
Abstract
Recent findings suggest that the mesolimbic dopamine neurons, known to promote cocaine-seeking behavior, are strongly inhibited by a newly characterized region of the midbrain known as the rostromedial tegmental nucleus (RMTg). The RMTg appears to be involved in generating reward-prediction error signals and inhibition of motivated behaviors, suggesting its potential involvement in the extinction of cocaine seeking as well. Therefore, to address this question, male Sprague–Dawley rats underwent surgeries for implantation of catheters and cannulas targeted at the RMTg. After cocaine self-administration, rats underwent modified extinction training. Pre- or post-training intra-RMTg microinjections of the allosteric AMPA receptor potentiator PEPA during the first 5 days of extinction training appeared to enhance the retention of the extinction learning. Following the extinction training, rats underwent cue-induced reinstatement or an ‘inactivation-alone’ extinction tests. RMTg inactivation before a cue-induced reinstatement session or inactivation alone before a standard extinction session increased overall lever pressing. To determine whether these effects generalized to other motivated behaviors, additional experiments examining food-seeking behavior were also conducted. The results from the food-seeking experiments indicate that PEPA microinjections into the RMTg did not influence the extinction of food seeking and that, at least in rats that had not been given PEPA during the extinction learning experiments, RMTg inactivation had no effect on lever pressing during the cue-induced reinstatement or inactivation-alone tests. These findings suggest that the RMTg provides general behavioral inhibition and is potentially involved in learning to extinguish cocaine-seeking behavior in rats.
Similar content being viewed by others

INTRODUCTION
Considerable research using the drug self-administration model has addressed the neural circuits that drive the reinstatement of cocaine-seeking behavior, yet less attention has been devoted to those systems that promote the extinction and ongoing suppression of cocaine-seeking behavior following extinction training. Evidence suggests that both the extinction of cocaine-seeking behavior as well as the ongoing suppression of cocaine seeking after extinction training require activity in the ventral part of the medial prefrontal cortex, known as the infralimbic (IL) cortex (LaLumiere et al, 2010; Peters et al, 2008). Following extinction training, IL inactivation induces cocaine-seeking behavior (Peters et al, 2008), whereas IL activation reduces cocaine-prime and cue-induced reinstatement (LaLumiere et al, 2012; Peters et al, 2008). IL inactivation during early extinction training also impairs the retention of extinction learning, whereas potentiating IL activity in the same time period enhances this retention (LaLumiere et al, 2010). Findings indicate that the suppression of cocaine-seeking behavior by IL activation is reversed by activation of the ventral tegmental area (VTA) dopamine neurons and activation of dopamine receptors in the nucleus accumbens (NA) shell, prelimbic cortex, and basolateral amygdala (LaLumiere et al, 2012). Thus, the suppression of cocaine-seeking behavior may depend, in part, on inhibition of the VTA-based dopamine system, although it is not clear how such inhibition may occur.
Recent work has identified a structure in the midbrain, known as the rostromedial tegmental nucleus (RMTg) or tail of the VTA, that has been suggested as a ‘master brake’ on the dopamine system (Barrot et al, 2012; Bourdy and Barrot, 2012). The RMTg lies immediately caudal to the posterior VTA and sends dense GABAergic projections to the VTA (Jhou et al, 2009b; Kaufling et al, 2009, 2010; Perrotti et al, 2005). Evidence suggests that a variety of aversive stimuli increase firing of RMTg neurons, including the omission of expected rewards (Barrot et al, 2012). Following training with fixed cues for either sucrose (Jhou et al, 2009a) or apple juice (Hong et al, 2011), a significant number of RMTg neurons increase firing rates in response to omission of expected reward delivery when compared with neurons outside of the RMTg. In contrast, rewards and cues predictive of rewards inhibit RMTg activity. These findings suggest that changes in RMTg activity may be a critical driver of VTA dopamine neuron activity as part of the reward-prediction error signal that VTA dopamine neurons have been hypothesized to produce (Barrot et al, 2012; Schultz, 2013). This signal is believed to be critical in learning the differences between expected and observed rewards (Steinberg et al, 2013). Indeed, neuronal responses in the VTA and RMTg appear to be opposite from each other depending on the reward expectations of the animal. Further evidence for this potential role for the RMTg comes from studies examining the lateral habenula. The lateral habenula provides a major glutamatergic input to the RMTg and appears to code for negative reward signals, for example, the omission of an expected reward (Jhou et al, 2009a, 2009b; Matsumoto and Hikosaka, 2007). Importantly, recent findings also indicate that the aversive properties of cocaine are mediated via the lateral habenula–RMTg pathway (Jhou et al, 2013). Thus, evidence suggests that lateral habenula inputs to the RMTg and RMTg inputs to the VTA are critical components of a reward-prediction error pathway (Barrot et al, 2012).
As extinction of cocaine-seeking behavior involves learning about the omission of an expected reward and, after training, the ongoing suppression of motivated behavior, the RMTg may be involved in these processes. To investigate this hypothesis, rats underwent cocaine self-administration followed by extinction training, using a modified extinction protocol that we have previously developed (LaLumiere et al, 2010), to determine whether altering RMTg activity modulates the extinction learning. Following the extinction training, the RMTg of rats was then inactivated before a cue-induced reinstatement test or before a standard extinction session to determine whether such inactivation could potentiate or even drive cocaine-seeking behavior. To determine whether these effects generalized to natural reward seeking, similar experiments were conducted in a food-seeking paradigm.
MATERIALS AND METHODS
Subjects
Male Sprague–Dawley rats (250–275 g at time of first surgery; Charles River, Wilmington, MA; n=75) were used for this study. All rats were single housed in a temperature-controlled environment under a 12-h reverse light/dark cycle (lights on at 1900 hours) and allowed to acclimate to the vivarium at least 3 days before surgery. Water was available ad libitum throughout all training and testing. All procedures used were in compliance with NIH guidelines for care of laboratory animals and approved by the University of Iowa Institutional Animal Care and Use Committee.
Surgery
For both cocaine- and food-seeking experiments, rats were placed in a stereotaxic instrument (Kopf Instruments, Tujunga, CA). Small surgical screws were implanted in the skull to serve as anchors, the guide cannula was implanted, and dental acrylic was used to secure the entire unit to the skull. A double-barreled cannula (1.2 mm width, 26 gauge, Plastics One, Wallingford, CT, USA) was aimed 3 mm dorsal to the RMTg (coordinates: A/P: −7.4 mm from Bregma and D/V: −5.5 mm from skull surface) based on previous work (Jhou et al, 2009b; Kaufling et al, 2009). Obdurators were used to maintain cannula patency and removed only during microinjections. Those rats that underwent cocaine self-administration also received intra-jugular venous catheters, as described previously (LaLumiere et al, 2012). Catheters were flushed with 0.1 ml 100 USP heparin and 0.1 ml of a glycerol lock solution (100 USP heparin, 70% sterile glycerol) daily throughout self-administration to maintain catheter patency. All animals were given 1 week to recover before behavioral procedures began.
