
Abstract
Desmoid-type fibromatosis (DTF, aggressive fibromatosis) is a non-metastasizing mesenchymal neoplasm of deep soft tissue with a tendency towards local recurrence. Genetic alterations affecting canonical Wnt/β-catenin signaling are reported in the majority of DTF. While most sporadic DTF harbor somatic mutations in CTNNB1, germline mutations in adenomatous polyposis coli (APC) are known to occur in hereditary DTF types (FAP, Gardner-Syndrome). Additional single nucleotide variants (SNVs) in AKT1 (E17K) and BRAF (V600E) were reported in pediatric DTF with potential clinical implications. We performed targeted next-generation sequencing (NGS) in a large cohort of 204 formalin-fixed DTF samples, comprising 22 pediatric cases (patients age ≤18 years). The mutational status was correlated with clinicopathological characteristics. Overall, deleterious CTNNB1 mutations were detected in 89% of DTF, most frequently affecting the serine/threonine phosphorylation sites T41 and S45 of β-catenin. While the T41A CTNNB1 mutation was significantly more often identified in the mesenterial localization, DTF originating from extra-intestinal sites more frequently harbored the S45P CTNNB1 alteration. Beyond common mutations in CTNNB1, additional SNVs were demonstrated in 7% of the DTF cohort and in 18% of the pediatric DTF subgroup. The mutational spectrum included deleterious mutations in AKT1 (G311S/D and T312I), ALK (R806H and G924S), AR (A159T), EGFR (P848L), ERBB2 (H174Y), IDH2 (H354Y), KIT (V559D), RET (T1038A), SDHA (R325M), and SDHD (R115W), as characterized by in silico prediction tools. In conclusion, our study indicates that DTF may harbor a broader mutational spectrum beyond CTNNB1 mutations, comprising targetable alterations including the herewith first reported imatinib-sensitive KIT V559D mutation in DTF.
Similar content being viewed by others

Introduction
Desmoid-type fibromatosis (aggressive fibromatosis, desmoid tumor, DTF) is an infiltrating, locally aggressive myofibroblastic neoplasm of intermediate malignant potential highly prone to local recurrence without the potential for metastatic spread. The tumors typically arise in deep soft tissue compartments of intra- and extra-abdominal localization. Pediatric forms usually affect the extremities, while in adults also the mesenterium and the abdominal wall are commonly involved sites; the latter predominantly affected in women1.
The majority of DTF harbor mutations affecting the canonical Wnt/β-catenin signaling pathway2. While in patients with familial adenomatous polyposis (FAP) β-catenin is not degraded through inactivating mutations in APC, most sporadic DTF harbor alterations in CTNNB1; both leading to a nuclear accumulation of β-catenin and an oncogenic activation of the Wnt/β-catenin signal transduction pathway3,4,5,6. In the sporadic subtype, alterations in CTNNB1 seem to be focused on the serine/threonine phosphorylation sites T41 and S457,8 with a higher risk of local recurrence reported in association with the S45F CTNNB1 mutation8,9.
Currently, no evidence-based approach for the treatment of DTF is established10 and the clinical course is still unpredictable with spontaneous regression as well as progression and long-lasting stable disease being reported for individual cases. Given the broad spectrum of clinical presentation, initial treatment decisions are highly individualized. However, primary en bloc surgery is no longer regarded as mandatory, as high recurrence rates (up to 60%) have been reported11. Currently, a shift towards a “wait and see” strategy is assessed by selected clinical trials and recommended by different studies10,12,13. In a retrospective study, Penel et al. reported no significant difference with regard to the outcome between surgical vs. non-surgical treatment approaches14. However, these results still have to be evaluated in prospective trials.
Several trials evaluated the benefit of additional systemic therapies in DTF. The multitargeted receptor tyrosine kinase inhibitor sorafenib prolonged progression-free survival in a double-blind phase 3 trial including 87 patients with progressive, symptomatic or recurrent DTF15. Furthermore, in a phase 2 study, the tyrosine kinase inhibitor imatinib was shown to represent a treatment option in CTNNB1 mutated DTF, especially in tumors carrying the S45F alteration16. Another phase 2 study reported on the efficacy of the Notch pathway inhibitor PF-03084014 in patients with pre-treated, progressive and symptomatic DTF17. The rationale to inhibit the Notch signaling pathway is based on the demonstration of signaling cross talk between the Notch- and the Wnt/β-catenin pathways17,18. There seem to be several intrinsic (i.e. mutational status or DTF tumor localization) and extrinsic (i.e. initial treatment decision) factors that may influence the biology of the disease.
The majority of genetic studies published so far applied conventional Sanger sequencing of CTNNB1 (limited to exon 3), focusing predominantly on the mutational subtype of sporadic DTF. Recently, Meazza et al. analyzed a small cohort of DTF employing NGS for mutational analysis and reported potential clinical implications for pediatric (patients age ≤18 years) DTF harboring gain-of-function mutations in AKT1 and BRAF19. These findings suggest a more complex mutational spectrum of DTF than expected so far, which may potentially be associated with therapeutic implications.
