
Abstract
Carbon ion therapy is a promising evolving modality in radiotherapy to treat tumors that are radioresistant against photon treatments. As carbon ions are more effective in normal and tumor tissue, the relative biological effectiveness (RBE) has to be calculated by bio-mathematical models and has to be considered in the dose prescription. This review (i) introduces the concept of the RBE and its most important determinants, (ii) describes the physical and biological causes of the increased RBE for carbon ions, (iii) summarizes available RBE measurements in vitro and in vivo, and (iv) describes the concepts of the clinically applied RBE models (mixed beam model, local effect model, and microdosimetric-kinetic model), and (v) the way they are introduced into clinical application as well as (vi) their status of experimental and clinical validation, and finally (vii) summarizes the current status of the use of the RBE concept in carbon ion therapy and points out clinically relevant conclusions as well as open questions. The RBE concept has proven to be a valuable concept for dose prescription in carbon ion radiotherapy, however, different centers use different RBE models and therefore care has to be taken when transferring results from one center to another. Experimental studies significantly improve the understanding of the dependencies and limitations of RBE models in clinical application. For the future, further studies investigating quantitatively the differential effects between normal tissues and tumors are needed accompanied by clinical studies on effectiveness and toxicity.
Export citation and abstract BibTeX RIS

Original content from this work may be used under the terms of the Creative Commons Attribution 3.0 licence. Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
1. Introduction
1.1. Ion beam radiotherapy
In recent years, ion beam radiotherapy using protons as well as carbon ions has gained significant interest (Castro et al 1994, Schulz-Ertner and Tsujii 2007, Kamada 2012, Combs and Debus 2013, Kamada et al 2015). There are several underlying reasons for this development: Although modern treatment techniques with photons such as intensity modulated in combination with image-guided radiotherapy improved dose distributions to a large extent, clinical results are still unsatisfying for some tumors and dose escalation in the tumor while respecting the radiation tolerance of the surrounding normal tissues is still expected to improve cure rates. Monoenergetic ion beams, on the other hand, exhibit an 'inverted' depth dose profile (Bragg curve) together with a finite range in tissue (Suit et al 2010). Due to the Coulomb interaction, ions deposit only a small dose in the entrance region (plateau) of the Bragg curve, which increases towards a maximum (Bragg peak) at the end of the ion range. Beyond the Bragg peak the dose drops off quickly leading to a sparing of the respective normal issue behind the target volume. For clinical applications, several monoenergetic Bragg peaks are superimposed to form a so-called spread-out Bragg peak (SOBP), which covers the complete tumor with high doses. Although this superposition increases the plateau dose significantly, it remains below that of the tumor and applying several SOBPs from different directions increases the degree of conformity, mainly because of the very steep lateral and distal dose gradients of the individual SOBPs. These physical properties of ions allow for dose escalation in the tumor relative to photons while it still spares the surrounding normal tissue.
Ion beams have been applied since the 1950s using the so-called passive beam delivery techniques (Castro et al 1992, Kanai et al 1999, Torikoshi et al 2007), however, it was only in the mid-1990s when the innovative beam scanning technique was clinically introduced for protons (Pedroni et al 1999, Pedroni et al 2005) and carbon ions (Haberer et al 1993, Kraft 2000). This further increased the accuracy of ion beam radiotherapy and the scanning technique is now considered as state of the art. Worldwide, 62 proton and 11 carbon ion centers are in clinical operation and additional facilities are under construction. By the end of 2015, 131 240 patients have been treated with protons and 22 963 were irradiated with other heavy charged particles, 19 376 of them with carbon ions (www.ptcog.ch).
1.2. Biological effectiveness of ions relative to photons
As compared to photons, irradiations with ion beams are empirically found to be biologically more effective if the same absorbed dose is applied. Quantitatively, this feature is expressed by the relative biological effectiveness (RBE)

which is defined as the ratio of a photon dose ( and an ion dose
and an ion dose  leading to the same biological effect under otherwise identical conditions. It has to be pointed out that these 'identical conditions' especially include the same number of fractions (Fx) as otherwise effects from beam quality and fractionation would be mixed-up.
leading to the same biological effect under otherwise identical conditions. It has to be pointed out that these 'identical conditions' especially include the same number of fractions (Fx) as otherwise effects from beam quality and fractionation would be mixed-up.
To determine the RBE with equation (1), a biological endpoint has to be specified. Although in principle any biological endpoint may be used, it should be well-detectable and relevant for the investigated radiobiological or clinical question. For radiotherapy, sterilization of tumor cells is most important and therefore, the endpoint cell survival is commonly used for in vitro experiments. In this case, irradiations with photons and ions are considered to be isoeffective if the dose-dependent survival fractions measured in the clonogenic assay are the same (figure 1(a)). For tumors, related endpoints such as local tumor control (figure 1(b)) or growth delay (figures 1(c) and (d)) are used accordingly. In this case, two doses, leading to the same tumor control probability (TCP) in dose-response experiments (figure 1(b)) or to the same growth delay (figures 1(c) and (d)) are considered as isoeffective.

Figure 1. Schematic illustration of the most frequently used methods to determine the RBE: (a) cell survival curves in vitro fitted by the LQM. Irradiations with photons and carbon ions are considered as isoeffective if the survival fractions are the same. The dose dependence of the RBE results from the different shapes of the photon and carbon ion curve and leads to different RBEs at different survival levels. The different survival levels are considered as different endpoints. (b) In vivo, dose response curves for the endpoints NTCP or TCP may be adjusted to the observed incidence at different dose levels. In this case, irradiations with photons and carbon ions are considered as isoeffective if the probability for the selected endpoint is the same. Here, the dose dependence of the RBE is introduced by the underlying fractionation schedule. If the curves for photons and carbon ions have different slopes, an additional dose dependence results from the comparison of RBE values at different effect levels. It has to be noted, however, that this dose dependence originates from the comparison of different rather than identical endpoints. (c,d) In-vivo growth delay experiments are used only for tumors. Two irradiations are considered as isoeffective, if the growth delay is the same. To measure the growth delay, the relative volume increase has to be specified. As it is difficult to obtain exactly the same growth delay for both radiation types, the isoeffective dose has to be determined by interpolation (d). The dose dependence of the RBE is introduced by the dose level selected for the reference irradiation with photons. It has to be noted that while growth delay experiments are performed with subtherapeutic doses, therapeutic doses are necessary to generate a complete dose-response curve in a tumor control assay. This has to be considered, when comparing RBEs from these two types of experiments.
Download figure:
Standard image High-resolution imageAlthough cell survival is in principle also relevant for normal tissue, its significance related to the occurrence of side effects is less clear due to the hierarchical structure of normal tissue. While cell survival experiments may measure the intrinsic radiosensitivity of normal proliferating cells, the radiation tolerance of the tissue will strongly depend on the irradiated volume and the interaction of substructures (e.g. vascular and parenchymal cells). In addition, early and late effects show different characteristics and the influence of treatment parameters like dose, fractionation, and beam modality on tissue response may be different. Normal tissue reactions may also be quantified by dose-response experiments using morphological, functional, or clinical criteria for the definition of the biological endpoint. Two doses are considered as isoeffective if the normal tissue complication probability (NTCP) is the same (figure 1(b)).
1.3. Dose prescription in ion beam radiotherapy
For a given absorbed ion dose, the  allows the calculation of the isoeffective photon dose
allows the calculation of the isoeffective photon dose

This allows the estimation of the biological effect of an ion irradiation on the basis of the response to a photon reference beam quality, for which the response is known. Experimentally, often 250 kV x-rays or 60Co are used as a reference while high-energy x-ray beams (e.g. 6 MV) are clinically more important. To distinguish the absorbed ion dose (also termed as physical dose) from the isoeffective photon dose, the latter is often termed as biologically effective dose (or biological dose) in units of GyE (Gray equivalent) or CGE (cobalt Gray equivalent if 60Co is used as a reference). However, as GyE and CGE are not SI units and since the term 'isoeffective dose' is also used in the context of other radiobiological effects such as fractionation (Wambersie et al 2006), the term 'RBE-weighted dose' is now widely adopted in the literature and numerical values are expressed in Gy (RBE) (ICRU Report 78 2008). Here, the amendment '(RBE)' reminds one that the dose (still measured in units of Gy) refers to an isoeffective photon dose rather than an absorbed ion dose.
1.4. Determinants of the RBE
The dose prescription in terms of RBE-weighted rather than absorbed dose requires the prediction of the RBE in treatment planning, however, the RBE is a complex quantity, which depends on the physical parameters of the irradiation as well as on the biological parameters of the irradiated biological system. In the following, the most important parameters are briefly discussed.
1.4.1. Ion type
Although the impact of multiple Coulomb scattering is larger for protons than for carbon ions, both ion types allow for highly conformal tumor irradiations. In comparison to protons, however, the RBE of carbon ions is significantly increased and varies much more for different physical and biological parameters (see below). For this reason, more detailed RBE models are required for carbon ions (Suit et al 2010). In contrast, the RBE of protons varies only very little over the SOBP and although there is some discussion about an increased RBE at the distal edge of the SOBP (Paganetti 2014, 2015), a fixed RBE value of 1.1 is currently adopted over the whole SOBP for all proton treatments.
1.4.2. Beam quality
In radiobiological studies, beam quality is commonly characterized by the linear energy transfer (LET). As the RBE increases with LET up to a certain value, the RBE in the Bragg peak is higher than in the entrance region. This is the main rationale for the clinical application of carbon ions. As the LET is only uniquely defined in monoenergetic beams, the LET for a position in a SOBP refers to an average value over the energy spectrum of the particles. This average LET value is either stated as the fluence-averaged LET (also termed as track-averaged LET)