Microinjection Procedures
Double barrel microinjectors (33 gauge) were connected to PE20 tubing that was connected to 10 μl Hamilton syringes. Syringes were controlled by an infusion pump. Microinjectors extended 3 mm beyond the end of the cannula and into the RMTg. All microinjections occurred over 60 s (0.3 μl/side) and injectors were left in place an additional 120 s to allow for diffusion. The following drugs were used throughout the experiments: the AMPA receptor potentiator PEPA (3 ng/side) or the GABAB and GABAA receptor agonists baclofen and muscimol (1 and 0.1 mM, respectively; BM), respectively. BM was dissolved in artificial cerebral spinal fluid (aCSF), whereas PEPA was initially dissolved in 50% aCSF/50% DMSO, which was then diluted to 90% aCSF/10% DMSO before use. PEPA was selected because it acts as an allosteric AMPA receptor potentiator and therefore does not directly activate the receptor but, rather, requires endogenous glutamate activation of the receptor (Nakagawa et al, 1999). Doses of all drugs were based on previous work (LaLumiere et al, 2012; Peters et al, 2008).
Behavioral Training and Testing
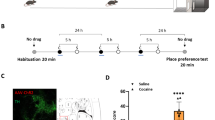
All behavioral training and testing occurred in a standard operant chamber (Med Associates, Fairfield, VT) containing two retractable levers and a house light, as well as a cue light and tone generator (4500 Hz) above the active lever. Figure 1a and b provides outlines of the order of the training and testing, described in detail below.

(a, b) Timelines illustrating order of behavioral training and testing in the cocaine and food self-administration experiments, respectively. As indicated in the Materials and Methods section, not all animals completed all steps. (c) Active and inactive lever presses and infusions during cocaine self-administration. (d) Active and inactive lever presses and number of food pellets during food self-administration. (e) Diagram showing microinjection tracks terminating in the RMTg for both the cocaine- and food-seeking groups. Figures adapted from Paxinos and Watson (2007) and A/P coordinates (in mm) are given relative to Bregma. Owing to the lack of clear structural boundaries for the RMTg, inclusion of data from animals was based on a conservative analysis and thus only needle tracks confirmed to be within the RMTg were included. In all cases, there was little evidence of tissue damage at the injection sites.
Cocaine Self-Administration
Before cocaine self-administration training began, rats underwent 24 h of food deprivation followed by a 15-h, overnight food-training session, as done previously (LaLumiere et al, 2012; Peters et al, 2008). Following food training, rats were maintained on a limited quantity of food (∼20 g) per day, given immediately after every self-administration, extinction, or reinstatement session.
Cocaine self-administration procedures were identical to those described previously (LaLumiere et al, 2012). In brief, active lever presses delivered a single 200 μg infusion of cocaine (dissolved in 0.9% saline; cocaine–HCl kindly provided by NIDA) on an FR1 schedule with a 20-s timeout period following each infusion. Each infusion was also paired with the presentation of the cue light and tone for 5 s. After a minimum of 12 days of self-administration with at least 10 days of >10 infusions, including the last 3 days, and discrimination between the active and inactive levers, animals moved into extinction.
Cocaine-Seeking Experiments
Extinction experiments
To examine the role of the RMTg in extinction learning and consolidation, we used procedures that we have used previously (LaLumiere et al, 2010). Throughout the extinction sessions, active lever presses produced no drug infusion or light/tone cues. During the first 5 days of extinction, rats received intra-RMTg microinjections of either PEPA or vehicle either immediately before or after the shortened (15 min) extinction session (modified non-cue extinction). Group assignments were matched based on infusions during the last 3 days of self-administration. For the next 7 days, animals were placed in the operant chamber for 2 h extinction sessions (standard non-cue extinction) without microinjections to assess the retention of extinction learning from the previous sessions.
Reinstatement experiments
Rats from the extinction experiments were then used in one of the following reinstatement experiments: cue-induced reinstatement test or inactivation-alone test. Reinstatements utilized a within-subjects design so that each animal received BM and aCSF on separate tests. Whether the rats received BM or aCSF first was counterbalanced based on their prior treatment group (PEPA vs aCSF). Active lever pressing was re-extinguished for a minimum of 3 days between reinstatement tests, and rats were required to meet the extinction criteria described below before undergoing the next test. Intra-RMTg microinjections of either BM or vehicle were given immediately before the session. Sessions were 2 h in length.
Because it was hypothesized that RMTg activity was at least partially responsible for suppressing cocaine-seeking behavior, we wanted to determine whether RMTg inactivation could potentiate cue-induced reinstatement. Therefore, a subset of rats (n=12) from the extinction experiments underwent five consecutive days of 2-h cue extinction sessions that served to reduce cue-driven lever pressing in order to reveal a potentiation during later reinstatement tests. This standard cue extinction utilized a program that was identical to the self-administration session program (ie, cue tone and light turned on with an active lever press), although rats did not receive any cocaine infusions. Following the standard cue extinction procedures, rats then underwent a minimum of 3 days of standard non-cue extinction sessions (at least two consecutive days with 25 or fewer active lever presses required before undergoing reinstatement tests), which served as the extinction baseline. The rats then underwent cue-induced reinstatement tests again by using the program that was identical to the self-administration training but without cocaine infusions. During the cue-induced reinstatement test, active lever presses produced the cue light and tone on an FR1 schedule with a 20-s timeout. Reinstatement tests were also 2 h in length and microinjections of BM or vehicle were given immediately before the sessions.
Based on the results from the cue-induced reinstatement tests, we wanted to determine whether RMTg inactivation alone could increase active lever pressing. This inactivation-alone test was examined in a separate subset of cocaine-seeking animals (n=7) that had also undergone the extinction experiments and was part of a second cohort of rats following the first cohort that underwent the cue-induced reinstatement tests. The inactivation-alone test used a standard extinction program (2 h, no light/tone cues or cocaine infusion). Microinjections of BM or vehicle were given immediately before the sessions.
Locomotor Activity
A subset of animals (n=4; three of the four underwent the extinction experiments but no reinstatement whereas the fourth completed both extinction and cue-induced reinstatement testing) was used in an open-field test to evaluate the effects of BM administration into the RMTg on locomotor activity, using a within-subjects design. The chamber consisted of 17″ × 17″ × 12″ height arena with 16 evenly spaced infrared transmitters and receivers around the edges (Med Associates). Animals were allowed a 30-min acclimation session to the testing chamber before receiving vehicle or BM microinjections and immediately being placed back in the chamber for a 2-h locomotor activity test. Animals were given 1 day off between open-field testing sessions, and microinjections were counterbalanced.
Food-Seeking Experiments
Following surgery and recovery, rats were given 20 g rat chow per day following each self-administration, extinction, and reinstatement session. The food-seeking experiments followed previous protocols established by McFarland et al (2004) in order to ensure robust reinstatement responding later during testing. Initially, active lever presses produced a single food pellet (45 mg; BioServ) on an FR1 schedule along with the same light/tone cues used in the cocaine self-administration studies. As training progressed, the reinforcement schedule, including both the food pellet and the cues, increased to FR3 and then FR5. Rats were required to receive 100 pellets per day for at least 3 days before moving onto the next schedule. Extinction procedures began after stable responding was achieved on the FR5 schedule.
Extinction experiments
The procedures for the extinction experiments for food seeking were identical to those of the cocaine-seeking studies, except that only the effects of pre-training intra-RMTg microinjections of PEPA or vehicle were assessed.