In our present genetic study, we performed an NGS-based comprehensive molecular characterization of 204 DTF cases comprising approximately 10% of pediatric samples. Genotyping results were correlated with clinical and pathological features with special emphasis on the mutational spectrum beyond CTNNB1 in pediatric and adult DTF.
Results
Spectrum of CTNNB1 alterations in DTF samples
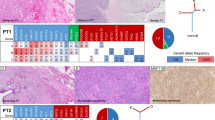
As pathogenic mutations in the CTNNB1 gene can be responsible for the constitutive activation of the canonical Wnt/β-catenin signaling cascade, we analyzed the entire CTNNB1 coding region by targeted NGS followed by validation via Sanger sequencing. Overall, the complete cohort of 204 DTF cases was successfully analyzed (Table 1 and Supplementary Table S1). In total, deleterious CTNNB1 mutations were detected in n = 181/204 (88.7%) DTF samples, with a minor fraction detected in the pediatric (77.3%) compared to the adult (89.9%) subgroup (Fig. 1A and Table 2). All four tumors in patients with familial adenomatous polyposis (FAP) analyzed in this study, were assigned to the CTNNB1 wild type group (n = 23/204; 11.3%). The majority of deleterious alterations in the CTNNB1 gene were restricted to the serine/threonine phosphorylation sites T41 and S45, including T41A (n = 111/204; 54.4%), S45F (n = 40/204; 19.6%), S45P (n = 18/204; 8.8%) or T41I (n = 5/204; 2.5%) amino acid exchanges (Fig. 1B and Table 2). Detected allelic frequencies were in the range from minimal 6% (S45P) to 13% (T41I) and maximal 34% (T41I) to 58% (T41A) (Fig. 1C). Less frequent CTNNB1 mutations were detected in individual DTF cases in the adult subgroup, comprising S33T (n = 1/204), S33L (n = 1/204), G34R (n = 1/204) and S45KR (n = 1/204) amino acid exchanges. In addition, biallelic mutations (S45T/S45Y, S45T/S45F and S45P/S45F) were identified in four adult samples. Comparing the pediatric (≤18 y/a) to the adult (>18 y/a) age groups, the fraction of CTNNB1 wildtype cases was larger in the pediatric (n = 5/22; 22.7%) vs. the adult (n = 18/170; 10.5%) group. Deleterious T41A (n = 97/170; 56.7% vs. n = 8/22; 36.4%) and S45P (n = 16/170; 9.4% vs. n = 1/22; 4.5%) β-catenin alterations were detected more often in adult DTF patients, whereas three out of four T41I variants were demonstrated in the pediatric subgroup (n = 1/170; 0.6% vs. n = 3/22; 13.6%). All pediatric DTF cases were localized in extra-intestinal sites in contrast to the frequent occurrence of extra-intestinal localization in adult DTF cases (n = 20/20; 100% vs. n = 86/164; 52.4%, p < 0.05).

(A) Frequency of detected CTNNB1 mutations in the pediatric and adult DTF subgroup (n = 204). (B) Spectrum of detected CTNNB1 mutations. (C) Range of allelic frequencies for the most common CTNNB1 mutation subtypes.
Correlating the individual CTNNB1 mutational status and tumor site, we observed a significantly higher incidence of deleterious T41 alterations in intra-abdominal compared to abdominal and extra-abdominal DTF cases (n = 43/59; 72.9% vs. n = 63/125; 50.4%; p = 0.0042). In addition, β-catenin mutations affecting S45 were detected significantly more frequently in extra-abdominal cases than in intra-abdominal and abdominal DTF cases (n = 36/106; 34.0% vs. n = 16/78; 20.51%, p = 0.0222) (Table 2). Concerning a possible prognostic relevance of the mutational subtype, recurrences were reported in 18 cases, occurring in extra-intestinal localization in 17 cases.
Only n = 6/131 (4.3%) CTNNB1 mutations detected by NGS could not be validated by Sanger sequencing due to low allelic frequencies in the range of 6.0–17.6% (Fig. 2A,B). Tumor tissue for immunohistochemical analysis was available in 131 DTF cases. Nuclear β-catenin positivity correlated with the CTNNB1 mutational status in 92.4% (n = 121/131) of DTF cases (Fig. 2C,D).

(A) Concordance of CTNNB1 mutational status comparing NGS and Sanger sequencing results. (B) Representative visualization of NGS and Sanger sequencing results (case #60 and #31). (C) Concordance of CTNNB1 mutational status and nuclear β-catenin immunoreactivity. (D) Representative HE and nuclear β-catenin IHC staining (case #203).
In three cases, more than one missense mutation was detected in CTNNB1 (Supplemental Table S1). A 26-year old female with a DTF localized at the trunk (extra-intestinal) harbored both a S45T and a S45Y mutation in CTNNB1 (case #26). The combination of S45F and S45T CTNNB1 mutations was detected in a 32-year old female patient with abdominal DTF of unknown size (case #149). A combination of uncommon S33T and G34R CTNNB1 alterations was detected in a 43-year old female patient with a 4.8 cm abdominal DTF (case #119). No additional non-CTNNB1 mutations were detected by NGS in these cases.
