 secretion was assessed by intracellular cytokine staining. c) Naive CD4 T cells were differentiated into TH0 or TH9 cells with or without IL-1β and Irf4 and Sfpi1 expression was assessed by quantitative RT-PCR after 6 hours. d) Naive CD4 T cells were differentiated into TH0, TH1, TH2, TH17 or TH9 cells with or without IL-1β and Tbx21, Gata3 and Rorc expression was assessed by quantitative RT-PCR after 24 hours. e) Same as in (d) and Il4 expression level as well as IFN-γ and IL-17 secretion were respectively determined using quantitative RT-PCR and ELISA. *P < 0.05; **P < 0.01; ***P < 0.001 (Kruskal-Wallis test in a, c-e). Data are from one experiment representative of at least two independent experiments with similar results (mean and s.e.m. of three triplicates in a, c-e).
Supplementary Figure 2 IL-1β does not globally affect TH9 cell differentiation.
Scatter plot providing an evaluation of the correlation of gene expression of TH9 cells in the presence or not of IL-1β. RNA sequencing analysis of naive CD4+ T cells differentiated into TH9 cells in the presence or not of IL-1β was performed after 1 hour (panel a) or 3 days (panel b). The 95% confidence intervals for each linear regression curve are represented. FPKM (Fragments Per Kilobase of exon per Million fragments mapped). Data are from one experiment representative of at least three independent experiments with similar results.
Supplementary Figure 3 Downregulation of Irf1 prevents the IL-1β-driven enhancement of TH9 cell cytokine secretion.
IL-9, IL-10 and IL-21 secretion levels were determined using ELISA after a three-day culture of wild-type naive CD4 T cells transduced with control or Irf1 siRNA and polarized into effector cells (upper panel) or polarized into effector cells in the presence of Cyanidine-3-glucoside (C3G) (lower panel). ***P < 0.001 (Kruskal-Wallis test). Data are from one experiment representative of at least three independent experiments with similar results.
Supplementary Figure 4 TH9 cells can be identified in vivo by chemokine receptor expression.
WT mice were injected i.v. with B16F10 melanoma cells. CD4 T cells were stained using anti-CCR3, anti-CXCR3, anti-CCR6 and anti-CCR4 antibodies and sorted from the tumor draining lymph nodes of tumor-bearing mice. Il9, Ifng and Il17 expression was interrogated in CD4+ CCR3+ CXCR3+ CCR6+ CCR4- cells (TH9) and compared to TH9 depleted total CD4+ cells (CD4) using RT-PCR. **P < 0.01 (Mann-Whitney test). Data are from one experiment representative of two independent experiments with similar results (mean and s.e.m. of three triplicates).
Supplementary Figure 5 IL-1β-driven IL-9 and IL-21 secretion in TH9 cells depends on Fyn but not NF-κB.
a) Naive CD4 T cells were differentiated into TH0 or TH9 cells in the presence or absence of IL-1β without or without 10 nM bortezomib (Veh: Vehicle; Bor: Bortezomib). IL-9 and IL-21 secretion was determined by ELISA after a 3-day culture b) Wild-type naive CD4 T cells transfected with control (CT) or Fyn siRNA were differentiated into TH0 or TH9 cells with or without IL-1b. IL-9, IL-10 and IL-21 secretion was assessed by ELISA after a 3-day culture. **P < 0.01; ***P < 0.001 (Mann-Whitney test). Data are from one experiment representative of two independent experiments with similar results (mean and s.e.m. of three triplicates).
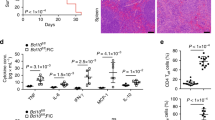
Supplementary Figure 6 Combined therapy using cyclophosphamide and the transfer of TH9 cells induced with IL-1β.
WT mice were injected s.c. with B16-OVA cells. At day 4, mice were treated with saline (CT) or cyclophosphamide (CTX). At days 5 and 10, mice were left untreated or received i.v. 2 million TH9 cells induced in the presence of IL-1β and tumor growth was monitored. Pooled data from two independent experiments with 5 mice per group are shown (error bars: mean and s.e.m.). *P < 0.05; ***P < 0.001 (Left panel Mann-Whitney test; Right panel Log rank test).
Supplementary Figure 7 IL-1β enhances the anticancer effect of classical TH9 cells in vivo.
Wild-type mice were injected iv with B16-OVA melanoma cells with or without effector OT-II TH9 cells. Twenty-four hours later, mice were treated either with IL-1b or PBS. Number of lung tumor foci was evaluated 13 days later. **P < 0.01; ***P < 0.001 (Kruskal-Wallis test). Pooled data from two independent experiments with five mice per group are shown (error bars: mean and s.e.m.).
Supplementary Figure 8 IL-21 dictates the anticancer effects of IL-1β-induced TH9 cells.
a) Wild-type (WT) or Il9r–/ – mice were injected i.v. with B16-OVA melanoma cells and effector OT-II TH9 cells differentiated with or without IL-1b and lung tumor foci were enumerated at day 13 b) Wild-type or Il21r–/– mice were injected i.v. with B16-OVA melanoma cells and effector OT-II TH9 cells differentiated with or without IL-1b and lung tumor foci were enumerated at day 13. c) Wild-type or KitW-sh mice were injected i.v. with B16-OVA melanoma cells and effector OT-II TH9 cells differentiated with or without IL-1b and lung tumor foci were enumerated at day 13 d) Wild-type mice were injected i.v. with B16-OVA melanoma cells and effector OT-II TH9 cells differentiated with or without IL-1β and subsequently treated with liposomal clodronate (LC) or control liposome (Veh) day -1, 0, 1, 3 and then every 3 days following CD4 T cell transfer for macrophage depletion. Lung tumor foci were enumerated at day 13 e) Wild-type mice were injected i.v. with B16-OVA melanoma cells and effector OT-II TH9 cells differentiated with or without IL-1β and subsequently treated with anti-CD19 or Ig control day -1, 0, 1, 3 and then every 3 days following CD4 T cell transfer for B cell depletion. Lung tumor foci were enumerated at day 13. **P < 0.01; ***P < 0.001 (Mann-Whitney test). Data are from one experiment representative of at least two independent experiments with similar results (5 mice per group; error bars: mean and s.e.m.).
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–8 and Tables 1–2 (PDF 1544 kb)
Rights and permissions
About this article

Cite this article
Végran, F., Berger, H., Boidot, R. et al. The transcription factor IRF1 dictates the IL-21-dependent anticancer functions of TH9 cells. Nat Immunol 15, 758–766 (2014). https://doi.org/10.1038/ni.2925
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ni.2925
This article is cited by
-
Epigenetic reprogramming of T cells: unlocking new avenues for cancer immunotherapy
Cancer and Metastasis Reviews (2024)
-
CD4+ T cells in cancer
Nature Cancer (2023)
-
IL-9 aggravates SARS-CoV-2 infection and exacerbates associated airway inflammation
Nature Communications (2023)
-
Opposing functions of circadian protein DBP and atypical E2F family E2F8 in anti-tumor Th9 cell differentiation
Nature Communications (2022)
-
Mouse pulmonary interstitial macrophages mediate the pro-tumorigenic effects of IL-9
Nature Communications (2022)
