
Bone Regeneration and Oxidative Stress: An Updated Overview
by
 , and
, and
Adrian Emilian Bădilă
1,2,†, 
Dragos Mihai Rădulescu
1,2,†,
Andrei Ilie
3,
Adelina-Gabriela Niculescu
4 ,
,
Alexandru Mihai Grumezescu
4,5,6,* and
and
Adrian Radu Rădulescu
1,2 1
“Carol Davila” University of Medicine and Pharmacy, 050474 Bucharest, Romania
2
Department of Orthopedics and Traumatology, Bucharest University Hospital, 050098 Bucharest, Romania
3
Faculty of Engineering in Foreign Languages, University Politehnica of Bucharest, 011061 Bucharest, Romania
4
Faculty of Applied Chemistry and Materials Science, University Politehnica of Bucharest, 060042 Bucharest, Romania
5
Research Institute of the University of Bucharest—ICUB, University of Bucharest, 050657 Bucharest, Romania
6
Academy of Romanian Scientists, 3 Ilfov Street, 050044 Bucharest, Romania
*
Author to whom correspondence should be addressed.
†
These authors equally contributed to this work and shared the first position.
Antioxidants 2022, 11(2), 318; https://doi.org/10.3390/antiox11020318
Submission received: 9 January 2022
/
Revised: 27 January 2022
/
Accepted: 2 February 2022
/
Published: 6 February 2022
(This article belongs to the Special Issue Oxidative Stress in Osteoclasts)
Abstract
:Bone tissue engineering is a complex domain that requires further investigation and benefits from data obtained over past decades. The models are increasing in complexity as they reveal new data from co-culturing and microfluidics applications. The in vitro models now focus on the 3D medium co-culturing of osteoblasts, osteoclasts, and osteocytes utilizing collagen for separation; this type of research allows for controlled medium and in-depth data analysis. Oxidative stress takes a toll on the domain, being beneficial as well as destructive. Reactive oxygen species (ROS) are molecules that influence the differentiation of osteoclasts, but over time their increasing presence can affect patients and aid the appearance of diseases such as osteoporosis. Oxidative stress can be limited by using antioxidants such as vitamin K and N-acetyl cysteine (NAC). Scaffolds and biocompatible coatings such as hydroxyapatite and bioactive glass are required to isolate the implant, protect the zone from the metallic, ionic exchange, and enhance the bone regeneration by mimicking the composition and structure of the body, thus enhancing cell proliferation. The materials can be further functionalized with growth factors that create a better response and higher chances of success for clinical use. This review highlights the vast majority of newly obtained information regarding bone tissue engineering, such as new co-culturing models, implant coatings, scaffolds, biomolecules, and the techniques utilized to obtain them.
1. Introduction
There are many circumstances in which bone defects can occur, such as trauma, congenital origins, or disease, impacting millions worldwide [1,2]. The defects present a great challenge for surgeons in cases of sizable osseous defects [3]. The lacunes existing in this area determined the necessity of comprehending the process of bone regeneration in order to generate better solutions [4].
Bone tissue is intensely vascularized, a particularity that influences growth, maturation, shaping, and regeneration [5,6]. Bone tissue can be of two types: cortical bone, which represents the external part of a bone, and cancellous bone, which is located inside the bone. The differences between these two layers are rigidity and porosity. The cortical bone is less porous and presents better mechanical stiffness than cancellous bone, which has approximately ten percent of the cortical bone stiffness [7,8,9]. Osteons inside the cortical bone form units with a Haversian system containing nerves and blood vessels [10]. Cancellous bones do not possess osteons with the Haversian system, but this aspect does not impact the blood vessel stream, which is a consequence of the high bone porosity. In the embryonic stage, bone can form in two distinct methods of ossification: intramembranous and enchondral [11].
Oxidative metabolism produces the reactive oxygen species (ROS) as a byproduct of energy-generating reactions that are largely generated in the mitochondria. A beneficial way in which reduced levels of ROS can operate is as signaling molecules that are vital for balancing cell differentiation, self-renewability, and proliferation. On the other hand, raised levels of ROS are damaging due to the interaction frequency with molecules such as proteins, RNA, and DNA, thus resulting in osteogenic lineage suppression [12]. One crucial factor in bone regeneration is maintaining bone homeostasis. In normal bone homeostasis, osteoblasts differentiation utilizing signal pathways such as fibroblast growth factor (FGF), bone morphogenetic protein, and hedgehog is facilitated; at the same time, the differentiation of osteoclasts is modulated with the aid of macrophage colony-stimulating factor (M-CSF) and the receptor activator of nuclear factor kappa-B ligand (RANKL) [13].
Therefore, this review aims to present the main tools of bone tissue engineering, precisely the scaffolds such as hydroxyapatite and bioactive glass, and the enhancing agents such as growth factors and biomolecules. Furthermore, the models of co-culturing and other studies concerning oxidative stress and reactive species of oxygen have been focused on to observe osteoclasts, osteocytes, and osteoblasts’ influence in the domain, both in single-cell form and together.
2. Bone Regeneration
Several methods address and enhance the regeneration of damaged tissue, which aim to bypass the limitations encountered by already utilized treatments, such as functionalization issues, lack of material compatibility to certain techniques, and translational success limitation [14,15]. Biomaterials and scaffolds can promote healthy tissue formation and are sought in fields such as bone regeneration [16]. Successful approaches orient towards materials that present a good biomimetic property and are bioactive to obtain similar structural features compared to the original extracellular matrix (ECM) [17,18,19].
Hydrogels are a great example when it comes to materials that influence the bone tissue engineering (BTE) field by releasing different types of growth factors (GFs) that aid neovascularization. There are three types of hydrogels that include various options: natural [16], semi-synthetic, and synthetic [6,20,21,22].
A class of materials that are commonly used for bone regeneration applications are bioceramics such as calcium sulfate (CS), hydroxyapatite (HA), and calcium silicate, which are frequently utilized because of their low cytotoxicity and high bioactivity and biocompatibility [23,24]. Bioceramic materials possess a microstructure that promotes ossification and vascularization growth, vital characteristics for osteointegration and osteoinduction [25,26]. This wide class of materials can be utilized as scaffolds which are characterized by properties such as pore shape and size, porosity, crystal distribution, sinter temperature, and phase composition [27,28,29]. Figure 1 presents the advantages and disadvantages of a selected array of vascularization strategies.
3. Hydroxyapatite for BTE
Among the strategies developed in the domain of regenerative medicine that strive to duplicate the tissue in order to obtain an efficient result, we can count materials such as bivalent aptamer-conjugated hydroxyapatite (Apt-HA) [30,31]. Apt-HA’s particularity is the specifically adsorbed vascular endothelial growth factor (VEGF), and it is utilized in synergetic regeneration and osteoconduction [32,33]. The functionalization with growth factors has been discovered recently, and it is realized by immersion; this led to a growth in research for the bioactivity enhancement of bioceramic scaffolds [34,35]. VEGF is one of the first proteins that presented links to osteogenesis and angiogenesis, the inactivation process being an instrument of observation for bone formation and vascularization [36,37]. VEGF and its receptors are expressed by osteoclasts, osteoprogenitors, and osteoblasts, and signal by promoting differentiation, activity, and recruitment [38]. Figure 2 displays the regulation that VEGF realizes to stimulate angiogenesis and osteogenesis.
There are multiple variations in which HA can be obtained depending on the source and quantities of the precursors. A suitable type for BTE can be carbonated hydroxyapatite (CHA), which serves as a coating on metallic substrates, preventing them from releasing metallic ions and protecting them from corrosion [39,40]. The material development is oriented towards mimetism, which guides parameters such as crystallinity, microstructure, and chemical composition. Mineral carbonate (CO3)2− is a component of natural bone that is present in a range of 2%–8% [41]. CHA is obtained by enriching the HA with carbonate minerals such as natural bone [42]. Several sources of bio-waste, such as eggshells, seashells, and animal bones, are rich in calcium, being suitable candidates for medical-grade sources of obtaining HA [43,44,45]. Several technological fluxes are available for obtaining a diversity of CHA with different properties, including co-precipitation [46] sol-gel, nanoemulsion [47], mechanochemical-hydrothermal [48], and mechanical alloying. The most sought method is co-precipitation due to the reduced costs and large-scale production capacity [49].
Another beneficial aspect of HA is the buffering mechanism ensured by PO43− and OH−; the ions of calcium and phosphate aid the remineralization, opposing the effect of calcium carbonate [50,51,52,53]. HA can influence the pH of the cariogenic biofilm, increasing it from 4.3 to 4.8. Apart from their use for mineralization enhancement, HA and CHA can also be used as buffering agents for organic acids [54,55].
Osteoporosis, which can be considered a discrepancy between osteoclasts-related bone resorption and osteoblasts-related bone development, can be caused by an estrogen deficiency [56,57]. The condition results in the mass reduction of cortical and trabecular bone, causing skeletal weakness and fractures. A suitable material for this application is nano HA (nHA), generating signals that stimulate the cell desired behavior and bone biomarker activity [58]. nHA is a great option due to the influence of the particle size on the strength of the implant coating; the scaffold should sustain bone deposition and the forces that are exerted [59].
4. Bioactive Glass
Silicate-based bioactive glass (Si-BaG) has become a very popular material for clinical BTE usage [60,61]. Another type of BaG that gained interest is phosphate BaG (P-BaG), which enables controlled ion release. Aspects such as specific surfaces can be utilized to incorporate or graft an array of biomolecules and curative agents [62,63]. The main advantages of Si-BaG and P-BaG are their modulation of the dissolution rate and incorporation of any desired ions into the composition [64]. The scaffold obtaining technique variations regarding BaG have been explored intensely, except for electrospinning technologies [65,66].
Several studies concluded that BaG accelerates the degradation of polylactides (PLA) as well as poly-L, DL-lactide (PLDLA) [67]. Other papers concluded that composites that included BaG showed enhanced bone generation during longer periods of time and celerity in the molecular weight decrease of PLDLA when utilized in composites such as PLDLA/13-93 BaG [68]. The PLDA/13-93 BaG was also involved in the regulatory process of endothelial markers [69].
Several classifications have been established for BaG in order to see the proper domain of application and the technological flux required to achieve the specifications [70,71]. One option is the sol-gel technique, which involves the addition of a surfactant forming the desired structure of the material. The process involves a calcination step with a temperature of 700 °C, which ensures the removal of organic components obtaining the cavitated material [72].
5. Mesenchymal Stem Cells Influence in BTE
Heterogenous cells do not survive at the center of grafts that exceed thickness due to the restriction of passive transport, which has a limited distance and ensures the flux of metabolites, gases, and nutrients, thus influencing the cell capability [75,76]. Mesenchymal stem cells (MSCs) generate osteoclastogenic cytokines, the receptor activator for nuclear factor kappa B ligand (RANKL), and macrophage colony-stimulating factor (M-CSF) in physiological circumstances. MSCs co-culturing also lowered the tumorigenicity for ovarian cancer cells, but cancer-associated MSCs (CA-MSCs) determine angiogenesis and tumor development when in direct contact with tumor cells or discharging growth factors, cytokines, and exosomes [77]. The discrepancies between MSCs and CA—MSCs provoked interest and proved the CA-MSCs’ distinct properties [78]. The proliferation and diversity of cells are modulated by elements such as pH, dissolved gas, and shear stress [79,80].
Bioreactors can be used to control physicochemical factors such as pH, pressure, humidity, temperature, dissolved oxygen, carbon dioxide, and shear stress [81]. Aspects such as cellular waste removal and the nutrients flux can be controlled by the fabricated medium. Consequently, bioreactors became a desirable option for BTE applications [82,83]. The microenvironment is responsible for the stemness and lineage diversity capacities of the stem cells [84]. Another factor that drastically impacts characteristics, such as morphology and cell viability, is the shear stress-induced alignment [85]. Cell activity can be modulated by the microstructure of the utilized material [86].
Some tested parameters such as runt-related protein 2 (Runx2) and collagen type I (Col1) observed with distinct shear stress indicate that differentiation happens with celerity due to the stress increase. Runx2 represents a vital marker of osteogenesis, and Col1 represents a factor of transcription that underlies the existence of bone cells [87,88]. MSCs are required to be lead to an osteogenic phenotype that can be realized by adding growth factors; another option is their direct cultivation on bone-derived ECM, which presents osteogenic features [89,90].
MSCs therapeutic potential has been observed in several clinical uses between phases I, II, and III, many studies focusing on the engraftment obstacles and diseases regarding hematopoietic stem cells. However, some MSCs such as bone marrow (BM-MSC)-derived ones can express cytokines and thus can be utilized in cancer treatments and therapeutic payloads [77].
There are several types of MSCs sources besides BM: adipose tissue (AT) and umbilical cord (UC) are suited for cell replacement therapy. In comparison to the BM harvested MSCs, MSCs from UC and AT can be collected utilizing less invasive methods. UCs present the advantage of high proliferation and can be cultivated for a long period. AT presents similarities with processed lipoaspirate cells, resulting in a large quantity of cells generated as a by-product of cosmetic liposuction, and can grow in standard culture conditions [77].
6. MicroRNAs in MSCs Differentiation
Since the first isolation of MSCs, their ability of differentiation has been studied and evaluated as being able to act as adipocytes, chondrocytes, and osteocytes, as is displayed in Figure 3 [91,92].
Several studies have been realized on establishing what factors and signaling pathways are involved in the MSC differentiation for the purpose of BTE [93,94]. Cell differentiation has some indispensable regulators such as signals from the ECM, cytokines, and endogenous GFs [95]. Osteogenesis can also be influenced by external aspects such as mechanical forces and electromagnetic fields [96,97]. MSC osteogenic differentiation is supported by the utilized biomaterials [98,99], but the supplementation with ions enhances the osteogenic scaffold potential [100]. Cell differentiation can also be enhanced with the aid of microRNAs (miRNAs) [101], GFs [102], and biophysical stimuli [103]. Some epigenetic factors and processes that modulate the differentiation of MSCs include acetylation and methylation, non-coding RNA (ncRNAs) molecules such as miRNAs [104] and long non-coding RNAs (lncRNA) [105], and DNA methylation [106].
miRNAs are single-stranded ncRNAs that are responsible for regulating 30–60% of protein-coding genes. The mRNA completes the complementary stage and suffers degradation, but in the case of partial mRNA, the protein levels decrease [107,108,109,110,111,112]. One of the main tasks realized by miRNA is to bind to mRNA; apart from this gene, regulation is realized due to molecular mechanisms. The regulation can positively or negatively impact osteogenic differentiation and general transcription factors [113,114]. New approaches find a point of interest in the options that miRNAs offer in BTE [115]. They can be utilized as bioactive factors [116] and scaffolds [117] in order to generate the desired response.