To the best of our knowledge, two uncommon alterations were detected in CTNNB1: (I) S33L (97_98delinsCT) in an intra-abdominal DTF of a 60-year old male without additional mutations (case #181) and (II) S45KA in a 74-year old male with an abdominal 10.5 cm DTF (case #51). Both variants were predicted as potentially deleterious by in silico analysis.
Spectrum of non-CTNNB1 alterations in DTF samples
All protein coding exons of 26 additional cancer-associated genes were analyzed by NGS including: AKT1, ALK, AR, BRAF, DDR2, EGFR, ERBB2, FGFR3, GNA11, GNAQ, GNAS, IDH1, IDH2, KIT, KRAS, MET, NRAS, PDGFRA, PIK3CA, PTEN, RET, SDHA, SDHB, SDHC, SDHD and TP53. In 31 out of 204 (15.2%) DTF cases, additional non-CTNNB1 alterations were detected (summarized in Fig. 3A,B and Supplementary Tables S1 and S2). No genetic alterations were identified in n = 3/22 (13.6%) pediatric and n = 18/170 (10.6%) adult DTF cases. Overall, 56.8% (n = 21/37) of the non-synonymous variants (in AKT1, ALK, AR, EGFR, ERBB2, IDH2, KIT, KRAS, RET, SDHA and SDHD) were predicted to have a potentially deleterious impact by ≥3 independent in silico tools (Supplementary Table S3). In contrast, all five variants identified in FGFR3, MET and PDGFR were classified in silico as potentially neutral/tolerated. Notably, deleterious mutations were more often demonstrated simultaneously to CTNNB1 alterations affecting the phosphorylation site T41 compared to S45 (n = 13/21; 61.9% vs. n = 5/21; 23.8%; p = 0.0278) with three deleterious mutations exclusively identified in two CTNNB1 wild type pediatric DTF cases. In total, deleterious non-CTNNB1 mutations were demonstrated in n = 4/22 (18.2%; in AKT1, ALK, IDH2, RET and SDHD) pediatric vs. n = 15/170 (8.8%; in AKT1, ALK, AR, EGFR, ERBB2, IDH2, KIT, KRAS, RET and SDHA) adult DTF cases.

(A) Clustered mutational profile of pediatric and adult DTF (n = 204). Alterations in different genes (rows) are indicated for each sample (columns). Clinicopathological information and in silico prediction results on the potential deleterious impact of detected gene variants are summarized according to the legend. (B) Spectrum of single nucleotide substitutions identified in DTF and defined by the sequence context immediately 3′ and 5′ to the mutated nucleobase.
In one case (case #202), an imatinib-sensitive KIT (V559D) mutation was detected in addition to the CTNNB1 (T41A) mutation. The tumor was localized in the Musculus rectus abdominis of a 43-year old female patient and displayed a diffuse infiltrative growth.
A combined S45P and S45F mutation was detected in a 25-year old male with an 8.0 cm DTF affecting the trunk (case #37). In this tumor, additional mutations were identified in AR (A159T) and SDHA (T36I). In silico analysis of the additional mutations indicated no pathogenic potential. Of note, in one case, a similar combination of S45P and S45F CTNNB1 alterations and an A159T mutation in AR was detected with no clinical data available (case #38).
The known KIT polymorphism M541L was demonstrated in 16% of the analyzed DTF cases. Analyzing the mutational spectrum of single nucleotide substitutions from all n = 183/204 mutated DTF samples, C > T substitution was identified as major process (Fig. 3B).
DISCUSSION
In this study, we attempted to refine the mutational spectrum in DTF beyond the well-established alterations affecting the canonical Wnt/β-catenin signaling pathway2,3,4,5,6. We conducted a comprehensive molecular characterization of 204 DTF cases applying NGS.
Based on our previous results analyzing mesenteric DTF20, we first examined the mutational CTNNB1 status in correlation with clinicopathological features. Previous studies reported genomic CTNNB1 alterations in 67–92% of sporadic DTF cases demonstrated by conventional Sanger sequencing and limited to CTNNB1 exon 37,8,9,19,20,21 (summarized in Table 3). In our present study, we conducted NGS with an increased sensitivity (≥10% allelic frequency) and detected deleterious CTNNB1 mutations in an enlarged fraction of n = 181/204 (89%) DTF cases. Due to low allelic frequencies, 4.3% of CTNNB1 mutations were exclusively traceable by NGS (The mean region coverage depth was 377×) and not detected by Sanger sequencing. With regard to clinical data, 4 cases (one pediatric) were known to be associated with FAP and harbored, as expected, no mutation in CTNNB1.
As reported by previous studies, the majority of deleterious alterations in the CTNNB1 gene were restricted to the serine/threonine phosphorylation sites T41 and S45. In our study, T41A (n = 111/204; 54.4%), S45F (n = 40/204; 19.6%) and S45P (n = 18/204; 8.8%) were the most frequent amino acid exchanges affecting sites involved in the modulation of the interaction of β-catenin with several kinases: Sequential phosphorylation at T41, S33 and S37 is mediated by the glycogen synthase kinase 3 beta (GSK-3β), while S45 is phosphorylated by the casein kinase-1 alpha (CK1α) targeting β-catenin for ubiquitination and subsequently proteasomal degradation5,22,23. Mutations affecting these phosphorylation sites lead to stabilization and translocation of β-catenin into the nucleus where it acts as a transcriptional regulator. A schematic overview of β-catenin indicating relevant protein domains, phosphorylation sites, interaction partners and associated cellular functions is shown in Fig. 4.