or as the dose-averaged LET

As outlined in Guan et al (2015) for protons, Sel is the unrestricted electronic stopping power of an ion with the kinetic energy E, Φ(E, z) is the fluence of the ions with the kinetic energy E at the location z and D(E, z) is the respective dose. In the case of carbon ions, the primary charged particles may produce secondary fragments and the contributions of the different ion types have to be added as indicated by the summation over the index i. It is important to note that averaging over fluence or dose will lead to different numerical values for the LET at the same position in the SOBP. When stating averaged LET values of experimental studies, it is therefore important to specify clearly to which definition these values refer. The exact numerical values of the averaged LET may also depend on the model used for the calculation of the stopping power and the particle spectrum. As the biological effect in cells or tissue is determined by the total effect of the dose contributions of the different LET components building-up the radiation field, the dose-averaged LET is normally used to specify the beam quality in radiobiological experiments and if not otherwise stated, LET values provided in this review refer to the dose-averaged LET.
Although the dose-averaged LET is a practical and frequently used quantity to specify the beam quality, it has to be noted that different ion types at the same dose-averaged LET (and otherwise identical conditions, see sections 1.4.3 and 1.4.4) will in general exhibit different RBEs (Friedrich et al 2013a, 2013b). This has to be considered when relating dose-averaged LET and RBE in radiobiological experiments.
1.4.3. Dose
From figure 1(a), it can be seen that the shapes of the survival curves differ for photons and carbon ions. This implies a dependence of the RBE values on the selected survival fraction (i.e. the biological endpoint) and as the latter depends on dose, also a dose dependence of the RBE. Generally, the RBE increases with decreasing dose, however, the amount of this increase depends strongly on LET.
In contrast to cell culture experiments, in vivo studies (e.g. figure 1(b)) often use fractionated irradiations, and in this case two different dose dependencies may occur: (i) Using the same effect probability (e.g. 50%) and the same number of fractions for both radiation modalities (see RBE definition in section 1.2), a higher fraction number will increase the respective tolerance doses (i.e. TD50) for photons more than for high-LET carbon ions, which in turn leads to an increasing RBE with decreasing fractional dose. (ii) On the other hand, comparing the dose-response curves for photons and carbon ions at a fixed fraction number, it is seen in most cases that the curve is steeper for carbon ions than for photons. As a consequence, the RBE increases with an increasing level of effect (e.g. 50% versus 80%) and thus with increasing dose. Although both cases represent a dose dependence of the RBE, it has to be noted that the probability of the effect and thus the biological endpoint is fixed in the first case while it is subject to change in the second.
1.4.4. Biological factors
Generally, any biological factor that influences the response (i.e. cell survival, effect probability or growth delay) of the biological system differentially for photons and carbon ions will impact the RBE. In particular, this includes the definition of the biological endpoint as well as the detection method. But even if these are fixed, various cell- and/or tissue-specific factors may influence the RBE significantly. The most important biological parameters are the intrinsic radiosensitivity and the fractionation dependence given by the α- and α/β-values of the linear-quadratic model (Fowler 1989). Especially for tumors, however, there may be additional factors that are responsible for radioresistance. Not all of these factors are known, but candidates are slowly growing or quiescent tumor subpopulations as well as changes in micro-environmental conditions like hypoxia, which modulates the radiosensitivity against photons and carbon ions differently.
1.5. Scope of this review
This topical review summarizes current knowledge on carbon ion RBE, explicitly excluding the RBE of protons and other heavier ions. For an outline on the important question whether the global proton RBE of 1.1 should be replaced by a more sophisticated model, we refer to the recent articles by Paganetti (2014, 2015). Although of clinical interest, heavy ions other than carbon are also not addressed as they are currently not applied in patients and since only very few experimental data are available. Also planning studies are not addressed as they only apply RBE models, but do not contribute to their validation.
This topical review is organized as follows: in section 2, the physical and potential biological causes for an increased effectiveness of carbon ions are summarized. This essentially contains a description of measurable biological endpoints, which are produced by different doses of photons or carbon ions, respectively, and which as a consequence result in an increased RBE. Section 3 summarizes the available in vitro and in vivo RBE data. Section 4 outlines the most important RBE models focusing on those that are clinically applied. Here, only a basic overview of the most important features and parameters rather than a complete description of all technical details is provided. For the latter, the reader is referred to the original publications on these models. Section 5 describes how these RBE models are applied in the clinic. As all RBE models were initially developed based on in vitro data, special emphasis is put on their transfer into clinical application and the associated selection of model-parameters. Section 6 finally discusses current attempts to validate RBE models experimentally and clinically. As the clinical applications in different centers use specific model features to a varying extent, the validation may be more experimentally than clinically oriented in some cases. Section 7 finally summarizes and discusses the current status of the use of the RBE concept in carbon ion therapy and points out clinically relevant conclusions as well as open questions.
2. Physical and biological causes for the increased RBE
2.1. Track structure
The underlying reason for the altered biological effectiveness of carbon ions as compared to photons is the different way they transfer their energy on a microscopic scale. While high-energetic photons transfer a substantial fraction of their energy to secondary electrons, predominantly via the Compton-effect, ions most likely undergo Coulomb-interactions and release a large number of secondary electrons with very low energies, mostly in the keV range. In both cases, the secondary electrons lose their kinetic energy along their residual range, which is typically 1–3 cm for photons, but much less for carbon ions. As a consequence, the secondary electrons released by photons spread their energy over large distances from the primary interaction point while the secondary electrons originating from ions deposit their energy essentially within a small radius around the primary ion track (Krämer and Kraft 1994a, 1994b). This maximum radius decreases from 156 µm to 3 µm and 0.06 µm for carbon ion energies of 100, 10, and 1 MeV u−1, respectively (Elsässer et al 2008). As the maximum track radius decreases and the LET increases with decreasing energy, the local dose around the carbon ion track (i.e. the specific energy) increases strongly towards the distal edge of the SOBP.
Biologically, the increase of the locally absorbed doses causes a higher density of direct radiation damage and on a microscopic scale it has been shown that the molecular response of cells is spatially correlated with the location of the ion track (Scholz et al 2001, Jakob et al 2002, 2003). As a consequence, the response in terms of cell survival (in vitro) or the probability of an effect (in vivo) increases relative to photons corresponding to an increased RBE. In the following, the known biological effects of carbon ions in single cells and tissues are summarized.
2.2. Basic radiobiological effects
2.2.1. Primary DNA damage pattern
Charged particles with their higher ionization density along the radiation tracks more effectively produce DNA damage such as double-strand breaks (DSBs), congestions of two or more abasic sites as well as other forms of base damage, or single-strand breaks in close vicinity of defect bases. Damage to DNA in the form of multiple lesions are referred to as being clustered (Goodhead 1994, Prise 1994) and are defined as two or more closely associated DNA lesions, involving both strands (bi-stranded lesions) usually within one or two helical turns (Sutherland et al 2001, Hada and Georgakilas 2008).
Based on Monte Carlo simulations the expected number of individual DNA lesions per unit of absorbed dose is about the same for low- and high-LET radiations (Semenenko and Stewart 2006). Yet, with increasing LET the proportion of complex lesions accumulate (from 30%–40% for low-LET to >90% for high-LET radiation) and they are distributed within smaller segments of DNA, resulting in a higher level of cluster complexity. Moreover, while, indirect actions via the generation of reactive oxygen species (ROS) due to the radiolysis of water molecules plays an important role in conventional low-LET irradiation, direct effects on DNA is the dominant process of high-LET radiation, which might be one explanation why intrinsic cellular factors (e.g. oxygen) are of minor importance for cellular inactivation (Hirayama et al 2009; for a review see Georgakilas et al (2013)). Yet, beside DNA damage, oxidative stress to membranes and cellular organelles represents an additional pathway that enhances radiation-induced damage. Mitochondria are both a major source of ROS and a target of cellular ROS. There is also evidence suggesting that oxidative stress plays a major role especially after high-LET radiation due to the more pronounced structural and functional alteration of the mitochondrial membranes as well as by the more pronounced up-regulation of genes involved in the oxidative stress pathway (Laurent et al 2013, Sun et al 2014).
In mammalian cells, a number of different repair systems exist that deal with DNA damage. The two most important types of double-strand break repair processes are the homologous recombination repair (HRR) and the nonhomologous end-joining (NHEJ) pathways. Various studies have revealed that when energy is deposited in localized regions of either the DNA, the nucleosome, or in chromatine fibers, damage processing is less efficient and consequently renders the repair of DNA damage slower, less complete, and with lower accuracy. All these events are LET dependent with the maximum damage at 150–200 keV µm−1 and are considered to be the main cause for the higher biological effect per unit dose (Ritter et al 1977, Roots et al 1979, Weber and Flentje 1993, Ward 1994, Taucher-Scholz et al 1996, Rydberg et al 2005). Recently, it has also been shown that the spacing, but not the quantity, of the DSBs within the DNA molecule likely influences the efficacy of DNA repair (Lorat et al 2016).
One of the most obvious underlying molecular mechanisms for the refractory repair is the radiation-induced release of short DNA fragments (100–2000 base pairs), a striking characteristic feature of clustered DNA damage created by carbon ions (Rydberg 1996). DSBs are primarily repaired by the dominant, fast but error prone NHEJ repair pathway. The key player in this pathway is the Ku70/Ku80 heterodimer, which binds to DNA termini with high affinity, helps to protect DNA ends from degradation, and is responsible for the recruitment of additional repair factors (Fell and Schild-Poulter 2015). High-LET-induced short DNA fragments are unable to be bound by the Ku70/Ku80 heterodimer leaving a higher proportion of damage unrepaired by this most dominant repair system, which consequently indicates a considerable higher impact of HRR mechanisms following high-LET-induced DNA damage (Okayasu et al 2006, Wang et al 2008, Anderson et al 2010, Gerelchuluun et al 2015). Much less is known about the role of LET on radiation-induced damage to nuclear proteins, which are mandatory for proper DNA processing, structuring, and packaging.
2.2.2. Cell cycle and mode of cell death
The DNA damage response pathway is a multistep process which involves damage recognition, information transfer, cell cycle regulation via checkpoints, and the activation of appropriate repair systems. When proliferative active cells are irradiated, DNA lesions induced by radiation activate checkpoints that temporarily slow down cell cycle preventing or limiting the entry of cells into the S phase or mitosis in the presence of unrepaired damage and hence, allowing time for genomic reconstitution. In the case of unrepairable DNA lesions, cells undergo permanent cell-cycle arrest or cell death.
Besides the described repair-associated cell cycling interference, an additional cell cycle dependent radiation effect exists. Mammalian cells reveal a differential response to low-LET radiation, when treatment occurs in well-defined cell cycle phases with a maximal radiosensitivity shortly before and during mitosis and with resistance rising during the DNA synthesis, reaching a peak in the late part of the S phase. Low cycling or quiescent cells, in human tumors often localized in oxygen- and nutrition-deprived tissue sites, are thought to be most resistant to radiation because of their larger capacity to recover from radiation-induced DNA damage (Masunaga and Ono 2002, Pawlik and Keyomarsi 2004).
In vitro studies focusing on cell cycle effects after high-LET irradiation have shown a considerably more pronounced cell cycle arrest induced by carbon ions, especially in the G2-phase, predominantly induced by the occurrence of clustered DNA damage and the more sophisticated demands for DNA repair (Lucke-Huhle et al 1979, Fournier and Taucher-Scholz 2004, Hu et al 2014).
In addition, when synchronized cells are irradiated with high-LET particles the variation in cell cycle-related radiosensitivity is reduced. Especially cells in the phase of DNA synthesis are considered to be more sensitive to high-LET radiation, presumably due to dysfunctional repair pathways. For increasing LET values, a general decrease in the variation of radiosensitivity, depending on the phase, has been observed (Bird and Burki 1975, Blakely et al 1989, Fournier and Taucher-Scholz 2004, Wang et al 2008, Wang et al 2009).
Beside cell cycle arrest, DNA damage induced by radiation generates a complex cascade of events leading to transcriptional and post-transcriptional activation of a subset of genes triggering cell death if cells fail to repair DNA damage. For radiation, mitotic cell death, necrosis, and premature-senescence have been described as the major processes of cell inactivation. Even the induction of autophagy, an evolutionarily conserved cellular recycling process (Ohsumi 2014), was observed in tumor cells after carbon ion treatment (Hino et al 2010, Jinno-Oue et al 2010).
Mitotic cell death is often found in p53-mutated tumors that are resistant to genotoxic damage. The process is initially characterized by chromosome missegregation followed by aberrant mitosis or imperfect chromosome segregation, leading to the formation of multinucleated cells. Moreover, the involvement of apoptotic cell death or mitosis restitution has been suggested at the end of this process (Erenpreisa and Cragg 2001).
In tumors harboring mutated p53, resistance to radiation therapy is thought to result from the failure of x-rays to induce a sufficient level of apoptosis. In contrast, high-LET radiation can induce apoptosis more effectively regardless of the cellular p53 gene status. The precise mechanism involved is presently not clear. There is evidence that following high-LET exposure, caspases, a family of protease enzymes playing an essential role in cell inactivation, are upregulated via an alternative p53-independent pathway (Yamakawa et al 2008). More recent studies point out the impact of ceramide as an upstream key regulator (Alphonse et al 2013). As this bioactive lipid is implicated in a variety of physiological functions including apoptosis, cell growth arrest, differentiation, and senescence, it puts a spotlight on cellular membranes as an alternative target to DNA in radiation-induced cell-response. The RBE values for the surviving fraction and the induction of apoptosis were increased in a LET-dependent manner (Takahashi et al 2004, Mori et al 2009), yet, it should be kept in mind that in general, biological functions are strongly dependent on the intrinsic cellular genetic profile and hence the mode of cell death is not influenced solely by radiation quality.
2.2.3. Therapeutic response
The exposure of biological systems to irradiation is characterized by a sequence of events of physical, physical–chemical, and biological processes, the sum of them determining the therapeutic outcome. A simplified view is, whether a cell is inactivated or survives treatment either without any change or at the cost of genetic alterations. Cellular viability, defined as the ability of a cell to preserve its physical and metabolic integrity as well as its clonogenicity, defined as the ability of a cell to undergo cell division, are the most important measures of cell survival. Both endpoints can be assessed by simple and rapid techniques such as vital staining, evidence of metabolic activity, and clonogenic assays. The latter are mainly used to establish radiation dose-survival curves. Typically, these curves express cellular survival fraction S(D) as a function of the irradiation dose,  , (figure 1(a)). While for low-LET radiation, the survival curve is characterized by an initial shoulder region followed by an exponential survival decrease, survival curves obtained for cells exposed to high-LET radiation exhibit no shoulder and are entirely exponential, which is represented by a straight line (Cox et al 1977, Blakely et al 1979). Many intrinsic factors contribute to the radiation tolerance of cells. Of outstanding importance is the genetic background of a cell, especially the status of the DNA damage checkpoints, the signaling pathways, and the proficiency and the capacity of the various repair systems. Biological consequences are ambivalent. While certain deficiencies in DNA repair pathways lead to radiosensitization, up-regulated oncogenes, mutated tumor suppressor genes, and dysregulated cell death pathways are able to increase radiation resistance. Other parameters which influence the radio-induced cellular lethality include the number and rate of cell proliferation, the cell cycle distribution, the concentration of radical scavengers and anti-oxidative enzymes, the metabolic status as well as the intracellular micromilieu. Monolayer cell cultures possessing well defined genetic characteristics, either intrinsically acquired or modulated by genetic manipulation, have been intensively used to decipher underlying radiobiological mechanisms and to determine the differential effectiveness of low- versus high-LET radiation. More recently, complex three-dimensional (3D) organotypic cell cultures and multicellular spheroids, which aim to better simulate the in vivo characteristics of intact organs have gained importance (Eke and Cordes 2011). Finally, animal models are necessary to validate in vitro findings, focusing on tumor and normal tissue response with biological endpoints, reflective of clinical outcomes in the human disease. Meanwhile, a variety of models exist, including syngeneic tumors and human tumor xenografts as well as transgenic tumor models (Teicher 2006). The most relevant assays for quantifying the radiation response in vivo are summarized in figures 1(b)–(d).
, (figure 1(a)). While for low-LET radiation, the survival curve is characterized by an initial shoulder region followed by an exponential survival decrease, survival curves obtained for cells exposed to high-LET radiation exhibit no shoulder and are entirely exponential, which is represented by a straight line (Cox et al 1977, Blakely et al 1979). Many intrinsic factors contribute to the radiation tolerance of cells. Of outstanding importance is the genetic background of a cell, especially the status of the DNA damage checkpoints, the signaling pathways, and the proficiency and the capacity of the various repair systems. Biological consequences are ambivalent. While certain deficiencies in DNA repair pathways lead to radiosensitization, up-regulated oncogenes, mutated tumor suppressor genes, and dysregulated cell death pathways are able to increase radiation resistance. Other parameters which influence the radio-induced cellular lethality include the number and rate of cell proliferation, the cell cycle distribution, the concentration of radical scavengers and anti-oxidative enzymes, the metabolic status as well as the intracellular micromilieu. Monolayer cell cultures possessing well defined genetic characteristics, either intrinsically acquired or modulated by genetic manipulation, have been intensively used to decipher underlying radiobiological mechanisms and to determine the differential effectiveness of low- versus high-LET radiation. More recently, complex three-dimensional (3D) organotypic cell cultures and multicellular spheroids, which aim to better simulate the in vivo characteristics of intact organs have gained importance (Eke and Cordes 2011). Finally, animal models are necessary to validate in vitro findings, focusing on tumor and normal tissue response with biological endpoints, reflective of clinical outcomes in the human disease. Meanwhile, a variety of models exist, including syngeneic tumors and human tumor xenografts as well as transgenic tumor models (Teicher 2006). The most relevant assays for quantifying the radiation response in vivo are summarized in figures 1(b)–(d).
2.2.4. Genomic instability and mutagenicity
When radiation-induced DNA damage is extensive and check-point functions are impaired as often found in tumor cells, unrepaired DNA could pass through these control units, leading to significant chromosomal aberrations during the mitotic phase (Löbrich and Jeggo 2007, Asaithamby et al 2011). In the case of particles, the energy deposition along the track not only causes damage on the DNA molecule itself but also on chromatin fibers or in adjacent chromosome territories. Due to the higher number and the large scale distribution of more complex DSBs, the number of chromosome breaks differs substantially between low- and high-LET irradiation. Moreover, the type and the complexity of aberrations are altered all together having a dramatic impact on cell viability. High-LET radiation induces a higher frequency of chromatid breaks than low-LET radiation. The kinetics of chromatid break rejoining consists of two exponential components having a rapid and a slow time constant, which appears to be similar for low- and high-LET radiations. However, after high-LET radiation exposures, the rejoining process for isochromatid breaks influences the repair kinetics of chromatid-type breaks. This plays an important role in the assessment of chromatid break rejoining in the G2 phase of the cell cycle resulting in more severe biological effects (Durante et al 1998, Kawata et al 2004).
Recent studies on how radiation-induced chromosome breaks are formed have shown that also conformational processes associated with the accessibility and functionality of DNA repair enzymes contribute to chromatin breaks. Changes such as unfolding of chromatin lead to an unfavorable energy state and eventually to chromosome breaks by mechanical stress, which cannot be restituted. If no conformational chromatin alteration occurs, a correct DNA repair may take place or misrepair may lead to chromosome exchanges and rearrangements (Pantelias and Terzoudi 2010).
2.2.5. Signaling and gene expression
Existing evidence suggests that radiation exposure affects global gene expression associated with various biological effects in mammalian cells. As a result, a large amount of data has been collected on low-LET-induced changes in gene expression (Khodarev et al 2001). Much less is known about the effect of high-LET particle irradiation on global gene expression in mammalian cells. In general, the expression of many genes takes place independently of the radiation quality. Examples are the overexpression of genes responsible for cell regulation, cell inactivation, oxidative stress response, signal transduction, and energy pathway. Yet, the magnitude of the change in expression is dependent on the LET. Genes specifically altered by carbon ion irradiation but not by x-rays include SPHK1, a secondary messenger in cellular proliferation and survival (Higo et al 2006), the transforming growth factor in the ß-signaling pathway (TGF-ß) and cell cycle regulators (Fushimi et al 2008, Matsumoto et al 2008) as well as genes involved in stress-response, cell-communication, and motility processes (Imadome et al 2008, Suetens et al 2014). Although most of these results are performed on cultured cells possessing a clear cell- and tissue-type dependency, they might provide the underlying molecular basis which would help to explain the increased efficacy of carbon ions.
3. RBE measurements
3.1. Cellular systems
In cell cultures many studies have shown that high-LET irradiation is more effective than low-LET radiation for the process of cell inactivation (Cox et al 1977, Blakely et al 1979). In spite of possible influences from interlaboratorial biological and technical differences which might considerably aggravate the interpretation and comparison of measured RBE values, extensive data collections have provided a rather reliable data set of RBE values. For various human cell lines, RBE values ranged between 1.06 and 1.32 for the entrance region (LET = 13.3 keV µm−1) and 2.00 and 3.01 for the Bragg peak region (LET = 77 keV µm−1), detected as reproductive cell death using colony-formation assays (Suzuki et al 2000, Belli et al 2008). While there exists a clear therapeutic effect correlated with LET with nearly no distinguishable difference between normal and tumors cells, a differential response for the same LETs is less obvious (Ando and Kase 2009).
In many studies, the RBE for carbon ions clearly correlates with photon radiosensitivity, exemplified by cell lines deficient in DNA repair pathways, which are more sensitive to photons and hence, reveal an RBE close to unity as shown for cell killing in NHEJ defective cells (Eguchi-Kasai et al 1998, Weyrather et al 1999).
Conversely, many tumors are characterized by genetic alterations, which confer radioresistance. Some mutations in the tumor suppressor gene p53, a protein known to play a central role in DNA repair, cell cycle regulation, and apoptosis (Levine 1997), are able to cause a restricted response to cellular stress, often associated with increased resistance to low-LET ionizing radiation. Similarly, in cancerous cells overexpressing survivin or BCL-2, both proteins involved in the suppression of apoptosis render cells therapy resistant (Jin et al 2008, Hamada et al 2008a). Compared to conventional radiation, cell inactivation after exposure to high-LET particles is more pronounced in such systems, which results in a higher RBE, giving the genetic background a minor importance for some but not all tumor cells. Recent findings indicate the existence of glioblastoma cells with radioresistance to both low- and high-LET irradiation, presumably associated with the status of their intrinsic genome integrity (Dokic et al 2015). Likewise, some sub-clones of cancer cells generated radiation resistant by repeated exposure to x-rays and also reveal resistance to carbon ions, pointing to possible common radiation quality independent damage-response pathways (Sato et al 2014).
Friedrich et al established a large particle irradiation data ensemble based on published cell survival curves after irradiation with different ion types and analyzed the RBE as a function of LET and  , the frequency distributions of
, the frequency distributions of  ,
,  and
and  for photon irradiations as well as the correlation between
for photon irradiations as well as the correlation between  - and
- and  -values for photons (Friedrich et al 2013b). However, no numerical values for RBE,
-values for photons (Friedrich et al 2013b). However, no numerical values for RBE,  or
or  are provided for further analysis.
are provided for further analysis.
3.2. Animal models
In the clinical situation, there is still a lack of knowledge about the responses of various tumor types, as well as of early and late responding normal tissues. While cell lines are predominantly useful to search for underlying mechanisms, whole animal models are helpful to validate tolerance doses, fractionation effects, and clinical efficiency.
3.2.1. Normal tissues
Determining normal tissue RBEs is of outstanding relevance because (i) correlation of LET and RBE is not necessarily linear across a complete range and may also depend on dose/fraction, (ii) in spite of the high physical accuracy, a non-negligible volume of normal tissue surrounding the tumor is in close proximity to the planning target volume (PTV) and even occasionally included in the (high-LET) volume, and (iii) all solid tumors contain normal tissue structures, e.g. stromal structures, vascular elements etc. and many tumor cells are intertwined in a substrate of normal tissue.
Radiation-induced normal tissue lesions reveal differences in their temporal appearance. Early effects occurring with the onset of radiation are associated with cell inactivation of rapidly proliferating radiosensitive cells, increased endothelial cell swelling, vascular permeability, and edema as well as lymphocyte adhesion and infiltration. Recovery processes start with repair and repopulation of stem cell pools within individual tissue compartments and, depending on the turnover time of the tissue, continue for months. In contrast, late reactions occur months to years after radiation and are characterized by the induction and sustained up-regulation of specific molecular signaling pathways, the depletion of tissue-specific stem cells and vascular damage, leading to tissue remodeling, organ dysfunction, and fibrosis (Hall 2001, Stone et al 2003, Rodemann and Blaese 2007).
Although several studies have been performed to determine RBE values in vivo, the available data is still limited. Table 1 compiles the existing data.
Table 1. Compilation of in vivo studies measuring the RBE of carbon ions in normal tissue.
| Organ | Host | Endpoint | Fx | LET (keV µm−1) | RBE | Reference |
|---|---|---|---|---|---|---|
| Skin | Golden Syrian hamster | Average ventral thoracic skin reaction level | 1 | n.r. | 1.60 | Leith et al (1981) |
| 2 | Modified Bragg peak | 1.75 | ||||
| 5 | 1.90 | |||||
| Spinal cord | CDF1 mice | Radiation-induced myelopathy |
1 | 10 | 1.45 | Leith et al (1982b) |
| 80 | 1.48 | |||||
| 4 | 10 | 1.31 | ||||
| 80 | 1.95 | |||||
| Skin | CDF1 mice | Residual skin damage 1 year after RT (ED50 equivalent) |
4 | 10 | 1.04 | Leith et al (1982a) |
| 4 | 80 | 1.53 | ||||
| Skin | C3H/HeMsNrsf mice | Moist des-quamation |
1 | 14 | 1.45 |
Ando et al (1998) |
| 20 | 1.75 | |||||
| 42 | 2.15 | |||||
| 77 | 2.50 | |||||
| 2 | 14 | 1.35 | ||||
| 20 | 1.40 | |||||
| 42 | 1.50 | |||||
| 77 | 2.40 | |||||
| 4 | 14 | 1.40 | ||||
| 20 | 1.60 | |||||
| 42 | 1.85 | |||||
| 77 | 3.20 | |||||
| 8 | 14 | 1.60 | ||||
| 20 | 1.90 | |||||
| 42 | 2.25 | |||||
| 77 | 3.20 | |||||
| Spinal cord | SLC Wistar rats | Radiation-induced myelopathy |
1 | 70 | 1.38 | Okada et al (1998) |
| Developing brain | SLC Wistar rats | Microcephaly and histology |
1 | 50 | 1.3–1.6 | Inouye et al (2000) |
| Liver (after partial hepatectomy) | Balb/c mice | Hepatic failure LD50/60 (50% lethal dose within 60 d) |
1 | 50,7 | 1.86 | Tomizawa et al (2000) |
| Brain | Copenhagen rats | MRI contrast enhancement 20 months post RT (50% effect probability level) |
1 | 155 | 1.95 | Karger et al (2002) |
| Intestine | Balb/c mice | Intestinal crypt regeneration |
1 | 13,7 | 1.3 | Gueulette et al (2004) |
| 40,9 | 1.6 | |||||
| 49,4 | 1.7 | |||||
| 70,7 | 1.9 | |||||
| Intestine | Balb/c mice | Intestinal crypt regeneration |
1 | 42 | 1.47j | Uzawa et al (2009) |
| 50 | 1.63 | |||||
| 74 | 1.80 | |||||
| 3 | 42 | 1.71 | ||||
| 50 | 1.95 | |||||
| 74 | 2.24 |
|||||
| Spinal cord | Sprague Dawley rats | Radiation-induced myelopathy (ED50) |
1 | 13 | 1.44 | Debus et al (2003) |
| 91 | 1.77 | |||||
| 2 | 13 | 1.37 | ||||
| 91 | 2.17 | |||||
| Spinal cord | Sprague Dawley rats | Radiation-induced myelopathy (ED50) |
6 | 13 | 1.33 | Karger et al (2006) |
| 91 | 2.97 | |||||
| 18 | 13 | 1.42 | ||||
| 91 | 5.04 | |||||
| Spinal cord | Sprague Dawley rats | Radiation-induced myelopathy (ED50) |
1 | 16 | 1.26 | Saager et al (2014) |
| 21 | 1.33 | |||||
| 36 | 1.39 | |||||
| 45 | 1.52 | |||||
| 66 | 1.68 | |||||
| 99 | 1.83 | Saager et al (2016) | ||||
| Spinal cord | Sprague Dawley rats | Radiation-induced myelopathy (ED50) |
2 | 16 | 1.28 | Saager et al (2015) |
| 21 | 1.43 | |||||
| 36 | 1.52 | |||||
| 45 | 1.71 | |||||
| 66 | 1.94 | |||||
| 99 | 2.30 | |||||
| Skin | CDF1 mice | Radiation-induced fibrosis (FD50) |
1 | 65 | 1.50 | Sørensen et al (2015) |
Photon reference: aCs-137 γ-rays. bX-rays (140 KV). cX-rays (200 KV). dX-rays (230 KV). eX-rays (240 KV). fCobalt-60. gPhotons (6 MeV). hPhotons (15 MeV). iAll RBEs recalculated from figure 6 in Ando et al (1998). jAll RBEs determined at the Helmholtz Centre for Heavy Ion Research (GSI). n.r.: not reported.
From a biological point of view, the response of normal tissue to radiation is mainly influenced by the proliferative and functional tissue organization, the regional differences in radiosensitivity within an organ, and possible interactions between organs, while biophysical factors consist of the total dose, the timing, and the irradiated volume. Early and late radiation-induced normal tissue response as well as volume effects are clearly determined by the structural tissue organization, but nowadays most tissues are considered to be organized in mixed structures consisting of both parallel and serial arrangements. In conventional low-LET radiotherapy, applying low doses per fraction over an extended time period was the preferred strategy to allow for repair and repopulation and to maximize normal tissue sparing.
In contrast, two different objections play a role, when high-LET carbon ion response to normal tissue is evaluated: (i) normal tissue located proximal to the PTV in the entrance channel of the beam, and (ii) a non-negligible volume of normal tissue surrounding the tumor might be included in the (high-LET) volume, due to uncertainties in defining the irradiated treatment volume and to physiological organ movement in the body. This is of special importance when organs of risk, such as optic nerves, large arteries, or radiosensitive organs are in close proximity to the safety margins. Both aspects have already been investigated in the very early pilot studies with early and late responding tissues. A possible favorable differential RBE of skin between the target volume and the entrance channel of the beam was demonstrated by Leith et al. when the dose was applied in four fractions (Leith et al 1982a). The first extensive systematic studies were performed end of the 1990s by Ando et al using the qualitative grading of skin reactions in mice in a single and fractionated animal experiment (Ando et al 1998). Although acute skin reactions occurred after lower doses and were mostly reversible, the authors could demonstrate for the first time a differential fractionation effect of carbon ions compared to photons, which resulted in a clear increase of RBEs with decreasing dose per fraction. The intestinal crypt assay, another early responding tissue system turned out to be a practical in vivo assay to compare the biological efficacy of varying beams, operating in different institutions and countries (Gueulette et al 2004).
The neutron experience taught us that late effects, which exhibit a more complicated pathophysiology and are difficult to cure, should be prevented. Yet, because long range clinical observations for carbon ions are still pending, the early collected radiobiological knowledge to late radiation effects in the rat spinal cord as an established animal model was extended. The first systematic examination was performed in a 1 cm SOPB at two different LETs with 1, 2, 6, and 18 fractions using radiation-induced myelopathy as a strong, non-reversible biological endpoint, which occurred in rats up to 300 d post irradiation (Debus et al 2003, Karger et al 2006). The results demonstrated that carbon ion irradiations are significantly more effective in the peak than in the plateau region. While the relatively low RBEs in the entrance channel, which ranged from 1.3–1.4, still retain the normal tissue sparing effect, the measured RBEs in the SOBP increased from 1.8 in a single fraction to 5 after 18 fractions. The more prominent the sparing effect of photons, the higher the increase in RBE in the SOBP, which is attributed to the low fractionation effect of carbon ions, showing little change in isoeffective total dose for myelopathy with fractionation. Recently, results of the single dose experiments were confirmed in radiation-induced fibrosis, another irreversible late damage effect of irradiation resulting in closely related RBEs (Sørensen et al 2015). Because for high-LET particles as carbon ions, the modulated beam is heterogeneous in RBE along the beam path in an extended Bragg peak, the small size of the serially organized spinal cord tissue allowed the study of the dependence of RBE on LET. For that purpose, the RBE for induced myelopathy in the rat spinal cord after 1 and 2 fractions of carbon ion doses was measured at six different positions of a 6 cm spread-out Bragg-peak. In the investigated dose range, the data suggest an obviously linear relation between RBE and LET for late effects in the cervical spinal after 1 and 2 fractions (Saager et al 2014, 2015, 2016).
It should be noted that the different studies used various reference beams. This however is considered to have only a minor influence on RBE in the range of 10%–20%. Additional studies are urgently needed, determining RBEs of late effects for different fractionation schemes and tissues of varying cell repair mechanisms including a better understanding and prediction of carcinogenic risk for pediatric patients.
3.2.2. Solid tumors
Determining RBEs in animal tumor models is more complex. In addition to the inherent or acquired genetic alterations of the tumor cells, structural elements such as the basement membrane, fibroblasts, extracellular matrix, immune cells, vasculature, and functional parameters like blood flow, nutrition, and oxygen supply gain importance. Furthermore, radiation generates ROS, promotes tumor cell repopulation, exerts pro-angiogenic effects via the activation of prosurvival signaling cascades, and triggers an immune response (Good and Harrington 2013, Junttila and de Sauvage 2013). In contrast to in vitro studies, it is presently not feasible to fully assess the significance or the contribution of each biological factor to the complex dynamics of therapy resistance in vivo, where all factors interact simultaneously. Both intrinsic and extrinsic factors emerge as biological properties that lead to resistance and shape intra-tumoral heterogeneity, and consequently influence therapeutic response. Moreover, the selected biological endpoint significantly influences treatment strategies and hence RBE values. While measurement of tumor growth delay reflects a generalized kill of tumor cells in the lower dose range, tumor control assays depend solely on the inactivation of clonogenic cells (Zips 2009).
As compared to normal tissues, much less studies investigated the RBE of carbon ions in experimental solid tumors. A compilation of the available data is given in table 2.
Table 2. Compilation of in vivo studies measuring the RBE of carbon ions in tumors.
| Tumor | Host | Endpoint | Fx | LET (keV µm−1) | RBE | Reference |
|---|---|---|---|---|---|---|
| 9L brain tumor | Fisher 344 rats | In situ/clonogenic survival | 1 | n.r. | 1.33 |
Wheeler et al (1979) |
| Rhabdomyo-sarcoma | WAG/Rij rats | Growth delay |
1 | 12 | 1.3 | Tenforde et al (1981) |
| 1 | 80 | 2.3 | ||||
| Human esophagus carcinoma | BalbC/Ajcl/nu mice | Growth delay |
1 | 70 | 2.02 | Takahashi et al (1998) |
| NFSa fibrosarcoma | C3H/He mice | Growth delay |
1 | 14 | 1.4 | Koike et al (2002) |
| 1 | 44 | 1.8 | ||||
| 1 | 74 | 2.4 | ||||
| 4 | 44 | 2.3 | ||||
| 4 | 74 | 3.0 | ||||
| 6 | 44 | 2.3 | ||||
| 6 | 74 | 3.0 | ||||
| Dunning prostate carcinoma | Copenhagen rats | Tumor control assay |
1 | 75 | 2.30 | Peschke et al (2011), Karger et al (2013) |
| 2 | 75 | 2.39 | ||||
| R3327-AT | 6 | 75 | 2.67 | |||
| Dunning prostate carcinoma | Copenhagen rats | Tumor control assay |
Glowa et al (2016) | |||
| R3327-HI | 1 | 75 | 2.08 | |||
| R3327-H | 1 | 75 | 1.62 | |||
| C3H mammary carcinoma | C3H/He mice | Tumor control assay |
1 | 65 | 1.48 | Sørensen et al (2015) |
Photon reference: aCs-137 γ-rays. bX-rays (200 KV). cX-rays (220 KV). dX-rays (240 KV). ePhotons (6 MV). fRelative RBE (RBESOBP/RBEplateau) under oxic conditions. n.r.: not reported.
Most of the early studies were performed with fast growing tumors using single dose experiments and growth delay as biological endpoint to define RBEs. As tumors experience heterogeneous LETs across their treatment volume, average LETs are indicated as a measure of radiation quality. Even with the lower doses applied in all growth delay studies, most of the tumors showed RBEs ranging around 2. The only fractionated experiment, which investigated a mouse NFSa fibrosarcoma model using up to 6 Fx and different LET values, was performed by Koike et al (2002). These authors did not observe a fractionation dependence of the RBE above 4 Fx at a LET value of 74 keV µm−1 and only a minimal impact on fractionation was seen at and below 20 keV µm−1.
More recent studies (Peschke et al 2011, Karger et al 2013, Sørensen et al 2015, Glowa et al 2016) are based on local tumor control assays. This experimental design is not only demanding much higher therapeutic doses but is also considered to be of higher clinical relevance because even the last surviving tumor cells have to be eradicated. For single doses, at least four different tumor models show RBEs between 1.48 and 2.4 for an LET range of 65–80 keV µm−1 (table 2). The only fractionated experiment with a radioresistant prostate adenocarcinoma exhibited an increase of RBE with higher fraction number with only minor differences to the RBE curve measured by Koike et al for the same LET (Peschke et al 2011, Karger et al 2013). An interesting finding was that the increase of the tumor RBEs with decreasing dose was much weaker than for the spinal cord as a late reacting normal tissue (Karger et al 2006, 2013). In an additional study, the influence of structural or functional intratumoral characteristics on the RBE was examined in three sublines of an experimental prostate tumor differing in grading (Glowa et al 2016). Radiation doses required for local tumor control differed significantly less and the dose-response curves were steeper for carbon ions, as compared to photons. Both results indicate a minor impact of tumor heterogeneity on therapy outcome after carbon ion therapy. Furthermore, the increase of RBE with tumor grading is primarily caused by a higher radiation resistance against photons, while the tolerance against carbon ions remains almost unaffected. Which of the key biological features, such as hypoxia, DNA damage repair, angiogenesis/vasculogenesis, cancer stem cells, tumor stroma, and the immune response pathways is governing tumor responses to photon radiation is presently under investigation.
3.2.3. Influence of hypoxia
Extensive studies have been performed in the past to elucidate the impact of oxygen deprivation on the radiation response of tumors. Tissue hypoxia results from the inadequate supply of oxygen that compromises biologic functions. In solid tumors, oxygen delivery to neoplastic and stromal cells is frequently reduced or even abolished by increased vessel distances, severe structural abnormalities of tumor microvessels, and disturbed microcirculation. Cancer cells have adapted these pathways, allowing tumors to survive and even grow under hypoxic conditions, and hence tumor hypoxia is now recognized as a major aspect of malignant progression, poor prognosis, and resistance to therapy (Hockel and Vaupel 2001, Harris 2002).
The effect of oxygen in sensitizing cells to radiation is quantified by a factor called oxygen enhancement ratio (OER)