There are several miRNAs involved in the generation of ROS, and thus in the oxidative stress process. Radiosensitivity can be achieved by miR-328-3p when overexpressed due to suppression of H2AX (a subtype of histone) in vitro and in vivo. NAD-dependent deacetylase sirtuin-1 (SIRT1) is capable of cell functional regulation for processes such as oxidative stress, aging, and apoptosis utilizing the deacetylation of a variety of substrates. The upregulation of SIRT1 is realized by miRNA, precisely the miR-199a, which targets the gene [118].
7. Osteoclast Bone Models
Osteoclasts can be defined as large, multinucleated cells which differ from monocytes and macrophages by presenting M-CSF and RANKL [119]. The co-culture techniques became popular tests for investigating bone cells such as osteoclasts, osteocytes, and osteoblasts for purposes such as bone metabolism and disease behavior [120,121,122]. Many articles have approached the co-culturing systems with osteoclasts and osteoblasts in vitro [123,124]. The main reason for the co-culture tests is to establish the interactions between cells and test different approaches in a controlled environment to generate information that can further be applied in the BTE domain [125]. The predominance of osteoblast/osteoclast co-culture studies frequency is due to the restricted availability of osteocytes. The lack of osteocytes availability resides in the difficult process of isolation for these post-mitotic cells [126,127]. There is a lack of primary osteocyte co-cultured studies with osteoclasts, with only two models reported at present, but some studies have treated the subject using murine sources for osteocytes [128]. In most cases, there are only two bone-related cell species in co-culturing techniques. The main reason is the difficulties in terms of requirements needed because of the different medium condition preparation, this being a very important step for a successful study [129]. Some results show the presence of osteoclasts, osteocytes, and osteoblasts after 21 days, even if at the beginning of the research the precursors utilized were for primary human osteoclasts and osteoblasts [125]. The setback of this study was the inability to generate data for single-cell species to investigate the matter further. Because vascularization is necessary for BTE, there has been co-culturing research involving the use of osteoclasts, osteoblasts, and endothelial cells. It has been stated in several studies that cell cultures behave in different ways in comparison with similar cells but are co-cultured, and the co-culturing technique became accepted as a common practice in biomaterial research [130]. There have been new models established that operate with triple cultures in order to obtain data on bone cellular infrastructures [131].
In the established triple cultures, all cell species showed their typical morphology and there were no obvious morphological differences between single and triple cultures. A good balance between the three cell species is a prerequisite to use those triple cultures in the future to investigate the influence of bioactive molecules, drugs, and biomaterial extracts. As expected, due to the signaling between the cells, there were detected differences between single and triple cultures on the mRNA level. Table 1 presents details regarding the techniques that are frequently used [123,128,132].
The latest model was based on generating a three-dimensional environment permitting cell–cell interactions of osteoclasts, osteoblasts, and osteocytes where the analysis can occur for all cell types as well as for single-cell species. This kind of model aims to generate a perspective on complex systems that include all the major species of bone cells. Collagen can be utilized in order to separate osteocytes from the other cells, allowing a gradually obtained result [125].
Bone cells are responsible for the development and metabolism of the bone, but there is a whole system influencing the result, which involves cytokines, GFs, receptors, transcription factors, ligands, and cell-specific enzymes [133]. The in vitro studies have a controlled medium and various options available, which also imply many variable factors that need to be addressed. Thus, the studies have many challenges to overcome. Existent models have studied vasculogenesis, bone marrow, and tumor models [134]. The models also went in a direction similar to microfluidics; approaches such as bone-on-a-chip and organ-on-a-chip lead towards a system that facilitates the investigation of osteoclasts or osteoblasts responding to osteocyte-prepared medium [135,136]. However, since microfluidics work with a limited number of samples, the popular choice remains a simpler co-culturing model strongly impacted by the collagen gel, which allows obtaining a 3D environment [137,138].
Materials such as copper are used due to the great properties they possess, such as angiogenesis and osteogenesis enhancement, as well as antimicrobial effects [139,140]. This material is used to generate enhanced BaG, titanium implants, cement, and ceramics [141,142]. The osteoclasts are influenced by the material due to the potential oxidative stress exerted at different concentrations [138].
There are four main categories of 3D co-culturing models: hydrogels, cell aggregation, scaffolds, and dynamic models. Some of them can be divided once more into subcategories, thus ensuring diversity and options for several new applications. For hydrogels we have the source, which can be natural or synthetic, the first being a network composed out of naturally originated monomers and the latter being synthetic. Both present different advantages and disadvantages according to the source and profile, even though diffusion plays a large role in the success of both. The cell aggregation can be classified with the technological flux, one being obtained with hanging drop plates and the other being generated with 3D Petri dishes. The first one generates 3D spheroid aggregates and the second technique molds agarose in order to obtain defined shapes by cell agglomeration. The scaffolds can also be differentiated with the source of the material dividing it into synthetic and natural 3D structures [123].
There are several new theories that will increase the complexity of the existing 3D models. Three-dimensional models may replicate the in vivo conditions while using human cells in order to obtain valid information regarding the hypothesis obtained with the animal trials which require further analysis. Furthermore, techniques such as perfusion and biomechanical mimetism have created room for studies that generate similar conditions to the in vivo environment, thus becoming a preferred model for researchers. Also, they ensure a high compatibility rate and diversity, alongside good chemical properties [123].
For instance, Zhang and colleagues [143] proposed a complex 3D structure-based co-culture platform that mimics the Haversian bone, with osteogenic cells angiogenic/neurogenic cells distributed at a specific location for active bone tissue engineering. The authors have used digital laser processing-based 3D printing technology to ensure the precision of their scaffolds, allowing the creation of custom-designed structures. The as-described system displayed significantly improved osteogenic and angiogenic effects as compared with the unicellular delivery system both in vitro and in vivo, holding great promise for tissue regeneration.
Moreover, recent progress has been reported in moving from preclinical 3D models to clinical models [144]. For example, Pauli et al. [145] have described the development of a precision cancer care platform integrating whole-exosome sequencing with a living biobank that enables high-throughput drug screens on patient-derived tumor organoids. The scientists investigated 56 tumor-derived organoid cultures and 19 patient-derived xenograft models, including bone models, from 769 patients enrolled in an Institutional Review Board-approved clinical trial. Their tremendous effort may serve as a basis for discovering novel personalized therapeutic options, especially for patients where standard clinical options have been exhausted.
Several other perspectives in the field of 3D bone culturing models include automating cell seeding procedures towards attaining a safe and standardized production of engineered tissue constructs, using nondestructive live-monitoring techniques to obtain unique insights into cellular interactions, and moving from single culture to multiorgan models in order to understand the molecular communications between the bone tissue model and other tissues/organs [146].
8. Oxidative Stress Influence in BTE
Osteoblasts and osteoclasts are vital in bone remodeling; thus, they have been investigated to better comprehend the bone regeneration mechanism [147,148]. After the discovery of osteoprotegerin (OPG), RANKL, and RANK, a pathway was formed which supplies data for the osteoblast regulation of osteoclasts utilizing bone matrix interaction, paracrine factors, and cell–cell contact [123,149]. Osteoclasts differentiation (Figure 4) starts with the attachment of RANKL to the RANK receptor with the aid of a receptor-associated factor (TRAF6) which activates a wide range of mitogen-activated protein kinases (MAPKs) that trigger the nuclear factor of activated T cells 1 (NFATc1), leading to osteoclasts differentiation [148].
Reactive oxygen species (ROS) are molecules and free radicals (e.g., superoxide anion, hydrogen peroxide, hydroxyl radical) mainly resulting as byproducts of leaked electrons from the mitochondrial electron transport chain during aerobic respiration [118]. ROS are necessary for regulating cellular processes, including proliferation, survival, metabolism, apoptosis, and differentiation. ROS are moieties of interest in bone regeneration as they can be used for dual purposes. Their beneficial aspect resides in the ability to act as an intracellular marking agent, being essential for the transmission of cell signals [148].
However, ROS can produce cellular imbalance in reduction–oxidation reactions when their level increases due to age or inflammatory states, leading to oxidative stress. Oxidative stress modulates fundamental cellular physiological responses via signal transduction, transcription factors, and ncRNAs, promoting nuclear and mitochondrial DNA damage and initiating DNA repair pathways [118,148]. Continued oxidative stress was demonstrated to generate diseases, leading to bone destruction and cellular death [149].
ROS are also vital components in osteoclasts’ regulation of differentiation [150]. It had been assumed that the extreme production of osteoclasts induced by local inflammation could be prevented by limiting the excessive production of intracellular ROS. The impact of ROS in the relationship of osteoclasts and osteoblasts was studied with the help of co-cultured models [151]. In particular, maintaining bone homeostasis is critical in preserving an optimum balance between formation and resorption, influencing bone mass and strength. Specifically, these properties begin to reduce with aging, with an enhanced osteoclast activity and a decreased osteoblast activity [148].
Cellular senescence results in a response to persistent stress; it is characterized by a stable cell-cycle arrest, the expression of senescence-associated β-galactosidase (SA-β-gal), the increased expression of the cell cycle inhibitor p16Ink4a, the overexpression of the cyclin-dependent kinase (CDK) inhibitor, senescence-associated secretory phenotype (SASP), telomere shortening, and a persistent DNA damage response (DDR) [118,149]. This is relevant in the context of oxidative stress as ROS provoke cell senescence by stimulating the DDR pathway to stabilize p53 and promote CDK inhibitor gene expression. Particularly, p53 represents a major regulator of cellular response to oxidative stress. On the one hand, it can decrease ROS levels and repair DNA damage in cells; on the other hand, p53 can increase ROS production and promote the apoptosis or senescence of the cells [118].
Moreover, several conditions can occur due to the patient’s age, such as postmenopausal osteoporosis, diabetes, cirrhosis, cancer, and neurogenerative disease, which can be delayed with antioxidants related to chemo-preventive and curative therapies such as glutathione [152,153,154,155]. The redox indicators that are usually analyzed are reduced glutathione (GSH) and oxidized glutathione (GSSG) and their balance. In addition, some studies noted the increase of osteoclasts differentiation in the presence of GSH [156].
One of the popular antioxidants used for clinical studies and cell cultures is N-acetyl cysteine (NAC) [157,158]. Studies that involve NAC treatments concluded a reduction in cellular processes, and ROS have some involvement in the matter. NAC and ascorbate have beneficial effects in reducing stimulus for the loss of bone mass, osteoblast apoptosis, oxidative stress, and osteoclastogenesis after gonadectomy [159,160]. NAC has also been utilized in the analysis of mitochondrial ROS and physiological involvement [161]. Important results have been also obtained by the oral administration of Ewha-18278 (a pyrazole derivative). It was reported that Ewha-18278 protected ovariectomy-induced osteoporosis in mice by NADPH oxidase (NOX) inhibition and ROS suppression. This anti-osteoporotic agent aided in the recovery of bone parameters, leading to improved bone strength and an increased number of osteoblasts [148]. Another beneficial effect was registered from vitamin K managing to reduce oxidative stress and ROS production. Specifically, vitamin K was observed to protect cells from H2O2-induced changes in protein expression, being able to support bone tissue formation, remodeling, and mineralization [162,163].
Phosphoinositide 3-kinase (PI3K)/protein kinase B (AKT) represents a signaling pathway that regulates cells’ proliferation, survival, and death and the osteoblastic and osteoclastic functions altering the formation, differentiation, and apoptosis. The deficiency of AKT2 translates into a decrease in RUNX2 expression. Thus, the AKT pathway promotes RUNX2 gene expression [164]. Osteogenic differentiation is promoted by the regulation of PI3K/AKT and RUNX2, modulating the RUNX2 activity [165].
Table 2 comprises several clinical studies that are being realized, researched, or revisited. The trials reflect the focus growth that the domain is gaining due to the continuous discoveries in the field. In time, the data and the enhancement of cell isolation will allow more clinical trials to take place. However, the stages in which some studies reside are incipient and still focus on side effects and dosage for emerging solutions.
9. Discussion
Bone regeneration requires the top performance of several components such as efficient analysis, materials that possess special properties, suitable GFs, and molecules to ensure a successful overall process.
Oxidative stress plays a vital part in bone regeneration as it is known to alter the process of bone remodeling. Through the unbalance it creates between osteoblasts and osteoclasts activity, persistent exposure to high levels of oxidative stress may further result in cellular senescence, bone diseases, and skeletal system disorders. Thus, special consideration must be given to maintaining oxidative stress at optimum levels. Particularly, the destructive potential of ROS can be reduced by the administration of antioxidant agents that can restore bone parameters, supporting bone tissue formation, remodeling, and mineralization.
Nonetheless, ROS production can also be considered relevant as a therapeutic target under certain conditions, thus being a beneficial aspect if tackled properly. More exactly, ROS can act as an intracellular marking agent, being involved in the transmission of cell signals. Hence, its detection and targeting can contribute to a better understanding of cellular interactions and approaching pathophysiological conditions in a more specific manner. Therefore, oxidative stress needs to be further regulated and researched in order to benefit from good results in the field.
Bone regeneration can be enhanced by use of specifically engineered scaffolds. Materials such as BaG and hydroxyapatite remain good performers among others which possess advantageous properties, such as a low cytotoxicity and good biocompatibility. This provides a strong basis for the functionalization and regeneration of the tissue.
Culturing systems are also important tools in analyzing the mechanisms of bone regeneration, bone homeostasis, and ROS generation and effectively evaluating the various cells’ response to emerging BTE strategies. In particular, the 3D techniques generate new data and legitimization to animal studies that were never performed on humans before. Also, the mimetism of the in vivo domain makes it a good perspective to take into consideration when it is time to start clinical studies.
Growth factors such as VEGF modulate the process of angiogenesis and osteogenesis. Hence, they represent valuable assets to any strategy when used properly. Figure 5 displays the main fields that take part in BTE.