Schematic overview of β-catenin indicating relevant protein domains, phosphorylation sites, interaction partners and associated cellular functions.
Several genotype-phenotype correlations have been published so far with the S45F alterations found to predict a higher tendency for local recurrence in DTF7,8,9. In accordance with results published by Colombo et al., we detected CTNNB1 mutations in association with S45 significantly more often in extra-abdominal than in intra-abdominal or abdominal DTF cases (n = 36/106; 36.0% vs. n = 16/78; 20.51%, p = 0.0222). As previously indicated for mesenteric DTF20, we observed a significantly higher incidence of deleterious T41 alterations in intra-abdominal compared to abdominal and extra-abdominal DTF cases (n = 43/59; 72.9% vs. n = 63/125; 50.4%; p = 0.0042). Recently, the French Sarcoma group published data from a nationwide prospective study indicating that tumor localization constitutes a major prognostic factor. Two-year event free survival was significantly higher in favorable localizations than in unfavorable sites which was mainly extra-intestinal localization (except the lower limb)14.
We observed phenotypic and genetic differences comparing the pediatric and adult DTF groups. All pediatric DTF were localized extra-intestinal (100%; n = 20/20), while only 52.4% (n = 86/164) of DTF in adult patients occurred extra-intestinal. With regard to the data from the French Sarcoma group, an increased risk for local recurrence is implicated for the pediatric DTF group. For both DTF subgroups, the majority of deleterious alterations in the CTNNB1 gene were restricted to the threonine phosphorylation site T41, with T41A being the most common variant in the pediatric (n = 8/22; 36.4%) and the adult (n = 97/170; 56.7%) DTF subgroup. Interestingly, three of the less frequent CTNNB1 T41I variant were predominately demonstrated in pediatric DTF patients (n = 3/22; 13.6%), compared to only a single adult DTF case (n = 1/170; 0.6%) which has not been reported by other studies including pediatric DTF patients before19,24,25.
Crago et al. performed whole-exome sequencing of DTF and demonstrated additional genomic alterations in association with the canonical Wnt/β-catenin signaling pathway (i.e. loss of chromosome 6 or BMI1 mutations), further substantiating the idea that β-catenin is the major oncogenic driver in desmoid tumor development and progression. However, in a small cohort of 26 DTF cases focusing mainly on pediatric DTF, Meazza et al. reported deleterious aberrations in AKT1 (E17K; n = 8/26, 30.7%) and BRAF (V600E; n = 5/26,19.2%)19. These findings suggest a more complex mutational spectrum occurring at least in a subset of DTF than expected so far, including potentially targetable alterations that could be exploited for therapeutic benefit. To define the spectrum of non-CTNNB1 alterations in our large DTF cohort, all protein coding exons of 26 additional cancer-associated genes were analyzed by NGS. In 15.2% of DTF cases, additional non-CTNNB1 alterations could be detected, and 56.8% of the non-synonymous variants in AKT1, ALK, AR, EGFR, ERBB2, IDH2, KIT, KRAS, RET, SDHA and SDHD were predicted to have a potentially deleterious impact by ≥3 independent in silico tools. Overall, deleterious non-CTNNB1 mutations were demonstrated in 18.2% (AKT1, ALK, IDH2, RET and SDHD) pediatric vs. only 8.8% (AKT1, ALK, AR, EGFR, ERBB2, IDH2, KIT, KRAS, RET and SDHA) in adult DTF cases. For further evidenced-based variant categorization, we aim to classify all detected somatic variants into four tiers based on currently available clinical and experimental evidence to determine the level of clinical significance in DTF patients for each genomic alteration as proposed in the Standards and Guidelines for the Interpretation and Reporting of Sequence Variants in Cancer26. In contrast to the results published by Meazza et al., only a single case of 22 pediatric DTF patients harbored a deleterious AKT1 (G311D/S) mutation with unknown therapeutic relevance. A second deleterious AKT1 (T312I) mutation was discovered in an adult DTF patient. In our entire cohort, no pathogenic BRAF (V600E) mutation was detected.