given as the ratio of doses in the absence and presence of oxygen, respectively, needed to obtain the same biological effect. For anoxic cells irradiated with photons in vitro, the OER is about 3. With increasing LET, the OER decreases and reaches unity at high dose-averaged LET values of ~500 keV µm−1, suggesting a minor dependence on oxygen concentration for tumor cell killing by high-LET radiation (Barendsen 1968, Furusawa et al 2000, Antonovic et al 2013).
Spheroids are considered as avascular multicellular 3D tumor systems and were irradiated with x-rays or carbon ion beams under elevated, ambient, or restricted oxygen supply conditions. The response in the SOBP is characterized by a high RBE (4.31) and a relatively low OER in the range of 1.4 (Walenta and Müller-Klieser 2016).
Studies in animal tumor models are rare. The very early experiments were performed in 9L brain tumors on the basis of in vivo/in vitro assays, where tumors were irradiated either in anesthetized animals breathing air or 5 min after the animals were sacrificed. Immediately after irradiation, the tumors were excised, minced, trypsinized, and plated for colony formation. While the OER of carbon ions in the entrance region was similar to the OER of x-rays (~2), the OER in the SOBP was reduced to 1.7 ± 0.1 (Wheeler et al 1979).
A similar technique was used by Hirayama et al (2013, 2015). Briefly, implanted solid SCCVII tumors were irradiated with either carbon ions or photons under clamped and ambient conditions. Cell survival was analyzed using a colony formation or a micronuclei formation assay. After photon irradiation an OER of 1.87 ± 0.13 was obtained. Carbon ion OERs ranged between 1.43 ± 0.19 and 1.52 ± 0.10, yet surprisingly with no significant change of the OER across a 6 cm spread-out Bragg peak.
Using an elegant technique, Masunaga et al (2008) were able to detect and analyze the RBE of intra-tumor quiescent cells, a subpopulation of tumor cells often localized in hypoxic areas (Masunaga et al 2008). The RBE was calculated for radiation-induced micronucleus formation in solid tumors differing in their hypoxic status. Considerably high RBE values up to 4.5 were detected in hypoxic quiescent cells 12 h after high-LET treatment. As oxygenated and hypoxic cells in these tumors exhibited almost the same radio-sensitivity to carbon ions the authors concluded that these beams might be useful for suppressing the dependence on the heterogeneity within solid tumors.
3.2.4. Influence of stem cells
In recent years another subpopulation of tumor cells has attracted notice because of their capacity to self-renew and to generate the heterogeneous lineages of cancer cells that comprise the tumor. As permanent local tumor control is the ultimate goal for curative radiotherapy, the inactivation of such cancer stem cells (CSCs) is considered to be of major importance (Baumann et al 2008). If CSCs are more radioresistant than their non-clonogenic counterparts is presently a matter of intensive research and is highly dependent on the identification and discrimination of CSCs from non-stem cells. To date there is some evidence for innate radioresistance mechanisms based on a higher DNA-repair capacity and an enhanced reactive oxygen species defense. On the other hand, studies have shown that ionizing radiation itself is able to induce a process of reprogramming in non-tumorigenic cancer cells, assigning the complex network of micro-environmental signaling within the tumor microenvironment an important role (Woodward and Bristow 2009, Ogawa et al 2013, Vlashi and Pajonk 2015).
The more efficient DNA repair machinery of CSCs, their slower cycling, their enhanced protection against ROS, and their upregulated survival signaling (Held et al 2016) make this tumor subpopulation a key target for particle therapy.
Studies on cultured cells are ambivalent. Beneficial effects after carbon ion treatment have been reported for human colon stem-like cancer cells showing RBE values at the 10% survival level between 1.63 and 1.74 as well as for pancreatic cancer stem cell-like cells with RBEs from 1.85–2.10 (Cui et al 2011, Oonishi et al 2012). RBEs for human hematopoietic stem and progenitor cells (HSPCs) are also lower (1.4–1.7) suggesting that with respect to apoptosis and chromosomal aberrations, mature lymphocytes reflect the respective radiation responses of their proliferating progenitors (Becker et al 2009). In contrast, head-and-neck cancer CSCs as well as bone marrow-derived mesenchymal stem cells are resistant to both photon and carbon-ion irradiations (Bertrand et al 2014, Nicolay et al 2015).
Two animal studies were published using a human colon carcinoma cell line (Cui et al 2011) and a human pancreatic cancer cell line (Sai et al 2015), xenotransplanted into nude mice. Radiation quality dependent treatment effects on CSCs were quantified on the histological level, showing a dose-dependent significant decrease of marker positive putative cancer stem cells after carbon ion irradiation as compared to x-rays, however, without quantifying RBEs. Clearly, additional detailed studies are urgently needed, including tumor control assays, the in vivo characterization and quantification of CSCs, and the analysis of tumor samples from patients to better understand the potential therapeutic effect of charged particles on CSCs.
3.2.5. Additional influential factors
Many additional biological processes, supporting the superior efficacy of carbon ion radiotherapy, have been reported, yet without assignment of RBE values. Carbon ions induce more pronounced damage to vascular endothelial cells and this results in a differential pO2 distribution after treatment with either photons or carbon ions (Ando et al 1999). Moreover, there is evidence that carbon ions are able to better suppress angiogenic processes (Takahashi et al 2003, Kamlah et al 2011).
To avoid metastasis, the spread of tumor cells from the primary tumor to other organ sites in the body, either naturally or induced by the treatment itself, is another important aspect for the improvement of cancer treatment, and the benefits from particles have been investigated intensively. Several in vitro studies with tumor lines differing in origin demonstrated a reduced migration activity after carbon ion treatment. The extent of migratory response in tumor cells is dependent on radiation dose, cell type, and radiation quality (Goetze et al 2007, Akino et al 2009, Ogata et al 2011).
Tumor cell migration, mainly with regard to irradiation of glioblastomas (GBMs) was of special interest due to a planned patient study. In GBMs, carbon ion irradiation was significantly more effective in interfering with cellular functions related to metastasis, such as cell motility and invasion, than photon RT (Rieken et al 2012, Walenta and Müller-Klieser 2016). The only presently available in vivo study used a mouse squamous cell carcinoma model and quantified the number of metastatic nodules in the lung, induced by treatment of primary tumors with either x-rays or carbon ions. This study could not find any differences in the incidence of metastatic events (Tamaki et al 2009).
In the last couple of years, a more detailed knowledge of biomolecular processes underlying radiation effects, including immunological aspects and their implication on therapeutic outcome, has been obtained. Meanwhile, it is well accepted that radiation is able to modify the tumor immune microenvironment leading to both, immune-stimulatory properties and adaptive antitumor effects. As ionizing irradiation-induced abscopal and bystander effects interact with the immune system of cancer patients, the role of high-LET particles has been questioned (Burnette and Weichselbaum 2013).
Currently, there is very little experimental information on bystander effects available. The effects of carbon ions to induce bystander/non-targeted effects in vitro have been reported (Fournier et al 2007, Hamada et al 2008b, Wang et al 2015). In addition, considerable peripheral immune system dysfunction, preferentially in proportion of distinct subsets of T-lymphocytes in thymus and spleen, have been described for neural cell injury caused by carbon ion irradiation (Lei et al 2015). A similar effect was reported in a syngeneic model, where carbon ions not only efficiently eliminated the primary tumor but also reduced tumor formation after secondary tumor challenge at a contra-lateral site. The antitumor effect was the result of a tumor-specific, long-lasting antitumor immunity through CD8-positive T lymphocytes (Matsunaga et al 2010).
3.3. Clinical RBE
Due to the complex dependencies of the RBE on physical and biological factors, RBE values always refer to the specific irradiation conditions, the biological system as well as to the selected biological endpoint. The resulting RBE is therefore more accurately termed as 'experimental RBE'. With this respect, all RBE values discussed in sections 3.1 and 3.2 are experimental RBEs and when referencing these values, the experimental conditions have to be clearly specified.
In contrast, the 'clinical RBE' describes the ratio of prescribed absorbed doses of a photon and a high-LET irradiation, which are believed to result in clinically equivalent results (Wambersie 1999). In contrast to the experimental RBE, the clinical RBE is an operational concept, which involves a medical decision on the basis of all presently available experimental and clinical information (Wambersie 1999) and with increasing experience, the clinical RBE may be adjusted.
The concept of a clinical RBE was previously used in neutron therapy to adjust for its increased biological effectiveness. Although the neutron RBE depends on the energy spectrum and dose, it is rather constant within the treatment field if these parameters are fixed. In contrast, the RBE within carbon ion fields varies much more and to compensate for this, the absorbed dose has to be modulated using predictions of RBE models (section 4) to achieve a homogeneous biological effect within the SOBP. This has important consequences for carbon ion RT: (i) there is no uniquely defined prescription of the absorbed dose, which can be used to determine the clinical RBE. (ii) If a clinical RBE value shall still be specified, it consequently refers to a certain particle spectrum and dose level, i.e. to a certain position within the SOBP, which may be indicated as a reference point. This approach was followed by Japanese centers for passive beam delivery, where the RBE for carbon ions at a dose-averaged LET of 80 keV µm−1 was set to the neutron value of 3.0 (Kanai et al 1999, Gueulette and Wambersie 2007). (iii) If model-based absolute RBE values are used, as done at the European carbon ion centers for scanned carbon ion beams (Krämer and Scholz 2000), the isoeffectiveness of the absorbed dose distribution depends on the correctness of the model. In this case, the clinical RBE cannot be specified independently of the RBE model and the selected model parameters. The clinical RBE is thus implicitly defined by the selected model and its input parameters.
3.4. Conclusions on available RBE data
In experimental studies, RBE values can be measured for specific irradiation conditions. None of these conditions will truly reflect the conditions in patients and also, RBE values from in vitro and in vivo-studies will differ, especially in the case of tumors with their additional influential factors on RBE and their large intrinsic heterogeneities. Nevertheless, experimental studies can help to study the functional dependencies of the RBE, which at least partly can be included in the RBE models (section 4). For safe and effective application of carbon ion RT, special concepts for the application of RBE models are necessary. These concepts are model-dependent as discussed in section 5.
4. RBE modeling
4.1. Aim of RBE models
In treatment planning, the increased biological effectiveness of carbon ions relative to photons has to be considered. Using the RBE concept allows the transformation of the absorbed ion doses into isoeffective photon doses (RBE-weighted doses), for which most clinical knowledge on dose-response is related to. Due to the complex dependencies of the RBE of carbon ions on physical and biological parameters (section 3), the RBE has to be described by more or less sophisticated models rather than by a single factor. The following subsections describe the most important RBE models with a focus on those that are clinically applied. It is a common feature of all RBE models that they are conceptionally developed based on in vitro data. Special considerations, which are necessary to apply the models in patients and thus to fix the clinical RBE, are described in section 5.
4.2. RBE based on survival curves for carbon ions and photons
As discussed in the experimental data in section 3.1, the increased effectiveness of carbon ions leads to a cell survival curve that exhibits (i) an increased initial slope at low doses and (ii) a less pronounced or even vanishing shoulder at larger doses (figure 1(a)). In terms of the parameters of the linear-quadratic model (LQM) (Fowler 1989), this relates to an increased intrinsic radiosensitivity α and a reduced repair capacity (i.e. an increased α/β-value), which both are LET-dependent. As a direct consequence of the different shapes of the survival curves, the RBE becomes dependent on dose.
It has to be noted that since the survival curves in figure 1(a) refer to single dose experiments, the concept of the RBE also refers to the differential response to single fractions. In a fractionated treatment, each fraction is considered as being independent and thus has the same RBE (an assumption which may not be true for tumors). Using the total rather than fractional doses for calculating the RBE (equation (1)) is therefore only allowed if the number of fractions is the same for photons and ions. Otherwise, effects resulting from radiation quality and fractionation effects would be mixed.
Using the expression of the LQM (Fowler 1989) for the cell survival fraction  separately for photons and ions, the RBE can be expressed as (Joiner 1989, 2009):
separately for photons and ions, the RBE can be expressed as (Joiner 1989, 2009):
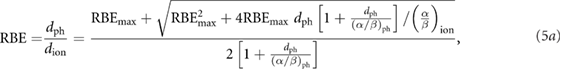
or

