
Abstract
Objectives were to evaluate the role of canonical WNT signaling in development of the preimplantation embryo. Signaling was activated with 2-Amino-4-(3,4-(methylenedioxy)benzylamino)-6-(3-methoxyphenyl)pyrimidine (AMBMP) and inhibited with Dickkopf-related protein 1 (DKK1). Treatment of bovine embryos with AMBMP at day 5 after insemination decreased development to the blastocyst stage at day 7 and reduced numbers of trophectoderm and inner cell mass cells. At high concentrations, AMBMP caused disorganization of the inner cell mass. DKK1 blocked actions of AMBMP but did not affect development in the absence of AMBMP. Examination of gene expression in day 6 morulae by microarray revealed expression of 16 WNT genes and other genes involved in WNT signaling; differences in relative expression were confirmed by PCR for 7 genes. In conclusion, the preimplantation embryo possesses a functional WNT signaling system and activation of the canonical pathway can inhibit embryonic development.
Similar content being viewed by others

Introduction
The WNT signaling system plays important roles in directing developmental processes including maintenance of pluripotency1,2, cell migration during gastrulation and neurulation3 and axis formation4,5. WNT proteins regulate cell function through a variety of cell signaling systems. The canonical pathway involves binding of WNT proteins to a membrane receptor, Frizzled (FZD) and co-receptor, LDL receptor-related protein (LRP) 5 and 6. This signal activates the downstream messenger Dishevelled that blocks action of the β-catenin destruction complex, resulting in accumulation of β-catenin in the cytoplasm and translocation to the nucleus, where it activates transcription factors such as T-cell factor and lymphocyte enhancer factor6,7,8. Non-canonical signaling pathways are activated by binding of WNTs to various receptors including FZD, ROR2 and RYK9,10.
The canonical pathway has been associated with regulation of cell fate while non-canonical pathways have been implicated in regulation of polarity, asymmetric cell divisions and cell movements during gastrulation11. While individual WNT proteins are often classified as to whether they activate canonical or non-canonical pathways, activation of a specific pathway is dependent not only on the ligand but also receptor availability, cell type, specific stage of embryonic development and agonistic or antagonistic effects of other WNT proteins8,10,11.
Little is known about the role of WNT signaling in embryonic development during the preimplantation period. Evidence from the mouse indicates that the Wnt system is present and activated as early as the two-cell stage12,13,14,15. However, inhibition of Wnt signaling does not compromise development to the blastocyst stage14 and, therefore, activation of this signaling system may not be a requirement for preimplantation development. Activation of WNT signaling in bovine embryos by inhibitors of glycogen synthase kinase 3 (GSK3) had inconsistent effects on development to the blastocyst stage. One inhibitor, LiCl2, blocked development to the blastocyst stage while another, CT99021, increased the percent of zygotes that developed to the blastocyst stage16.
Here we tested the hypothesis that activation of WNT signaling during early embryonic development inhibits development of embryos to the blastocyst stage. The basis for the hypothesis was the observation that colony stimulating factor-2 (CSF2), which alters gene expression in a way that would inhibit WNT signaling17, can enhance the competence of embryos to develop to the blastocyst stage and to establish pregnancy when transferred into females18. WNT signaling was activated by administration of 2-Amino-4-(3,4-(methylenedioxy)benzylamino)-6-(3-methoxyphenyl)pyrimidine (AMBMP), a WNT agonist that activates canonical signaling19,20. The timing of agonist administration, day 5 after insemination, represents a period in development after the embryo has undergone major genome activation21 and is at the morula stage of development. Moreover, it is identical to the time of development when CSF2 acts to enhance embryonic development. To determine whether actions of AMBMP were mediated by a canonical WNT pathway, it was tested whether effects of AMBMP could be blocked by Dickkopf-related protein 1 (DKK1), which interferes with binding of WNTs to the co-receptor LRP 5/622,23. Administration of DKK1 was also used to assess the effects of signaling of endogenous WNTs through LRP 5/6 on early embryonic development. Finally, the capacity of the preimplantation embryo to express genes involved in WNT signaling was evaluated by querying a microarray database generated using mRNA from the bovine morula for expression of genes involved in WNT signaling.
Results
Effect of AMBMP on development to the blastocyst stage
The first experiment was designed to test if activation of canonical WNT signaling blocked development of embryos to the blastocyst stage. Addition of AMBMP to culture medium at day 5 after insemination caused a concentration-dependent decrease (P < 0.001) in the proportion of oocytes that reached the blastocyst stage at day 7 after insemination (Table 1). The minimum concentration necessary to obtain a significant inhibitory effect was 0.7 μM AMBMP.
