Dose-Dependent Sorafenib-Induced Immunosuppression Is Associated with Aberrant NFAT Activation and Expression of PD-1 in T Cells

 ,
,
Abstract
:1. Introduction
2. Results
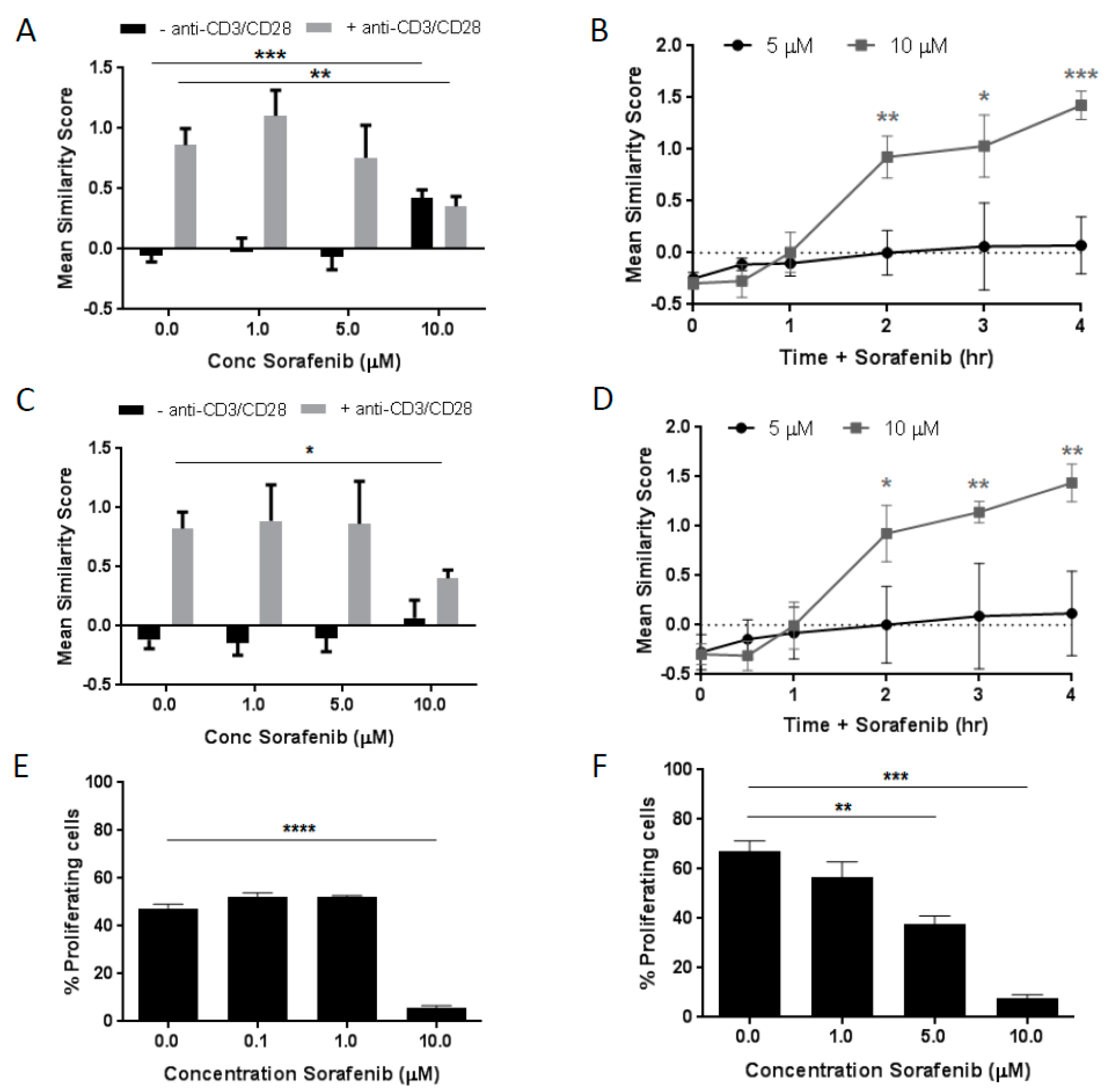