Reinstatement experiments
For the reinstatement tests with food-seeking behavior, the procedures were identical to those used with the cocaine-seeking studies. However, because the amount of active lever pressing in the food-seeking studies was considerably higher than those in the cocaine-seeking studies, the extinction criterion of 25 or fewer active lever presses for two consecutive days before undergoing reinstatement was found to be too strict. Therefore, the extinction criterion was that rats’ active lever presses was required to be <10% of the active lever presses from the last 2 days of food self-administration for two consecutive extinction sessions before undergoing a reinstatement test. In contrast to the cocaine-seeking studies, all rats that underwent the extinction experiments also underwent 5 days of 2-h standard cue extinction sessions, the cue-induced reinstatement tests, and the inactivation-alone tests. As with the cocaine experiments, animals had to meet extinction criteria before moving onto the next stage of the experiments, and multiple reinstatement or inactivation-alone tests were conducted in a counterbalanced, within-subjects design.
Histology
Rats were given an overdose of sodium pentobarbital (100 mg/kg, i.p.) and transcardially perfused with 0.9% saline. Brains were removed and stored in 3.7% formaldehyde. Coronal sections (75 μm) were taken, mounted onto gelatin-coated slides, and stained using Cresyl violet. Injection sites were verified under a light microscope according to the atlas of Paxinos and Watson (2007).
Data Analysis
Extinction data (days 1–5 and 6–12 analyzed separately) were analyzed using two-way repeated-measures analyses of variance (ANOVA) with post hoc t-tests between groups at each time point with Bonferroni corrections to the p-value, using GraphPad Prism software. Multiplicity-adjusted p-values (ie, the p-value for each test was multiplied by the number of post hoc tests) of <0.05 were considered significant. Reinstatement session lever presses were analyzed using a one-way ANOVA and a Tukey post hoc test was used when appropriate. Locomotor activity was analyzed using a Mann–Whitney test. p-Values of <0.05 were considered significant. All measures are expressed as mean±SEM with each group’s n indicated in the figure.
RESULTS
A total of 50 rats were included in the final data while another 25 were excluded for the following reasons: failure to acquire self-administration (one rat), incorrect cannula placements (15 rats, cocaine; 5 rats, food), lost head caps (three rats), and clogged cannula (one rat). Figure 1c and d shows the infusions or food and active and inactive lever presses during cocaine and food self-administration, respectively, from any animals included in the final data.
Histology
Figure 1e shows the location of the microinjector tips in the RMTg for all animals whose data were used in the final analysis (both cocaine- and food-seeking experiments).
Cocaine-Seeking Experiments
Extinction experiments
Figure 2a shows the active lever presses across the 12 days of the extinction experiment with the post-training microinjections. For the modified (15 min) non-cue extinction sessions on days 1–5, a two-way repeated-measures ANOVA found no significant effect of group (F(1,24)=1.077, p>0.05), a trend toward a significant effect of time (F(4,24)=2.482, p<0.08), and no significant interaction (F(4,24)=1.467, p>0.05). For the standard (2 h) non-cue extinction sessions on days 6–12, a two-way repeated-measures ANOVA revealed no significant effect of group (F(1,36)=1.903, p>0.05), a trend toward a significant effect of time (F(6,36)=2.164, p<0.07), and a significant interaction between group and time (F(6,36)=2.55, p<0.05). Those rats that had received PEPA immediately following training on days 1–5 showed a more rapid reduction in active lever pressing on days 6–12 compared with those rats that had received aCSF. Post hoc analysis of days 6–12 did not reveal any significant differences between the groups on any individual day. There were no significant effects of PEPA on inactive lever pressing (data not shown).

Effect of RMTg activation on extinction following cocaine self-administration. (a) Active lever presses (mean±SEM) during extinction sessions for those rats receiving post-training intra-RMTg microinjections of PEPA or aCSF. Modified non-cue extinction sessions on days 1–5 were 15 min in length with microinjections given immediately following each session, whereas standard non-cue extinction sessions on days 6–12 were 2 h in length with no microinjections. Those rats that had received PEPA immediately after each session during the modified non-cue extinction sessions showed a more rapid reduction in their active lever pressing on days 6–12. Two-way, repeated-measures ANOVAs of inactive lever pressing found no significant effects of time, group, or interaction for days 1–5 or 6–12 (p>0.05 in all cases; data not shown). (b) Active lever presses (mean±SEM) during extinction sessions for those rats receiving pre-training intra-RMTg microinjections of PEPA or aCSF. Those rats that had received PEPA immediately before each modified non-cue extinction session showed a faster reduction in their active lever pressing on days 6–12 (standard non-cue extinction sessions). Post hoc analysis of the results from days 6 to 12 indicated a significant difference between groups on day 7, the second day of the standard non-cue extinction sessions (p<0.05). Separate two-way repeated-measures ANOVAs of inactive lever pressing found no effect of time, group, or interaction on days 1–5 (p>0.05 in all cases; data not shown) and a significant effect of time (F(6,132)=2.579, p<0.05) and no significant effect of group or interaction on days 6–12 (p>0.05; data not shown). (c) Combined active lever presses (mean±SEM) during extinction sessions for those rats receiving pre- and post-training microinjections. Rats that had previously received PEPA during the modified non-cue extinction sessions (days 1–5), regardless of whether given before or after training, had a more rapid reduction in active lever pressing during the standard non-cue extinction sessions on days 6–12. Two-way repeated-measures ANOVAs were also conducted on inactive lever pressing for days 6–12 of extinction on the combined groups (p>0.05 in all cases; data not shown). *p<0.05, indicating a significant interaction between group and time for days 6–12 in the panel.
Figure 2b shows the active lever presses across the 12 days of the experiment for the pre-training microinjections. For the modified (15 min) non-cue extinction sessions on days 1–5, a two-way repeated-measures ANOVA found no significant effect of group (F(1,88)=0.023, p>0.05), a significant effect of time (F(4,88)=3.479, p<0.05), and no significant interaction (F(4,88)=0.348, p>0.05). For standard (2 h) non-cue extinction sessions on days 6–12, a two-way repeated-measures ANOVA revealed no significant effect of group (F(1,132)=1.112, p>0.05), a significant effect of time (F(6,132)=11.13, p<0.0001), and a significant interaction between group and time (F(6,132)=2.988, p<0.01). Those rats that had received PEPA immediately before training on days 1–5 showed a more rapid reduction in active lever pressing on days 6–12 compared with those rats that had received aCSF. Post hoc analysis of the results from days 6–12 indicated a significant difference between groups on day 7, the second day of 2 h extinction sessions (p<0.05). There were no significant effects of PEPA treatment on inactive lever presses (data not shown).
Although the post-training control group and pre-training control groups do not appear to show identical extinction curves on days 6–12, this is likely owing to the considerable variability observed across days. Indeed, our previous work has found large day-to-day variability on days 6–12 using identical behavioral paradigms (LaLumiere et al, 2010). Therefore, because there was such variability and because there was no difference in active lever pressing for the pre- vs post-training microinjections on the critical first 5 days of extinction, we also combined the data for additional analysis of days 6–12 (Figure 2c). For days 6–12 (standard non-cue extinction sessions) of the combined data, a two-way repeated-measures ANOVA revealed no effect of group (F(1,180)=2.747, p>0.05), a significant effect of time (F(6,180)=12.37, p<0.0001), and a significant interaction (F(6,180)=2.926, p<0.01). Those rats that had received PEPA during days 1–5 of extinction training had a more rapid reduction in active lever pressing during the retention tests on days 6–12. Moreover, post hoc analysis revealed a significant difference between groups on day 7 (p<0.05). There were no significant effects of PEPA treatment on inactive lever presses (data not shown).