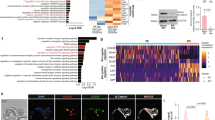
AMBMP reduced significantly total cell number at concentrations of 0.7 μM and higher (Table 1). Similarly, AMBMP reduced significantly numbers of trophectoderm (TE) cells at 1.4 μM and higher and numbers of ICM cells at 0.7 and 2.8 μM (Table 1). The ratio of ICM/TE cells was not significantly affected (P = 0.33) by AMBMP but the ratios were numerically highest at the two highest concentrations of AMBMP. At the highest concentrations of AMBMP, the ICM appeared disorganized with several cells not labeled with CDX2 (i.e. ICM) being located outside the cluster of cells characterized by the ICM (Figure 1).

Representative images of labeling of inner cell mass and trophectoderm cells.
Blastocysts were harvested at day 7 and subjected to differential immunofluorescence. Blue: Hoescht 33342 staining (all nuclei); green: CDX2+ cells (labeled with mouse anti-CDX2 and FITC-conjugated anti mouse IgG). Trophectoderm cells were labeled with Hoescht and anti CDX2 while inner cell mass cells were labeled with Hoescht only.
Inhibition of actions of AMBMP by CSF2
An experiment was conducted to determine whether AMBMP would inhibit embryonic development when embryos were exposed to CSF2, which has been reported to alter genes involved in WNT signaling. Embryos cultured with CSF2 (10 ng/ml) beginning at Day 5 were exposed to AMBMP (0.7 μM) at Day 6. Despite the preesence of CSF2, AMBMP decreased (P = 0.04) blastocyst development at day 7 (percent of embryos becoming blastocysts = 41.5 ± 3.7% vs 29.9% ± 3.7%, for embryos treated with vehicle and AMBMP, respectively).
Inhibition of actions of AMBMP by the WNT antagonist DKK1
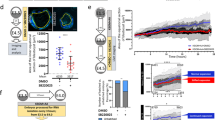
A third experiment was designed to test whether AMBMP blocked development by acting through the WNT signaling pathway. The approach was to determine whether DKK1, which prevents binding of the co-receptor LRP 5/6 to the agonist-receptor complex, blocks effects of AMBMP on development. As shown in Figure 2, AMBMP decreased (P = 0.02) the proportion of embryos that developed to the blastocyst stage at day 7. Moreover, blastocyst development was affected by the interaction between AMBMP and DKK1 at day 7 (P = 0.01). This interaction occurred as a result of blockage of the negative effect of AMBMP on blastocyst development by DKK1 (Figure 2); combination of AMBMP and DKK1 resulted in 22.2 ± 2.1% of embryos developing to blastocysts at day 7 compared to 15.9 ± 2.1% for AMBMP alone. In the absence of AMBMP, however, DKK1 tended to reduce blastocyst development at day 7 (29.9 versus 21.1 ± 2.1% for control and 100 ng/ml of DKK1, respectively).

Co-treatment of bovine embryos at day 5 after insemination with 100 ng/ml Dickkopf-related protein 1 (DKK1) reduces the inhibitory effect of a WNT agonist (AMBMP) on development to the blastocyst stage.
Data are least-squares means ± SEM of results from 5 replicates. The percent of oocytes becoming blastocysts at day 7 was affected by agonist (P = 0.02) and the agonist by DKK1 interaction (P = 0.01).
Effect of DKK1 on blastocyst development
Because there was a tendency of DKK1 to decrease development, an additional experiment was performed to determine whether DKK1 itself has deleterious effects on embryo competence to develop to the blastocyst stage. Embryos were cultured with concentrations of DKK1 ranging from 0 to 400 ng/ml, beginning at day 5 after insemination. As shown in Figure 3, addition of DKK1 at concentrations of 50, 100, 200 and 400 ng/ml did not change (P = 0.47) the proportion of embryos reaching the blastocyst stage at day 7 of development as compared to embryos cultured without DKK1.

Effect of administration of different concentrations of Dickkopf related protein 1 (DKK1) at day 5 after insemination on blastocyst development at day 7.
Data are least-squares means ± SEM of results from 7 replicates. No difference in the percent of ooyctes that became blastocysts at day 7 after insemination was observed following treatment with DKK1 at concentrations between 50 and 400 ng/ml (P = 0.47).
Expression of genes involved in WNT signaling
Examination of gene expression in the day 6 embryo using microarray analysis (Table 2) revealed that the embryo expresses at least 16 WNT genes at this stage, with greatest hybridization signal for WNT1, WNT2B, WNT11, WNT8A and WNT10B. Moreover, genes for WNT receptors were expressed including 11 FZD genes, the co-receptors LRP5 and LRP6 and non-canonical receptors RYK and ROR2. Genes encoding signal transduction molecules for both canonical and non-canonical pathways were expressed, such as CTNNBL1 (gene encoding β-catenin), as were a variety of genes encoding for proteins that modulate WNT signaling (such as DKK1).