2.1. Dose- and Time-Dependent Effects of Sorafenib on NFAT1 Nuclear Translocation in T Cells
2.2. Dose-Dependent Effects of Sorafenib on T Cell Proliferation
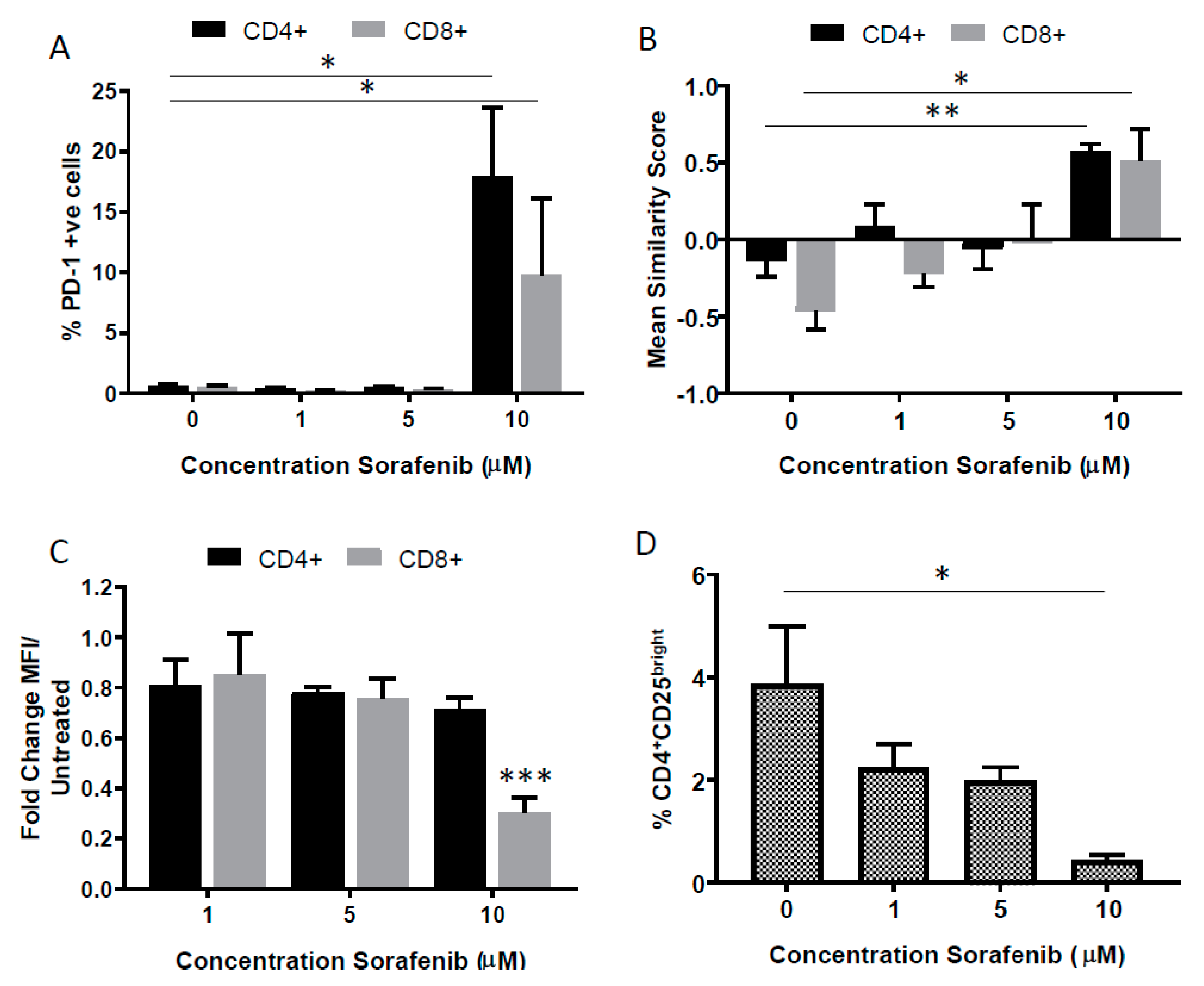