To the best of our knowledge, we herewith report the first DTF case with a synchronous CTNNB1 (T41A) and imatinib-sensitive KIT (V559D) mutation (case #202). The tumor was localized in the Musculus rectus abdominis of a 43-year old female patient, displayed a diffusely infiltrative growth pattern and showed nuclear expression of β-catenin. The imatinib-sensitive KIT (V559D) alteration is known to cause ligand-independent tyrosine kinase activity and is supposed to play a central role in the pathogenesis of GIST27,28,29. If this KIT (V559D) mutation represented an additional oncogenic alteration in DTF, one could hypothesize that imatinib therapy might be effective. However, the clinical impact of additional deleterious mutations in the case of CTNNB1 mutated DTF cases remains uncertain. Tyrosine kinase inhibition has been evaluated in a phase 2 study of the German Interdisciplinary Sarcoma Group (GISG-01)16. The authors describe a positive correlation of CTNNB1 mutational status and progression arrest rate after imatinib treatment, especially in DTF tumors harboring a CTNNB1 S45F alteration which were overrepresented in that study. In accordance with previously published results30,31, the known KIT polymorphism M541L was demonstrated in 16% of all analyzed DTF cases. In a prospective series by Dufresne et al., no predictive effect of the M541L variant on the efficacy of imatinib therapy for patients with DTF was seen30.
As the current study was retrospectively designed and focused on the mutational spectrum of different DTF subgroups, one of the major limitations of this study is the sparse clinical follow-up information available to us. Complementary studies comprising larger DTF cohorts are needed to correlate the mutational spectrum of both, the pediatric as well as the adult DTF subgroup to clinical outcome taking into consideration e.g. the primary localization as prognostic factor. Additional information of potential therapeutic benefit will further be gained by expanding the NGS panel used in this study (limited to the exonic region of only 27 genes) to include additional genes with targetable alterations. Furthermore, germline APC mutations were not validated for patients with FAP.
In conclusion, we report deleterious CTNNB1 mutations in n = 181/204 (89%) patients with DTF confirming previous studies. Further mutational analysis indicates that a subset of DTF harbor a more complex mutational spectrum comprising not only deleterious alterations in CTNNB1 but also potentially targetable alterations in non-Wnt/β-catenin signal transduction effectors that could be further exploited for therapeutic approaches. As an example, we herewith report the first imatinib-sensitive KIT (V559D) mutation in DTF which deserves further clinical investigation.
Materials and Methods
Tumor specimens/clinicopathological features
In total, 204 DTF tissue specimens were selected from the archive of the GIST/Sarcoma Registry (University Münster, Germany) and the Gerhard-Domagk-Institute of Pathology (Münster University Hospital, Germany). According to the current WHO classification1, all DTF diagnoses were reviewed by four experienced pathologists (EW, IG, SH, WH) based on morphological criteria, clinical information, immunohistochemical and molecular analyses as described before20. Scientific and retrospective analysis of the cohort of mesenchymal tumors was approved by the Ethics Review Board (University of Münster, 2016-091-f-S). Written informed consent from the patients was not requested and was waived by the Ethics Review Board. The study did comply with the principles set out in the United States Department of Health and Human Services Belmont Report and the World Medical Association Declaration of Helsinki. Clinicopathological data were available for 192 (94%) DTF patients (female: n = 118, 61% and male: n = 74, 39%). The cohort included 22 pediatric (≤18 y/a; mean years of age 11.9) and 170 adult (>18 y/a; mean years of age 44.9) DTF patients. With regard to clinical data, 4 cases (2%; one pediatric) were known to be associated with FAP and 200 (98%) cases were sporadic. Median tumor size was 7.0 cm (range 1-28 cm). Tumor sites were categorized into (I) extra-intestinal (n = 106, 58%), (II) intra-abdominal/mesenteric (n = 59, 32%) and (III) abdominal/abdominal wall (n = 19, 10%). All tumors within the pediatric subgroup were exclusively localized extra-intestinally. Detailed clinicopathological patient and tumor characteristics are summarized in Table 1 and individualized in Supplementary Table S1.
Next-generation sequencing (NGS)/Sanger sequencing
A customized Qiagen GeneRead DNAseq Mix-n-Match V2 panel (in total 1028 amplicons covering 63.8 kb) was used to sequence the complete exonic region of 27 genes (AKT1, ALK, AR, BRAF, CTNNB1, DDR2, EGFR, ERBB2, FGFR3, GNA11, GNAQ, GNAS, IDH1, IDH2, KIT, KRAS, MET, NRAS, PDGFRA, PIK3CA, PTEN, RET, SDHA, SDHB, SDHC, SDHD and TP53) that are commonly mutated in cancer. As described before32,33,34, Target enrichment was conducted by means of the GeneRead DNAseq Panel PCR V2 Kit (Qiagen). Purification and size selection steps were processed utilizing Agencourt AMPure XP magnetic beads (Beckman Coulter). The GeneRead DNA Library I Core Kit (Qiagen) was applied to perform end repair, A-addition and ligation to NEXTflex-96 DNA barcodes (Bioo Scientific). The HiFi PCR Master Mix (GeneRead DNA I Amp Kit, Qiagen) and NEXTflex oligonucleotides (Bioo Scientific) were used for the amplification of adapter-ligated DNA. Sequencing was performed on the MiSeq system (Illumina; 12.5 pM library pools, 2% PhiX V3 control and MiSeq Reagent v2 chemistry). We used the Quantitative Multiplex FFPE Reference Standard (Horizon Discovery, ref.no. HD200, 11 somatic variants verified at 0.8–24.5% allelic frequency) as isogenic quality control for routine performance evaluation and monitoring of NGS workflow integrity (pre analytical DNA extraction, NGS workflow and post-analytical bioinformatics). The CLC Biomedical Genomics Workbench software (CLC bio, Qiagen) was used for NGS data analysis. Validation by Sanger sequencing (CTNNB1 exon 3) was performed using the BigDye Terminator Cycle Sequencing Kit (v3.1, Life Technologies). Following primer set was applied: 5′-CTG ATT TGA TGG AGT TGG ACA TGG CCA TG-3′ (forward) and 5′-CCA GCT ACT TGT TCT TGA GTG AAG GAC TGA G-3′ (reverse).