Analysis of a separate set of embryos at day 6 of development by qPCR confirmed results of the microarray. A total of 7 genes were analyzed and all but one (WNT3) were detectable by qPCR (Figure 4A). Moreover, the amount of mRNA, expressed as the fold-change relative to WNT11 (the least-abundant detectable gene) was highly correlated (r = 0.83, P = 0.02) with amount of mRNA detected by microarray analysis (Figure 4B).

Genes related to WNT signaling are expressed in day 6 morulae.
(A) Agarose gel electrophoretogram of reverse transcriptase PCR products for CTNNBL1, DKK1, FZD7, LRP6, RYK, WNT11, SDHA and GAPDH. Note that another gene, WNT3, was not detectable. (B) Correlation between expression results for day 6 morulae obtained from microarray analysis and from real-time PCR. WNT3 was undetectable and the amounts of other mRNA were expressed as a fold-change relative to amount of WNT11.
Discussion
Using the bovine embryo as a model, we demonstrate here that WNT signaling affects embryonic development to the blastocyst stage. Activation of canonical WNT signaling at day 5 of development decreased the number of embryos reaching the blastocyst stage. This conclusion is based on the observation that AMBMP decreased the proportion of embryos that developed to the blastocyst stage in a concentration-dependent manner. Likewise, this action of AMBMP was blocked by administration of DKK1 to interfere with FZD-LRP 5/6 receptor signaling.
The mechanism by which AMBMP inhibits development is not known. One possibility is that AMBMP blocked proliferation. Such an explanation is consistent with the reduction in blastocyst cell number caused by AMBMP. While canonical WNT signaling is often associated with an increase in cell proliferation24,25,26, inhibition of canonical WNT signaling by DKK1 can induce cell proliferation in some cells such as endothelial colony-forming cells27.
In addition to its role as the intra-cellular effector of canonical WNT signaling, β-catenin is also a main component of adherens junctions28, linking the adhesive glycoprotein E-cadherin to actin filaments29. Knockout of E-cadherin is lethal to mice embryos, as it impairs formation of the blastocoele and subsequent hatching of the embryo30. Perhaps, over-activation of WNT signaling through AMBMP results in recruitment of adherens junctions associated β-catenin to the nucleus, decreasing E-cadherin mediated cell-cell adhesion and disrupting formation of the blastocoele.
It is also possible that the decreased blastocyst formation caused by AMBMP is the result of maintenance of pluripotency and inhibition of blastomere differentiation. There is abundant evidence that canonical WNT signaling can promote pluripotency2,31,32,33,34. Consistent with this idea is that there was a tendency for WNT to affect numbers of TE cells to a greater extent than numbers of ICM cells.
Given the inhibitory effect of activation of the canonical pathway achieved by AMBMP and the expression of canonical WNTs by the day 6 embryo, it was surprising that inhibition of endogenously produced canonical WNTs by addition of DKK1 did not increase the percent of embryos that developed to the blastocyst stage. DKK1 inhibits the canonical WNT pathway through binding and inactivation of the LRP 5/6 WNT co-receptors22,35,36. However, DKK1 has more recently been associated with inhibition of non-canonical pathways in human cancer cell lines37 and with inhibition of both canonical and non-canonical pathways altering axis formation in the Xenopus embryo38. Thus, DKK1 could conceivably affect activation of canonical and non-canonical pathways activated by endogenously produced WNTs. Also, actions of one WNT can depend on the presence of another8,39 so global inhibition of WNT signaling might cause actions different than actions of individual WNTs. In the mouse, inhibition of endogenous canonical WNT signaling with either recombinant DKK1 or PKF115-584 and CGP049090, two small chemical molecules that inhibit the β-catenin/TCF complex, had no effect on development to the blastocyst stage14. Inhibition did, however, compromise development through implantation. Conducting experiments to determine long-term consequences of regulation of WNT signaling during the preimplantation period in cattle will provide additional insight into the role of WNTs in the preimplantation period.