To summarize the discussion on bone regeneration and oxidative stress, Table 3 synthesizes the roles and effects of the various described cells, growth factors, biomolecules, signal pathways, and bone engineering scaffolds.
10. Conclusions and Future Perspectives
Bone regeneration depends on several other domains to generate the best results, with many discoveries and breakthroughs owed to interdisciplinary research studies. Bioactive materials have been researched independently and used under the form of composites to obtain synergistic results monitored through in vitro and in vivo tests. The materials response leads to the discussion of functionalization with biomolecules and growth factors to accelerate the healing process with the full potential of cell growth. Aspects and particularities such as tensile strength, pore dimension, and composition are frequently modulated to obtain a data flux that can help decide the desired path to be followed.
As bone cells are scarce, not much research is available in clinical settings. Thus, studies mostly concern the employment of culture models for investigating bone cell interactions, bone metabolism, and disease behavior. In this respect, particular attention has been drawn to coculturing osteoclasts, osteocytes, and osteoblasts into biomimetic platforms. The co-cultured models have also sparked the idea of microfluidic attempts to generate bone-on-a-chip and ensure reliable data, but lack the ability to obtain a wide array of samples to date. Interesting results are also expected from the future development of multiorgan models that would help comprehend the importance of bone homeostasis in relation to other tissues and organs.
Considering every aspect that the domain provides, the existing solutions are continuously enhanced with the availability of new data streams. Oxidative stress and its associated diseases are being analyzed, and new antioxidants emerge as promising solutions. The beneficial aspects of ROS and the damaging concentrations of the molecules are being tested to have the greatest benefits from the data obtained.
To conclude, multiple bone scaffolding materials and functionalization molecules are emerging from recent studies and might soon start being utilized for obtaining a better clinical experience. Thus, we can presume that developing interdisciplinary studies lead to the emergence of new technologies for tissue engineering, oxidative stress control, osteogenesis promotion, and bone disease prevention.
Author Contributions
A.E.B., D.M.R., A.I., A.-G.N., A.M.G. and A.R.R. participated in the review, writing, and revision. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chen, C.-Y.; Chen, C.-C.; Wang, C.-Y.; Lee, A.K.; Yeh, C.-L.; Lin, C.-P. Assessment of the Release of Vascular Endothelial Growth Factor from 3D-Printed Poly-ε-Caprolactone/Hydroxyapatite/Calcium Sulfate Scaffold with Enhanced Osteogenic Capacity. Polymers 2020, 12, 1455. [Google Scholar] [CrossRef]
- Zyuz’kov, G.N. Targeted Regulation of Intracellular Signal Transduction in Regeneration-Competent Cells: A new Direction for Therapy in Regenerative Medicine. Biointerface Res. Appl. Chem. 2021, 11, 12238–12251. [Google Scholar] [CrossRef]
- He, J.; Chen, G.; Liu, M.; Xu, Z.; Chen, H.; Yang, L.; Lv, Y. Scaffold strategies for modulating immune microenvironment during bone regeneration. Mater. Sci. Eng. C Mater. Biol. Appl. 2020, 108, 110411. [Google Scholar] [CrossRef]
- Wang, W.; Nune, K.C.; Tan, L.; Zhang, N.; Dong, J.; Yan, J.; Misra, R.D.K.; Yang, K. Bone regeneration of hollow tubular magnesium-strontium scaffolds in critical-size segmental defects: Effect of surface coatings. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 100, 297–307. [Google Scholar] [CrossRef]
- Sharma, A.; Sharma, P.K.; Malviya, R. Stimuli-responsive supramolecules for bone tissue engineering. Biointerface Res. Appl. Chem. 2020, 10, 5122–5127. [Google Scholar] [CrossRef]
- Divband, B.; Samiei, M.; Davaran, S.; Roshangar, L.; Shahi, S.; Aghazadeh, M. Synthesis and in vitro Evaluation of Thermosensitive PLA-g-P(HEM-co-NIPAAM) Hydrogel Used for Delivery of VEGF. Biointerface Res. Appl. Chem. 2021, 11, 8043–8051. [Google Scholar] [CrossRef]
- Simunovic, F.; Finkenzeller, G. Vascularization Strategies in Bone Tissue Engineering. Cells 2021, 10, 1749. [Google Scholar] [CrossRef]
- Morgan, E.F.; Unnikrisnan, G.U.; Hussein, A.I. Bone Mechanical Properties in Healthy and Diseased States. Annu. Rev. Biomed. Eng. 2018, 20, 119–143. [Google Scholar] [CrossRef]
- Shadman, B.; Asadi, A.; Zahri, S.; Barzegar, A.; Marzban, A. Design of PLGA-based Scaffolds for Developing and Differentiating Mesenchymal Stem Cells (MSCs). Biointerface Res. Appl. Chem. 2021, 11, 12732–12742. [Google Scholar] [CrossRef]
- Chang, B.; Liu, X. Osteon: Structure, Turnover, and Regeneration. Tissue Eng. Part B Rev. 2021. [Google Scholar] [CrossRef]
- Salhotra, A.; Shah, H.N.; Levi, B.; Longaker, M.T. Mechanisms of bone development and repair. Nat. Rev. Mol. Cell Biol. 2020, 21, 696–711. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Yan, Y.; Zhao, C.; Xu, Y.; Wang, Q.; Xu, N. Resveratrol improves osteogenic differentiation of senescent bone mesenchymal stem cells through inhibiting endogenous reactive oxygen species production via AMPK activation. Redox Rep. Commun. Free Radic. Res. 2019, 24, 62–69. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, K.; Kho, D.H.; Yanagawa, T.; Zimel, M.; Heath, E.; Hogan, V.; Raz, A. Galectin-3 in bone tumor microenvironment: A beacon for individual skeletal metastasis management. Cancer Metastasis Rev. 2016, 35, 333–346. [Google Scholar] [CrossRef] [PubMed]
- Montalbano, G.; Tomasina, C.; Fiorilli, S.; Camarero-Espinosa, S.; Vitale-Brovarone, C.; Moroni, L. Biomimetic Scaffolds Obtained by Electrospinning of Collagen-Based Materials: Strategies to Hinder the Protein Denaturation. Materials 2021, 14, 4360. [Google Scholar] [CrossRef]
- Johnson, C.D.L.; Ganguly, D.; Zuidema, J.M.; Cardinal, T.J.; Ziemba, A.M.; Kearns, K.R.; McCarthy, S.M.; Thompson, D.M.; Ramanath, G.; Borca-Tasciuc, D.A.; et al. Injectable, Magnetically Orienting Electrospun Fiber Conduits for Neuron Guidance. ACS Appl. Mater. Interfaces 2019, 11, 356–372. [Google Scholar] [CrossRef] [PubMed]
- Ghassemi, T.; Shahroodi, A.; Ebrahimzadeh, M.H.; Mousavian, A.; Movaffagh, J.; Moradi, A. Current Concepts in Scaffolding for Bone Tissue Engineering. Arch. Bone Jt. Surg. 2018, 6, 90–99. [Google Scholar]
- Zhang, D.; Wu, X.; Chen, J.; Lin, K. The development of collagen based composite scaffolds for bone regeneration. Bioact. Mater. 2018, 3, 129–138. [Google Scholar] [CrossRef]
- Piard, C.; Luthcke, R.; Kamalitdinov, T.; Fisher, J. Sustained delivery of vascular endothelial growth factor from mesoporous calcium-deficient hydroxyapatite microparticles promotes in vitro angiogenesis and osteogenesis. J. Biomed. Mater. Res. Part A 2021, 109, 1080–1087. [Google Scholar] [CrossRef]
- Leonardo, C.; Giorgio, L.I.; Sara, R.; Sabrina, G.; Andrea, V.; Annapaola, P.; Alessandro, S.; Barbara, D.; Paolo, M.; Adriano, P.; et al. Nanostructured surface bioactive composite scaffold for filling of bone defects. Biointerface Res. Appl. Chem. 2020, 10, 5038–5047. [Google Scholar] [CrossRef]
- Adibfar, A.; Amoabediny, G.; Baghaban Eslaminejad, M.; Mohamadi, J.; Bagheri, F.; Zandieh Doulabi, B. VEGF delivery by smart polymeric PNIPAM nanoparticles affects both osteogenic and angiogenic capacities of human bone marrow stem cells. Mater. Sci. Eng. C 2018, 93, 790–799. [Google Scholar] [CrossRef]
- Bakshi, R.; Hokugo, A.; Khalil, D.; Wang, L.; Shibuya, Y.; Zhou, S.; Zhang, Z.; Rezzadeh, K.; McClendon, M.; Stupp, S.I.; et al. A Chemotactic Functional Scaffold with VEGF-Releasing Peptide Amphiphiles Facilitates Bone Regeneration by BMP-2 in a Large-Scale Rodent Cranial Defect Model. Plast. Reconstr. Surg. 2021, 147, 386–397. [Google Scholar] [CrossRef] [PubMed]
- Lü, L.; Deegan, A.; Musa, F.; Xu, T.; Yang, Y. The effects of biomimetically conjugated VEGF on osteogenesis and angiogenesis of MSCs (human and rat) and HUVECs co-culture models. Colloids Surf. B Biointerfaces 2018, 167, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Jha, P.; Danewalia, S.S.; Sharma, G.; Singh, K. Antimicrobial and bioactive phosphate-free glass-ceramics for bone tissue engineering applications. Mater. Sci. Eng. C Mater. Biol. Appl. 2018, 86, 9–17. [Google Scholar] [CrossRef]
- Massit, A.; El Yacoubi, A.; Rezzouk, A.; El Idrissi, B.C. Thermal Behavior of Mg-Doped Calcium-Deficient Apatite and Stabilization of beta Tricalcium Phosphate. Biointerface Res. Appl. Chem. 2020, 10, 6837–6845. [Google Scholar] [CrossRef]
- Hong, S.; Kang, E.Y.; Byeon, J.; Jung, S.-H.; Hwang, C. Embossed Membranes with Vascular Patterns Guide Vascularization in a 3D Tissue Model. Polymers 2019, 11, 792. [Google Scholar] [CrossRef] [Green Version]
- Francisco, E.M.; Zoccolotti, J.D.; Tiomnova, O.T.; Tolaba, A.G.; Chanfrau, J.E.R.; Jorge, J.H.; Basmaji, P.; Guastaldi, A.C. Sterilization of Scaffolds of Calcium Phosphates and Bacterial Cellulose for their Use in Tissue Regeneration. Biointerface Res. Appl. Chem. 2021, 11, 10089–10098. [Google Scholar] [CrossRef]
- Shie, M.-Y.; Shen, Y.-F.; Astuti, S.D.; Lee, A.K.; Lin, S.-H.; Dwijaksara, N.L.; Chen, Y.-W. Review of Polymeric Materials in 4D Printing Biomedical Applications. Polymers 2019, 11, 1864. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.H.; Chuang, T.Y.; Chiang, W.H.; Chen, I.P.; Wang, K.; Shie, M.Y.; Chen, Y.W. The synergistic effects of graphene-contained 3D-printed calcium silicate/poly-ε-caprolactone scaffolds promote FGFR-induced osteogenic/angiogenic differentiation of mesenchymal stem cells. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 104, 109887. [Google Scholar] [CrossRef]
- Tsai, C.H.; Hung, C.H.; Kuo, C.N.; Chen, C.Y.; Peng, Y.N.; Shie, M.Y. Improved Bioactivity of 3D Printed Porous Titanium Alloy Scaffold with Chitosan/Magnesium-Calcium Silicate Composite for Orthopaedic Applications. Materials 2019, 12, 203. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Lee, S.; Choi, Y.; Choi, J.; Kang, B.J. Sustained Release of Bone Morphogenetic Protein-2 through Alginate Microbeads Enhances Bone Regeneration in Rabbit Tibial Metaphyseal Defect Model. Materials 2021, 14, 2600. [Google Scholar] [CrossRef]
- Dang, M.; Saunders, L.; Niu, X.; Fan, Y.; Ma, P.X. Biomimetic delivery of signals for bone tissue engineering. Bone Res. 2018, 6, 25. [Google Scholar] [CrossRef]
- Son, J.; Kim, J.; Lee, K.; Hwang, J.; Choi, Y.; Seo, Y.; Jeon, H.; Kang, H.C.; Woo, H.M.; Kang, B.J.; et al. DNA aptamer immobilized hydroxyapatite for enhancing angiogenesis and bone regeneration. Acta Biomater. 2019, 99, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Dreyer, C.H.; Kjaergaard, K.; Ding, M.; Qin, L. Vascular endothelial growth factor for in vivo bone formation: A systematic review. J. Orthop. Translat. 2020, 24, 46–57. [Google Scholar] [CrossRef]
- Chen, Y.-W.; Shen, Y.-F.; Ho, C.-C.; Yu, J.; Wu, Y.-H.A.; Wang, K.; Shih, C.-T.; Shie, M.-Y. Osteogenic and angiogenic potentials of the cell-laden hydrogel/mussel-inspired calcium silicate complex hierarchical porous scaffold fabricated by 3D bioprinting. Mater. Sci. Eng. C 2018, 91, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Hao, F.; Qin, L.; Liu, J.; Chang, J.; Huan, Z.; Wu, L. Assessment of calcium sulfate hemihydrate-Tricalcium silicate composite for bone healing in a rabbit femoral condyle model. Mater. Sci. Eng. C Mater. Biol. Appl. 2018, 88, 53–60. [Google Scholar] [CrossRef]