In order to undergo the standard cue extinction training and reinstatement tests, rats had to complete a minimum of 7 days of standard (2 h) non-cue extinction and to achieve two consecutive days of fewer than 25 active lever presses. (Although not all rats from the extinction experiments actually continued into the subsequent reinstatement tests, they all underwent extinction training until criteria were reached). An additional analysis using a Mann–Whitney test (owing to the non-parametric distribution of data), therefore, was also conducted to determine whether there was a difference in the number of days of extinction the rats underwent before reaching the criteria. Those animals that received PEPA had a trend to more days of extinction compared with aCSF controls (p<0.1), but there were two outliers (one in each group) that skewed the results. When they were excluded, there was a significant difference, as the PEPA-treated rats underwent fewer days of extinction (mean days±SEM=7.125±0.125) compared with aCSF-treated rats (mean days±SEM=8.643±0.887; p<0.05).
Reinstatement experiments
As described in the Materials and Methods section, rats underwent cue extinction procedures before reinstatement tests. There was no difference in active lever pressing during the cue extinction sessions based on prior treatment (Supplementary Figure S1; statistics in the legend). Following re-extinction, rats then underwent cue-induced reinstatement tests. Figure 3a and b shows the active and inactive lever presses, respectively, during the cue-induced reinstatement tests of those rats that had received intra-RMTg microinjections of BM or aCSF. A one-way ANOVA of the active lever presses in panel A revealed a significant effect (F(2,33)=6.853, p<0.01). A one-way ANOVA of the inactive lever presses in panel B also revealed a significant effect (F(2,33)=8.463, p<0.01). In both cases, those rats that had received BM microinjections had significantly more lever presses during the cue-induced reinstatement test compared with the extinction baseline and those rats that had received aCSF (p<0.01 in all cases). Those rats receiving aCSF did not show a significant increase in active lever presses above baseline, indicating the effectiveness of the prior cued extinction. In addition, analyses (described in the legend in Figure 3) indicated that prior exposure to PEPA during extinction training had no effect on later reinstatement tests (data not shown).

Increased lever-pressing behavior and locomotor activity in rats whose RMTg was inactivated. (a, b) Active and inactive lever presses, respectively, (mean±SEM) during the cue-induced reinstatement session. Post hoc tests revealed that those rats that had received BM microinjections had significantly more active and inactive lever presses during the cue-induced reinstatement test compared with the extinction baseline and those rats that had received aCSF (p<0.01 in all cases). The aCSF group was not significantly different from the extinction baseline, indicating the effectiveness of the prior cue extinction. In addition, a two-way repeated-measures ANOVA was completed based on previous treatment during extinction training (aCSF vs PEPA) and reinstatement treatment (aCSF vs BM). There was no significant effect of prior treatment group (F(1,10)=1.158, p>0.05), a significant effect of drug (BM vs aCSF) given during reinstatement (F(1,10)=10.56, p<0.01), and no significant interaction between previous treatment group and drug treatment during reinstatement (F(1,10)=1.458, p>0.05; data not shown). Thus, prior treatment with PEPA did not affect lever-pressing behavior during cue-induced reinstatement testing. (c, d) Active and inactive lever presses, respectively, (mean±SEM) during a standard extinction session before which rats received intra-RMTg microinjections of either BM or aCSF. Post hoc tests in both cases revealed that those rats that had received BM microinjections had significantly more lever presses compared with the extinction baseline and those rats that had received aCSF (p<0.01 in all cases). In addition, a two-way repeated-measures ANOVA was completed based on previous treatment during extinction training (aCSF vs PEPA) and reinstatement treatment (aCSF vs BM). There was no significant effect of prior treatment group (F(1,5)=0.703, p>0.05), a significant effect of drug (BM vs aCSF) given during reinstatement (F(1,5)=17.48, p<0.01), and no significant interaction between previous treatment group and drug treatment during reinstatement (F(1,5)=0.588, p>0.05; data not shown). Thus, prior treatment with PEPA did not affect lever-pressing behavior during the inactivation-alone testing. (e) Total distance traveled (mean±SEM) during a locomotor activity test. Rats received intra-RMTg microinjections of BM or aCSF immediately before being placed in the locomotor activity test chamber for 2 h. A Mann–Whitney test revealed a significant difference between the two groups (p<0.05). *p<0.01 compared with extinction baseline and the aCSF group. #p<0.05 compared with the aCSF group.
As described in the Materials and Methods section, a separate subset of rats was given microinjections of BM or aCSF immediately before a full-length (2 h) standard non-cue extinction session. Figure 3c and d shows the active and inactive lever presses, respectively, from the BM-alone inactivation test. A one-way ANOVA of the active lever presses in panel C revealed a significant effect (F(2,18)=16.8, p<0.0001). A one-way ANOVA of the inactive lever presses in panel D revealed a significant effect (F(2,18)=14.91, p<0.001). Statistical analysis in both cases revealed that those rats that had received BM microinjections had significantly more lever presses compared with the extinction baseline and those rats that had received aCSF (p<0.01 in all cases).
Locomotor Activity
Because RMTg inactivation increased both active and inactive lever pressing in both of the above experiments, we considered the possibility that RMTg inactivation increases locomotor activity. Therefore, after completing the cocaine-seeking experiments, a subset of rats underwent a locomotor activity test, in which they received microinjections immediately before a 2-h open-field test. Rats’ assignments to their experimental groups were counterbalanced so that each animal received BM or vehicle on separate tests, using a within-subjects design. A Mann–Whitney test revealed that RMTg inactivation significantly increased total distance traveled during the test (p<0.05; Figure 3e).
Misplaced Injection Sites
To determine the degree of site specificity for the microinjections, a separate analysis of the cocaine-seeking data from animals with misplaced cannulas was conducted. No effect of PEPA microinjections on extinction learning or retention was observed for misplaced cannulations (Supplementary Figure S2). In contrast, regardless of the site of the misplacement, BM microinjections appeared to increase lever pressing during cue-induced reinstatement tests (Supplementary Figure S3). Supplementary Figure S4 shows the estimated locations of the microinjector tips outside the RMTg based on the site of misplaced microinjections as described in Supplementary Figure S3. An additional analysis (Supplementary Figure S5A and B) was conducted in which the distance between the RMTg and the site of the misplaced microinjection was estimated and data were based on whether the incorrect site was ‘proximal’ (⩽0.5 mm from RMTg) or ‘distal’ (>0.5 mm from RMTg). Both proximal and distal BM microinjections appeared to increase active lever pressing, although the distal microinjections appeared to have more variability in the behavioral response. Supplementary Figure S5C and D shows the estimated locations of the microinjector tips outside the RMTg based on the proximal vs distal site of misplaced microinjections. For the subset of cocaine-seeking rats that received the inactivation-alone test, there were no misplaced sites.