Present results demonstrate that the machinery for WNT signaling is present in the preimplantation embryo. The microarray data demonstrate that genes for canonical and non-canonical WNTs, non-WNT ligands such as Norrie disease protein and R-spondin, FZD and alternative WNT receptors such as RYK and ROR2 and a great variety of molecules involved in WNT signaling regulation are present in Day 6 bovine morulae. This finding was confirmed by qPCR using a subset of genes and different pools of day 6 morulae. Norrie disease protein, also called Norrin40 and R-spondin have been identified as alternative FZD ligands that are capable of activating canonical WNT signaling41. Likewise, ROR and RYK are transmembrane tyrosine kinases that have been recently identified as alternative (non-FZD) WNT receptors41. There is additional evidence in the literature indicating the presence of WNT signaling systems in the early embryo. Genes involved in WNT signaling pathways are expressed in both bovine oocytes and blastocysts42. Aparicio and coworkers16 demonstrated the presence of GSK3 protein, a key component of the β-catenin destruction complex, in bovine embryos from the 2-cell to the blastocyst stage. A variety of Wnts are expressed in murine blastocysts, with Wnt3a, Wnt6, Wnt 7b, Wnt 9a and Wnt 10b showing the highest expression levels13 and Wnts are expressed as early as the 2-cell stage12. Kemp and collaborators13 also reported the expression of Wnt antagonists such as Sfrps and Dkks, with greatest detection of Dkk1, in murine blastocysts.
It is likely that maternal regulation of preimplantation embryonic development involves regulation of embryonic WNT signaling. One such maternal signal might be CSF2, which improves ability of bovine embryos to develop to the blastocyst stage and to establish pregnancy after transfer into recipients18. One action of CSF2, when administered at the morula stage, is alteration of gene expression in a manner that was interpreted as causing inhibition of WNT signaling17. For example, CSF2 decreased the non-canonical WNT16 and increased expression of genes such as secreted frizzled-related protein 4 (SFRP4) that inhibit canonical WNT signals17. Our present findings indicate, however, that AMBMP inhibits development in the presence of CSF2. This result is suggestive that the positive action of CSF2 on embryonic survival is not mediated by inhibition of canonical WNT signaling.
In conclusion, our findings demonstrate that WNT signaling is present in pre-implantation embryos and plays a role in the regulation of development to the blastocyst stage. Activation of canonical signaling decreases development and blastocyst cell numbers; however, inhibition of WNT signaling by DKK1 is not beneficial to blastocyst development. The exact role for regulation of preimplantation development by WNTs is not yet clear. It has been postulated that the roles of different WNTs are variable according to developmental stage and factors such as cell environment, presence and abundance of different receptors and effector molecules8. It is very likely that regulation of embryonic development by WNTs involves a plethora of endogenous WNTs, receptors and signaling molecules, as well as a fine regulation of canonical and non-canonical pathways to balance maintenance of pluripotency, self-renewal and cell-fate commitment.
Methods
Reagents
Calbiochem (San Diego, CA, USA) was the source of AMBMP. The molecule was diluted in dimethylsulfoxide (DMSO) to a concentration of 2.8 mM. Aliquots of 100 μl-volume were stored at −20°C and protected from light until used. Human recombinant DKK1 was from R&D Systems (Minneapolis, MN). It was reconstituted using Dulbecco's phosphate buffered saline (DPBS) with 0.1% (w/v) bovine serum albumin (BSA) to a concentration of 100 μg/ml. Aliquots of 5 μl were stored at −20°C until use. Recombinant bovine CSF2 was donated by Novartis (Basel, Switzerland). Mouse monoclonal antibody (ascites) against CDX2 was purchased from Biogenex (Fremont, CA, USA) and affinity-purified goat anti-mouse IgG coupled to fluorescein (FITC) was from Abcam (Cambridge, MA, USA).
Production of embryos
Bovine embryos were produced by in vitro fertilization from oocytes obtained from abattoir-collected ovaries. All procedures for embryo production followed techniques previously described43.
Effect of AMBMP on development to the blastocyst stage
Following fertilization, putative zygotes (a mix of fertilized and unfertilized oocytes) were randomly placed in groups of 30 in 45 μl-microdrops of culture medium (SOF-BE1)43 covered with mineral oil. On day 5 after fertilization, AMBMP was added to each drop in a volume of 5 μl to achieve a final concentration of 0, 0.35, 0.7, 1.4 or 2.8 μM AMBMP and 0.1% (v/v) DMSO. Development to blastocyst stage was evaluated at day 7 and blastocysts were harvested and processed for determination of total cell number and number of ICM and TE cells. The experiment was conducted in 6 replicates using a total of 1,102 embryos.