- Diomede, F.; Marconi, G.D.; Fonticoli, L.; Pizzicanella, J.; Merciaro, I.; Bramanti, P.; Mazzon, E.; Trubiani, O. Functional Relationship between Osteogenesis and Angiogenesis in Tissue Regeneration. Int. J. Mol. Sci. 2020, 21, 3242. [Google Scholar] [CrossRef]
- Rumney, R.M.H.; Lanham, S.A.; Kanczler, J.M.; Kao, A.P.; Thiagarajan, L.; Dixon, J.E.; Tozzi, G.; Oreffo, R.O.C. In vivo delivery of VEGF RNA and protein to increase osteogenesis and intraosseous angiogenesis. Sci. Rep. 2019, 9, 17745. [Google Scholar] [CrossRef] [Green Version]
- Diomede, F.; Marconi, G.D.; Cavalcanti, M.; Pizzicannella, J.; Pierdomenico, S.D.; Fonticoli, L.; Piattelli, A.; Trubiani, O. VEGF/VEGF-R/RUNX2 Upregulation in Human Periodontal Ligament Stem Cells Seeded on Dual Acid Etched Titanium Disk. Materials 2020, 13, 706. [Google Scholar] [CrossRef] [Green Version]
- Sari, M.; Kristianto, N.A.; Chotimah; Ana, I.D.; Yusuf, Y. Carbonated Hydroxyapatite-Based Honeycomb Scaffold Coatings on a Titanium Alloy for Bone Implant Application—Physicochemical and Mechanical Properties Analysis. Coatings 2021, 11, 941. [Google Scholar] [CrossRef]
- Singh, A.; Singh, G.; Chawla, V. Characterization and mechanical behaviour of reinforced hydroxyapatite coatings deposited by vacuum plasma spray on SS-316L alloy. J. Mech. Behav. Biomed. Mater. 2018, 79, 273–282. [Google Scholar] [CrossRef]
- Permatasari, H.A.; Supii, A.I.; Suparta, G.B.; Yusuf, Y. Characteristics of Abalone Mussel Shells (Halioitis asinina) with Calcination Temperature Variations as a Basic Material for Synthesis of Carbonated Hydroxyapatite. Key Eng. Mater. 2019, 818, 31–36. [Google Scholar] [CrossRef]
- Safarzadeh, M.; Ramesh, S.; Tan, C.Y.; Chandran, H.; Noor, A.F.M.; Krishnasamy, S.; Alengaram, U.J.; Ramesh, S. Effect of multi-ions doping on the properties of carbonated hydroxyapatite bioceramic. Ceram. Int. 2019, 45, 3473–3477. [Google Scholar] [CrossRef]
- Dhanaraj, K.; Suresh, G. Conversion of waste sea shell (Anadara granosa) into valuable nanohydroxyapatite (nHAp) for biomedical applications. Vacuum 2018, 152, 222–230. [Google Scholar] [CrossRef]
- Núñez, D.; Elgueta, E.; Varaprasad, K.; Oyarzún, P. Hydroxyapatite nanocrystals synthesized from calcium rich bio-wastes. Mater. Lett. 2018, 230, 64–68. [Google Scholar] [CrossRef]
- Ramesh, S.; Loo, Z.Z.; Tan, C.Y.; Chew, W.J.K.; Ching, Y.C.; Tarlochan, F.; Chandran, H.; Krishnasamy, S.; Bang, L.T.; Sarhan, A.A.D. Characterization of biogenic hydroxyapatite derived from animal bones for biomedical applications. Ceram. Int. 2018, 44, 10525–10530. [Google Scholar] [CrossRef]
- Anggraini, R.M.; Supii, A.I.; Suparta, G.B.; Yusuf, Y. The Effect of pH on the Characteristics of Carbonate Hydroxyapatite Based on Pearl Shell (Pinctada maxima). Key Eng. Mater. 2019, 818, 44–49. [Google Scholar] [CrossRef]
- Ezekiel, I.; Kasim, S.R.; Ismail, Y.M.B.; Noor, A.-F.M. Nanoemulsion synthesis of carbonated hydroxyapatite nanopowders: Effect of variant CO32−/PO43− molar ratios on phase, morphology, and bioactivity. Ceram. Int. 2018, 44, 13082–13089. [Google Scholar] [CrossRef]
- Kong, W.; Zhao, K.; Gao, C.; Zhu, P. Synthesis and characterization of carbonated hydroxyapatite with layered structure. Mater. Lett. 2019, 255, 126552. [Google Scholar] [CrossRef]
- Sari, M.; Hening, P.; Chotimah, A.I.D.; Yusuf, Y. Bioceramic hydroxyapatite-based scaffold with a porous structure using honeycomb as a natural polymeric Porogen for bone tissue engineering. Biomater. Res. 2021, 25, 2. [Google Scholar] [CrossRef] [PubMed]
- Meyer, F.; Enax, J.; Epple, M.; Amaechi, B.T.; Simader, B. Cariogenic Biofilms: Development, Properties, and Biomimetic Preventive Agents. Dent. J. 2021, 9, 88. [Google Scholar] [CrossRef]
- Cieplik, F.; Rupp, C.M.; Hirsch, S.; Muehler, D.; Enax, J.; Meyer, F.; Hiller, K.-A.; Buchalla, W. Ca2+ release and buffering effects of synthetic hydroxyapatite following bacterial acid challenge. BMC Oral Health 2020, 20, 85. [Google Scholar] [CrossRef] [Green Version]
- Enax, J.; Fabritius, H.-O.; Fabritius-Vilpoux, K.; Amaechi, B.T.; Meyer, F. Modes of action and clinical efficacy of particulate hydroxyapatite in preventive oral health care—State of the art. Open Dent. J. 2019, 13, 274–287. [Google Scholar] [CrossRef]
- Enax, J.; Fabritius, H.-O.; Amaechi, B.T.; Meyer, F. Hydroxylapatit als biomimetischer Wirkstoff für die Remineralisation von Zahnschmelz und Dentin. ZWR-Das Dtsch. Zahnärzteblatt 2020, 129, 277–283. [Google Scholar] [CrossRef]
- Amaechi, B.T.; AbdulAzees, P.A.; Okoye, L.O.; Meyer, F.; Enax, J. Comparison of hydroxyapatite and fluoride oral care gels for remineralization of initial caries: A pH-cycling study. BDJ Open 2020, 6, 9. [Google Scholar] [CrossRef] [PubMed]
- Hejazi, F.; Ebrahimi, V.; Asgary, M.; Piryaei, A.; Fridoni, M.J.; Kermani, A.A.; Zare, F.; Abdollahifar, M.A. Improved healing of critical-size femoral defect in osteoporosis rat models using 3D elastin/polycaprolactone/nHA scaffold in combination with mesenchymal stem cells. J. Mater. Sci. Mater. Med. 2021, 32, 27. [Google Scholar] [CrossRef] [PubMed]
- Scribante, A.; Dermenaki Farahani, M.R.; Marino, G.; Matera, C.; Rodriguez y Baena, R.; Lanteri, V.; Butera, A. Biomimetic Effect of Nano-Hydroxyapatite in Demineralized Enamel before Orthodontic Bonding of Brackets and Attachments: Visual, Adhesion Strength, and Hardness in In Vitro Tests. BioMed Res. Int. 2020, 2020, 6747498. [Google Scholar] [CrossRef] [Green Version]
- Scalize, P.H.; de Sousa, L.G.; Gonçalves, L.M.N.; Pitol, D.L.; Palinkas, M.; Coppi, A.A.; Righeti, M.A.; Ricardo, V.; Bombonato-Prado, K.F.; Regalo, S.C.H.; et al. Low-level laser therapy enhances the number of osteocytes in calvaria bone defects of ovariectomized rats. Animal Model. Exp. Med. 2019, 2, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Lowe, B.; Hardy, J.G.; Walsh, L.J. Optimizing Nanohydroxyapatite Nanocomposites for Bone Tissue Engineering. ACS Omega 2020, 5, 1–9. [Google Scholar] [CrossRef]
- Du, M.; Chen, J.; Liu, K.; Xing, H.; Song, C. Recent advances in biomedical engineering of nano-hydroxyapatite including dentistry, cancer treatment and bone repair. Compos. Part B Eng. 2021, 215, 108790. [Google Scholar] [CrossRef]
- Lyyra, I.; Leino, K.; Hukka, T.; Hannula, M.; Kellomäki, M.; Massera, J. Impact of Glass Composition on Hydrolytic Degradation of Polylactide/Bioactive Glass Composites. Materials 2021, 14, 667. [Google Scholar] [CrossRef]
- Fernandes, H.R.; Gaddam, A.; Rebelo, A.; Brazete, D.; Stan, G.E.; Ferreira, J.M.F. Bioactive Glasses and Glass-Ceramics for Healthcare Applications in Bone Regeneration and Tissue Engineering. Materials 2018, 11, 2530. [Google Scholar] [CrossRef] [Green Version]
- Pontremoli, C.; Boffito, M.; Fiorilli, S.; Laurano, R.; Torchio, A.; Bari, A.; Tonda-Turo, C.; Ciardelli, G.; Vitale-Brovarone, C. Hybrid injectable platforms for the in situ delivery of therapeutic ions from mesoporous glasses. Chem. Eng. J. 2018, 340, 103–113. [Google Scholar] [CrossRef]
- Fiorilli, S.; Molino, G.; Pontremoli, C.; Iviglia, G.; Torre, E.; Cassinelli, C.; Morra, M.; Vitale-Brovarone, C. The Incorporation of Strontium to Improve Bone-Regeneration Ability of Mesoporous Bioactive Glasses. Materials 2018, 11, 678. [Google Scholar] [CrossRef] [Green Version]
- Lapa, A.; Cresswell, M.; Jackson, P.; Boccaccini, A.R. Phosphate glass fibres with therapeutic ions release capability—A review. Adv. Appl. Ceram. 2020, 119, 1–14. [Google Scholar] [CrossRef]
- Montalbano, G.; Fiorilli, S.; Caneschi, A.; Vitale-Brovarone, C. Type I Collagen and Strontium-Containing Mesoporous Glass Particles as Hybrid Material for 3D Printing of Bone-Like Materials. Materials 2018, 11, 700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montalbano, G.; Borciani, G.; Cerqueni, G.; Licini, C.; Banche-Niclot, F.; Janner, D.; Sola, S.; Fiorilli, S.; Mattioli-Belmonte, M.; Ciapetti, G.; et al. Collagen Hybrid Formulations for the 3D Printing of Nanostructured Bone Scaffolds: An Optimized Genipin-Crosslinking Strategy. Nanomaterials 2020, 10, 1681. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, P.; Sun, D.; Luo, Y.; Chen, C.; Tang, Z.; Liao, Y.; Cao, X.; Cheng, C.; Liu, W.; et al. Mechanical and degradative properties of PLDLA biodegradable pins with bioactive glass fibers in a beagle model. Biomed. Mater. 2020, 15, 035010. [Google Scholar] [CrossRef] [PubMed]
- Houaoui, A.; Lyyra, I.; Agniel, R.; Pauthe, E.; Massera, J.; Boissière, M. Dissolution, bioactivity and osteogenic properties of composites based on polymer and silicate or borosilicate bioactive glass. Mater. Sci. Eng. C 2020, 107, 110340. [Google Scholar] [CrossRef]
- Ojansivu, M.; Mishra, A.; Vanhatupa, S.; Juntunen, M.; Larionova, A.; Massera, J.; Miettinen, S. The effect of S53P4-based borosilicate glasses and glass dissolution products on the osteogenic commitment of human adipose stem cells. PLoS ONE 2018, 13, e0202740. [Google Scholar] [CrossRef] [Green Version]
- Salètes, M.; Vartin, M.; Mocquot, C.; Chevalier, C.; Grosgogeat, B.; Colon, P.; Attik, N. Mesoporous Bioactive Glasses Cytocompatibility Assessment: A Review of In Vitro Studies. Biomimetics 2021, 6, 9. [Google Scholar] [CrossRef]
- Mocquot, C.; Attik, N.; Pradelle-Plasse, N.; Grosgogeat, B.; Colon, P. Bioactivity assessment of bioactive glasses for dental applications: A critical review. Dent. Mater. Off. Publ. Acad. Dent. Mater. 2020, 36, 1116–1143. [Google Scholar] [CrossRef]
- Fernando, D.; Attik, N.; Cresswell, M.; Mokbel, I.; Pradelle-Plasse, N.; Jackson, P.; Grosgogeat, B.; Colon, P. Influence of network modifiers in an acetate based sol-gel bioactive glass system. Microporous Mesoporous Mater. 2018, 257, 99–109. [Google Scholar] [CrossRef]
- Wang, X.; Liu, Q.; Chen, W.; Liu, L. FGF adsorbed mesoporous bioactive glass with larger pores in enhancing bone tissue engineering. J. Mater. Sci. Mater. Med. 2019, 30, 48. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.-T.; Patel, D.-K.; Dutta, S.-D.; Ganguly, K. Fluid Flow Mechanical Stimulation-Assisted Cartridge Device for the Osteogenic Differentiation of Human Mesenchymal Stem Cells. Micromachines 2021, 12, 927. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Wei, Z.; He, S.; Bi, Y.; Cao, Y.; Wang, W. Mesoporous Bioactive Glass Scaffold Delivers Salvianolic Acid B to Promote Bone Regeneration in a Rat Cranial Defect Model. Curr. Drug Deliv. 2021, 18, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Ravichandran, A.; Liu, Y.; Teoh, S.H. Review: Bioreactor design towards generation of relevant engineered tissues: Focus on clinical translation. J. Tissue Eng. Regen. Med. 2018, 12, e7–e22. [Google Scholar] [CrossRef]
- Khalil, C.; Moussa, M.; Azar, A.; Tawk, J.; Habbouche, J.; Salameh, R.; Ibrahim, A.; Alaaeddine, N. Anti-proliferative effects of mesenchymal stem cells (MSCs) derived from multiple sources on ovarian cancer cell lines: An in-vitro experimental study. J. Ovarian Res. 2019, 12, 70. [Google Scholar] [CrossRef] [PubMed]
- Le Naour, A.; Prat, M.; Thibault, B.; Mével, R.; Lemaitre, L.; Leray, H.; Joubert, M.V.; Coulson, K.; Golzio, M.; Lefevre, L.; et al. Tumor cells educate mesenchymal stromal cells to release chemoprotective and immunomodulatory factors. J. Mol. Cell Biol. 2020, 12, 202–215. [Google Scholar] [CrossRef]
- Yi, T.; Huang, S.; Liu, G.; Li, T.; Kang, Y.; Luo, Y.; Wu, J. Bioreactor Synergy with 3D Scaffolds: New Era for Stem Cells Culture. ACS Appl. Bio Mater. 2018, 1, 193–209. [Google Scholar] [CrossRef]