where  is the maximum
is the maximum  given by the initial slopes of the survival curves and
given by the initial slopes of the survival curves and  and
and  are the repair capacities for photons and ions, respectively. Equations (5a) and (5b) directly describe the dependence on fractional dose and differ only in the selection of the absorbed dose variable
are the repair capacities for photons and ions, respectively. Equations (5a) and (5b) directly describe the dependence on fractional dose and differ only in the selection of the absorbed dose variable  for photons or
for photons or  for ions, respectively.
for ions, respectively.
For treatment planning, however, the dependence of the RBE on LET is also required and this dependence is only implicitly included in equations (5a) and (5b) via the LET dependence of the parameters  and
and  , which vary also with the type of ions.
, which vary also with the type of ions.
To model this LET dependence for proton beams, equation (5b) can be reformulated into

where  is the asymptotic RBE at high doses.
is the asymptotic RBE at high doses.
As discussed before (section 1.4.2), different ion types will in general exhibit different RBEs, even for the same dose averaged LET, dose, and biological system (Friedrich et al 2013a, 2013b). In the case of protons, a simple phenomenological approach may be used by modeling  as a linear function of dose-averaged LET using a slope, which is inversely related to
as a linear function of dose-averaged LET using a slope, which is inversely related to  (Carabe et al 2012, Wedenberg et al 2013, McNamara et al 2015). This implies that
(Carabe et al 2012, Wedenberg et al 2013, McNamara et al 2015). This implies that  also depends linearly on dose-averaged LET with a
also depends linearly on dose-averaged LET with a  -dependent slope. This
-dependent slope. This  -dependent slope has also been derived from statistical considerations in the microdosimetric kinetic model (Hawkins 1998). Different assumptions were made for the dependence of
-dependent slope has also been derived from statistical considerations in the microdosimetric kinetic model (Hawkins 1998). Different assumptions were made for the dependence of  (and thus
(and thus  ) on dose-averaged LET (linear versus no dependence) and
) on dose-averaged LET (linear versus no dependence) and  (Ödén et al 2017). As a result of the above assumptions, an essentially linear dependence of the RBE on dose-averaged LET is obtained for given values of dose and
(Ödén et al 2017). As a result of the above assumptions, an essentially linear dependence of the RBE on dose-averaged LET is obtained for given values of dose and  , which fits well to the cell survival data (McNamara et al 2015).
, which fits well to the cell survival data (McNamara et al 2015).
While this linearity covers the whole range of dose-averaged LET values of clinical proton beams (Wilkens and Oelfke 2004), this is not the case for carbon ions due to the so-called 'overkill effect' at high LET values, where the RBE decreases again (Weyrather et al 1999). Therefore, more sophisticated phenomenological or mechanistic models are used for carbon ions. Whether  should be considered as LET-dependent is still a matter of discussion (Jones 2010) and the RBE models differ in this respect.
should be considered as LET-dependent is still a matter of discussion (Jones 2010) and the RBE models differ in this respect.
In general, the RBE at a certain position within the SOBP will be determined not only by the primary carbon ions, but also by the spectra of secondary particles (fragments) produced in the primary beam. Lühr et al investigated the impact of these fragments and found that although a modification of the inelastic cross section by ±20% may alter the absorbed as well as the RBE-weighted dose distribution substantially by up to 15%, the impact on the RBE distribution is only small (<1.5% in the SOBP and <3% beyond) (Lühr et al 2012). Thus, in contrast to the absorbed and RBE-weighted doses, the RBE as a relative quantity depends only weakly on the details of the secondary particle spectra.
Sections 4.3–4.5 provide a detailed description of the clinically applied RBE models. Independent of their specific features, they all attempt to describe at least the LET dependence up to high values with clinically acceptable accuracy.
4.3. Mixed beam model
For carbon ion therapy with passive delivery techniques, a phenomenological model was developed at the National Institute of Radiological Sciences (NIRS) (Kanai et al 1997). In this model, the RBE is calculated from the cell survival curves for photons and ions using a specified survival level (figure 1(a)). The basic idea of the model is to determine the LET dependence of the curve parameters  and
and  in monoenergetic ion beams in vitro and to transform these values into effective values
in monoenergetic ion beams in vitro and to transform these values into effective values  and
and  for a LET spectrum ('mixed beams') located at the depth
for a LET spectrum ('mixed beams') located at the depth  within an SOBP by
within an SOBP by

and

In equation (6),  is the dose contribution of beam
is the dose contribution of beam  at the depth
at the depth  and
and  is the respective total dose of all contributions to the SOBP. The values of the curve parameters
is the respective total dose of all contributions to the SOBP. The values of the curve parameters  and
and  for beam
for beam  at the depth
at the depth  reflect the LET dependence of the survival curve, which can be directly obtained from tabulated in vitro data measured in monoenergetic carbon ion beams (Inaniwa et al 2015). The LET distribution in an SOBP was then determined by an analytic code assuming only the presence of carbon ions. More recently, the model was refined by calculating the amount of fragments with Geant4 and by considering the biological response to helium ions as being representative of the fragments produced in the primary carbon ion beam (Sakama et al 2012).
reflect the LET dependence of the survival curve, which can be directly obtained from tabulated in vitro data measured in monoenergetic carbon ion beams (Inaniwa et al 2015). The LET distribution in an SOBP was then determined by an analytic code assuming only the presence of carbon ions. More recently, the model was refined by calculating the amount of fragments with Geant4 and by considering the biological response to helium ions as being representative of the fragments produced in the primary carbon ion beam (Sakama et al 2012).
Equation (6) was derived based on a previous work by Zaider and Rossi (1980), who calculated the combined radiation response of two different LET components using the theory of dual radiation action. Although the model makes some assumptions on the interaction of sublesions within cells, it presents a phenomenological approach in the sense that no specific information on the microscopic energy deposition pattern is required.
 and
and  describe the effective cell survival fraction for a specific particle spectrum produced by the contributions of all monoenergetic beams with a total dose
describe the effective cell survival fraction for a specific particle spectrum produced by the contributions of all monoenergetic beams with a total dose  at the depth
at the depth  . Comparing the ion dose,
. Comparing the ion dose,  , with the respective photon dose,
, with the respective photon dose,  , (also termed as biological dose,
, (also termed as biological dose,  ) at the same survival level
) at the same survival level  (e.g. 10%) allows the calculation of the RBE (Inaniwa et al 2015):
(e.g. 10%) allows the calculation of the RBE (Inaniwa et al 2015):

4.4. Local effect model (LEM)
In contrast to the phenomenological mixed beam model, the LEM uses microscopic features of the energy deposition of the ions around their tracks to explain the increased RBE and it is thus termed as 'track structure model' (Scholz and Kraft 1992, 1996, Scholz 1996, Scholz et al 1997). The basic assumption of the LEM is that the biological effect (e.g. cell death) is induced by a single ('lethal') event caused by a local energy transfer and that the damage probability depends only on the amount of the locally deposited energy, no matter whether the energy deposition originates from photons or ions. The increased effectiveness of carbon ions relative to photons is thus considered as a consequence of a different microscopic distribution of the energy deposition within the critical target (section 2.1).
4.4.1. Basic principle
The LEM assumes that the critical target of the cell is uniformly distributed over the volume of the cell nucleus, V, without any variation of radiosensitivity within the nucleus or between nuclei of different cells (Scholz and Kraft 1992). Dividing the cell nucleus into many small subvolumes, each of them having the typical size of a lethal damage, the cell will survive only if none of the subvolumes experiences a lethal damage.
For a photon irradiation with a uniform dose, D, it is assumed that the local dose is also uniformly distributed over the nucleus. In this case, the damage probability is equal for all subvolumes and the survival probability of the cell can then be calculated from Poisson statistics, assuming zero lethal events in the nucleus (Scholz and Kraft 1992):

where  is the average number of lethal events in the nucleus at a given dose
is the average number of lethal events in the nucleus at a given dose  . Resolving equation (8), the average number of lethal events can be expressed as
. Resolving equation (8), the average number of lethal events can be expressed as

As  describes the survival probability of the cell at the dose
describes the survival probability of the cell at the dose  , it also describes the survival fraction of a cell population after photon irradiation at the same dose and can thus be identified with any mathematical expression describing the cell survival curve for photons (figure 1(a)).
, it also describes the survival fraction of a cell population after photon irradiation at the same dose and can thus be identified with any mathematical expression describing the cell survival curve for photons (figure 1(a)).
If photons are replaced by the same dose of heavy ions, the same amount of energy is distributed highly non-uniformly on a microscopic scale, leading to a heterogeneous local dose distribution  . The reason for this are the low-energy and thus short-ranging secondary electrons originating from the Coulomb interaction of the ions within the tissue. Considering a very small and thus uniformly irradiated subvolume,
. The reason for this are the low-energy and thus short-ranging secondary electrons originating from the Coulomb interaction of the ions within the tissue. Considering a very small and thus uniformly irradiated subvolume,  , the average number of lethal events scales with this volume by
, the average number of lethal events scales with this volume by  (Scholz and Kraft 1996). The total number of lethal events in the non-uniformly irradiated nucleus is then obtained by integration over the nucleus volume:
(Scholz and Kraft 1996). The total number of lethal events in the non-uniformly irradiated nucleus is then obtained by integration over the nucleus volume:

For a given fluence pattern of particles in the nucleus, which generates the local dose  , equation (10) gives the average number of lethal events and inserting this number into equation (8) allows the calculation of the survival probability after ion irradiation.
, equation (10) gives the average number of lethal events and inserting this number into equation (8) allows the calculation of the survival probability after ion irradiation.
It has to be noted that the local dose is a stochastic quantity as it is produced by the random fluence pattern of ions and therefore  (equation (8)) is of stochastic nature as well. The average survival probability is therefore obtained by averaging
(equation (8)) is of stochastic nature as well. The average survival probability is therefore obtained by averaging  over many cells using Monte Carlo methods (Scholz et al 1997). Comparing the ion dose, which leads to this average survival probability with the photon dose leading to the same survival probability, allows the calculation of the RBE according to equation (1).
over many cells using Monte Carlo methods (Scholz et al 1997). Comparing the ion dose, which leads to this average survival probability with the photon dose leading to the same survival probability, allows the calculation of the RBE according to equation (1).
For efficient RBE calculations in treatment planning, the initial slope of the survival curves given by the parameter  are pre-calculated for all ion types,
are pre-calculated for all ion types,  , and energies,
, and energies,  , and stored as a table in the treatment planning database (Scholz et al 1997, Krämer and Scholz 2000, 2006). For a spectrum of different ion types and energies, the effective
, and stored as a table in the treatment planning database (Scholz et al 1997, Krämer and Scholz 2000, 2006). For a spectrum of different ion types and energies, the effective  can then be calculated as a dose-weighted average. While these 'initial
can then be calculated as a dose-weighted average. While these 'initial  -values determine the RBE for the limit of very low doses (section 4.2), the increase of the slope of the photon survival curve has to be considered at higher doses. This can be done either by evaluating equation (8) or by applying an approximation for the
-values determine the RBE for the limit of very low doses (section 4.2), the increase of the slope of the photon survival curve has to be considered at higher doses. This can be done either by evaluating equation (8) or by applying an approximation for the  -value (Scholz et al 1997, Krämer and Scholz 2000, 2006).
-value (Scholz et al 1997, Krämer and Scholz 2000, 2006).
4.4.2. Input parameter
For the calculation of the effectiveness of ion irradiations, the LEM uses three types of input data: (i) the size of the cell nucleus as the critical target given by its volume, V, or radius,  ; (ii) the energy and particle type dependent radial dose distribution, which describes the local doses deposition in the nucleus, and (iii) the cell survival curve for photons, which is used to calculate the contribution of the local energy deposition by the ion to the overall biological effect.
; (ii) the energy and particle type dependent radial dose distribution, which describes the local doses deposition in the nucleus, and (iii) the cell survival curve for photons, which is used to calculate the contribution of the local energy deposition by the ion to the overall biological effect.
- (i)Radius of the cell nucleus: The LEM assumes a uniform spatial distribution of the critical target over the volume of the target and neglects spatial or temporal variations in radiosensitivity (Scholz and Kraft 1996, 1992). In addition, no specific assumption is made about the nature of the radiation-induced damage. The radius of the nucleus,
 , is fixed to 5 µm for all clinical applications and to comparable values for in vitro experiments (Scholz et al 1997, Elsässer and Scholz 2007)
, is fixed to 5 µm for all clinical applications and to comparable values for in vitro experiments (Scholz et al 1997, Elsässer and Scholz 2007) - (ii)Radial dose distribution: To calculate the local dose distribution,), an energy and particle type dependent radial dose profile is assumed according to the following parameterization (Scholz and Kraft 1996):
where is the distance from the particle track, and are cut-off parameters, and is a normalization constant, which is adjusted to ensure that the integral over the track cross section reveals the LET at the respective energy. The maximum radius is selected according to the range of the secondary electrons with the highest energy according to , with and (Elsässer and Scholz 2007, Elsässer et al 2008) while is set to 10 nm. A general problem with the parameterization of the radial dose distribution is that experimental data below 0.8 nm are not available and that all data were obtained in gas rather than condensed matter (Elsässer and Scholz 2007).
- (iii)Photon survival curve: To determine the local effect of the energy deposition, the photon survival curve is required. Although the basic version of the LQM (Fowler 1989) may be considered as a convenient description, the local doses deposited by ions may be very high and experimental data suggest that the quadratic shape of the survival curve becomes linear beyond a certain 'transition dose' . is therefore parametrized by Scholz et al (1997):
where is the local dose, and are the cell-specific radiosensitivity parameters of the LQM (Fowler 1989) for photon irradiations, and is the maximum slope occurring at and beyond . While and may in principle be measured in clonogenic survival assays, determination of is generally very difficult as it requires cell survival measurements at very high doses (Elsässer et al 2008).























4.4.3. Dependencies of the RBE
From equation (10), it can be seen that an increased number of lethal events and thus a higher efficiency of ion relative to photon irradiations results only for a shouldered photon survival curve (Scholz 1996). In this case, high local doses will decrease the survival over-proportionally relative to smaller local doses. Vice versa, if the photon survival curve is strictly linear, the integration in equation (10) corresponds to an evaluation of the photon survival probability at the mean dose within the whole nucleus, which corresponds to a uniform local dose distribution and in this case the survival probability will be the same as for a photon irradiation with the same dose. As the extent of the shoulder of the survival curve is described by the ratio  , the LEM predicts an increased RBE for decreasing
, the LEM predicts an increased RBE for decreasing  -values. Generally, the RBE increases with increasing LET and decreasing fractional dose and the extent of these dependencies is governed by the
-values. Generally, the RBE increases with increasing LET and decreasing fractional dose and the extent of these dependencies is governed by the  -value for photons (Scholz 1996).
-value for photons (Scholz 1996).
This indicates that cell lines with a high repair capacity after photon irradiation exhibit a higher RBE in carbon ion beams than repair-deficient cells. As compared to the dependence on  , the RBE has been shown to be much less dependent on absolute value of
, the RBE has been shown to be much less dependent on absolute value of  (Scholz 1996). Finally, the actually selected value for the radius of the nucleus,
(Scholz 1996). Finally, the actually selected value for the radius of the nucleus,  , is important only for very high LET values, where the track diameter becomes much smaller than the nucleus and thus contributes much to the non-uniformity of the local dose (Scholz 1996); for lower LET values the value of
, is important only for very high LET values, where the track diameter becomes much smaller than the nucleus and thus contributes much to the non-uniformity of the local dose (Scholz 1996); for lower LET values the value of  is less important. As the sensitive volume of the nucleus may be smaller than its geometrical volume, the size of the nucleus is considered as an effective volume (Elsässer et al 2008).
is less important. As the sensitive volume of the nucleus may be smaller than its geometrical volume, the size of the nucleus is considered as an effective volume (Elsässer et al 2008).
4.4.4. Further developments
The LEM version as described in the previous paragraphs is termed as LEM I. LEM I has been integrated into treatment planning systems at different carbon ion therapy centers (Krämer and Scholz 2000, Combs et al 2010, Rossi 2015) and by 2015, it had been applied for RBE-weighted dose optimization in more than 3200 patients (www.ptcog.ch). Although LEM I describes the main dependencies of the RBE, some systematic deviations were found in the comparison with the experimental data (section 6). This gave rise to further developments which resulted in the more recent versions LEM II, LEM III, and finally LEM IV. Up to now, however, only LEM I has been used clinically and whether LEM I or LEM IV describes the RBE better is matter of scientific analysis.
Following the initial development of LEM I (Scholz et al 1997), LEM II additionally introduced cluster effects and a modified radial dose profile (Elsässer and Scholz 2007). While LEM I did not make any specific assumption about the type of the damage, LEM II considers double strand breaks (DSBs) of the DNA as the relevant damage for the endpoint cell survival. In contrast, single strand breaks (SSBs) are only considered to be of relevance if two of them are combined into a DSB (SSB cluster damage). This is modeled by Monte Carlo methods assuming a linear genome of a certain size and a random distribution of an experimentally found number of 1250 SSBs/(Gy cell). A DSB will then be formed if two SSBs are separated by less than 25 base pairs. These additional DSBs increase the biological effect, which leads to corrections at higher local doses (Elsässer and Scholz 2007).
As a further modification in LEM II, the radial dose profile of equation (11) was modeled in more detail by considering: (i) the dose profile of the initial energy transfer, which is assumed to increase continuously with decreasing distance from the ion track down to  , and (ii) the diffusion of radiation-induced radicals, which is described by a convolution of the initial profile with a Gaussian distribution using a width of
, and (ii) the diffusion of radiation-induced radicals, which is described by a convolution of the initial profile with a Gaussian distribution using a width of  . As a result, the initial profile is broadened and exhibits higher local doses at distances close to the ion track and lower doses at larger distances (
. As a result, the initial profile is broadened and exhibits higher local doses at distances close to the ion track and lower doses at larger distances ( ) as compared to the parameterization used in LEM I (Elsässer and Scholz 2007). As a further modification, LEM III parameterizes the inner radius of the ion tracks by
) as compared to the parameterization used in LEM I (Elsässer and Scholz 2007). As a further modification, LEM III parameterizes the inner radius of the ion tracks by  , where
, where  is the ion velocity relative to the speed of light and
is the ion velocity relative to the speed of light and  is a constant, which is set to
is a constant, which is set to  (Elsässer et al 2008).
(Elsässer et al 2008).
As the most recent development, LEM IV proposes a generalization of the previous LEM concept assuming that equal local DSB densities rather than equal local doses lead to equal biological effects, independent of the applied radiation quality (Elsässer et al 2010, Friedrich et al 2012). For this, the DSBs are divided into isolated (iDSB) or clustered DSBs (cDSB), respectively, depending on whether exactly one or more than one DSB is produced in a subvolume of the nucleus, which is assumed to correspond to the size of a so-called 'giant loop' of the DNA and which is modeled by a cubic volume with an edge length of  . The complexity of the local damage pattern is then defined by the cluster index
. The complexity of the local damage pattern is then defined by the cluster index  , where
, where  is the number of subvolumes with the respective damage type. The number of iDSB and cDSB is calculated by Monte Carlo methods using an experimentally found number of 30 DSBs/(Gy cell). Irradiations with photons and ions are then considered as isoeffective if the cluster index is the same (Friedrich et al 2012). As photons in comparison to ions distribute their energy uniformly over a sensitive target, the same photon dose will in average distribute the same number of DSBs over a larger volume and the probability of having two or more DSBs (i.e. a cDSB) within a subvolume is therefore reduced. A comparable cluster index for photon irradiations will therefore be obtained at higher doses. This photon dose is termed as equivalent dose and includes the RBE.
is the number of subvolumes with the respective damage type. The number of iDSB and cDSB is calculated by Monte Carlo methods using an experimentally found number of 30 DSBs/(Gy cell). Irradiations with photons and ions are then considered as isoeffective if the cluster index is the same (Friedrich et al 2012). As photons in comparison to ions distribute their energy uniformly over a sensitive target, the same photon dose will in average distribute the same number of DSBs over a larger volume and the probability of having two or more DSBs (i.e. a cDSB) within a subvolume is therefore reduced. A comparable cluster index for photon irradiations will therefore be obtained at higher doses. This photon dose is termed as equivalent dose and includes the RBE.
To speed-up the RBE calculations, the same pre-calculations and approximation as for LEM I can be used (Scholz et al 1997, Krämer and Scholz 2000, 2006). As compared to LEM I, LEM IV exhibits a stronger dependence on LET leading to significantly higher RBE values at the distal edge of the SOBP (Grün et al 2012).
4.5. Microdosimetric kinetic model (MKM)
While the mixed beam model represents a purely phenomenological approach and the LEM assumes a continuous radial profile fixed to the track of the ions, the MKM follows microdosimetric principles. Based on the theory of dual radiation action (TDRA), the number of lethal events in a small volume of the cell nucleus was postulated to be proportional to the square of the specific energy,  (Kellerer and Rossi 1978, Zaider and Rossi 1980). In the development of the MKM, Hawkins (1994) generalized this postulate, assuming a linear-quadratic dependence on
(Kellerer and Rossi 1978, Zaider and Rossi 1980). In the development of the MKM, Hawkins (1994) generalized this postulate, assuming a linear-quadratic dependence on  . In addition, the MKM assumes the cell nucleus to be partitioned into so-called domains of small but finite size (assuming cylindrical volumes with radii
. In addition, the MKM assumes the cell nucleus to be partitioned into so-called domains of small but finite size (assuming cylindrical volumes with radii  and
and  for the nucleus and the domain, respectively) and based on the stochastic properties of the energy deposition events in the domains, the average number
for the nucleus and the domain, respectively) and based on the stochastic properties of the energy deposition events in the domains, the average number  of lethal events after high-LET radiation is derived as
of lethal events after high-LET radiation is derived as

with

In equation (13),  is the delivered dose and
is the delivered dose and  and
and  are the coefficients of the LQM. It is an intrinsic feature of the MKM that
are the coefficients of the LQM. It is an intrinsic feature of the MKM that  is considered as independent of the beam quality while the coefficient
is considered as independent of the beam quality while the coefficient  is composed of
is composed of  referring to the limit of
referring to the limit of  (realized by photon irradiations) and a second term, which is proportional to
(realized by photon irradiations) and a second term, which is proportional to  . The proportionality constant
. The proportionality constant  characterizes the beam quality and is related to the variance of the microscopic energy deposition to the domains as well as to the nucleus as a whole (Hawkins 1994), i.e. the more heterogeneous the microscopic energy deposition is, the larger is the variance and thus also the initial slope of the survival curve. Only in the limit of a completely uniform energy deposition (i.e.
characterizes the beam quality and is related to the variance of the microscopic energy deposition to the domains as well as to the nucleus as a whole (Hawkins 1994), i.e. the more heterogeneous the microscopic energy deposition is, the larger is the variance and thus also the initial slope of the survival curve. Only in the limit of a completely uniform energy deposition (i.e.  ), the response reduces to that of photons.
), the response reduces to that of photons.
In this formulation the MKM leads to a linear increase of  and and thus RBE with LET in the limit of low doses, while experimental data shows a saturation of the RBE at values of around 200 keV µm−1 and a decrease beyond (termed as the 'overkill-effect'). This behavior is attributed to limitations of the applied Poisson approximation and to consider the 'overkill-effect' in the model for monoenergetic beams, the MKM was further developed (Hawkins 2003). To apply the MKM at NIRS for SOBPs generated by superposition of scanned beams with different energies, a different formulation of the saturation correction was introduced and in this version (Kase et al 2006), the MKM was implemented into the clinical treatment planning system (Inaniwa et al 2010). According to this implementation, the average number of lethal events is given by Inaniwa et al (2010):
and and thus RBE with LET in the limit of low doses, while experimental data shows a saturation of the RBE at values of around 200 keV µm−1 and a decrease beyond (termed as the 'overkill-effect'). This behavior is attributed to limitations of the applied Poisson approximation and to consider the 'overkill-effect' in the model for monoenergetic beams, the MKM was further developed (Hawkins 2003). To apply the MKM at NIRS for SOBPs generated by superposition of scanned beams with different energies, a different formulation of the saturation correction was introduced and in this version (Kase et al 2006), the MKM was implemented into the clinical treatment planning system (Inaniwa et al 2010). According to this implementation, the average number of lethal events is given by Inaniwa et al (2010):

with  denoting the saturation-corrected dose mean specific energy of the domain produced by a single event, which is calculated from the respective density distribution,
denoting the saturation-corrected dose mean specific energy of the domain produced by a single event, which is calculated from the respective density distribution,  , of the specific energy:
, of the specific energy:

in which  is given by
is given by

with the characteristic saturation parameter

While equation (14c) reproduces the unsaturated values of  for low values, it maps high
for low values, it maps high  -values to an isoeffective lower value of
-values to an isoeffective lower value of  .
.
For the application of the MKM, the values of  ,
,  ,
,  , and
, and  have to be specified and if these parameters are fixed, the beam quality,
have to be specified and if these parameters are fixed, the beam quality,  , is solely determined by the distribution
, is solely determined by the distribution  of the microscopic quantity
of the microscopic quantity  (equation (14b)). In principle,
(equation (14b)). In principle,  can be directly determined from the distribution of
can be directly determined from the distribution of  , which can be measured by a tissue-equivalent proportional counter (TEPC) (Kase et al 2006). Practically, however,
, which can be measured by a tissue-equivalent proportional counter (TEPC) (Kase et al 2006). Practically, however,  is obtained by averaging the nominator and denominator of equation (14b) over the impact parameter of the ions relative to the center of the domain using the Kiefer–Chatterjee track structure model for
is obtained by averaging the nominator and denominator of equation (14b) over the impact parameter of the ions relative to the center of the domain using the Kiefer–Chatterjee track structure model for  and
and  (Inaniwa et al 2010). To calculate
(Inaniwa et al 2010). To calculate  for the mixed beam situation within the SOBP, the Monte Carlo code Geant4 is additionally used.
for the mixed beam situation within the SOBP, the Monte Carlo code Geant4 is additionally used.
Using this implementation of the MKM, the RBE at a given cell survival fraction,  , can then be calculated by
, can then be calculated by