Blastocyst cell numbers were determined as follows. All steps were conducted at room temperature unless otherwise described. Blastocysts were harvested from culture drops, washed three times in DPBS with 0.2% (w/v) polyvinylpyrrolidone (DPBS-PVP) and fixed in 4% (w/v) paraformaldehyde in DPBS-PVP for 15 min. Embryos were washed again three times in DPBS–PVP and permeabilized with 0.25% (v/v) Triton X-100 in DPBS for 20 min. After washing 3 times and incubation with a blocking buffer of 5% (w/v) BSA in DPBS (w/v) for 1 h, embryos were incubated with anti-CDX2 at 4°C overnight (used at the working concentration provided by the manufacturer). Embryos were washed with antibody buffer [10 mM Tris, pH 7.5 containing 0.15 M NaCl, 0.1% bovine serum albumin and 0.1% (v/v) Tween-20] and incubated with 1 μg/ml FITC-labeled goat anti-mouse IgG diluted in antibody buffer for 1 h at room temperature in the dark. Embryos were then incubated with 5 μg/ml Hoechst 33342 in DPBS-PVP for 15 min in the dark. Embryos were washed three times with DPBS–PVP, placed on clean microscope slides with Prolong® anti-fade solution (Invitrogen, Carlsbad, CA, USA), covered with a cover slip and examined for fluorescence using a Zeiss Axioplan 2 epifluorescence microscope (Zeiss, Gottingen, Germany) with a 40× objective and using Zeiss filter set 02 (DAPI filter) and Zeiss filter set 03 (FITC filter). Digital images were acquired using AxioVision software (Zeiss) and a high-resolution black and white Zeiss AxioCam MRm digital camera. Total cell number was determined by counting nuclei staining with Hoescht 33342, number of TE cells determined by counting nuclei positive for CDX2 (FITC) and number of ICM cells determined by subtracting number of TE cells from total cell number.
Inhibition of actions of AMBMP by CSF2
Following fertilization, putative zygotes were randomly allocated in groups of 30 in 40 μl-culture drops. At Day 5 after fertilization, embryos received 5 μl of CSF2 in culture medium to a final concentration of 10 ng/ml. At day 6, an additional 5 μl of medium was added containing either vehicle or AMBMP (final concentration = 0.7 μM). Blastocyst development was evaluated and recorded at Day 7 after insemination. This experiment was performed in five replicates using a total of 394 embryos.
Inhibition of actions of AMBMP by the WNT antagonist DKK1
Following fertilization, putative zygotes were randomly allocated in groups of 30 in 40 μl-culture drops. At day 5 after fertilization, AMBMP (diluted as in the previous experiment) or vehicle was added in a volume of 5 μl. In addition, DKK1, diluted in SOF-BE1 was added in a volume of 5 μl. The final concentration of DKK1 was 100 ng/ml. Drops of embryos were treated with vehicle only [0.1% (v/v) DMSO in SOF – control group], 0.70 μM of AMBMP, 100 ng/ml of DKK1, or 0.70 μM of AMBMP plus 100 ng/ml DKK1 (final concentrations). Development to the blastocyst stage was evaluated at day 7 after insemination. The experiment was performed in 5 replicates using a total of 1,487 embryos.
Effect of DKK1 on blastocyst development
Following fertilization, putative zygotes were randomly allocated in groups of 30 in 45 μl-culture drops. Dilution of DKK1 was performed as described in the previous experiment. At day 5 after fertilization, drops were treated with vehicle only [SOF-BE1 – control group] or 50, 100, 200 or 400 ng/ml of DKK1 (final concentrations). Development to the blastocyst stage was evaluated at day 7. The experiment was replicated 7 times using a total of 1,467 embryos.
Expression of genes involved in WNT signaling
A previously reported microarray database obtained by analysis of four separate pools of mRNA from bovine embryos at Day 6 of development with the Bos taurus Two Color Microarray Chip (Agilent Technologies, Santa Clara, CA, USA)17 was examined for expression of WNT genes as well as genes involved in WNT signaling. The relative abundance of each gene was determined by the intensity of the hybridization signal. Expression of four genes used as housekeeping genes in the bovine embryo (GAPDH, H2A, LMNB1, and ACTB)45 was also assessed to compare intensity of hybridization with that of genes involved in WNT signaling.
Real-time quantitative polymerase chain reaction (qPCR) of WNT related genes
Of the genes shown by microarray to be expressed in day 6 embryos, seven that encode main components of the WNT pathway were chosen for qPCR: WNT3, WNT11, FZD7, DKK1, LRP6, RYK and CTNNBL1. GAPDH and SDHA were used as housekeeping genes.