- Tiomnova, O.T.; Coelho, F.; Pellizaro, T.A.G.; Enrique, J.; Chanfrau, R.; de Oliveira Capote, T.S.; Basmaji, P.; Pantoja, Y.V.; Guastaldi, A.C. Preparation of scaffolds of amorphous calcium phosphate and bacterial cellulose for use in tissue regeneration by freeze-drying process. Biointerface Res. Appl. Chem. 2021, 11, 7357–7367. [Google Scholar] [CrossRef]
- Lim, K.-T.; Patel, D.K.; Seonwoo, H.; Kim, J.; Chung, J.H. A fully automated bioreactor system for precise control of stem cell proliferation and differentiation. Biochem. Eng. J. 2019, 150, 107258. [Google Scholar] [CrossRef]
- Rimington, R.P.; Capel, A.J.; Chaplin, K.F.; Fleming, J.W.; Bandulasena, H.C.H.; Bibb, R.J.; Christie, S.D.R.; Lewis, M.P. Differentiation of Bioengineered Skeletal Muscle within a 3D Printed Perfusion Bioreactor Reduces Atrophic and Inflammatory Gene Expression. ACS Biomater. Sci. Eng. 2019, 5, 5525–5538. [Google Scholar] [CrossRef] [PubMed]
- Paez-Mayorga, J.; Hernández-Vargas, G.; Ruiz-Esparza, G.U.; Iqbal, H.M.N.; Wang, X.; Zhang, Y.S.; Parra-Saldivar, R.; Khademhosseini, A. Bioreactors for Cardiac Tissue Engineering. Adv. Healthc. Mater. 2019, 8, 1701504. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Lü, D.; Zheng, L.; Zhang, F.; Zhang, X.; Lü, S.; Zhang, C.; Jia, X.; Shu, X.; Li, P.; et al. Hepatic differentiation of human embryonic stem cells by coupling substrate stiffness and microtopography. Biomater. Sci. 2021, 9, 3776–3790. [Google Scholar] [CrossRef]
- Babaliari, E.; Kavatzikidou, P.; Mitraki, A.; Papaharilaou, Y.; Ranella, A.; Stratakis, E. Combined effect of shear stress and laser-patterned topography on Schwann cell outgrowth: Synergistic or antagonistic? Biomater. Sci. 2021, 9, 1334–1344. [Google Scholar] [CrossRef] [PubMed]
- Limongi, T.; Dattola, E.; Botta, C.; Coluccio, M.L.; Candeloro, P.; Cucè, M.; Scopacasa, B.; Gallo Cantafio, M.E.; Critello, C.D.; Pullano, S.A.; et al. Influence of the Fabrication Accuracy of Hot-Embossed PCL Scaffolds on Cell Growths. Front. Bioeng. Biotechnol. 2020, 8, 84. [Google Scholar] [CrossRef] [PubMed]
- Guneta, V.; Zhou, Z.; Tan, N.S.; Sugii, S.; Wong, M.T.C.; Choong, C. Recellularization of decellularized adipose tissue-derived stem cells: Role of the cell-secreted extracellular matrix in cellular differentiation. Biomater. Sci. 2018, 6, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Corrigan, M.A.; Johnson, G.P.; Stavenschi, E.; Riffault, M.; Labour, M.-N.; Hoey, D.A. TRPV4-mediates oscillatory fluid shear mechanotransduction in mesenchymal stem cells in part via the primary cilium. Sci. Rep. 2018, 8, 3824. [Google Scholar] [CrossRef]
- Simunovic, F.; Winninger, O.; Strassburg, S.; Koch, H.G.; Finkenzeller, G.; Stark, G.B.; Lampert, F.M. Increased differentiation and production of extracellular matrix components of primary human osteoblasts after cocultivation with endothelial cells: A quantitative proteomics approach. J. Cell. Biochem. 2019, 120, 396–404. [Google Scholar] [CrossRef] [Green Version]
- Onishi, T.; Shimizu, T.; Akahane, M.; Omokawa, S.; Okuda, A.; Kira, T.; Inagak, Y.; Tanaka, Y. Osteogenic extracellular matrix sheet for bone tissue regeneration. Eur. Cells Mater. 2018, 36, 68–80. [Google Scholar] [CrossRef]
- Mazziotta, C.; Lanzillotti, C.; Iaquinta, M.R.; Taraballi, F.; Torreggiani, E.; Rotondo, J.C.; Otòn-Gonzalez, L.; Mazzoni, E.; Frontini, F.; Bononi, I.; et al. MicroRNAs Modulate Signaling Pathways in Osteogenic Differentiation of Mesenchymal Stem Cells. Int. J. Mol. Sci. 2021, 22, 2362. [Google Scholar] [CrossRef]
- Mortada, I.; Mortada, R. Epigenetic changes in mesenchymal stem cells differentiation. Eur. J. Med. Genet. 2018, 61, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Iaquinta, M.R.; Mazzoni, E.; Bononi, I.; Rotondo, J.C.; Mazziotta, C.; Montesi, M.; Sprio, S.; Tampieri, A.; Tognon, M.; Martini, F. Adult Stem Cells for Bone Regeneration and Repair. Front. Cell Dev. Biol. 2019, 7, 268. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Li, J.; Zou, Q.; Zuo, Y.; Cai, B.; Li, Y. Enhanced bone tissue regeneration of a biomimetic cellular scaffold with co-cultured MSCs-derived osteogenic and angiogenic cells. Cell Prolif. 2019, 52, e12658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klontzas, M.E.; Reakasame, S.; Silva, R.; Morais, J.C.F.; Vernardis, S.; MacFarlane, R.J.; Heliotis, M.; Tsiridis, E.; Panoskaltsis, N.; Boccaccini, A.R.; et al. Oxidized alginate hydrogels with the GHK peptide enhance cord blood mesenchymal stem cell osteogenesis: A paradigm for metabolomics-based evaluation of biomaterial design. Acta Biomater. 2019, 88, 224–240. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Madaleno, C.; Jatzlau, J.; Knaus, P. BMP signalling in a mechanical context—Implications for bone biology. Bone 2020, 137, 115416. [Google Scholar] [CrossRef]
- Varani, K.; Vincenzi, F.; Pasquini, S.; Blo, I.; Salati, S.; Cadossi, M.; De Mattei, M. Pulsed Electromagnetic Field Stimulation in Osteogenesis and Chondrogenesis: Signaling Pathways and Therapeutic Implications. Int. J. Mol. Sci. 2021, 22, 809. [Google Scholar] [CrossRef]
- Mazzoni, E.; Mazziotta, C.; Iaquinta, M.R.; Lanzillotti, C.; Fortini, F.; D’Agostino, A.; Trevisiol, L.; Nocini, R.; Barbanti-Brodano, G.; Mescola, A.; et al. Enhanced Osteogenic Differentiation of Human Bone Marrow-Derived Mesenchymal Stem Cells by a Hybrid Hydroxylapatite/Collagen Scaffold. Front. Cell Dev. Biol. 2021, 8, 1658. [Google Scholar] [CrossRef]
- Mazzoni, E.; Iaquinta, M.R.; Lanzillotti, C.; Mazziotta, C.; Maritati, M.; Montesi, M.; Sprio, S.; Tampieri, A.; Tognon, M.; Martini, F. Bioactive Materials for Soft Tissue Repair. Front. Bioeng. Biotechnol. 2021, 9, 613787. [Google Scholar] [CrossRef]
- Sprio, S.; Dapporto, M.; Preti, L.; Mazzoni, E.; Iaquinta, M.R.; Martini, F.; Tognon, M.; Pugno, N.M.; Restivo, E.; Visai, L.; et al. Enhancement of the Biological and Mechanical Performances of Sintered Hydroxyapatite by Multiple Ions Doping. Front. Mater. 2020, 7, 224. [Google Scholar] [CrossRef]
- Yang, C.; Liu, X.; Zhao, K.; Zhu, Y.; Hu, B.; Zhou, Y.; Wang, M.; Wu, Y.; Zhang, C.; Xu, J.; et al. miRNA-21 promotes osteogenesis via the PTEN/PI3K/Akt/HIF-1α pathway and enhances bone regeneration in critical size defects. Stem Cell Res. Ther. 2019, 10, 65. [Google Scholar] [CrossRef]
- Alarçin, E.; Lee, T.Y.; Karuthedom, S.; Mohammadi, M.; Brennan, M.A.; Lee, D.H.; Marrella, A.; Zhang, J.; Syla, D.; Zhang, Y.S.; et al. Injectable shear-thinning hydrogels for delivering osteogenic and angiogenic cells and growth factors. Biomater. Sci. 2018, 6, 1604–1615. [Google Scholar] [CrossRef] [PubMed]
- Fassina, L.; Bloise, N.; Montagna, G.; Visai, L.; Mognaschi, M.E.; Benazzo, F.; Magenes, G. Biomaterials and biophysical stimuli for bone regeneration. J. Biol. Regul. Homeost. Agents 2018, 32, 41–49. [Google Scholar]
- Lanzillotti, C.; De Mattei, M.; Mazziotta, C.; Taraballi, F.; Rotondo, J.C.; Tognon, M.; Martini, F. Long Non-coding RNAs and MicroRNAs Interplay in Osteogenic Differentiation of Mesenchymal Stem Cells. Front. Cell Dev. Biol. 2021, 9, 742. [Google Scholar] [CrossRef] [PubMed]
- Ju, C.; Liu, R.; Zhang, Y.-W.; Zhang, Y.; Zhou, R.; Sun, J.; Lv, X.-B.; Zhang, Z. Mesenchymal stem cell-associated lncRNA in osteogenic differentiation. Biomed. Pharmacother. 2019, 115, 108912. [Google Scholar] [CrossRef] [PubMed]
- Cakouros, D.; Gronthos, S. Epigenetic Regulators of Mesenchymal Stem/Stromal Cell Lineage Determination. Curr. Osteoporos. Rep. 2020, 18, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Chirayil, R.; Kincaid, R.P.; Dahlke, C.; Kuny, C.V.; Dälken, N.; Spohn, M.; Lawson, B.; Grundhoff, A.; Sullivan, C.S. Identification of virus-encoded microRNAs in divergent Papillomaviruses. PLoS Pathog. 2018, 14, e1007156. [Google Scholar] [CrossRef] [Green Version]
- Rotondo, J.C.; Mazzoni, E.; Bononi, I.; Tognon, M.; Martini, F. Association between Simian Virus 40 and Human Tumors. Front. Oncol. 2019, 9, 670. [Google Scholar] [CrossRef]
- Vojtechova, Z.; Tachezy, R. The Role of miRNAs in Virus-Mediated Oncogenesis. Int. J. Mol. Sci. 2018, 19, 1217. [Google Scholar] [CrossRef] [Green Version]
- Ledda, B.; Ottaggio, L.; Izzotti, A.; Sukkar, S.G.; Miele, M. Small RNAs in eucaryotes: New clues for amplifying microRNA benefits. Cell Biosci. 2020, 10, 1. [Google Scholar] [CrossRef] [Green Version]
- Finotti, A.; Fabbri, E.; Lampronti, I.; Gasparello, J.; Borgatti, M.; Gambari, R. MicroRNAs and Long Non-coding RNAs in Genetic Diseases. Mol. Diagn. Ther. 2019, 23, 155–171. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bär, C.; Thum, T.; de Gonzalo-Calvo, D. Circulating miRNAs as mediators in cell-to-cell communication. Epigenomics 2019, 11, 111–113. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Peng, R.; Wang, J.; Qin, Z.; Xue, L. Circulating microRNAs as potential cancer biomarkers: The advantage and disadvantage. Clin. Epigenet. 2018, 10, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desantis, V.; Saltarella, I.; Lamanuzzi, A.; Melaccio, A.; Solimando, A.G.; Mariggiò, M.A.; Racanelli, V.; Paradiso, A.; Vacca, A.; Frassanito, M.A. MicroRNAs-Based Nano-Strategies as New Therapeutic Approach in Multiple Myeloma to Overcome Disease Progression and Drug Resistance. Int. J. Mol. Sci. 2020, 21, 3084. [Google Scholar] [CrossRef] [PubMed]
- Arriaga, M.A.; Ding, M.H.; Gutierrez, A.S.; Chew, S.A. The Application of microRNAs in Biomaterial Scaffold-Based Therapies for Bone Tissue Engineering. Biotechnol. J. 2019, 14, e1900084. [Google Scholar] [CrossRef] [PubMed]
- Iaquinta, M.R.; Mazzoni, E.; Manfrini, M.; D’Agostino, A.; Trevisiol, L.; Nocini, R.; Trombelli, L.; Barbanti-Brodano, G.; Martini, F.; Tognon, M. Innovative Biomaterials for Bone Regrowth. Int. J. Mol. Sci. 2019, 20, 618. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.H. MicroRNA Networks Modulate Oxidative Stress in Cancer. Int. J. Mol. Sci. 2019, 20, 4497. [Google Scholar] [CrossRef] [Green Version]
- Cipriani, C.; Colangelo, L.; Santori, R.; Renella, M.; Mastrantonio, M.; Minisola, S.; Pepe, J. The Interplay between Bone and Glucose Metabolism. Front. Endocrinol. 2020, 11, 122. [Google Scholar] [CrossRef] [Green Version]
- Ono, T.; Nakashima, T. Recent advances in osteoclast biology. Histochem. Cell Biol. 2018, 149, 325–341. [Google Scholar] [CrossRef]
- Terpos, E.; Ntanasis-Stathopoulos, I.; Dimopoulos, M.A. Myeloma bone disease: From biology findings to treatment approaches. Blood 2019, 133, 1534–1539. [Google Scholar] [CrossRef] [Green Version]
- Maeda, K.; Kobayashi, Y.; Koide, M.; Uehara, S.; Okamoto, M.; Ishihara, A.; Kayama, T.; Saito, M.; Marumo, K. The Regulation of Bone Metabolism and Disorders by Wnt Signaling. Int. J. Mol. Sci. 2019, 20, 5525. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Ehnert, S.; Rouß, M.; Häussling, V.; Aspera-Werz, R.H.; Chen, T.; Nussler, A.K. From the Clinical Problem to the Basic Research-Co-Culture Models of Osteoblasts and Osteoclasts. Int. J. Mol. Sci. 2018, 19, 2284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borciani, G.; Montalbano, G.; Baldini, N.; Cerqueni, G.; Vitale-Brovarone, C.; Ciapetti, G. Co–culture systems of osteoblasts and osteoclasts: Simulating in vitro bone remodeling in regenerative approaches. Acta Biomater. 2020, 108, 22–45. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, A.; Skottke, J.; von Witzleben, M.; Gelinsky, M. Triple Culture of Primary Human Osteoblasts, Osteoclasts and Osteocytes as an In Vitro Bone Model. Int. J. Mol. Sci. 2021, 22, 7316. [Google Scholar] [CrossRef] [PubMed]