In silico tools to predict the potential deleterious impact of detected variants
The potential impact of NGS-detected variants in the coding regions was predicted using following in silico tools as reported32,33,34: PolyPhen-2 (v2.2.2r398)35, Protein Variation Effect Analyzer (PROVEAN; v1.1.3)36, Sorting Intolerant From Tolerant (SIFT; Ensembl 66)37,38, Mutation Assessor (release 3)39 and Combined Annotation Dependent Depletion (CADD; v.1.3)40. The PolyPhen-2 method utilizes physical and evolutionary comparative considerations to predict amino acid changes on protein structure and function. Scoring is in the range from 0 (neutral) to 1 (deleterious) and potential functional significance is categorized into benign, possibly damaging, and probably damaging. The PROVEAN web-based algorithm classified gene variants as either potentially neutral or deleterious (cutoff −2.5). SIFT was used with the default settings, classifying gene variants from 0 (damaging) to 1 (tolerated). The CADD algorithm is trained to differentiate 14.7 million high-frequency human-derived alleles for integration of diverse annotations into a single measure (C score). Scoring correlates with allelic diversity, annotations of pathogenicity, disease severity, complex trait associations, and experimentally measured regulatory effects. Calculated C scores rank a pathogenic alteration relative to all potential substitutions of the human genome. To increase the prediction accuracy and the level of confidence, a combination of in silico algorithms based on functional parameters, protein structure and evolutionary information was used. We combined the individual output from ≥3 of five in silico prediction tools and produced a single consensus outcome summarized in Supplementary Table S3.
Immunohistochemistry (IHC)
IHC was performed on 3 µm sections from paraffin-embedded tissue using the automated Ventana BenchMark ULTRA IHC staining system (Roche). Tissue sections were deparaffinized and pre-treated with Cell Conditioning 1 solution (CC1, Ventana/Roche) for 24–64 minutes at 95–100 °C. The primary antibodies were then incubated for 16–32 minutes at 36 °C. Immunoreactions were visualized via the Optiview DAB IHC detection kit (ref.no. 760–700, Ventana/Roche). Tissue sections were counterstained with hematoxylin (ref.no. 790-2208, Ventana/Roche) and bluing solution (ref.no. 760-2037, Ventana/Roche). The mouse polyclonal β-catenin antibody (ref.no. CMC 22421040, clone 14, CellMarque) was applied. Only nuclear localization of β-catenin was considered positive for the purpose of the study, and the percentage of stained tumor cells was estimated in 10 high-power microscopic fields (400x magnification). DTF cases with ≥10% immunoreactive cells were regarded as positive. Following antibodies were additionally applied to exclude differential diagnoses: CD34 (ref.no. 790-2927, clone QBEnd10, Ventana/Roche), DOG1 (ref.no. CMC 24431050, clone SP31, CellMarque), CDK4 (ref.no. AHZ0202, DCS-31, Invitrogen/Thermo Fisher Scientific) and MDM2 (ref.no.33-7100, IF2, Invitrogen/Thermo Fisher Scientific).
Statistical analysis
Clinical parameters of the cohort are based on descriptive statistics (comprising numbers and percentages, median and mean standard deviation). Chi-square-, Fisher’s exact- and t-tests (two-tailed with a 95% confidence interval) were calculated (SPSS 20 software, IBM and GraphPad Prism).
References
Fletcher, C. D. M. & Organization, W. H. WHO Classification of Tumours of Soft Tissue and Bone. (IARC Press, 2013).
Crago, A. M. et al. Near universal detection of alterations in CTNNB1 and Wnt pathway regulators in desmoid-type fibromatosis by whole-exome sequencing and genomic analysis. Genes, chromosomes & cancer 54, 606–615, https://doi.org/10.1002/gcc.22272 (2015).
Jilong, Y., Jian, W., Xiaoyan, Z., Xiaoqiu, L. & Xiongzeng, Z. Analysis of APC/beta-catenin genes mutations and Wnt signalling pathway in desmoid-type fibromatosis. Pathology 39, 319–325, https://doi.org/10.1080/00313020701329823 (2007).
Giardiello, F. M. et al. APC gene mutations and extraintestinal phenotype of familial adenomatous polyposis. Gut 40, 521–525, https://doi.org/10.1136/gut.40.4.521 (1997).
Tejpar, S. et al. Predominance of beta-catenin mutations and beta-catenin dysregulation in sporadic aggressive fibromatosis (desmoid tumor). Oncogene 18, 6615–6620, https://doi.org/10.1038/sj.onc.1203041 (1999).