Food-Seeking Experiments
Extinction experiments
To determine whether the findings with cocaine-seeking behavior generalized to a non-drug reward, we conducted additional experiments examining food-seeking behavior. Figure 4a shows the active lever presses from the modified non-cue extinction sessions (days 1–5), in which microinjections were made immediately before the session, as well as the 7 days of standard non-cue extinction sessions (days 6–12), with no microinjections given. For the modified (15 min ) non-cue extinction sessions on days 1–5, a two-way repeated-measures ANOVA found no significant effect of group (F(1,16)=0.2118, p>0.05), a significant effect of time (F(4,64)=14.16, p<0.01), and no significant interaction (F(4,74)=1.076, p>0.05). For the standard (2 h) non-cue extinction sessions on days 6–12, a two-way repeated-measures ANOVA revealed no significant effect of group (F(1,16)=0.008, p>0.05), a significant effect of time (F(6,96)=20.76, p<0.0001), and a significant interaction (F(6,96)=2.337, p<0.05). However, post hoc tests for days 6–12 found no significant differences. Visual inspection of the data suggests that the significant interaction is primarily being driven by the relatively large decrease in active lever presses in the control group from days 6 to 7, vis-à-vis the PEPA group, on those days. However, this trend is opposite from what would be expected based on the cocaine-seeking data. Analyses of the inactive lever presses on days 1–5 and days 6–12 revealed no significant effects (data not shown).

Effect of RMTg activation on extinction following food self-administration and effects of RMTg inactivation on lever pressing during reinstatement tests. (a) Active lever presses (mean±SEM) during extinction sessions for those rats receiving pre-training intra-RMTg microinjections of PEPA or aCSF. Modified non-cue extinction sessions on days 1–5 were 15 min in length with microinjections given immediately before each session, whereas standard non-cue extinction sessions on days 6–12 were 2 h in length with no microinjections. There were no effects of PEPA treatment on active lever pressing for either the modified or standard non-cue extinction sessions. Two-way repeated-measures ANOVAs were also conducted on inactive lever pressing for days 1–5 and 6–12 of extinction learning and found no differences (p>0.05; data not shown). (b, c) Active and inactive lever presses, respectively, (mean±SEM) during a cue-induced reinstatement test for rats that had previously received aCSF during extinction training. (One rat from the extinction experiment did not reach extinction criteria after the cued extinction training in order to undergo reinstatement testing, reducing the total n for the reinstatement tests). Although initial analyses of the data suggested that RMTg inactivation potentiated food-seeking behavior, as with the cocaine seeking, further analyses based on prior aCSF/PEPA treatment during extinction training indicated an interaction between the prior treatment and the inactivation during the reinstatement tests. Therefore, the data from the reinstatement tests were split based on whether the rats had received aCSF or PEPA treatment during extinction (aCSF (b, c, f, g) and PEPA (d, e, h, i). In b, c, there were no significant differences among groups in either panel. (d, e) Active and inactive lever presses, respectively, (mean±SEM) during a cue-induced reinstatement session for rats that had previously received PEPA during extinction training. RMTg inactivation before the cue-induced reinstatement produced a non-significant trend toward increased active and inactive lever pressing. (f, g) Active and inactive lever presses, respectively, (mean±SEM) during the inactivation-alone session for rats that had previously received aCSF. RMTg inactivation had no effects on active lever pressing but produced a trend toward increased inactive lever pressing. (h, i) Active and inactive lever presses, respectively, (mean±SEM) during the inactivation-alone session for rats that had previously received PEPA. Analysis indicated trends toward significantly increased active and inactive lever pressing in those rats whose RMTg was inactivated. *p<0.05 compared with the extinction baseline. #p<0.1 compared with aCSF group. ^p<0.09 compared with extinction baseline and the aCSF group.
As with the cocaine-seeking studies, an additional analysis using a Mann–Whitney test was also conducted to determine whether there was a difference in the number of days of extinction the rats underwent before reaching the criteria to move onto the cued extinction training. There was no significant difference between those rats that had received PEPA (mean days±SEM=12.00±2.963) and those rats that had received aCSF (mean days±SEM=10.11±1.679) during extinction training (p>0.999). There was no difference in active lever pressing during the 2-h standard cued extinction sessions based on prior treatment (Supplementary Figure S1B; statistics in legend). There were not sufficient misplaced cannulas/injections in the food-seeking studies for analysis purposes.
Reinstatement experiments
Following the standard cue extinction procedures, rats underwent cue-induced reinstatement tests for food-seeking behavior. Although initial analyses suggested an effect of RMTg inactivation on lever pressing during the cue-induced reinstatement test, further analysis, surprisingly and in contrast to the cocaine-seeking experiments, indicated that prior treatment with PEPA interacted with the inactivation. Therefore, the reinstatement test data were analyzed separately based on prior PEPA vs aCSF treatment. As shown in Figure 4b–e, RMTg inactivation had no effect on lever pressing in control rats (ie, rats that had previously received aCSF during the extinction learning experiments) but appeared to increase lever pressing in those rats that had previously received PEPA. For rats that had received aCSF during extinction training, a one-way repeated-measures ANOVA of the active lever pressing (Figure 4b) during the cue-induced reinstatement test indicated a trend toward a significant effect (F(2,16)=3.659, p<0.08). However, post hoc analyses revealed no significant differences or trends toward differences (p>0.1). A one-way repeated-measures ANOVA of the inactive lever presses during the cue-induced reinstatement test (Figure 4c) found no significant effects (F(2,16)=0.6691, p>0.05). For rats that had received PEPA during extinction training, a one-way repeated-measures ANOVA of the active lever pressing during the cue-induced reinstatement tests (Figure 4d) found a significant effect (F(2,14)=6.687, p<0.05). Post hoc analyses found trends toward significant differences between the BM group and the aCSF group and the BM group and the extinction baseline (p<0.09 in both cases). A one-way repeated-measures ANOVA of the inactive lever pressing during the cue-induced reinstatement tests (Figure 4e) found a trend toward significant effect (F(2,14)=5.211, p<0.06). Post hoc analyses found a trend toward a significant difference between the aCSF and BM groups (p<0.1) but no other differences.
The same pattern of results found with cue-induced reinstatement tests was also found with the inactivation-alone tests (Figure 4f–i). For the aCSF rats, a one-way repeated-measures ANOVA of the active lever pressing during the inactivation-alone tests (Figure 4f) found no significant effect (F(2,16)=1.585, p>0.05). A one-way repeated-measures ANOVA of the inactive lever presses (Figure 4g) found a trend toward a significant effect (F(2,16)=4.226, p<0.07). Post hoc analyses found a trend toward a significant difference between the aCSF and BM groups (p<0.08) but no other differences. For the PEPA rats, a one-way repeated-measures ANOVA of the active lever pressing during the inactivation-alone tests (Figure 4h) found a significant effect (F(2,14)=6.785, p<0.05). Post hoc analyses found trends toward significant differences between the BM group and the aCSF group and the BM group and the extinction baseline (p<0.09 in both cases). A one-way repeated-measures ANOVA of the inactive lever pressing during the inactivation-alone tests (Figure 4i) found a significant effect (F(2,14)=6.689, p<0.05). Post hoc analyses found a significant difference between the BM group and the extinction baseline (p<0.05) and no other differences. Thus, RMTg inactivation appeared to increase lever pressing but primarily only in those rats that had previously received PEPA.