- Prideaux, M.; Stern, A.R.; Bonewald, L.F. Isolation of Murine and Human Osteocytes. In Osteoporosis and Osteoarthritis; Humana Press: New York, NY, USA, 2021; pp. 3–13. [Google Scholar]
- Bernhardt, A.; Wolf, S.; Weiser, E.; Vater, C.; Gelinsky, M. An improved method to isolate primary human osteocytes from bone. Biomed. Tech. Biomed. Eng. 2020, 65, 107–111. [Google Scholar] [CrossRef]
- Bernhardt, A.; Österreich, V.; Gelinsky, M. Three-Dimensional Co-culture of Primary Human Osteocytes and Mature Human Osteoclasts in Collagen Gels. Tissue Eng. Part A 2020, 26, 647–655. [Google Scholar] [CrossRef]
- Vis, M.A.M.; Ito, K.; Hofmann, S. Impact of Culture Medium on Cellular Interactions in in vitro Co-culture Systems. Front. Bioeng. Biotechnol. 2020, 8, 911. [Google Scholar] [CrossRef]
- Pagani, S.; Torricelli, P.; Veronesi, F.; Salamanna, F.; Cepollaro, S.; Fini, M. An advanced tri-culture model to evaluate the dynamic interplay among osteoblasts, osteoclasts, and endothelial cells. J. Cell. Physiol. 2018, 233, 291–301. [Google Scholar] [CrossRef]
- Grémare, A.; Aussel, A.; Bareille, R.; Paiva Dos Santos, B.; Amédée, J.; Thébaud, N.B.; Le Nihouannen, D. A Unique Triculture Model to Study Osteoblasts, Osteoclasts, and Endothelial Cells. Tissue Eng. Part C Methods 2019, 25, 421–432. [Google Scholar] [CrossRef]
- Wein, F.; Bruinink, A. Human triple cell co-culture for evaluation of bone implant materials. Integr. Biol. 2013, 5, 703–711. [Google Scholar] [CrossRef]
- Plotkin, L.I.; Bruzzaniti, A. Molecular signaling in bone cells: Regulation of cell differentiation and survival. Adv. Protein Chem. Struct. Biol. 2019, 116, 237–281. [Google Scholar] [CrossRef] [PubMed]
- Scheinpflug, J.; Pfeiffenberger, M.; Damerau, A.; Schwarz, F.; Textor, M.; Lang, A.; Schulze, F. Journey into Bone Models: A Review. Genes 2018, 9, 247. [Google Scholar] [CrossRef] [Green Version]
- George, E.L.; Truesdell, S.L.; York, S.L.; Saunders, M.M. Lab-on-a-chip platforms for quantification of multicellular interactions in bone remodeling. Exp. Cell Res. 2018, 365, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Truesdell, S.L.; George, E.L.; Van Vranken, C.C.; Saunders, M.M. A Lab-On-A-Chip Platform for Stimulating Osteocyte Mechanotransduction and Analyzing Functional Outcomes of Bone Remodeling. J. Vis. Exp. JoVE 2020. [Google Scholar] [CrossRef]
- Chen, K.; Zhou, Q.; Kang, H.; Yan, Y.; Qian, N.; Li, C.; Wang, F.; Yang, K.; Deng, L.; Qi, J. High Mineralization Capacity of IDG-SW3 Cells in 3D Collagen Hydrogel for Bone Healing in Estrogen-Deficient Mice. Front. Bioeng. Biotechnol. 2020, 8, 864. [Google Scholar] [CrossRef] [PubMed]
- Nasello, G.; Alamán-Díez, P.; Schiavi, J.; Pérez, M.Á.; McNamara, L.; García-Aznar, J.M. Primary Human Osteoblasts Cultured in a 3D Microenvironment Create a Unique Representative Model of Their Differentiation Into Osteocytes. Front. Bioeng. Biotechnol. 2020, 8, 336. [Google Scholar] [CrossRef]
- Bernhardt, A.; Bacova, J.; Gbureck, U.; Gelinsky, M. Influence of Cu2+ on Osteoclast Formation and Activity In Vitro. Int. J. Mol. Sci. 2021, 22, 2451. [Google Scholar] [CrossRef]
- Vincent, M.; Duval, R.E.; Hartemann, P.; Engels-Deutsch, M. Contact killing and antimicrobial properties of copper. J. Appl. Microbiol. 2018, 124, 1032–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elrayah, A.; Zhi, W.; Feng, S.; Al-Ezzi, S.; Lei, H.; Weng, J. Preparation of Micro/Nano-Structure Copper-Substituted Hydroxyapatite Scaffolds with Improved Angiogenesis Capacity for Bone Regeneration. Materials 2018, 11, 1516. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wu, H.; He, F.; Wu, T.; Zhou, L.; Ye, J. Concentration-dependent osteogenic and angiogenic biological performances of calcium phosphate cement modified with copper ions. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 99, 1199–1212. [Google Scholar] [CrossRef]
- Zhang, M.; Lin, R.; Wang, X.; Xue, J.; Deng, C.; Feng, C.; Zhuang, H.; Ma, J.; Qin, C.; Wan, L.; et al. 3D printing of Haversian bone–mimicking scaffolds for multicellular delivery in bone regeneration. Sci. Adv. 2020, 6, eaaz6725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortini, M.; Baldini, N.; Avnet, S. New advances in the study of bone tumors: A lesson from the 3D environment. Front. Physiol. 2019, 10, 814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauli, C.; Hopkins, B.D.; Prandi, D.; Shaw, R.; Fedrizzi, T.; Sboner, A.; Sailer, V.; Augello, M.; Puca, L.; Rosati, R. Personalized in vitro and in vivo cancer models to guide precision medicine. Cancer Discov. 2017, 7, 462–477. [Google Scholar] [CrossRef] [Green Version]
- Hadida, M.; Marchat, D. Strategy for achieving standardized bone models. Biotechnol. Bioeng. 2020, 117, 251–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Wang, Z.; Duan, N.; Zhu, G.; Schwarz, E.M.; Xie, C. Osteoblast-osteoclast interactions. Connect. Tissue Res. 2018, 59, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Agidigbi, T.S.; Kim, C. Reactive Oxygen Species in Osteoclast Differentiation and Possible Pharmaceutical Targets of ROS-Mediated Osteoclast Diseases. Int. J. Mol. Sci. 2019, 20, 3576. [Google Scholar] [CrossRef] [Green Version]
- Collins, J.A.; Diekman, B.O.; Loeser, R.F. Targeting aging for disease modification in osteoarthritis. Curr. Opin. Rheumatol. 2018, 30, 101–107. [Google Scholar] [CrossRef]
- Schröder, K. NADPH oxidases in bone homeostasis and osteoporosis. Free Radic. Biol. Med. 2019, 132, 67–72. [Google Scholar] [CrossRef]
- Ohyama, Y.; Ito, J.; Kitano, V.J.; Shimada, J.; Hakeda, Y. The polymethoxy flavonoid sudachitin suppresses inflammatory bone destruction by directly inhibiting osteoclastogenesis due to reduced ROS production and MAPK activation in osteoclast precursors. PLoS ONE 2018, 13, e0191192. [Google Scholar] [CrossRef] [Green Version]
- Wojsiat, J.; Zoltowska, K.M.; Laskowska-Kaszub, K.; Wojda, U. Oxidant/Antioxidant Imbalance in Alzheimer’s Disease: Therapeutic and Diagnostic Prospects. Oxidative Med. Cell. Longev. 2018, 2018, 6435861. [Google Scholar] [CrossRef] [Green Version]
- Gaucher, C.; Boudier, A.; Bonetti, J.; Clarot, I.; Leroy, P.; Parent, M. Glutathione: Antioxidant Properties Dedicated to Nanotechnologies. Antioxidants 2018, 7, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, H.; Zhen, C.; Liu, J.; Yang, P.; Hu, L.; Shang, P. Unraveling the Potential Role of Glutathione in Multiple Forms of Cell Death in Cancer Therapy. Oxidative Med. Cell. Longev. 2019, 2019, 3150145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Wang, G.; Gong, L.; Sun, G.; Shi, B.; Bao, H.; Duan, Y. Isopsoralen regulates PPAR-γ/WNT to inhibit oxidative stress in osteoporosis. Mol. Med. Rep. 2018, 17, 1125–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, H.; Ochi, M.; Ono, M.; Aoyama, E.; Ogino, T.; Kondo, Y.; Ohuchi, H. Glutathione accelerates osteoclast differentiation and inflammatory bone destruction. Free Radic. Res. 2019, 53, 226–236. [Google Scholar] [CrossRef]
- Mu, P.; Hu, Y.; Ma, X.; Shi, J.; Zhong, Z.; Huang, L. Total flavonoids of Rhizoma Drynariae combined with calcium attenuate osteoporosis by reducing reactive oxygen species generation. Exp. Ther. Med. 2021, 21, 618. [Google Scholar] [CrossRef] [PubMed]
- Shieh, P.; Jan, C.R.; Liang, W.Z. The protective effects of the antioxidant N-acetylcysteine (NAC) against oxidative stress-associated apoptosis evoked by the organophosphorus insecticide malathion in normal human astrocytes. Toxicology 2019, 417, 1–14. [Google Scholar] [CrossRef]
- Chen, L.; Wang, G.; Wang, Q.; Liu, Q.; Sun, Q.; Chen, L. N-acetylcysteine prevents orchiectomy-induced osteoporosis by inhibiting oxidative stress and osteocyte senescence. Am. J. Transl. Res. 2019, 11, 4337–4347. [Google Scholar]
- Raffaele, M.; Barbagallo, I.; Licari, M.; Carota, G.; Sferrazzo, G.; Spampinato, M.; Sorrenti, V.; Vanella, L. N-Acetylcysteine (NAC) Ameliorates Lipid-Related Metabolic Dysfunction in Bone Marrow Stromal Cells-Derived Adipocytes. Evid.-Based Complement. Altern. Med. 2018, 2018, 5310961. [Google Scholar] [CrossRef]
- Dludla, P.V.; Dias, S.C.; Obonye, N.; Johnson, R.; Louw, J.; Nkambule, B.B. A Systematic Review on the Protective Effect of N-Acetyl Cysteine Against Diabetes-Associated Cardiovascular Complications. Am. J. Cardiovasc. Drugs Drugs Devices Other Interv. 2018, 18, 283–298. [Google Scholar] [CrossRef]
- Muszyńska, M.; Ambrożewicz, E.; Gęgotek, A.; Grynkiewicz, G.; Skrzydlewska, E. Protective Effects of Vitamin K Compounds on the Proteomic Profile of Osteoblasts under Oxidative Stress Conditions. Molecules 2020, 25, 1990. [Google Scholar] [CrossRef]
- Ambrożewicz, E.; Muszyńska, M.; Tokajuk, G.; Grynkiewicz, G.; Žarković, N.; Skrzydlewska, E. Beneficial Effects of Vitamins K and D3 on Redox Balance of Human Osteoblasts Cultured with Hydroxyapatite-Based Biomaterials. Cells 2019, 8, 325. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Luo, R.; Yang, W.; Zhou, Z.; Li, C. miR-363-3p is activated by MYB and regulates osteoporosis pathogenesis via PTEN/PI3K/AKT signaling pathway. In vitro cellular & developmental biology. Animal 2019, 55, 376–386. [Google Scholar] [CrossRef]
- Zhang, Y.; Cao, X.; Li, P.; Fan, Y.; Zhang, L.; Li, W.; Liu, Y. PSMC6 promotes osteoblast apoptosis through inhibiting PI3K/AKT signaling pathway activation in ovariectomy-induced osteoporosis mouse model. J. Cell. Physiol. 2020, 235, 5511–5524. [Google Scholar] [CrossRef] [PubMed]
- Detsch, R.; Boccaccini, A.R. The role of osteoclasts in bone tissue engineering. J. Tissue Eng. Regen. Med. 2015, 9, 1133–1149. [Google Scholar] [CrossRef]
- Cao, W.; Helder, M.N.; Bravenboer, N.; Wu, G.; Jin, J.; Ten Bruggenkate, C.M.; Klein-Nulend, J.; Schulten, E.A.J.M. Is There a Governing Role of Osteocytes in Bone Tissue Regeneration? Curr. Osteoporos. Rep. 2020, 18, 541–550. [Google Scholar] [CrossRef]
- Zhang, W.; Zhu, C.; Wu, Y.; Ye, D.; Wang, S.; Zou, D.; Zhang, X.; Kaplan, D.L.; Jiang, X. VEGF and BMP-2 promote bone regeneration by facilitating bone marrow stem cell homing and differentiation. Eur. Cells Mater. 2014, 27, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Aghadavod, E.; Khodadadi, S.; Baradaran, A.; Nasri, P.; Bahmani, M.; Rafieian-Kopaei, M. Role of Oxidative Stress and Inflammatory Factors in Diabetic Kidney Disease. Iran. J. Kidney Dis. 2016, 10, 337–343. [Google Scholar]
- Murahashi, Y.; Yano, F.; Nakamoto, H.; Maenohara, Y.; Iba, K.; Yamashita, T.; Tanaka, S.; Ishihara, K.; Okamura, Y.; Moro, T.; et al. Multi-layered PLLA-nanosheets loaded with FGF-2 induce robust bone regeneration with controlled release in critical-sized mouse femoral defects. Acta Biomater. 2019, 85, 172–179. [Google Scholar] [CrossRef]
- Song, K.; Rao, N.J.; Chen, M.L.; Huang, Z.J.; Cao, Y.G. Enhanced bone regeneration with sequential delivery of basic fibroblast growth factor and sonic hedgehog. Injury 2011, 42, 796–802. [Google Scholar] [CrossRef] [PubMed]
- Starlinger, J.; Sarahrudi, K.; Kecht, M.; Koerbler, F.; Pietschmann, P.; Aharinejad, S. The influence of M-CSF on fracture healing in a mouse model. Sci. Rep. 2021, 11, 22326. [Google Scholar] [CrossRef]
- Zarins, J.; Pilmane, M.; Sidhoma, E.; Salma, I.; Locs, J. The Role of Strontium Enriched Hydroxyapatite and Tricalcium Phosphate Biomaterials in Osteoporotic Bone Regeneration. Symmetry 2019, 11, 229. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Advantages and disadvantages of vascularization strategies. Created based on information from [7].