Kinzler, K. W. et al. Identification of FAP locus genes from chromosome 5q21. Science 253, 661–665, https://doi.org/10.1126/science.1651562 (1991).
Domont, J. et al. High frequency of beta-catenin heterozygous mutations in extra-abdominal fibromatosis: a potential molecular tool for disease management. British journal of cancer 102, 1032–1036, https://doi.org/10.1038/sj.bjc.6605557 (2010).
Lazar, A. J. et al. Specific mutations in the beta-catenin gene (CTNNB1) correlate with local recurrence in sporadic desmoid tumors. The American journal of pathology 173, 1518–1527, https://doi.org/10.2353/ajpath.2008.080475 (2008).
Colombo, C. et al. CTNNB1 45F mutation is a molecular prognosticator of increased postoperative primary desmoid tumor recurrence: an independent, multicenter validation study. Cancer-Am Cancer Soc 119, 3696–3702, https://doi.org/10.1002/cncr.28271 (2013).
Kasper, B. et al. Management of sporadic desmoid-type fibromatosis: a European consensus approach based on patients’ and professionals’ expertise - a sarcoma patients EuroNet and European Organisation for Research and Treatment of Cancer/Soft Tissue and Bone Sarcoma Group initiative. European journal of cancer 51, 127–136, https://doi.org/10.1016/j.ejca.2014.11.005 (2015).
Crago, A. M. et al. A prognostic nomogram for prediction of recurrence in desmoid fibromatosis. Annals of surgery 258, 347–353, https://doi.org/10.1097/SLA.0b013e31828c8a30 (2013).
Penel, N. et al. Management of desmoid tumours: A nationwide survey of labelled reference centre networks in France. European journal of cancer 58, 90–96, https://doi.org/10.1016/j.ejca.2016.02.008 (2016).
Salas, S. et al. Prognostic factors influencing progression-free survival determined from a series of sporadic desmoid tumors: a wait-and-see policy according to tumor presentation. Journal of clinical oncology: official journal of the American Society of Clinical Oncology 29, 3553–3558, https://doi.org/10.1200/JCO.2010.33.5489 (2011).
Penel, N. et al. Surgical versus non-surgical approach in primary desmoid-type fibromatosis patients: A nationwide prospective cohort from the French Sarcoma Group. European journal of cancer 83, 125–131, https://doi.org/10.1016/j.ejca.2017.06.017 (2017).
Gounder, M. M. et al. Sorafenib for Advanced and Refractory Desmoid Tumors. The New England journal of medicine 379, 2417–2428, https://doi.org/10.1056/NEJMoa1805052 (2018).
Kasper, B. et al. Correlation of CTNNB1 Mutation Status with Progression Arrest Rate in RECIST Progressive Desmoid-Type Fibromatosis Treated with Imatinib: Translational Research Results from a Phase 2 Study of the German Interdisciplinary Sarcoma Group (GISG-01). Annals of surgical oncology 23, 1924–1927, https://doi.org/10.1245/s10434-016-5132-4 (2016).
Shang, H. et al. Targeting the Notch pathway: A potential therapeutic approach for desmoid tumors. Cancer-Am Cancer Soc 121, 4088–4096, https://doi.org/10.1002/cncr.29564 (2015).
Kim, H. A. et al. Notch1 counteracts WNT/beta-catenin signaling through chromatin modification in colorectal cancer. The Journal of clinical investigation 122, 3248–3259, https://doi.org/10.1172/JCI61216 (2012).
Meazza, C. et al. AKT1 and BRAF mutations in pediatric aggressive fibromatosis. Cancer medicine 5, 1204–1213, https://doi.org/10.1002/cam4.669 (2016).
Huss, S. et al. beta-catenin (CTNNB1) mutations and clinicopathological features of mesenteric desmoid-type fibromatosis. Histopathology 62, 294–304, https://doi.org/10.1111/j.1365-2559.2012.04355.x (2013).
Amary, M. F. et al. Detection of beta-catenin mutations in paraffin-embedded sporadic desmoid-type fibromatosis by mutation-specific restriction enzyme digestion (MSRED): an ancillary diagnostic tool. The American journal of surgical pathology 31, 1299–1309, https://doi.org/10.1097/PAS.0b013e31802f581a (2007).
Gao, C. et al. Exon 3 mutations of CTNNB1 drive tumorigenesis: a review. Oncotarget 9, 5492–5508, https://doi.org/10.18632/oncotarget.23695 (2018).
Rubinfeld, B. et al. Stabilization of beta-catenin by genetic defects in melanoma cell lines. Science 275, 1790–1792, https://doi.org/10.1126/science.275.5307.1790 (1997).
Bo, N., Wang, D., Wu, B., Chen, L. & Ruixue, M. Analysis of beta-catenin expression and exon 3 mutations in pediatric sporadic aggressive fibromatosis. Pediatr Dev Pathol 15, 173–178, https://doi.org/10.2350/10-07-0866-OA.1 (2012).
Wang, W. L. et al. CTNNB1 genotyping and APC screening in pediatric desmoid tumors: a proposed algorithm. Pediatr Dev Pathol 15, 361–367, https://doi.org/10.2350/11-07-1064-OA.1 (2012).