Primers were designed and obtained from Integrated DNA Technologies (Coralville, Iowa, USA), except for the housekeeping genes SDHA44 and GAPDH45, which were previously published. Primer sequences are listed in Table 3. A standard curve was performed for each primer using bovine day 7 blastocysts. PCR amplicons were sequenced and the obtained products were mapped to the Bos taurus genome using the Basic Local Alignment Search Tool of NCBI. All amplicon sequences aligned to the genes for which primers were designed with identities between 93 and 99%. For qPCR of day 6 morulae, 2 pools of 40 embryos were used. RNA was isolated using PicoPure RNA Isolation Kit (Applied Biosystems, Foster City, CA, USA) following the manufacturer's instructions. RNA concentration and quality were assessed using NanoDrop (Thermo Fisher Scientific, Wilmington, DE, USA). DNase treatment was performed using Qiagen DNase reagents (Qiagen, Valencia, CA) and reverse transcription using High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA). qPCR reaction was performed using SsoFast EvaGreen Supermix reagent (Bio-Rad, Hercules, CA, USA). The Bio-Rad thermal cycler CFX96-Real-Time system (Bio-Rad, Hercules, CA, USA) was used to quantify abundance of gene transcripts. The following PCR protocol was used: 30 seconds at 95°C followed by 40 cycles each of 5 seconds at 95°C and 1 minute at 60°C. All samples were amplified in duplicate and mean cycle threshold (Ct value) was used to compare transcript fold change. Fold changes were calculated relative to the Ct value for WNT11.
Statistical analysis
Analyses were performed with SAS for Windows, version 9.2 (SAS Institute Inc., Cary, NC). Data were tested for normality of distribution using the Univariate procedure. The General Linear Model procedure was used to perform least-squares analysis of variance to determine treatment effects and interactions. Replicate was considered random and other main effects were considered fixed. Tests of significance were made using the appropriate error terms after calculating expected means squares. The pdiff procedure of SAS was used as a means-separation test for multiple degree-degree-of-freedom effects. Results are presented as least-squares means ± standard error of the mean. The level of significance for all analyses was 0.05.
References
Kielman, M. F., Rindapaa, M., Gaspar, C., van Poppel, N., Breukel, C., avn Leeuwen, S., Taketo, M. M., Roberts, S., Smits, R. & Fodde, R. Apc modulates embryonic stem-cell differentiation by controlling the dosage of β-catenin signaling. Nature Genet. 32, 594–605 (2002).
Sato, N., Meijer, L., Skaltsounis, L., Greengard, P. & Brivanlou, A. H. Maintenance of pluripotency in human and mouse embryonic stem cells through activation of Wnt signaling by a pharmacological GSK-3-specific inhibitor. Nature Med. 10, 55–63 (2004).
Tada, M. & Smith, J. C. Wnt11 is a target of Xenopus Brachyury: regulation of gastrulation movements via Dishevelled, but not through the canonical Wnt pathway. Development 127, 2227–2238 (2000).
Tada, M., Concha, M. L. & Heisenberg, C. P. Non-canonical Wnt signaling and regulation of gastrulation movements. Cell Dev. Biol. 13, 251–260 (2002).
Tao, Q., Yokota, C., Puck, H., Kofron, M., Birsoy, B., Yan, D., Asashima, M., Wylie, C. C., Lin, X. & Heasman, J. Maternal Wnt11 activates the canonical Wnt signaling pathway required for axis formation in Xenopus embryos. Cell 120, 857–871 (2005).
Reya, T. & Clevers, H. Wnt signaling in stem cells and cancer. Nature 434, 843–850 (2005).
Huang, H. & He, X. Wnt/β-catenin signaling: new (and old) players and new insights. Curr. Opin. Cell Biol. 20, 119–125 (2008).
Amerongen, R. & Nusse, R. Towards an integrated view of Wnt signaling in development. Development 136, 3205 – 3214 (2009).
Schambony, A. & Wedlich, D. Wnt-5A/Ror2 regulate expression of XPAPC through an alternative noncanonical signaling pathway. Dev. Cell 12, 779–792 (2007).
De, A. Wnt/Ca2+ signaling pathway: a brief overview. Acta Biochim. Biophys. Sin. 43, 745–756 (2011).
Chien, A. J., Conrad, W. H. & Moon, R. T. A Wnt survival guide: from flies to human disease. J. Invest. Dermatol. 129, 1614–1627 (2009).
Lloyd, S., Fleming, T. P. & Collins, J. E. Expression of Wnt genes during mouse preimplantation development. Gene Expr. Patterns 3, 309–312 (2003).
Kemp, C., Willems, E., Abdo, S., Lambiv, L. & Leyns, L. Expression of all Wnt genes and their secreted antagonists during mouse blastocyst and postimplantation development. Dev. Dyn. 233, 1064–1075 (2005).
Xie, H., Tranguch, S., Jia, X., Zhang, H., Das, S. K., Dey, S. K., Kuo, C. J. & Wang, H. Inactivation of nuclear Wnt-β-catenin signaling limits blastocyst competency for implantation. Development 135, 717–727 (2008).
Chen, Q., Zhang, Y., Lu, J., Wang, Q., Wang, S., Cao, W., Wang, H. & Duan, E. Embryo-uterine cross-talk during implantation: the role of Wnt signaling. Mol. Hum. Reprod. 15, 215–221 (2009).