Figure 1.
Advantages and disadvantages of vascularization strategies. Created based on information from [7].


Figure 3.
Representation of sources of MSCs and the differentiation in cell lines. Adapted from an open-access source [91].
Figure 3.
Representation of sources of MSCs and the differentiation in cell lines. Adapted from an open-access source [91].

Figure 4.
The signaling pathway for osteoclasts differentiation (adapted from an open-access source) [148].
Figure 4.
The signaling pathway for osteoclasts differentiation (adapted from an open-access source) [148].

Figure 5.
The fields involved in BTE.


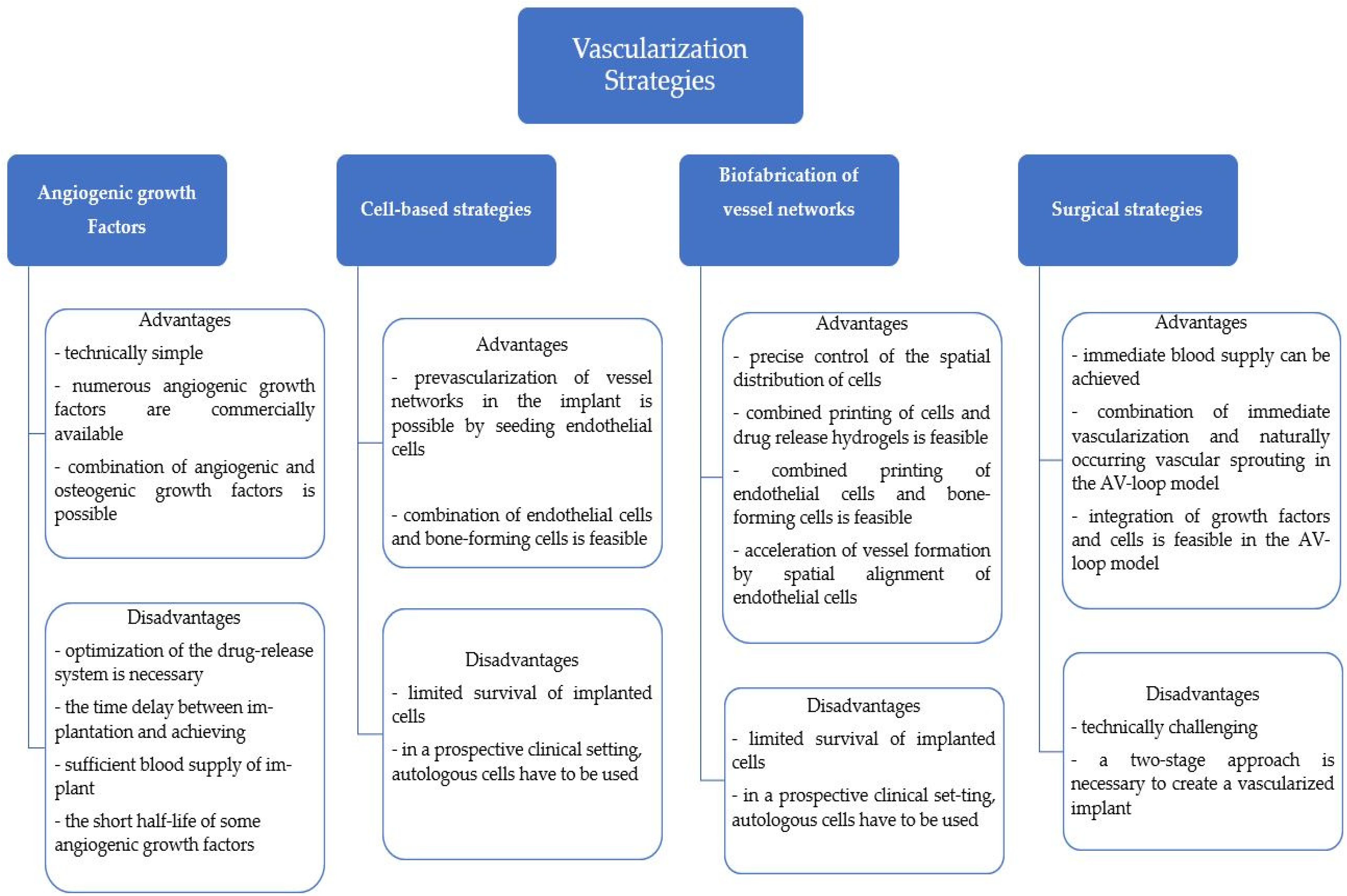{kind=link}
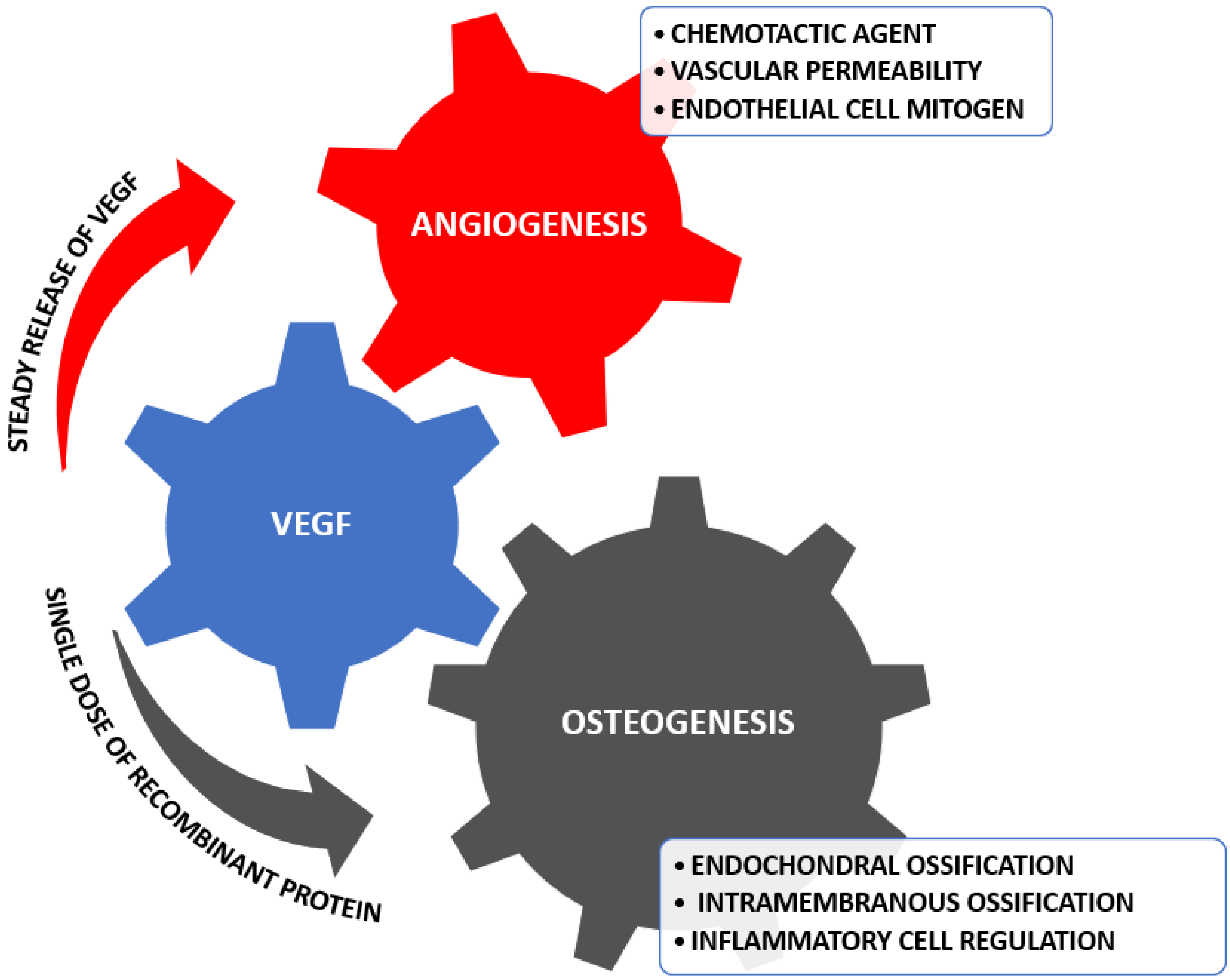{kind=link}
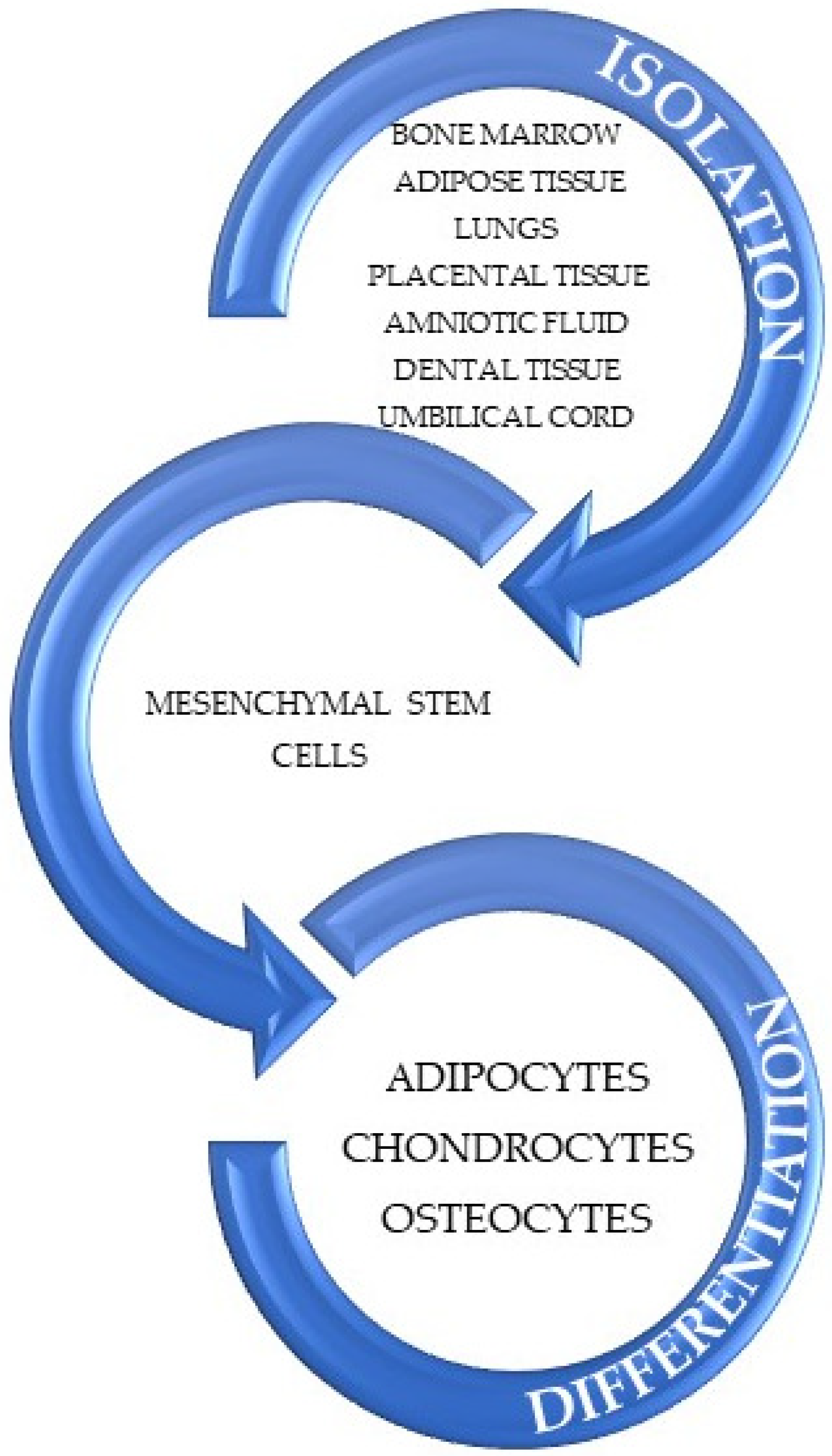{kind=link}
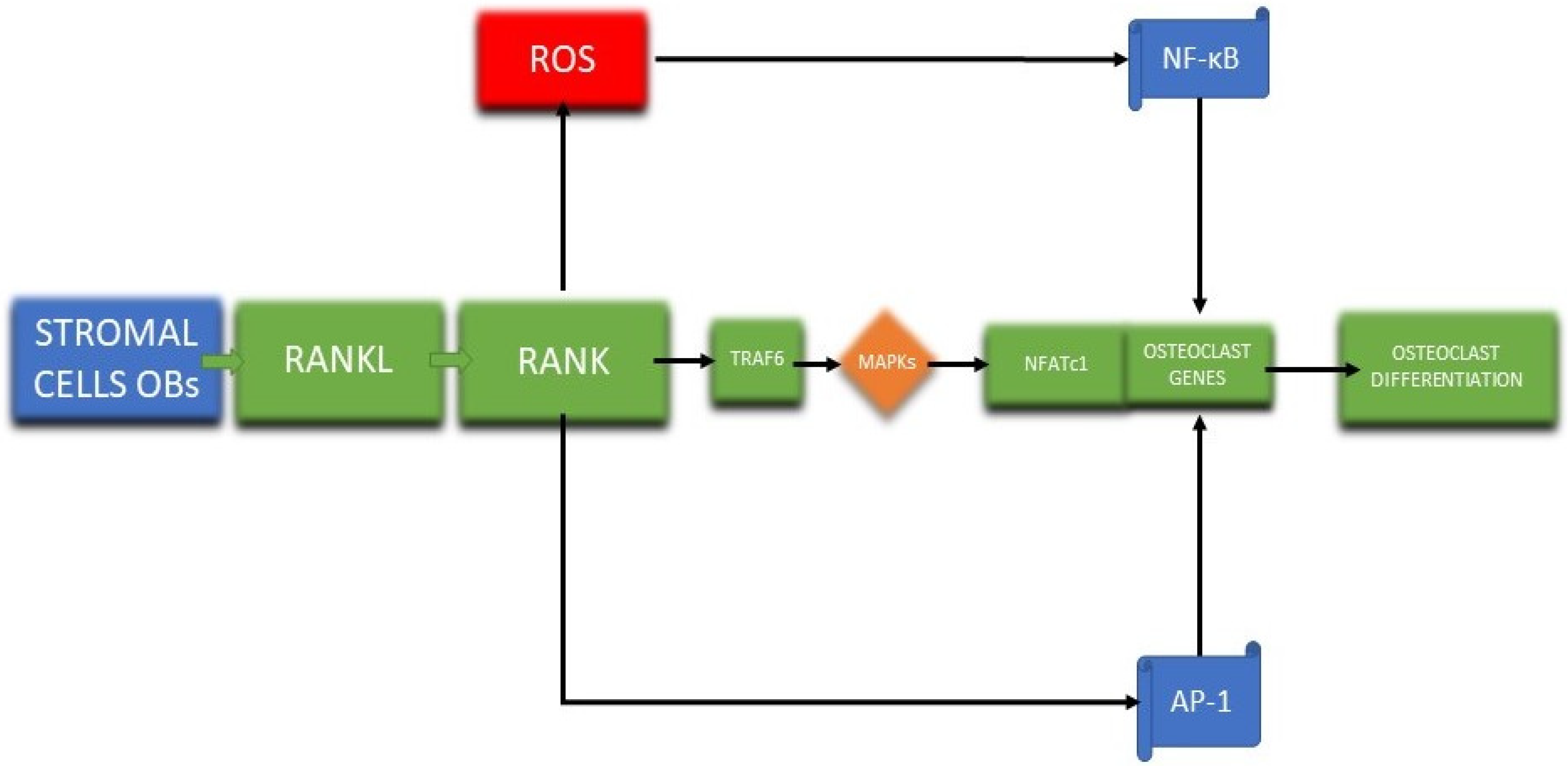{kind=link}
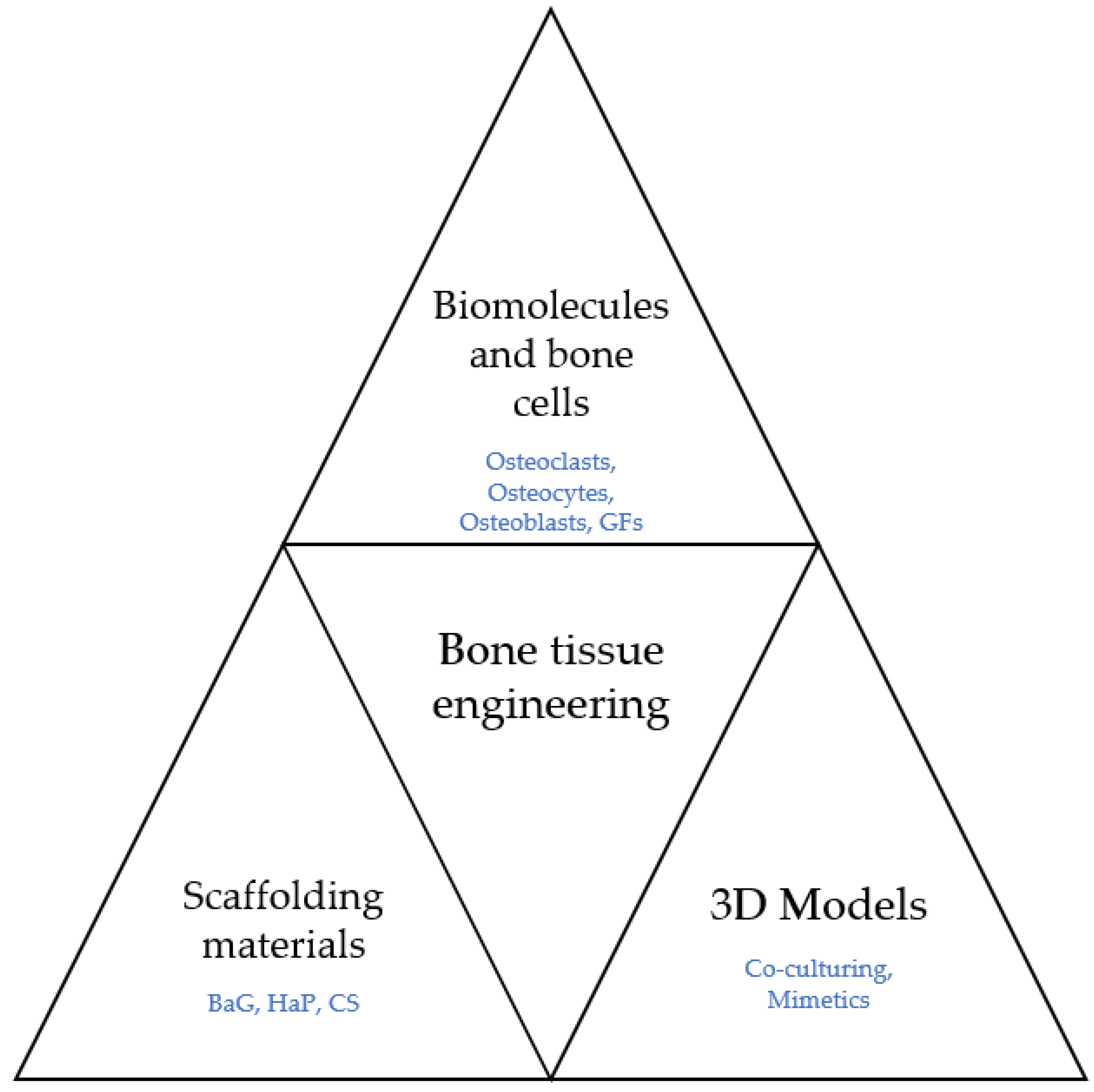{kind=link}
Table 1.
Advantages and limitations of the culturing techniques. Created based on information from [123,128].
| Culturing Type | Particularities | Observations | Advantages | Limitations |
|---|---|---|---|---|
| Triple culture of osteoblasts, osteoclasts, and osteocytes | • There are two techniques for seeding: one done individually called patterned seeding and the other being called mixed seeding | • Patterned seeding resulted in a better individual analysis • Mixed seeding generated better direct cell-cell contact | • Realistic model • Allows the study of the interactions of the cells • No major morphological differences • The ratio of cells to substance increases | • The models are not cultivated on resorbable membranes • Different serum concentrations are required for differentiation • Requires more substances and space |
| Co-cultured approaches with two different species of bone cells | • The test was realized with a porous membrane situated between the two cultures promoting the single cell type analysis | • The osteoclast gels present poor strength in comparison with the single culture underlining the degrading osteoblastic enzyme outcome | • Analytical aim of crosstalking cultures • Cost-effective | • The resorption capacities of the cultures are yet to be determined • Fewer variation opportunities resulting in isolated outcomes |
Table 2.
Examples of clinical studies in the domain of BTE.
| Clinical Trials • gov• Identifier | Official Title | Purpose of the Study | Data Availability |
|---|---|---|---|
| NCT03652753 | Pilon Fracture With Intra-articular Injection of N-Acetylcysteine (Pilon NAC) | Analyzation of the outcomes obtained from the amino acid NAC on cartilage cells in cases of intra-articular fracture of the ankle joint. | Estimated primary completion date: January 2023 Estimated study completion date: January 2024 |
| NCT03024008 | Enhancement of Bone Regeneration and Healing in the Extremities by the Use of Autologous BonoFill-II | Evaluation of the safety of an autologous bone-regenerating graft for the reconstruction of deficient bone. | Estimated primary completion date: June 2022 Estimated study completion date: September 2022 |
| NCT04498715 | Trochanteric Femur Fracture Operated With Dynamic Hip Screw System (DHS) Augmented With a Biphasic Apatite Sulphate Combined With Systemic or Local Bisphosphonate | Investigating the bone regeneration processes for a metal device utilizing bone substitute cement and bisphosphonate. | Estimated primary completion date: February 2021 Estimated study completion date: August 2021 |
| NCT02171104 | MT2013-31: Allo HCT for Metabolic Disorders and Severe Osteopetrosis | The assessment of the capacity to generate donor hematopoietic engraftment without growth in transplant-related mortality for patients with severe osteopetrosis and inherited metabolic disorders. | Estimated primary completion date: September 2021 Estimated study completion date: December 2021 |
| NCT04875767 | Cartilage Repair Using a Hyaluronic Acid-Based Scaffold With Bone Marrow Aspirate Compared With Microfracture for Focal Articular Cartilage Damage of the Hip (CHASE) | Investigating the results of the procedure for a 24-month timespan post-surgery in order to determine if any complications will occur. | First posted: 6 May 2021 Estimated study completion date: 31 December 2026 |
Table 3.
Overview of relevant elements for bone regeneration and oxidative stress.
| Key Elements | Roles/Effects in Bone Regeneration and Oxidative Stress | Refs. | |
|---|---|---|---|
| Category | Representatives | ||
| Cells | Osteoblasts | Can promote calcium salts deposition in the bone matrix, leading to bone reconstruction Their increased production is associated with improved fracture healing time Secrete OPG | [105,166] |