DISCUSSION
The present experiments investigated the role of the RMTg in the extinction and reinstatement of cocaine-seeking behavior. The findings indicate that RMTg activation, whether via pre- or post-training microinjections during the early modified non-cue extinction training after cocaine self-administration, altered active lever pressing in the subsequent standard non-cue extinction sessions. During cue-induced reinstatement tests or inactivation-alone tests, RMTg inactivation induced an increase in both active and inactive lever pressing in cocaine-seeking rats. The results from the locomotor tests suggest that RMTg inactivation increased overall motor activity. In contrast to the cocaine-seeking extinction, RMTg activation did not alter the extinction learning for food-seeking behavior. RMTg inactivation also had no effects during the food-seeking reinstatement tests, either cue-induced or inactivation alone, in control animals but increased lever pressing in those rats that had previously received PEPA during the extinction learning experiments. Together, the findings suggest that the RMTg has a role in behavioral inhibition and may be involved in the early extinction learning for cocaine-seeking behavior.
Previous work indicates a role for the RMTg as a contributor to the reward-prediction error signal of the VTA (Barrot et al, 2012; Bourdy et al, 2012). Such a signal would be expected to be critical for reward learning and, indeed, VTA dopamine neurons fire in a manner consistent with the idea that their activity serves to predict reward delivery (Glimcher, 2011; Schultz, 1998, 2013; Schultz et al, 1997). Moreover, dopamine neuron firing decreases below baseline when an expected reward is omitted, consistent with the reward-prediction error signal. Evidence suggests that negative stimuli, including footshocks and the omission of expected rewards, increase RMTg activity as well as Fos expression in RMTg neurons, whereas rewarding stimuli and cues predictive of reward decrease RMTg activity (Hong et al, 2011; Jhou et al, 2009a), a pattern opposite that of dopamine neurons. Combined with the other work mentioned, this strongly suggests that the RMTg is a critical component for modulating the reward-prediction error signal of VTA dopamine neurons. As learning about the omission of expected rewards would be expected to be a critical component of extinction learning, the present results, showing that potentiating RMTg activity during the modified non-cue extinction sessions appeared to alter the retention of extinction learning on the subsequent standard non-cue extinction sessions, are consistent with the hypothesized role of the RMTg. However, considering that work investigating the function of the RMTg is relatively new, it is critical to consider other possible explanations for the findings. It is possible that PEPA microinjections altered the animals’ memory regarding previous cocaine self-administration or desire for cocaine, rather than the extinction learning, producing a reduction in motivation to press the lever to receive cocaine. However, PEPA-injected rats and control rats show equivalent levels of lever pressing during the standard cued extinction sessions, suggesting that is not the case. Nonetheless, as studies continue to examine the function of this relatively newly described brain region, it will be critical for future work to consider all possibilities.
The lack of effect of RMTg manipulation on food-seeking extinction and, at least in control rats (ie, rats that had not previously received PEPA), on lever pressing during the reinstatement tests for food is surprising, considering that previous studies have shown that omission of expected food rewards increases RMTg activity (Jhou et al, 2009a). The reason for these discrepancies is not clear. As the current study used standard food pellets, rather than the sweet food (eg sucrose pellets) or drink (eg, apple juice) used in previous studies (Hong et al, 2011; Jhou et al, 2009a), it is possible that the palatability of the food/drink reward and/or the strength of the rewarding properties of the reinforcer accounts for the difference between the cocaine seeking. Nonetheless, in the present study, the rats self-administered the standard food pellets at robust levels. Another possibility is that cocaine administration altered RMTg plasticity and responsiveness in a manner different from that found with food self-administration. Indeed, the discovery of the RMTg as a distinct region began with the observation that psychostimulants such as cocaine induced a significant increase in Fos expression in a neuroanatomically distinct area that was eventually described as the RMTg (Geisler et al, 2008; Jhou et al, 2009b; Kaufling et al, 2009, 2010; Perrotti et al, 2005). Considering the aforementioned role of the RMTg in mediating aversive or ‘omission-of-expected-reward’ signals, it may be that cocaine activates the RMTg owing to its known aversive properties. Indeed, cocaine is known to have both rewarding and aversive components (Ettenberg, 2004; Guzman and Ettenberg, 2007; Su et al, 2013), and evidence suggests that the aversive properties are mediated by the lateral habenula–RMTg pathway. Lesions or inactivation of these regions prevent the avoidance behaviors that have been found with cocaine in a runway operant paradigm (Jhou et al, 2013). Based on these findings, the RMTg may be critically involved in the mediation of cocaine’s aversive properties, and, thus, cocaine may have a particular ability to influence plasticity in the structure, in contrast to the non-aversive food pellets, leading to the observed differences in the current work.
Indeed, this hypothesis may explain the differences in the food-seeking reinstatement tests between those rats that had previously received PEPA vs aCSF during extinction training. Repeated activation of the RMTg, either via cocaine or PEPA administration, may alter plasticity in the RMTg such that later inactivation produces changes in motivated behavior. Although PEPA does not directly activate AMPA receptors but rather potentiates their activation by endogenous glutamate (Sekiguchi et al, 1997), the present results suggest that such potentiation was sufficient to alter the functioning of the RMTg later during the reinstatement tests. The effect of prior treatment with PEPA in the food-seeking experiment, indicating an altered responsiveness of the RMTg to later inactivation and making those rats behave in a fashion similar to rats during cocaine-seeking reinstatement testing, raises this intriguing possibility. It is not known how such plasticity might occur in the RMTg or how it may be linked to ongoing behavioral suppression. If the lateral habenula–RMTg pathway is involved in both the negative reward-prediction error signal and general behavioral suppression, changes in synaptic strength at the lateral habenular inputs to the RMTg may produce greater activation of the circuit and thereby greater behavioral inhibition when inputs from other brain regions to the lateral habenula are activated. This could inhibit specific behaviors and/or general behavior depending on which other areas are activated for a given occasion. As the lateral habenula provides a glutamatergic input to the RMTg, future work should examine the potential for alterations in synaptic strength in these connections and attempt to determine whether such changes encode alterations in behavior.
Although it seems clear that the RMTg is involved in general behavioral inhibition, whether the RMTg is also involved in inhibiting specific motivated behavior is not clear based on the present data. RMTg inactivation had no effect on food-seeking behavior in the control rats (ie, those rats that had received aCSF during the extinction experiment), but RMTg inactivation during cocaine-seeking reinstatement tests increased both active and inactive lever pressing to a significant degree. Thus, the present findings suggest that RMTg inactivation produces a general behavioral disinhibition, at least under some circumstances. Prior work has found that RMTg lesions significantly reduce freezing during tone fear conditioning as well as freezing to a predator odor but increase defensive behavior for the same odor (Jhou et al, 2009a), suggesting a degree of direction for the increased behavior of the animal yet also a general increase in activity. Nonetheless, the present findings are distinctly opposite from those found with manipulations of the VTA itself, consistent with the findings indicating that the RMTg provides a strong inhibitory input to the VTA. Indeed, activation of VTA dopamine neurons induces or potentiates cocaine-seeking behavior, whereas inactivation or glutamate receptor blockade in the VTA blocks reinstatement of cocaine seeking (LaLumiere et al, 2012; Mahler et al, 2013; McFarland and Kalivas, 2001; Stewart, 1984). Thus, the present findings with inactivation suggest a role for the RMTg in general behavioral inhibition, although it should also be noted that, based on the analysis of misplaced microinjection sites, there may be multiple nuclei in the vicinity having a similar role (see ‘Methodological considerations’ below for a larger discussion).