where  refers to the dose of the reference beam quality and
refers to the dose of the reference beam quality and  contains all microdosimetric properties of the energy deposition by the ions (Inaniwa et al 2010).
contains all microdosimetric properties of the energy deposition by the ions (Inaniwa et al 2010).
The input parameters of the MKM were determined for human salivary gland (HSG) cells using a fixed value of  , which has been measured in 200 kVp x-rays. The values of the other parameter were determined by fitting the predicted doses corresponding to 10% HSG cell survival at different beam qualities. As a result,
, which has been measured in 200 kVp x-rays. The values of the other parameter were determined by fitting the predicted doses corresponding to 10% HSG cell survival at different beam qualities. As a result,  ,
,  and
and  were obtained (Inaniwa et al 2010).
were obtained (Inaniwa et al 2010).
5. Clinical application of RBE models
5.1. Relation between in vitro and clinical data
RBE models are always developed based on theoretical concepts using specific parameters for the radiation response, which themselves depend on the radiation quality of the beam (Hawkins 1994, Kanai et al 1997, Scholz et al 1997, Inaniwa et al 2010). As these response parameters can be measured only in vitro under well-defined conditions, the systematic dependencies of the model are usually benchmarked against cell experiments (Hawkins 1994, Kanai et al 1997, Weyrather et al 1999) (see also section 6.3). While cell inactivation is the endpoint usually considered in cell experiments, it represents only an approximation for the radiation response of complex tissues or tumors because the latter additionally depends on interaction with surrounding cells as well as with environmental factors like nutrition or oxygen supply (Tatum et al 2006, Junttila and de Sauvage 2013, Glowa et al 2016). Whether a clinically relevant radiation reaction will develop also depends on the number and types of killed cells (associated with dose, beam quality, and irradiated volume), the importance of these cells for the superordinated functional subunits as well as the global tissue architecture (parallel versus serial) and different endpoints (e.g. early versus late effects) may compete (Withers et al 1988, Dörr 2009, Joiner 2009). Using cellular parameters for the calculation of the RBE for clinical endpoints therefore introduces additional uncertainties in the quantitative estimation of the tissue response. For this reason, clinical application of RBE models always involves an intermediate step, in which the value of the most important in vitro parameter is replaced by an effective clinical value determined in patients and which corresponds to the clinically relevant biological endpoint (Scholz 1996, Inaniwa et al 2015). In this way, the uncertainty in the prediction of the RBE and thus of the prescribed RBE-weighted dose is reduced. Nevertheless, the RBE predictions may still contain significant uncertainties and dose escalation trials may be necessary to find the optimal prescribed dose. Which model parameter is replaced by its corresponding clinical value and which endpoint is considered depends on the model as well as on the available clinical information.
5.2. Mixed beam model
In the mixed beam model (section 4.3), RBE depth dose profiles were calculated for a set of clinically applied and passively generated SOBPs using HSG cells as the biological system and 10% cell survival as the biological endpoint (Kanai et al 1997). For the transition to clinical application, the following additional information was used (Kanai et al 1999, Gueulette and Wambersie 2007): (i) Experimental evidence indicated that the RBE of carbon ions in a 6 cm SOBP at a dose-averaged LET of 80 keV µm−1 is comparable to that of a previously applied neutron beam. (ii) The clinical RBE of this neutron beam was found to be 3.0 when 18 fractions of 0.9 Gy were applied. For patient treatments, this fractionation schedule was initially applied (Kanai et al 2006) and the RBE profile measured for HSG cells at the 10% survival level was normalized to the neutron-RBE at the position corresponding to a LET of 80 keV µm−1. This fractionation schedule was then subsequently modified within dose escalation and hypofractionation studies (Tsujii et al 2004).
With respect to the terminology used at NIRS, two RBE weighted doses are distinguished: While the biological dose,  describes the RBE-weighted dose for the endpoint 10% HSG cell survival, the clinical dose,
describes the RBE-weighted dose for the endpoint 10% HSG cell survival, the clinical dose,  , refers to the numerical dose value prescribed to patients after normalizing the RBE profile to the clinical neuron RBE.
, refers to the numerical dose value prescribed to patients after normalizing the RBE profile to the clinical neuron RBE.
In the dose prescription system at the NIRS, the clinical dose for a given SOBP can be expressed as follows:

In equation (16),  is the actually applied absorbed dose and
is the actually applied absorbed dose and  is the absorbed dose related to a predefined survival level of HSG cells (here 10%) evaluated either at the center,
is the absorbed dose related to a predefined survival level of HSG cells (here 10%) evaluated either at the center,  , or at any depth,
, or at any depth,  , of the SOBP, respectively.
, of the SOBP, respectively.  is the experimental RBE of the HSG cells at the same survival level. The factor 1.46 scales the biological to the clinical dose (Gueulette and Wambersie 2007, Matsufuji et al 2007).
is the experimental RBE of the HSG cells at the same survival level. The factor 1.46 scales the biological to the clinical dose (Gueulette and Wambersie 2007, Matsufuji et al 2007).
The last two terms of equation (16) describe the relative depth profiles of  and the absorbed dose, respectively. Both profiles are normalized to the respective values at the center of the SOBP. As the passive beam delivery system at the NIRS uses ridge filters to generate the absorbed depth dose profiles from monoenergetic beams (Torikoshi et al 2007), the shapes of these ridge filters are designed in a way that the product of the last two terms in equation (16) equals unity. As a result, a uniform clinical dose within the SOBP is obtained.
and the absorbed dose, respectively. Both profiles are normalized to the respective values at the center of the SOBP. As the passive beam delivery system at the NIRS uses ridge filters to generate the absorbed depth dose profiles from monoenergetic beams (Torikoshi et al 2007), the shapes of these ridge filters are designed in a way that the product of the last two terms in equation (16) equals unity. As a result, a uniform clinical dose within the SOBP is obtained.
It turns out that the shape of the  profile relative to the center of the SOBP is only weakly dependent on the dose level especially for early responding tissues. For patient treatments, the clinical RBE at the center of SOBP will be determined in dose escalation studies within clinical trials.
profile relative to the center of the SOBP is only weakly dependent on the dose level especially for early responding tissues. For patient treatments, the clinical RBE at the center of SOBP will be determined in dose escalation studies within clinical trials.
Strictly speaking, the  profile as well as the
profile as well as the  value at the center of the SOBP would have to be taken at the survival level produced by the absorbed dose. However, as the RBE profile of each SOBP is encoded in the corresponding ridge filter, specific hardware components would be required for each dose level. As this is impractical for clinical application, the dose dependence of the RBE in the mixed beam model is neglected and all RBE values refer to a survival level of 10%, even for strongly hypofractionated treatments.
value at the center of the SOBP would have to be taken at the survival level produced by the absorbed dose. However, as the RBE profile of each SOBP is encoded in the corresponding ridge filter, specific hardware components would be required for each dose level. As this is impractical for clinical application, the dose dependence of the RBE in the mixed beam model is neglected and all RBE values refer to a survival level of 10%, even for strongly hypofractionated treatments.
This has important consequences for the interpretation of the clinical doses prescribed with the mixed beam model: While these doses can be considered as conceptionally isoeffective to photon treatments for the initial fractionation schedule, for which the clinical neutron RBE (which refers to photons) was adopted, this is no longer the case for other fractionation schedules as the dose dependence of the RBE is neglected. Within the framework of the mixed beam model, the clinical dose is therefore treated only as an index for treatment effectiveness and the optimal doses and fractionation schedules are determined in clinical trials (Tsujii and Kamada 2012).
As a further consequence of disregarding the dependence of RBE on dose, only one field per day is applied as the superposition of multiple fields would change the particle spectrum as well as the absorbed dose distribution and thus also the RBE distribution. In the case of treatment plans with multiple beams, the individual beams are therefore delivered on consecutive days.
Finally, it has to be noted that the photon survival curve of the HSG cells shows only a small shoulder corresponding to a rather large  -value. The resulting RBE is therefore considered to be representative for early reacting normal tissues and for tumor response. This is in-line with the finding that RBE for early reactions in mouse skin were comparable for 80 keV µm−1 carbon ions and neutrons for irradiations with four fractions (Kanai et al 1999). This has to be considered when comparing RBE-weighted doses from the mixed beam model with RBE-weighted doses from the LEM, as the latter uses late effects in normal tissues as the biological endpoint (see also section 5.5).
-value. The resulting RBE is therefore considered to be representative for early reacting normal tissues and for tumor response. This is in-line with the finding that RBE for early reactions in mouse skin were comparable for 80 keV µm−1 carbon ions and neutrons for irradiations with four fractions (Kanai et al 1999). This has to be considered when comparing RBE-weighted doses from the mixed beam model with RBE-weighted doses from the LEM, as the latter uses late effects in normal tissues as the biological endpoint (see also section 5.5).
5.3. Local effect model
The LEM (section 4.4) was also primarily developed and tested based on in vitro data (Weyrather et al 1999) and it has been shown that the functional dependencies of the RBE are impacted the most by the  -ratio for photons while it depends less on the absolute value of the radiosensitivity-parameter
-ratio for photons while it depends less on the absolute value of the radiosensitivity-parameter  (Scholz 1996). For clinical application of the LEM, the cellular
(Scholz 1996). For clinical application of the LEM, the cellular  is therefore replaced by its clinical value which originates from comparison of different isoeffective fractionation schedules for photons and the selection of
is therefore replaced by its clinical value which originates from comparison of different isoeffective fractionation schedules for photons and the selection of  then depends on the irradiated tissue as well as on the clinically considered endpoint As the LEM was first applied for treatments of skull-base tumors (Schulz-Ertner et al 2005, 2007a, 2007b), late reactions of the central nervous system (CNS) tissue were considered as the most important endpoint and the
then depends on the irradiated tissue as well as on the clinically considered endpoint As the LEM was first applied for treatments of skull-base tumors (Schulz-Ertner et al 2005, 2007a, 2007b), late reactions of the central nervous system (CNS) tissue were considered as the most important endpoint and the  -ratio was accordingly set to a value of
-ratio was accordingly set to a value of  . As the RBE increases for decreasing
. As the RBE increases for decreasing  , this selection was considered as being conservative with respect to the clinical effectiveness of carbon ions in the CNS.
, this selection was considered as being conservative with respect to the clinical effectiveness of carbon ions in the CNS.
For practical reasons, the value of  has been used also for other tumor sites in almost all patients. As one exception, the
has been used also for other tumor sites in almost all patients. As one exception, the  -values for the treatment of prostate cancer was raised to
-values for the treatment of prostate cancer was raised to  for the tumor and the surrounding normal tissue within a clinical trial, since toxicity (Habl et al 2016) as well as effectiveness (a second manuscript is in preparation) in a preceding trial were lower than expected. In addition, values of
for the tumor and the surrounding normal tissue within a clinical trial, since toxicity (Habl et al 2016) as well as effectiveness (a second manuscript is in preparation) in a preceding trial were lower than expected. In addition, values of  within and
within and  outside the gross target volume (GTV) are currently used in a clinical trial in non-small-cell superior sulcus tumors (Hauswald et al 2015). With the exception of these cases, all patients up to now have been treated using the same
outside the gross target volume (GTV) are currently used in a clinical trial in non-small-cell superior sulcus tumors (Hauswald et al 2015). With the exception of these cases, all patients up to now have been treated using the same  -values for the tumor and the normal tissue.
-values for the tumor and the normal tissue.
In contrast to the Japanese approach with the mixed beam model, clinical application of the LEM considers late effects in normal tissue as the clinically relevant endpoint. As the LEM is applied in combination with the active beam scanning technique, the RBE distribution is not connected to any patient-specific hardware. The dose dependence of the RBE can therefore be easily considered by modulating the delivered absorbed dose distribution. Furthermore, the delivery of multiple fields per fraction is feasible as the LEM calculates the RBE for the local particle spectrum.
5.4. Microdosimetric kinetic model (MKM)
With the introduction of the more flexible beam scanning technique at NIRS in 2011, the MKM has been clinically introduced (Inaniwa et al 2015) and subsequently also replaced the phenomenological mixed beam model at the passive beam lines. The MKM was introduced to consider the dose-dependence of the RBE as well as the dependence of the RBE on the fragment spectrum produced by the primary ions (Inaniwa et al 2015). Both dependencies were not included in the mixed beam model. As an important consequence, the MKM can also be applied to multiple field optimizations in intensity modulated particle therapy (IMPT).
To make use of the extensive clinical experience with previous carbon ion treatments at NIRS delivered with passive beams, the introduction of the MKM has been performed in a way that it maintained the dose prescription system established with the mixed beam model (Inaniwa et al 2015). With the mixed beam model, the photon equivalence of the prescribed RBE-weighted dose for the initially applied fractionation scheme was obtained by normalization of the experimental RBE profile to the previously determined clinical neutron RBE (which itself refers to photon treatments) at the 'neutron-equivalent point' (depth corresponding to 80 keV µm−1, see section 5.2). As the dose dependence of the RBE is not considered in the mixed beam model, photon equivalence is no longer obtained when moving from the initial fractionation to hypofractionated treatments. Practically, this led to increased nominal prescription doses for hypofractionated carbon ion treatments as compared to the expected dose of an isoeffective photon treatment.
For the clinical introduction of the MKM, it was therefore decided to replace the photon reference radiation by a carbon ion reference radiation of a specified beam quality. This beam quality was selected as that at the center of a 6 cm SOBP produced by 350 MeV u−1 carbon ions (reference SOBP). To realize this approach, the biological dose was defined as (Inaniwa et al 2015):

In equation (17), the LQM coefficient  now refers to the carbon ion rather than photon reference radiation. As in the mixed beam model, the model parameters were determined from HSG cell survival curves, which resulted in
now refers to the carbon ion rather than photon reference radiation. As in the mixed beam model, the model parameters were determined from HSG cell survival curves, which resulted in  ,
,  ,
,  , and
, and  (Inaniwa et al 2015).
(Inaniwa et al 2015).
By construction, the biological dose,  , coincides with the absorbed dose,
, coincides with the absorbed dose,  , at all dose levels if
, at all dose levels if  is selected at the center of the reference SOBP (reference point). As
is selected at the center of the reference SOBP (reference point). As  is directly connected to the survival fraction,
is directly connected to the survival fraction,  , a uniform distribution of
, a uniform distribution of  within the SOBP indicates a uniform biological effect. In this case,
within the SOBP indicates a uniform biological effect. In this case,  describes the RBE relative to the reference point and equals unity there. Similarly, equation (17) may be used to optimize the absorbed dose distribution
describes the RBE relative to the reference point and equals unity there. Similarly, equation (17) may be used to optimize the absorbed dose distribution  for any configuration of treatment fields to achieve a uniform biological effect in the SOBP (i.e. the same
for any configuration of treatment fields to achieve a uniform biological effect in the SOBP (i.e. the same  or
or  ), which is the basis for the application of the MKM for multiple fields or IMPT treatments. It is important to note that the MKM conceptionally includes a dose dependence of the slope of the RBE depth dose profile as well as of the biological dose,
), which is the basis for the application of the MKM for multiple fields or IMPT treatments. It is important to note that the MKM conceptionally includes a dose dependence of the slope of the RBE depth dose profile as well as of the biological dose,  .
.
While  is solely used to prescribe a uniform biological effect within the SOBP by optimizing the absorbed dose distribution, a scaling factor is required to connect
is solely used to prescribe a uniform biological effect within the SOBP by optimizing the absorbed dose distribution, a scaling factor is required to connect  to the clinically prescribed dose
to the clinically prescribed dose  . This scaling factor is given by the clinical RBE at the reference point, which was determined to be 2.41 for 10% cell survival of HSG cells in the original dose prescription system with the mixed beam model (Inaniwa et al 2015). This results in
. This scaling factor is given by the clinical RBE at the reference point, which was determined to be 2.41 for 10% cell survival of HSG cells in the original dose prescription system with the mixed beam model (Inaniwa et al 2015). This results in