Aparicio, I. M., Garcia-Herreros, M., Fair, T. & Lonergan, P. Identification and regulation of glycogen synthase kynase-3 during bovine embryo development. Reproduction 140, 83–92 (2010).
Loureiro, B., Oliveira, L. J., Favoreto, M. G. & Hansen, P. J. Colony-stimulating factor 2 inhibits induction of apoptosis in the bovine preimplantation embryo. Am. J. Reprod. Immunol. 65, 578–588 (2011).
Loureiro, B., Bonilla, L., Block, J., Fear, J. M., Bonilla, A. Q. S. & Hansen, P. J. Colony-stimulating factor 2 (CSF-2) improves development and post-transfer survival of bovine embryos produced in vitro. Endocrinology 150, 5046–5054 (2009).
Liu, J., Wu, X., Mitchell, B., Kintner, C., Ding, S. & Schultz, P. G. A small-molecule agonist of the Wnt signaling pathway. Angew. Chem. Int. Ed. 44, 1987–1990 (2005).
Lim, J. C., Kania, K. D., Wijesuriya, H., Chawla, S., Sethi, J. K., Pulasky, L., Romero, I. A., Couraud, P. O., Weksler, B. B., Hladky, S. B. & Barrand, M. A. Activation of β-catenin signaling by GSK-3 inhibition increases p-glycoprotein expression in brain endothelial cells. J. Neurochem. 106, 1855–11865 (2008).
Memili, E. & First, N. L. Zygotic and embryonic gene expression in cow: a review of timing and mechanisms of early gene expression as compared with other species. Zygote 8, 87 – 96 (2000).
Bafico, A., Liu, G., Yaniv, A., Gazit, A. & Aaronson, S. A. Novel mechanisms of Wnt signaling inhibition mediated by Dickkopf-1 interaction with LRP6/Arrow. Nature Cell Biol. 3, 686–686 (2001).
Nusse, R. Making head or tail of Dickkopf. Nature 411, 255–256 (2001).
Tsukamoto, A. S., Grosschedl, R., Guzman, R. C., Parslow, T. & Varmus, H. E. Expression of the int-1 gene in transgenic mice is associated with mammary gland hyperplasia and adenocarcinomas in male and female mice. Cell 55, 619 – 625 (1988).
Reya, T., Duncan, A. W., Ailles, L., Domen, J., Scherer, D. C., Willert, K., Hintz, L., Nusse, R. & Weissman, I. L. A role for Wnt signalling in self-renewal of haematopoietic stem cells. Nature 423, 409–414 (2003).
Wang, H., Charles, P. C., Wu, Y., Ren, R., Pi, X., Moser, M., Barshishat-Kupper, M., Rubin, J. S., Perou, C., Bautch, V. & Patterson, C. Gene expression profile signatures indicate a role for Wnt signaling in endothelial commitment from embryonic stem cells. Circ. Res. 98, 1331–1339 (2006).
Smadja, D. M., d'Audigier, C., Weiswald, L. B., Badoual, C., Dangles-Marie, V., Mauge, L., Evrard, S., Laurendeau, I., Lallemand, F., Germain, S., Grelac, F., Dizier, B., Vidaud, M., Bieche, I. & Gaussen, P. The Wnt antagonist Dickkopf-1 increases endothelial progenitor cell angiogenic potential. Arterioscler. Thromb. Vasc. Biol. 30, 2544–2552 (2010).
Kohan-Ghadr, H. R., Smith, L. C., Arnold, D. R., Murphy, B. D. & Lefebvre, R. C. Aberrant expression of E-cadherin and β-catenin proteins in placenta of bovine embryos derived from somatic cell nuclear transfer. Rep. Fertil. Dev. 24, 588–598 (2011).
Kemler, R. From cadherins to catenins: cytoplasmic protein interactions and regulation of cell adhesion. Trends Genet. 9, 317–321 (1993).
Larue, L., Ohsugi, M., Hirchenhain, J. & Kemler, R. E-dacherin null mutant embryos fail to form a trophectoderm epithelium. Proc. Natl. Acad. Sci. USA 91, 8263–8267 (1994).
Cole, M. F., Johnstone, S. E., Newman, J. J., Kagey, M. H. & Young, R. A. Tcf3 is an integral component of the core regulatory circuitry of embryonic stem cells. Genes Dev. 22, 746 – 755 (2008).
Wend, P., Holland, J. D., Ziebold, U. & Birchmeiter, W. Wnt signaling in stem and cancer stem cells. Semin. Cell Dev. Biol. 21, 855 – 863 (2010).
Kelly, K. F., Ng, D. Y., Jayakumaran, G., Wood, G. A., Koide, H. & Doble, B. W. β-catenin enhances Oct-4 activity and reinforces pluripotency through a TCF-independent mechanism. Cell Stem Cell 8, 214–227 (2011).