That activation of the RMTg appears to influence the consolidation of extinction learning while RMTg inactivation also appears to produce general behavioral inhibition are not inconsistent with each other and may be owing, in part, to the use of different drugs. Whereas BM-induced inactivation would be expected to produce an overall inhibition of all neurons, PEPA would be expected to only potentiate active signaling, as PEPA requires endogenous glutamate release for its effects on the postsynaptic cells. Thus, PEPA may have acted to selectively increase specific activated inputs to the RMTg, in contrast to the less discrete effects of BM. Moreover, based on the emerging evidence regarding the RMTg, it is likely the RMTg may be both a component in the learning regarding the lack of reward and a mechanism by which other brain regions exert an inhibitory effect on motivated behavior. The apparent ability of the RMTg to influence extinction consolidation mirrors what has been previously found with the IL, suggesting a potential interaction between the IL and RMTg in mediating these effects, Peters et al (2008) found that IL inactivation induces cocaine-seeking behavior in rats that had previously undergone cocaine self-administration and extinction training. Later work demonstrated that IL activity regulates the consolidation of extinction learning that occurs during the early extinction process (LaLumiere et al, 2010). Because the RMTg receives inputs from the medial prefrontal cortex including the IL (Bourdy et al, 2012; Kaufling et al, 2009), IL activity could alter RMTg activity during extinction. However, in contrast to the IL, RMTg inactivation produced an overall increase in both active and inactive lever presses. Thus, if the IL interacts with the RMTg either during extinction learning or behavioral inhibition, it must do so utilizing mechanisms to produce a more specific behavioral output. Potentially, the role of the RMTg in behavioral inhibition may be more general, whereas suppression of specific behaviors may be controlled by forebrain structures that control activity in the RMTg more precisely than can be achieved with inactivating microinjections, as done in the present study.
Although the present discussion has assumed that the current results, including both the extinction learning and behavioral disinhibition results, originate from the effects of the RMTg on VTA dopamine neuron activity, the RMTg also provides an input to the lateral hypothalamus and the substantia nigra pars compacta (Kaufling et al, 2009), and these pathways in particular could account for the general behavioral disinhibition observed when the RMTg is inactivated. The lateral hypothalamus is known to be involved in driving motivated behavior, including drug-seeking behavior, as inactivation of the lateral hypothalamus reverses the alcohol-seeking behavior induced by NA shell inactivation (Millan et al, 2010, 2011). In addition, evidence indicates that activation of orexin neurons in the lateral hypothalamus drives drug-seeking behavior (Aston-Jones et al, 2009, 2010). The substantia nigra pars compacta is also known to provide dopaminergic innervation of structures involved in motivated behavior and motor activity (for review see Mercuri et al, 1992). Changes in dopamine neuron firing in the substantia nigra via RMTg inputs could thus influence dopamine levels in striatal regions and modulate cocaine-seeking behavior, motivated behavior, and/or motor activity. Indeed, RMTg inactivation may increase motivation and produce generalized behavioral activations via regulation of different pathways.
Methodological Considerations
An analysis of misplaced cannulas was also conducted for the cocaine experiments. There appeared to be no effect of PEPA microinjections outside the RMTg on extinction learning. In contrast, BM microinjections outside the RMTg appeared to increase overall lever pressing in the cue-induced reinstatement test. It may be that the behavioral disinhibition that occurs with RMTg inactivation is actually because of spreading of the BM to another nearby region. Further analysis of the sites of the misplaced microinjections, however, appeared to show no relationship between the location of the ‘misplacement’ and the resulting behavior. Possibly, the ‘misplaced’ microinjections enabled diffusion of the BM into the RMTg to inhibit a sufficient number of RMTg neurons to produce the overall increase in lever-pressing behavior. Indeed, because of the conservative approach used for determining the location of microinjections, it is likely that some of the misplaced microinjections actually terminated in the RMTg. However, analysis of the misplaced microinjections based on estimated proximity to the RMTg suggests that inactivation can produce increased lever pressing regardless of proximity of the microinjections, although there appeared to be more variability in the ‘distal’ missed microinjections. Considering that the distance between the distal microinjections and the RMTg were relatively large (>0.5 mm) and the aforementioned fact that the location of the misses had no relationship with the resulting behavior, it is possible that other nuclei in the vicinity may have functions similar to that of the RMTg and thus also be involved in behavioral inhibition, although our analysis of the literature has not led to any obvious candidates. Thus, these findings suggest that further studies will be required to parse out the functions of these brainstem regions, and that there may be even more heretofore undiscovered distinct nuclei in the vicinity with critical control over motivated behavior. The differences between the PEPA and BM effects with regard to missed placements suggest that the functions of these regions vis-à-vis the RMTg (eg, extinction learning vs behavioral inhibition) may not be completely identical.
As with most of the experiments in our prior study examining the role of the IL during extinction learning and consolidation (LaLumiere et al, 2010), the present studies did not find an effect of the microinjections during days 1–5 of extinction for cocaine seeking, likely owing to the high day-to-day variability that we have observed in the past with such short sessions. Of particular interest in the current study, however, is that the effect of the RMTg activation on the retention of the extinction learning was not apparent on the first day of the full-length (2 h) sessions but, rather, was only observed on subsequent days. This is in contrast to our previous work using this behavioral paradigm to investigate extinction learning wherein the retention effects were observed on the first day of the full-length sessions and were typically largest on that day (LaLumiere et al, 2010). Although it is unclear why the present study did not show a similar pattern, it may be that the relatively low lever pressing during the full-length extinction session, especially on the first full-length extinction day (day 6), compared with our previous work using this paradigm, reduced the likelihood of observing any differences.
CONCLUSIONS
The present findings indicate that RMTg activation during extinction training after cocaine, but not food, self-administration may alter the extinction process. The evidence also indicates, however, that RMTg inactivation produces a general behavioral disinhibition in rats that have undergone cocaine self-administration. These results suggest, therefore, that any role the RMTg has in inhibiting specific motivated behavior likely involves interactions with other systems that provide more specific control over the behavior.
FUNDING AND DISCLOSURE
Dr. RTL's work has been funded by the NIH. The authors declare no conflict of interest.
References
Aston-Jones G, Smith RJ, Moorman DE, Richardson KA (2009). Role of lateral hypothalamic orexin neurons in reward processing and addiction. Neuropharmacology 56 (Suppl 1): 112–121.
Aston-Jones G, Smith RJ, Sartor GC, Moorman DE, Massi L, Tahsili-Fahadan P et al (2010). Lateral hypothalamic orexin/hypocretin neurons: A role in reward-seeking and addiction. Brain Res 1314: 74–90.
Barrot M, Sesack SR, Georges F, Pistis M, Hong S, Jhou TC (2012). Braking dopamine systems: a new GABA master structure for mesolimbic and nigrostriatal functions. Journal Neurosci 32: 14094–14101.
Bourdy R, Barrot M (2012). A new control center for dopaminergic systems: pulling the VTA by the tail. Trends Neurosci 35: 681–690.
Ettenberg A (2004). Opponent process properties of self-administered cocaine. Neurosci Biobehav Rev 27: 721–728.
Geisler S, Marinelli M, Degarmo B, Becker ML, Freiman AJ, Beales M et al (2008). Prominent activation of brainstem and pallidal afferents of the ventral tegmental area by cocaine. Neuropsychopharmacology 33: 2688–2700.
Glimcher PW (2011). Understanding dopamine and reinforcement learning: the dopamine reward prediction error hypothesis. Proc Natl Acad Sci USA 108 (Suppl 3): 15647–15654.