Li, M. M. et al. Standards and Guidelines for the Interpretation and Reporting of Sequence Variants in Cancer: A Joint Consensus Recommendation of the Association for Molecular Pathology, American Society of Clinical Oncology, and College of American Pathologists. J. Mol. Diagn. 19, 4–23, https://doi.org/10.1016/j.jmoldx.2016.10.002 (2017).
Joensuu, H. et al. Effect of the tyrosine kinase inhibitor STI571 in a patient with a metastatic gastrointestinal stromal tumor. The New England journal of medicine 344, 1052–1056, https://doi.org/10.1056/NEJM200104053441404 (2001).
Rubin, B. P. et al. KIT activation is a ubiquitous feature of gastrointestinal stromal tumors. Cancer Res. 61, 8118–8121 (2001).
Hirota, S. et al. Gain-of-function mutations of c-kit in human gastrointestinal stromal tumors. Science 279, 577–580, https://doi.org/10.1126/science.279.5350.577 (1998).
Dufresne, A. et al. Impact of KIT exon 10 M541L allelic variant on the response to imatinib in aggressive fibromatosis: analysis of the desminib series by competitive allele specific Taqman PCR technology. BMC cancer 14, 632, https://doi.org/10.1186/1471-2407-14-632 (2014).
Grabellus, F. et al. The prevalence of the c-kit exon 10 variant, M541L, in aggressive fibromatosis does not differ from the general population. Journal of clinical pathology 64, 1021–1024, https://doi.org/10.1136/jcp.2011.090498 (2011).
Trautmann, M. et al. Phosphatidylinositol-3-kinase (PI3K)/Akt Signaling is Functionally Essential in Myxoid Liposarcoma. Molecular Cancer Therapeutics 18, 834–844, https://doi.org/10.1158/1535-7163.Mct-18-0763 (2019).
Rehkämper, J. et al. Diagnostic tools in the differential diagnosis of giant cell-rich lesions of bone at biopsy. Oncotarget 9, 30106 (2018).
Grunewald, I. et al. MDM2 and CDK4 amplifications are rare events in salivary duct carcinomas. Oncotarget 7, 75261–75272, https://doi.org/10.18632/oncotarget.12127 (2016).
Adzhubei, I. A. et al. A method and server for predicting damaging missense mutations. Nat. Methods 7, 248–249, https://doi.org/10.1038/nmeth0410-248 (2010).
Choi, Y., Sims, G. E., Murphy, S., Miller, J. R. & Chan, A. P. Predicting the functional effect of amino acid substitutions and indels. Plos One 7, e46688, https://doi.org/10.1371/journal.pone.0046688 (2012).
Kumar, P., Henikoff, S. & Ng, P. C. Predicting the effects of coding non-synonymous variants on protein function using the SIFT algorithm. Nature Protocols 4, 1073–1082, https://doi.org/10.1038/nprot.2009.86 (2009).
Ng, P. C. & Henikoff, S. Predicting deleterious amino acid substitutions. Genome Res. 11, 863–874, https://doi.org/10.1101/gr.176601 (2001).
Reva, B., Antipin, Y. & Sander, C. Determinants of protein function revealed by combinatorial entropy optimization. Genome Biol. 8, R232, https://doi.org/10.1186/gb-2007-8-11-r232 (2007).
Kircher, M. et al. A general framework for estimating the relative pathogenicity of human genetic variants. Nat. Genet. 46, 310–315, https://doi.org/10.1038/ng.2892 (2014).
Acknowledgements
This study was supported in part by grants from the Maria Möller Foundation to S.H.
Author information
Authors and Affiliations
Contributions
Conception and design: M.T., J.R., I.G. and S.H. Development of methodology, acquisition, analysis, and/or interpretation of data: M.T., J.R., H.G., J.B., E.W., W.H., I.G. and S.H. Contribution of administrative, experimental, analytic or material support: J.B., E.W. and W.H. Writing and/or revision of the manuscript: M.T., J.R., H.G. and S.H. Study supervision: M.T. and S.H. All authors reviewed and approved the manuscript for submission.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Trautmann, M., Rehkämper, J., Gevensleben, H. et al. Novel pathogenic alterations in pediatric and adult desmoid-type fibromatosis – A systematic analysis of 204 cases. Sci Rep 10, 3368 (2020). https://doi.org/10.1038/s41598-020-60237-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-60237-6
This article is cited by
-
Mesenchymale Tumoren im Gastrointestinaltrakt
Die Gastroenterologie (2024)
-
Aggressive gluteal fibromatosis: a rare case associated with spontaneous superior gluteal artery pseudoaneurysm
Skeletal Radiology (2023)
-
Clinical Prognostic Factors and Integrated Multi-Omics Studies Identify Potential Novel Therapeutic Targets for Pediatric Desmoid Tumor
Biological Procedures Online (2022)
-
Efficacy of auranofin as an inhibitor of desmoid progression
Scientific Reports (2022)
-
Recurrent CTNNB1 mutations in craniofacial osteomas
Modern Pathology (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