To be consistent with the mixed beam prescription system, this value is considered to be dose-independent also in the MKM prescription system. This assures that the same nominal prescribed doses lead to the same clinical effect in both systems. While the MKM approach now includes the dose dependence of the slope of the RBE profile in the SOBP, it does not consider the dose dependence of the clinical RBE and the latter is still included in the values of the clinically prescribed doses. This results from the dose-independent scaling factor of 2.41, which refers to 10% HSG cell survival, and the fact that different prescribed clinical doses,  , lead to different values of values of
, lead to different values of values of  (equation (18)) and thus to a different HSG cell survival,
(equation (18)) and thus to a different HSG cell survival,  , which in turn leads to a different absorbed dose profile
, which in turn leads to a different absorbed dose profile  (equation (17)).
(equation (17)).
5.5. Transfer of RBE-weighted doses between different centers
Most of the patients treated with carbon ion have been treated either at Japanese centers using the passive beam delivery technique in combination with the mixed beam model or at European centers using the beam scanning method in combination with the local effect model. In 2011, beam scanning was also introduced at NIRS in combination with the MKM and the MKM subsequently replaced the mixed beam model at the passive beam lines at NIRS, however, the MKM is still used with the same dose prescription system as it was the case with the mixed beam model.
As the calculation of the RBE with the different models is based on the consideration of different biological endpoints (early effects in normal and tumor tissues versus late effects in normal tissues) and since the dose dependence is considered only with the LEM but not with the mixed beam model, transfer of dose prescriptions and comparison of clinical results are very difficult. The underlying reason is that different RBE predictions by the two models cause different RBE-weighed doses for identical absorbed dose distributions and identical RBE-weighted dose distributions will be related to different absorbed dose distributions and thus to a different biological effectiveness. Therefore, prescribed doses in units of Gy (RBE) of centers using different RBE models cannot be considered as isoeffective, which is in contrast to photon radiotherapy.
As a consequence, these centers have developed their own clinical strategies in optimizing therapy results. These strategies include dose escalation studies as well as the transfer and adaption of prescribed doses between different tumor indications based on the clinical results of their own center or other centers using the same RBE model. A clear disadvantage of this approach is that results of large patient data sets of centers using a different RBE model are not accessible for treatment optimization.
To circumvent this problem, a simulation approach has been followed by several authors (Fossati et al 2012, Steinsträter et al 2012, Molinelli et al 2016). The basic idea is that identical absorbed dose distributions delivered at different centers will produce the same clinical effect, regardless of the applied RBE model. This is considered as a reasonable assumption, although it neglects that active and passive beam delivery systems may produce different secondary particles spectra with somewhat different biological effectiveness. However, it has been experimentally shown that these differences are rather small if the same absorbed dose is applied (Uzawa et al 2009).
As an example, Steinsträter et al (2012) used their LEM-based treatment planning system (TPS) to reconstruct the absorbed dose distributions of the SOBPs at the NIRS and to recalculate the RBE-weighted dose distribution with LEM instead of the mixed beam model based on the connected carbon ion fluence patterns. Comparing the RBE-weighted dose distributions of the two treatment plans then allows the determination of conversion factors for the RBE-weighted dose between the two centers. It has to be noted, however, that the recalculated RBE-weighted dose distribution will usually not be uniform in the target volume due to the different functional dependencies in both RBE models. Therefore, representative RBE-weighted doses have to be used as input for the conversion factors. Furthermore, the conversion factors will depend on the selected model parameters as well as on the considered dose level.
As the reproduction of the absorbed dose distribution may be achieved only with a research TPS (Steinsträter et al 2012) or by a Monte Carlo system with an integrated RBE model (Mairani et al 2010), a slightly different approach has been followed using a commercial TPS. Fossati et al (2012) used their LEM-based TPS to generate RBE-weighted SOBPs with the same extensions as those applied clinically at the NIRS. By using different RBE-weighted dose levels and by searching for the best agreement of the underlying absorbed dose distributions, isoeffective SOBPs and the corresponding conversion factors were determined for a set of geometrical target volumes and clinical treatment scenarios. Recently, the conversion factors were confirmed also for clinical cases (Molinelli et al 2016).
The main difference between the two approaches is that in latter case, both RBE-weighted SOBPs are uniform within the target volume and that differences in the RBE models are now reflected in the underlying absorbed dose distribution. Although both methods allow the transfer of clinical results between different centers, they have to be accompanied by careful clinical dose selection strategies.
Disregarding the different biological endpoints used for clinical application, the mixed beam model and the LEM lead to comparable shapes of the absorbed dose distributions for uniform RBE-weighted SOBPs for standard as well as for hypofractionated treatments (Fossati et al 2012, Steinsträter et al 2012). The problem of transferring clinical results from one center to another can therefore be practically solved by the above described conversion of prescription doses.
6. Validation of RBE models
6.1. Uncertainties of model-based RBE values
From the previous sections, it is obvious that model-based RBE predictions are associated with significant uncertainties. These uncertainties may be attributed to two different sources (figure 2): (i) errors in the RBE distribution and (ii) errors in the absolute RBE value (specified at a reference point within the patient). Errors in the RBE distribution may compromise the delivery of a biologically uniform dose to the tumor and moreover may jeopardize therapeutic success by under- or over-dosage of tumor or normal tissue, respectively. The RBE distribution can be measured for specific experimental settings in preclinical studies in vitro as well as in vivo. The results of these measurements can then be used to validate the predicted dependencies in the RBE model.