Sokol, S. Y. Maintaining embryonic stem cell pluripotency with Wnt signaling. Development 138, 4341 – 4350 (2011).
Semenov, M. V., Tamai, K., Brott, B. K., Kuhl, M., Sokol, S. & He, X. Head inducer Dickkopf-1 is a ligand for Wnt co-receptor LRP6. Curr. Biol. 11, 951–961 (2001).
Zorn, A. M. Wnt signaling: Antagonistic Dickkopfs. Curr. Biol. 11, R592–R595 (2001).
Lee, A. Y., He, B., You, L., Xu, Z., Mazieres, J., Reguart, N., Mikami, I., Batra, S. & Jablons, D. M. Dickkopf-1 antagonizes Wnt signaling independent of β-catenin in human mesothelioma. Biochem. Biophys. Res. Commun. 323, 1246–1250 (2004).
Cha, S.-W., Tadjuidje, E., Tao, Q., Wylie, C. & Heasman, J. Wnt5a and Wnt11 interact in a maternal Dkk-1-regulated fashion to activate both canonical and non-canonical signaling in Xenopus axis formation. Development 135, 3719–3729 (2008).
Maye, P., Zheng, J., Li, L. & Wu, D. Multiple mechanisms for Wnt11-mediated repression of the canonical Wnt signaling pathway. J. Biol. Chem. 279, 24659 – 24665 (2004).
Xu, S., Cheng, F., Liang, J., Wu, W. & Zhang, J. Manternal xNorrin, a canonical WNT signaling agonist and TGF-β antagonist, controls early neuroectoderm specification in Xenopus. PLoS Biol. 10, e1001286 (2012).
Hendrickx, M. & Leyns, L. Non-conventional Frizzled ligands and Wnt receptors. Dev. Growth Differ. 50, 229–243 (2008).
Adjaye, J., Herwig, R., Brink, T. C., Herrman, D., Greber, B., Sudheer, S., Groth, D., Carnwath, J. W., Lehrach, H. & Niemann, H. Conserved molecular portraits of bovine and human blastocysts as a consequence of the transition from maternal to embryonic control of gene expression. Physiol. Genomics 31, 315–327 (2007).
Fields, S. D., Hansen, P. J. & Ealy, A. D. Fibroblast growth factor requirements for in vitro development of bovine embryos. Theriogenology 75, 1466–1475 (2011).
Goossens, K., Van Poucke, M., Van Soom, A., Vandesompele, J., Van Zeveren, A. & Peelman, L. J. Selection of reference genes for quantitative real-time PCR in bovine preimplantation embryos. BMC Dev Biol 5, 27 (2005).
Robert, C., McGraw, S., Massicotte, L., Pravetoni, M., Gandolfi, F. & Sirard, M. A. Quantification of housekeeping transcript levels during the development of bovine preimplantation embryos. Biol Reprod 67,1465–1472 (2002).
Acknowledgements
This project was supported by Agriculture and Food Research Initiative Competitive Grants 2009-65203-05732 and 2011-67015-30688 from the USDA National Institute of Food and Agriculture. The authors thank Central Beef Packing Co. (Center Hill, FL, USA) for provision of ovaries, William Rembert for collecting and transporting ovaries, James Moss and Luciano Bonilla for assistance with embryo production, Manabu Ozawa for development of the CDX2 labeling protocol and Novartis for donation of CSF2.
Author information
Authors and Affiliations
Contributions
All authors were involved in experimental design, data analysis and interpretation and in writing the paper. AD, KD, KM and BL conducted the experiments.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article
Cite this article
Denicol, A., Dobbs, K., McLean, K. et al. Canonical WNT signaling regulates development of bovine embryos to the blastocyst stage. Sci Rep 3, 1266 (2013). https://doi.org/10.1038/srep01266
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep01266
This article is cited by
-
Extracellular vesicles-coupled miRNAs from oviduct and uterus modulate signaling pathways related to lipid metabolism and bovine early embryo development
Journal of Animal Science and Biotechnology (2024)
-
MicroRNA-451 is downregulated in the follicular fluid of women with endometriosis and influences mouse and human embryonic potential
Reproductive Biology and Endocrinology (2019)
-
A dose-dependent response to MEK inhibition determines hypoblast fate in bovine embryos
BMC Developmental Biology (2019)
-
Importance of prostate androgen-regulated mucin-like protein 1 in development of the bovine blastocyst
BMC Developmental Biology (2019)
-
Dickkopf-related protein 1 is a progestomedin acting on the bovine embryo during the morula-to-blastocyst transition to program trophoblast elongation
Scientific Reports (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