Guzman D, Ettenberg A (2007). Runway self-administration of intracerebroventricular cocaine: evidence of mixed positive and negative drug actions. Behav Pharmacol 18: 53–60.
Hong S, Jhou TC, Smith M, Saleem KS, Hikosaka O (2011). Negative reward signals from the lateral habenula to dopamine neurons are mediated by rostromedial tegmental nucleus in primates. J Neurosci 31: 11457–11471.
Jhou TC, Fields HL, Baxter MG, Saper CB, Holland PC (2009a). The rostromedial tegmental nucleus (RMTg), a GABAergic afferent to midbrain dopamine neurons, encodes aversive stimuli and inhibits motor responses. Neuron 61: 786–800.
Jhou TC, Geisler S, Marinelli M, Degarmo BA, Zahm DS (2009b). The mesopontine rostromedial tegmental nucleus: A structure targeted by the lateral habenula that projects to the ventral tegmental area of Tsai and substantia nigra compacta. J Comp Neurol 513: 566–596.
Jhou TC, Good CH, Rowley CS, Xu SP, Wang H, Burnham NW et al (2013). Cocaine drives aversive conditioning via delayed activation of dopamine-responsive habenular and midbrain pathways. J Neurosci 33: 7501–7512.
Kaufling J, Veinante P, Pawlowski SA, Freund-Mercier MJ, Barrot M (2009). Afferents to the GABAergic tail of the ventral tegmental area in the rat. J Comp Neurol 513: 597–621.
Kaufling J, Veinante P, Pawlowski SA, Freund-Mercier MJ, Barrot M (2010). gamma-Aminobutyric acid cells with cocaine-induced DeltaFosB in the ventral tegmental area innervate mesolimbic neurons. Biol Psychiatry 67: 88–92.
LaLumiere RT, Niehoff KE, Kalivas PW (2010). The infralimbic cortex regulates the consolidation of extinction after cocaine self-administration. Learn Mem 17: 168–175.
LaLumiere RT, Smith KC, Kalivas PW (2012). Neural circuit competition in cocaine-seeking: roles of the infralimbic cortex and nucleus accumbens shell. Eur J Neurosci 35: 614–622.
Mahler SV, Smith RJ, Aston-Jones G (2013). Interactions between VTA orexin and glutamate in cue-induced reinstatement of cocaine seeking in rats. Psychopharmacology (Berl) 226: 687–698.
Matsumoto M, Hikosaka O (2007). Lateral habenula as a source of negative reward signals in dopamine neurons. Nature 447: 1111–1115.
McFarland K, Davidge SB, Lapish CC, Kalivas PW (2004). Limbic and motor circuitry underlying footshock-induced reinstatement of cocaine-seeking behavior. J Neurosci 24: 1551–1560.
McFarland K, Kalivas PW (2001). The circuitry mediating cocaine-induced reinstatement of drug-seeking behavior. J Neurosci 21: 8655–8663.
Mercuri NB, Calabresi P, Bernardi G (1992). The electrophysiological actions of dopamine and dopaminergic drugs on neurons of the substantia nigra pars compacta and ventral tegmental area. Life Sci 51: 711–718.
Millan EZ, Furlong TM, McNally GP (2010). Accumbens shell-hypothalamus interactions mediate extinction of alcohol seeking. J Neurosci 30: 4626–4635.
Millan EZ, Marchant NJ, McNally GP (2011). Extinction of drug seeking. Behav Brain Res 217: 454–462.
Nakagawa T, Iino M, Sekiguchi M, Wada K, Ozawa S (1999). Potentiating effects of 4-[2-(phenylsulfonylamino)ethylthio]-2,6-difluoro-phenoxyaceta mide (PEPA) on excitatory synaptic transmission in dentate granule cells. Neurosci Res 35: 217–223.
Paxinos G, Watson C (2007) The Rat Brain in Stereotaxic Coordinates 6th edn. Elsevier Academic Press: Amsterdam.
Perrotti LI, Bolanos CA, Choi KH, Russo SJ, Edwards S, Ulery PG et al (2005). DeltaFosB accumulates in a GABAergic cell population in the posterior tail of the ventral tegmental area after psychostimulant treatment. Eur J Neurosci 21: 2817–2824.
Peters J, LaLumiere RT, Kalivas PW (2008). Infralimbic prefrontal cortex is responsible for inhibiting cocaine seeking in extinguished rats. J Neurosci 28: 6046–6053.
Schultz W (1998). Predictive reward signal of dopamine neurons. J Neurophysiol 80: 1–27.
Schultz W (2013). Updating dopamine reward signals. Curr Opin Neurobiol 23: 229–238.
Schultz W, Dayan P, Montague PR (1997). A neural substrate of prediction and reward. Science 275: 1593–1599.
Sekiguchi M, Fleck MW, Mayer ML, Takeo J, Chiba Y, Yamashita S et al (1997). A novel allosteric potentiator of AMPA receptors: 4–2-(phenylsulfonylamino)ethylthio–2,6-difluoro-phenoxyaceta mide. J Neurosci 17: 5760–5771.
Steinberg EE, Keiflin R, Boivin JR, Witten IB, Deisseroth K, Janak PH (2013). A causal link between prediction errors, dopamine neurons and learning. Nat Neurosci 16: 966–973.
Stewart J (1984). Reinstatement of heroin and cocaine self-administration behavior in the rat by intracerebral application of morphine in the ventral tegmental area. Pharmacol Biochem Behav 20: 917–923.
Su ZI, Wenzel J, Ettenberg A, Ben-Shahar O (2013). Prior extended daily access to cocaine elevates the reward threshold in a conditioned place preference test. Addict Biol 19: 826–837.
Acknowledgements
We thank members of the LaLumiere laboratory for their helpful comments on a previous version of this manuscript. In addition, we thank Anne Curbow, Nathan Myhre, Rachel Gordon, Julia Collison, Holly Steger, Alyse Gray, Amy Pietro, Alec Wilcox, Alexandra Toft, and Erin Naffziger. This research was supported by NIH grant DA034684 (RTL).
Author information
Authors and Affiliations
Corresponding author
Additional information
Supplementary Information accompanies the paper on the Neuropsychopharmacology website
Rights and permissions
About this article

Cite this article
Huff, M., LaLumiere, R. The Rostromedial Tegmental Nucleus Modulates Behavioral Inhibition Following Cocaine Self-Administration in Rats. Neuropsychopharmacol 40, 861–873 (2015). https://doi.org/10.1038/npp.2014.260
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/npp.2014.260
This article is cited by
-
Rostromedial tegmental nucleus nociceptin/orphanin FQ (N/OFQ) signaling regulates anxiety- and depression-like behaviors in alcohol withdrawn rats
Neuropsychopharmacology (2023)
-
The rostromedial tegmental nucleus RMTg is not a critical site for ethanol-induced motor activation in rats
Psychopharmacology (2023)
-
From head to tail (of the VTA): role of projections from prelimbic cortex to rostromedial tegmental nucleus in cocaine reinstatement
Neuropsychopharmacology (2021)
-
Prelimbic cortical projections to rostromedial tegmental nucleus play a suppressive role in cue-induced reinstatement of cocaine seeking
Neuropsychopharmacology (2021)
-
Inhibition of the rostromedial tegmental nucleus reverses alcohol withdrawal-induced anxiety-like behavior
Neuropsychopharmacology (2019)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}