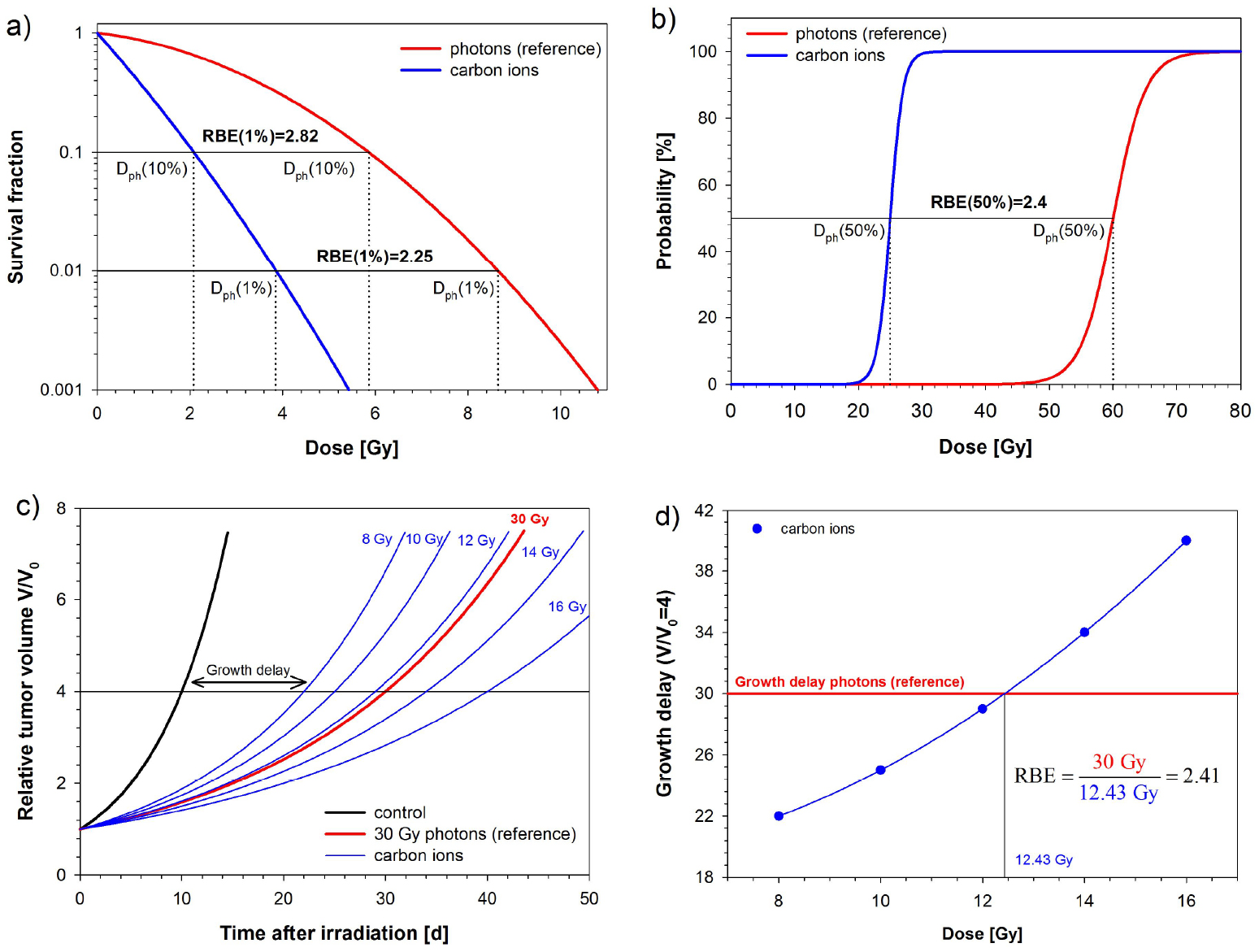{kind=link}
Figure 2. Schematic representation of the uncertainties of model-based RBE values. The RBE uncertainties may be separated in those of the absolute RBE and the RBE distribution. The RBE distribution may be validated only experimentally and is an intrinsic part of the RBE-weighted dose distribution delivered to the patient. Although significant uncertainties may be included in the absolute RBE in patient treatments, they may be compensated to some extent by the selection of the prescribed dose within a clinical dose finding approach.
Download figure:
Standard image High-resolution image{kind=link}
However, even if a perfect agreement between measured and calculated RBE is obtained in such experiments, the experimental RBE will very likely differ from that in patients. Although this difference may lead to under- or over-dosage of tumor and normal tissue, respectively, it may at least partly be compensated by the selection of the prescribed dose within a clinical approach. Therefore, only the combined effect of absolute RBE and dose prescription can be validated in clinical trials.
As illustrated in figure 2, both the absolute RBE as well as the RBE distribution depend on the selected model parameters or in the case of empirical RBE models, on experimental and clinical input data. By changing these input data, the accuracy of the RBE prediction may in principle be iteratively improved. However, due to the limited information, this is a very difficult process and whether adaptations are made will always be a clinical decision.
As an example, the clinical  -value in the LEM affects the absolute RBE as well as the RBE distribution. While the latter aims at delivering a uniform RBE-weighted dose distribution, which may be verified by dedicated preclinical experiments, the absolute RBE determines the effectiveness and safety of the treatment, which can only be validated in clinical trials. Adapting the model parameter
-value in the LEM affects the absolute RBE as well as the RBE distribution. While the latter aims at delivering a uniform RBE-weighted dose distribution, which may be verified by dedicated preclinical experiments, the absolute RBE determines the effectiveness and safety of the treatment, which can only be validated in clinical trials. Adapting the model parameter  , as has been done for prostate cancer treatments (see section 5.3), will modify the predictions of both the absolute RBE as well as the RBE distribution. In the case of the mixed beam model, the RBE distribution is based on cell-survival measurements with HSG cells and the absolute RBE is explicitly defined by introducing a clinical RBE from neutrons at a 'neutron-equivalent point' of the SOBP (see section 5.2).
, as has been done for prostate cancer treatments (see section 5.3), will modify the predictions of both the absolute RBE as well as the RBE distribution. In the case of the mixed beam model, the RBE distribution is based on cell-survival measurements with HSG cells and the absolute RBE is explicitly defined by introducing a clinical RBE from neutrons at a 'neutron-equivalent point' of the SOBP (see section 5.2).
6.2. Normal tissues versus tumors
Validation of RBE models involves the comparison of calculated and experimentally obtained RBE values. The focus of these studies is to test the LET dependence at different dose levels for different cell lines characterized by their radiosensitivity parameters  and
and  . While the functional dependencies of the RBE can be well investigated in this way, it cannot be expected that the differential RBE between normal and tumor cells will reflect the in vivo situation as the response of experimental tumors is strongly influenced by inter-cell and environmental factors. The impact of these factors can be studied only in vivo. However, as inter-cell and environmental factors are not considered by any RBE model, quantitative RBE validation studies are performed only in normal tissues.
. While the functional dependencies of the RBE can be well investigated in this way, it cannot be expected that the differential RBE between normal and tumor cells will reflect the in vivo situation as the response of experimental tumors is strongly influenced by inter-cell and environmental factors. The impact of these factors can be studied only in vivo. However, as inter-cell and environmental factors are not considered by any RBE model, quantitative RBE validation studies are performed only in normal tissues.
6.3. In vitro validation
In vitro validation of RBE models may be performed by comparing predicted and measured RBE values directly or by comparing the underlying survival fractions after ion beams irradiations. In both cases the survival curve parameters for photons are required as input parameters for the calculation. As compared to survival data, presenting the result in direct comparison of measured and calculated RBE values has the advantage that the clinical relevance of deviations in terms of dose is easier to assess.
The RBE prediction of the mixed beam model essentially relies on the calculation of the dose-averaged parameters  and
and  of the cell survival curve for the mixed beam situation from the respective parameters for monoenergetic beams (section 4.3). This approach has been validated for survival curves of V79 cells using differently weighted combined irradiations with monoenergetic helium and carbon ion beams (Kanai et al 1997). For the established clinical setting of the model, the shape of a 6 cm RBE-weighted SOBP was reproduced by the model and a flat biological response in the target region was predicted for different cell lines (Kanai et al 1999).
of the cell survival curve for the mixed beam situation from the respective parameters for monoenergetic beams (section 4.3). This approach has been validated for survival curves of V79 cells using differently weighted combined irradiations with monoenergetic helium and carbon ion beams (Kanai et al 1997). For the established clinical setting of the model, the shape of a 6 cm RBE-weighted SOBP was reproduced by the model and a flat biological response in the target region was predicted for different cell lines (Kanai et al 1999).
Most systematic validation studies for RBE models have been performed for the LEM using dedicated experiments with V79, CHO-K1, and XRS5 cells. In early studies (Scholz et al 1997, Weyrather et al 1999) measured RBE, and survival fractions of carbon ions were used to benchmark the predictions of the clinically applied LEM I at different dose levels (corresponding to survival fractions of 1%, 10%, and 50%) and a good agreement was found over a wide LET range. Based on benchmarking against in vivo data (section 6.4), the LEM was further developed and additional comparisons in terms of RBE or cell survival fraction were made for the different LEM versions (Elsässer and Scholz 2007, Elsässer et al 2008, Elsässer et al 2010, Friedrich et al 2012). Some of these studies were also extended to other cell lines (Elsässer et al 2008) and ion types (Elsässer and Scholz 2007, Elsässer et al 2010). To simulate a clinical scenario, LEM I and LEM II were also tested against cell survival experiments in 3D phantoms using a u-shaped target together with a multi-field optimization of two opposing SOBPs (Gemmel et al 2008). In a similar setting, LEM IV was tested in an opposing-beam geometry using single field optimization (Elsässer et al 2010). In both cases, the results of these studies were considered to support the new developments in the more recent LEM versions.
An initial validation of the MKM has been performed comparing predicted survival curves with published data on V79 cells irradiated with several monoenergetic proton, deuteron, and helium beams (Hawkins 1998). Later, direct comparisons of the predicted RBE with literature data have also been performed for the LET dependence of the RBE including a saturation correction (Hawkins 2003). At the NIRS, dedicated experiments with HSG cells were performed and the model parameters were determined for the MKM with a modified saturation correction (Kase et al 2006). With these parameters, the predicted and measured dependence of the α-value on the microdosimetric quantities was tested for the HSG cells. For this, irradiations were performed with a variety of ions (protons to iron) using either a monoenergetic Bragg peak or a 6 cm SOBP and a good accuracy was found up to 450 keV µm−1. When the MKM was integrated into the TPS at the NIRS, further benchmarking of the LET dependence of the MKM predictions in terms of survival was performed for a carbon ion SOBP and a good agreement with measurements was found (Inaniwa et al 2010).
6.4. In vivo validation
Although quite a large number of RBE measurements have been performed in vivo, only a very few dedicated experiments have been performed to benchmark predictions of clinically applied RBE models. The first in vivo experiment to validate the procedures for carbon ion treatment planning at the Helmholtz Centre for Heavy Ion Research (GSI) in pig lungs as a model for normal tissue effects revealed the problems associated with dose planning when complex moving structures are included in the beam channel (Dörr et al 1999).
For the mixed beam model, it has been shown that irradiations with an SOBP, which has been designed based on a 10% survival fraction of V79 cells also leads to a flat response of crypt cells in the SOBP after in vivo irradiations of the mouse intestine (Fukutsu et al 1997). In this study, only the shape of the RBE profile was validated in accordance with the concept of the mixed beam model to normalize the relative RBE profile to the clinical RBE obtained from fast neutrons (section 5.2).
Absolute comparisons of measured and calculated RBE values have only been performed for the LEM. Here, a series of dose-response studies were performed using the rat spinal cord in combination with the endpoint myelopathy °II within 300 d as a model for late normal tissue reactions. The spinal cord model was introduced decades ago (van der Kogel and Barendsen 1974) and has been applied in many low- and high-LET radiation response studies. Regarding RBE measurements, the spinal cord has the advantage of having a very small diameter, which allows the measurement of the response at a well-defined position within an SOBP. It is therefore an ideal model to measure the functional dependence on dose and LET.
A first series of experiments with 1, 2, 6, and 18 fractions was performed with the spinal cord positioned in the plateau region of a monoenergetic Bragg peak and at the center of a 1 cm SOBP, respectively, Debus et al 2003, Karger et al 2006). Using 15 MV photons from a linear accelerator, the RBE was determined from the ratio of the tolerance doses TD50 (dose at 50% probability of myelopathy °II) of photons and carbon ions. It was found that LEM I underestimates the RBE in the SOBP by about 25% over the whole dose range while in the plateau, it overestimates the RBE for low doses and underestimates it for high dose per fraction. Especially, the dose dependence predicted by the model for the plateau was not seen in the experimental data. These discrepancies gave rise to further developments of the LEM (section 4.4.4) and for LEM III and IV, a much better agreement was found for the SOBP irradiations at high-LET values as compared to LEM I and II (Elsässer et al 2008, Grün et al 2012), For the plateau region, however, systematic deviations were still observed.
To assess the LET dependence of the RBE in more detail, a large series of experiments was performed at six different positions of a 6 cm SOBP covering the dose-averaged LET range of 16–99 keV µm−1 and using 1, 2, or 6 fractions (Saager et al 2014, 2015, 2016). As a result of the 1 and 2 fraction experiments, a linear increase of RBE with LET was found, and the increase was higher for lower fractional doses (1.28–2.30 versus 1.26–1.83). Comparisons with predictions of LEM I and IV revealed a better agreement of LEM IV in the high-LET region while LEM I was more accurate at low-LET values and the agreement generally was dependent on the dose level. While LEM IV predicts RBE values near 1.0 at very low LET values, the experimental data still show increased values around 1.27 for both dose levels.
In these comparisons, LEM I calculations have been performed with the standard input parameter used also for patient treatments. It has to be noted that calculations with LEM IV do not only include modifications of the model but also modified model parameters, which have been selected to improve the description of the experimental data. Although various different sets of input data have been used to describe in vitro experiments, the in vivo experiments resulted in two standard input data sets for LEM I and IV, respectively, both using  (Grün et al 2012).
(Grün et al 2012).
6.5. Clinical validation
Direct validation of RBE models in patients is in general very difficult and limited by the fact that the absorbed dose distribution and the underlying particle spectrum depends strongly on the individual patient parameters, such as beam configuration, target volume or dose prescription. In addition, any observed endpoint represents an integral response of the irradiated organ or tumor, influenced by the individual radiosensitivity, and cannot be assigned to a well-defined absorbed dose or LET. Finally, a comparable patient collective treated with photons is often not available. Therefore, the RBE values estimated in patient collectives are associated with large uncertainties.
As an example, the clinical RBE was estimated in non-small cell lung cancer patients treated either with photons or carbon ions using a model-based approach (Kanai et al 2006). In this analysis, several assumptions and simplifications had to be made and the RBE of 1.75 at the TCP level of 50% agreed well with the value of the HSG cells of 1.63, but was lower than the clinical RBE of 2.38 resulting from the applied dose prescription system. At TCP levels of 80% and 90%, however, the RBE values of 1.94 and 2.14, respectively, were found to be closer to the clinical RBE. In the respective dose range, the clinically obtained local control rates after carbon ion and photon therapy were found to be comparable if the nominal prescribed dose for carbon ions calculated by the mixed beam model in terms of Gy (RBE) was the same as the prescribed photon dose expressed in Gy (Kanai et al 2006).
Using the estimated model parameters for the photon collective of this study as input, the dose-response curve for non-small cell lung cancer treatments was recalculated with the LEM as a function of the absorbed dose (Scholz et al 2006). Comparison with clinical data from NIRS revealed a ~10% higher dose for the model calculation.
As an alternative to a direct determination of the RBE in patients, it can be investigated, whether the prescribed RBE-weighted dose leads to the same effectiveness or toxicity as a photon treatment with the same dose. A confirmation would then support the RBE model including the selected parameters. However, if there is a disagreement, the prescribed dose or the value of dose-influencing model parameters rather than the model itself will be changed as no information on the specific weakness of the RBE model is gained. This pragmatic clinical approach has been followed when adjusting the  -ratio for prostate cancer treatments (see section 5.3).
-ratio for prostate cancer treatments (see section 5.3).
As a further example, 5y-local control rates of skull-base chordoma treated at different centers with photons, protons, or carbon ions could be described by a common dose-response curve using the prescribed RBE-weighted dose as a dosimetric variable (Schulz-Ertner et al 2007a). This finding supported the assumption that the RBE for the carbon ion treatment is not fundamentally wrong. However, as in the study of Kanai et al (2006), a limitation is that the patient characteristics differed between the centers and treatment modalities.
In another study, temporal lobe reactions in MRIs of patients after the treatment of skull base chordoma and chondrosarcoma were analyzed and a steep dose-response curve was established as a function of the LEM I-based RBE-weighted maximum dose in the temporal lobe, excluding the 1 cm3 volume with the highest dose (Schlampp et al 2010). Based on the absorbed dose distributions of the same patient collective, the calculation of the RBE-weighted dose distribution was repeated using LEM IV instead of LEM I and a second dose-response curve was derived (Gillmann et al 2014). Comparing both curves, a clinically significant difference of 9.5 Gy (RBE) was found for TD5. In principle, a comparison of these response curves with a corresponding curve for photon treatments would allow determination of whether LEM I or LEM IV describes the RBE more accurately. Unfortunately, no such dose-response curves for photons are available and also the TD5-value published by the QUANTEC analysis did not allow for a decision due to the large confidence intervals originating from the analysis of prescribed rather than local doses (Lawrence et al 2010). Ongoing analysis attempts to establish a low-LET curve by analyzing patient data after proton therapy.
In general, studies in patients may be used to investigate the consistency of RBE predictions with clinical results, but they are not able to detect specific weaknesses of the model in the functional dependence on physical and biological parameters. For this, dedicated in vitro and in vivo studies are required.
7. Summary and conclusion
7.1. Clinical value and limitations of the RBE concept
Carbon ion therapy has now been applied for two decades and using the RBE has proven to be a feasible and safe concept to consider the increased effectiveness of carbon ions relative to photons (Kamada et al 2015). Currently three RBE models are applied in patients and the foundation of these models range from relatively simple empirical relations (mixed beam model) to more complex models, which make use of advanced theoretical concepts (LEM and MKM). With all three models, the absorbed carbon ion dose distribution is transformed into a RBE-weighted dose distribution; however, although the use of the RBE conceptionally claims isoeffectiveness to a photon treatment, this is not always true for all cases or in all aspects for at least the following reasons:
- (i)Isoeffectiveness always refers to a specific biological or clinical endpoint and even in the best case of fully validated RBE models, isoeffectiveness may be expected only for the selected, but not for other, competing endpoints.
- (ii)Known dependencies on treatment parameters, which are not or only partially reflected by the RBE model will lead to additional uncertainties. Most important with this respect is the dose dependence, which is handled differently in all three models.
- (iii)The accuracy of RBE predictions of the models is associated with uncertainties. These are related to the selection of adequate model parameters as well as to the intrinsic capability of the model to describe the functional dependence on the most important physical and biological parameters.
- (iv)Finally, the RBE is affected by many biological factors, which are not included in any model and only some of them are known. This is especially relevant for tumors and potential candidates are all factors that affect the response to carbon ions and photons differently. Even if these tumor-specific factors are known, they may be heterogeneously distributed within the tumor and between tumors of different patients, even for the same tumor type, and this distribution may furthermore undergo temporal and/or spatial changes.
7.2. Clinical application of RBE models
It is important to understand that clinical carbon ion treatments in Japan and Europe are performed using different RBE models in combination with different biological endpoints and the models additionally treat the dose dependence in a different way. While European Centers use the LEM I and late normal tissue effects as the biological endpoint (Scholz et al 1997), the RBE in Japan is based on the radiation response of HSG cells, which are considered as representative of early responding tissues and tumors (Kanai et al 1999). Furthermore, the RBE profiles of the mixed beam model were initially normalized to the clinical neutron RBE of 3.0, which was obtained for a specific fractionation scheme, and this clinical RBE was then maintained also for all other fractionation schedules. This indicates that photon equivalence can be expected only for the initial fractionation schedule, but not for modified fractionations. As a clinically highly relevant consequence, identical prescription doses in terms of Gy (RBE) at European and Japanese centers will in general not lead to equivalent clinical results. This significantly complicates the transfer of results from clinical trials between these centers (for further discussion, see also Fossati et al (2018)). Although, the dose dependence is now considered to some extent by the clinical application of the MKM in Japan, the dose prescription system is still the same as for the mixed beam model. From the Japanese perspective this was necessary to maintain consistency with previous dose prescriptions (Inaniwa et al 2015).
The significant impact of the applied RBE model, its input parameters as well as the considered biological endpoint on treatment effectiveness also poses requirements on treatment reporting. This issue will also be addressed by an ICRU report for heavy ions, which is currently in preparation.
It has to be noted that even though clinical application of each model involves a transition step from in vitro to in vivo parameters (see section 5), dose prescription is not solely based on the calculated RBE. Rather, clinical application always includes a clinical dose finding process, which is based on judgement of clinical effects and which at least partially compensates for model inaccuracies.
7.3. Open clinical issues related to the RBE
The independent beneficial role of carbon ions or other high-LET ions in radiotherapy essentially depends on the question of whether the RBE for clonogenic tumor cell inactivation (in the given tumor microenvironment) is higher to a clinically relevant extent than the RBE for side effects in the surrounding normal tissue. Part of the required RBE increase is realized by the increase of the LET towards the distal edge of the SOBP. This increase, however, leads to the highest LET at therapeutic doses in the safety margin around the tumor. Whether this is critical for the included normal tissue depends on the volume of the safety margin and thus on the geometrical accuracy of dose delivery on the one side, but also on the biological properties of the involved tissues on the other side.
Preclinical data suggests that RBE is high for tumors with considerable resistance to photons and for normal tissues possessing a substantial sparing effect. In accordance with this finding, RBE models predict the highest differential RBE for low  -values in the tumor and high
-values in the tumor and high  -values in the normal tissue (Weyrather and Debus 2003), where the high
-values in the normal tissue (Weyrather and Debus 2003), where the high  -values refer to acute radiation effects. As acute and late radiation effects may occur competingly, tumor candidates for carbon ion therapy should have a comparable or lower
-values refer to acute radiation effects. As acute and late radiation effects may occur competingly, tumor candidates for carbon ion therapy should have a comparable or lower  -value in the tumor than in the surrounding normal tissue. Although the RBE-weighted dose distribution may be calculated for this scenario, clinical studies rather than treatment plan comparisons are required to prove clinical superiority of carbon ions over protons or photons. With this respect, there are two ongoing randomized Phase III trials comparing the effectiveness and side effects of carbon ions and proton treatments in skull base chordoma and chondrosarcoma (Nikoghosyan et al 2010a, 2010b). Many other trials are investigating the effectiveness and safety of carbon ions for other tumor localizations and combined treatments with photons, protons, or additive chemotherapy without specifically looking at the comparison of high- and low-LET radiation (for an overview of ongoing clinical studies, see Combs and Debus (2013), Tsujii and Kamada (2012), Kamada et al (2015), Fossati et al (2018)).
-value in the tumor than in the surrounding normal tissue. Although the RBE-weighted dose distribution may be calculated for this scenario, clinical studies rather than treatment plan comparisons are required to prove clinical superiority of carbon ions over protons or photons. With this respect, there are two ongoing randomized Phase III trials comparing the effectiveness and side effects of carbon ions and proton treatments in skull base chordoma and chondrosarcoma (Nikoghosyan et al 2010a, 2010b). Many other trials are investigating the effectiveness and safety of carbon ions for other tumor localizations and combined treatments with photons, protons, or additive chemotherapy without specifically looking at the comparison of high- and low-LET radiation (for an overview of ongoing clinical studies, see Combs and Debus (2013), Tsujii and Kamada (2012), Kamada et al (2015), Fossati et al (2018)).
Besides investigating different tumor entities and different treatment modalities, the therapeutic ratio of hypofractionated treatments is being investigated and very encouraging results have been obtained (Tsujii et al 2007, Tsujii and Kamada 2012, Kamada et al 2015). While these hypofractionated treatment concepts were developed mainly based on clinical experience and trials, there are meanwhile also preclinical studies that show that the increase of the RBE with fraction number is larger for normal tissue than for tumors (Koike et al 2002, Karger et al 2006, Karger et al 2013). Although this seems to suggest a radiobiological rationale for hypofractionation, the underlying reasons for this behavior are not fully understood.
In conventional low-LET radiotherapy, the application of low doses per fraction over an extended time period allows for repair and repopulation and maximizes normal tissue sparing, while for hypofractionation with particles two presumably closely associated mechanisms come into consideration: Firstly, the existence of characteristic structural-functional effects, which act differentially with respect to the radiation modality, and secondly, the recent finding that high doses per fraction per se induces severe vascular damage, which deteriorate the intratumor microenvironment by not only causing vast amounts of additional tumor cell death but also by triggering anti-tumor immunity and thereby suppressing recurrence and metastatic tumor growth (Song et al 2014). Both mechanisms are not reflected in any RBE models.
Integration of heterogeneously distributed biological tumor properties into radiotherapy, a heavily discussed issue in conventional photon therapy (Søvik et al 2010), is also of high relevance for carbon ion therapy. The most prominent example for such factors is hypoxia, but also quiescent or other resistant cell subpopulation may be mentioned. In photon therapy, concepts of biologically adapted radiotherapy include an increase of the local dose in these potentially resistant regions of the tumor, which is also termed as 'dose painting' (Thorwarth et al 2007, Kim and Tomé 2010). While the 'dose painting' capabilities for scanned carbon ion beams are even better than for photons, they additionally offer an increased LET. As there is evidence from preclinical studies that the response to high-LET radiation is much less dependent on inter- or intra-tumor heterogeneity (Glowa et al 2016), concentration of the high-LET components of the dose distribution to the most resistant regions of the tumor is considered as a promising approach. In analogy to 'dose painting', this modulation of the LET distribution is termed as 'LET painting' and its technical feasibility has been demonstrated in planning studies by combining high- and low-LET radiation modalities (Bassler et al 2014). In spite of its technical feasibility, clinical application of LET painting is not yet realized because of the lack of reliable quantitative input data from imaging on the spatial-temporal distribution of the parameters of interest. With this respect, further research and development of functional imaging technology and adequate interpretation of the imaging signals for the use in both ion beam as well as photon radiotherapy is necessary.
7.4. How can radiobiological studies contribute to improving carbon ion RT?
From the aforementioned sections, it is clear that a definitive proof of the feasibility, safety, and effectiveness of carbon ion treatments can only be obtained from carefully designed clinical trials. Nevertheless, preclinical radiobiological studies can provide important information for the application and further development of carbon ion therapy. The direction of this radiobiological research covers especially the following fields: (i) quantitative investigation of normal tissue response, (ii) identification of tumor-specific factors and quantitative assessment of their impact on tumor response, and (iii) deciphering of the underlying physiological and molecular mechanisms and identification of predictive factors for therapy outcome. All these investigations have to compare the responses to carbon ions and photons to identify differential effects, which would then directly impact the RBE of carbon ions.
Normal tissues exhibit a rather low heterogeneity within or between individual subjects. Therefore, they are well-suited to experimentally validate the dependence of RBE model predictions on the most important physical and biological parameters. Although this validation can be done also in cell culture to some extent, in vivo measurements are more relevant as they reflect the complexity of both, the structure and function of biological systems, and are thus located closer to the situations of patients. With this respect, the largest quantitative database results from irradiations of the rat spinal cord, a model, which has the advantage of measuring the response to a local particle spectrum (see table 1). Only few data are available for other late reacting normal tissues and research in this direction should be extended by establishing additional normal tissue models with clearly detectable biological endpoints.
It has to be noted that experimentally determined RBE values can never be directly transferred to patients as they always refer to the specific experimental conditions. As a consequence, RBE models can also be validated only for these specific experimental conditions. The underlying assumption is that a validated model then accurately calculates the RBE also for other experimental or clinical conditions.
In comparison to normal tissues, tumors exhibit a much larger heterogeneity between and even within individual subjects. This is due to tumor-associated factors (e.g. intrinsic and microenvironmental resistant factors), which often vary spatially as well as temporally. As these factors are not included in RBE models, comparison of experimental and calculated RBE is not the primary focus. Rather, the identification of these factors as well as the assessment of their impact on the RBE is of vital importance because it can help to select suitable tumors for carbon ion therapy and to stratify patients into different treatment concepts. Currently, quantitative studies are available only for a limited number of tumor types and fractionation schedules (table 2) and hence, it is of highest relevance to further extend such studies.
Besides comparisons between tumors or tumor sublines, another important line of research is the quantitative comparison of the tumor and normal tissue response to investigate the differential RBE between both tissues. Although some information may be gained by comparing the results of existing tumor and normal tissue studies, a limitation is that the combination of model systems and their radiation sensitivity does not reflect the clinical situation. Currently, there is only one single fraction study, which determines the RBE of carbon ions simultaneously for tumor as well as for acute and late normal tissue effects in the same animal model (Sørensen et al 2015). No such studies are available for fractionated irradiations. Fractionated studies, however, would be highly important as fractionation alters the RBE differently for normal tissues and tumors. Comparing the RBEs in the same animal model would thus allow for the identification of the fractionation schedule with the highest differential RBE for which the maximum therapeutic benefit for the patient is expected. As the RBE of tumors and its fractionation dependence may be influenced by a number of biological factors (Glowa et al 2016), which are not included in RBE models, a reliable model-based prediction of the differential RBE is currently not possible. From the experimental side, a further limitation is the lack of suitable normal tissue models with clearly detectable biological endpoints, especially for late radiation effects. Disregarding these difficulties, the investigation of the differential RBE between normal tissues and tumors can help to experimentally prove the independent role of the high-LET effect in carbon ion therapy by demonstrating the existence of a beneficial differential RBE between tumor and normal tissue. This may then be the starting point for the design of corresponding clinical trials.
Finally, it is important to identify differences in physiological and molecular response parameters. This may help to identify clinical situations, where carbon ion treatments can be expected to be superior over photon irradiations, to optimize carbon ion treatment schedules as well as to identify potential target structures for combined therapeutic treatment modalities. With this respect, the ultimate goal is to find predictive parameters for treatment outcome and thereby to increase the confidence of the assignment of patients to a treatment with carbon ions, photons, or a combination of both, eventually including supportive drugs.
7.5. Conclusion
The RBE has proven to be a valuable concept and it is considered as state-of-the-art for clinical practice in carbon ion therapy. In treatment planning, the RBE has to be calculated by biomathematical models, which—in spite of all validation efforts—still involve significant sources of uncertainty. Therefore, use of these models is always connected to a clinical dose prescription approach, which allows for safe and effective treatments. Although different models are used, this approach has been successfully implemented in Japanese and European centers, however, prescription doses cannot be directly exchanged between these centers and therefore, great care has to be taken when transferring results from clinical studies. In parallel to clinical trials, preclinical radiobiological studies are an integral part in the process of validation, optimization, and further development of clinical carbon ion therapy.
Acknowledgment
This study was supported by the German Research Foundation (DFG, KFO 214, KA 2679/3-1, GL 893/1-1) and by the German Cancer Aid (111434).