| Osteoclasts | Responsible for bone resorption Involved in bone remodeling, which is an essential process for regeneration of bone defects Express M-CSF and RANKL Their increased cellular activity has been associated with bone diseases, such as osteoporosis, rheumatoid arthritis, and osteoarthritis | [119,148,166] | |
| Osteocytes | Regulate osteoclast and osteoblast activity Optimally used and stimulated osteocytes lead to improved bone regeneration Can enhance osteogenesis of stem cells Feedback from osteocytes limits the size of the bone-forming unit by the secretion of sclerostin | [150,167] | |
| MSCs | May differentiate into osteocytes Generate osteoclastogenic cytokines, RANKL, and M-CSF Inducing MSC osteogenesis promotes bone formation and bone regeneration | [77,91,105] | |
| Growth factors, biomolecules, and signal pathways | VEGF | Regulates osteoclast activity Modulates angiogenesis and osteogenesis Facilitates MSCs homing and differentiation Elevates intracellular ROS level | [36,37,166,168,169] |
| FGF | Active role in bone repair process Modulates osteoblasts differentiation Enhances bone regeneration in bone defects and clinical fractures | [13,170] | |
| BMP | Regulator of cartilage and bone formation Modulates osteoblasts differentiation Facilitates MSCs homing and differentiation | [13,105,168] | |
| Shh | Upregulates BMPs Modulates osteoblasts differentiation Stimulates a cascade of downstream genes involved in bone development Enhances regeneration in bone defects | [13,171] | |
| M-CSF | Modulates osteoclasts differentiation Role in recruiting stem cells to the fracture site Impacts hard callus formation | [13,172] | |
| RANKL | Modulates osteoclasts differentiation Can enhance osteoclastogenesis and improve bone remodeling when added to biomaterials Can induce ROS formation | [13,150,166] | |
| OPG | Antagonist receptor for RANKL Decreases osteoclast formation and bone resorption activity | [166,173] | |
| NOX | One of the main sources of ROS NOX2-derived O2− enhances RANKL-induced NFATc1 expression in osteoclast signalling; NOX2 inhibition exerts protective effects and may prevent bone loss NOX4 contribues to osteoclastogenesis and bone homeostasis; NOX4 is involved in bone loss, representing a potential therapeutic target for osteoporosis treatment | [148] | |
| P53 | Regulates cell cycle, apoptosis, growth, and metabolism of target genes Master regulator in the cellular response to oxidative stress Reduces intracellular ROS levels by promoting antioxidant reactions | [118] | |
| miRNA | Modulates osteogenic differentiation and general transcription factors Can regulate processes such as oxidative stress, aging, and apoptosis Involved in the generation of ROS Several miRNAs are involved in cellular senescence | [101,118] | |
| Scaffolds | Hydrogels | Delivery and controlled release of growth factors that aid neovascularization Enhance proliferation of HUVEC when incorporated with these cells Support co-culturing of bone cells | [21,34,123] |
| Bioceramics | Their microstructure promotes ossification and vascularization growth Well-defined pore architecture improves cell seeding efficiency, cell viability, migration, proliferation, and differentiation, enhancing bone regeneration Can be functionalized with growth factors that aid in osteogenesis and angiogenesis | [25,34,35,49] | |
| Bioactive glass | Enhances bone regeneration during longer periods of time Involved in the regulatory process of endothelial markers Material porosity influences remineralization | [68,69,74] | |
Abbreviations: MSCs—mesenchymal stem cells; VEGF—vascular endothelial growth factor; FGF—fibroblast growth factor; BMP—bone morphogenic protein; Shh—sonic hedgehog; M-CSF—macrophage colony-stimulating factor; RANKL—receptor activator of nuclear factor kappa-B ligand; OPG—osteoprotegerin; NOX—NADPH oxidase; NFATc1—nuclear factor of activated T cells 1; HUVEC—human umbilical vein endothelial cells.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bădilă, A.E.; Rădulescu, D.M.; Ilie, A.; Niculescu, A.-G.; Grumezescu, A.M.; Rădulescu, A.R. Bone Regeneration and Oxidative Stress: An Updated Overview. Antioxidants 2022, 11, 318. https://doi.org/10.3390/antiox11020318
AMA Style
Bădilă AE, Rădulescu DM, Ilie A, Niculescu A-G, Grumezescu AM, Rădulescu AR. Bone Regeneration and Oxidative Stress: An Updated Overview. Antioxidants. 2022; 11(2):318. https://doi.org/10.3390/antiox11020318
Chicago/Turabian StyleBădilă, Adrian Emilian, Dragos Mihai Rădulescu, Andrei Ilie, Adelina-Gabriela Niculescu, Alexandru Mihai Grumezescu, and Adrian Radu Rădulescu. 2022. "Bone Regeneration and Oxidative Stress: An Updated Overview" Antioxidants 11, no. 2: 318. https://doi.org/10.3390/antiox11020318
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.