2.3. Effects of High Dose Sorafenib on PD-1 Expression on T Cells
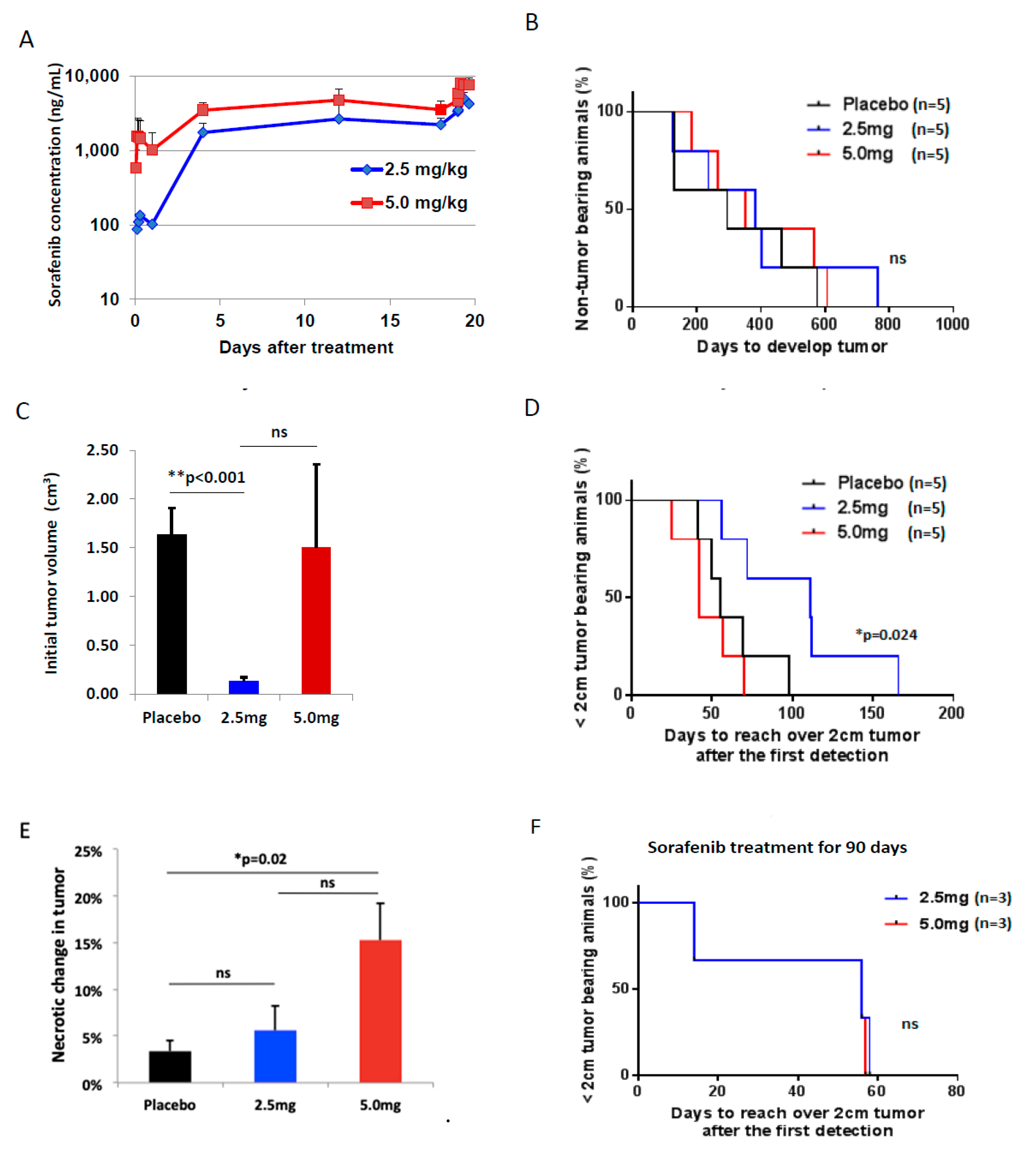
2.4. Pharmacokinetics with Different Doses of Sorafenib Treatment
2.5. Toxicity of Sorafenib Treatment In Vivo Model of HCC
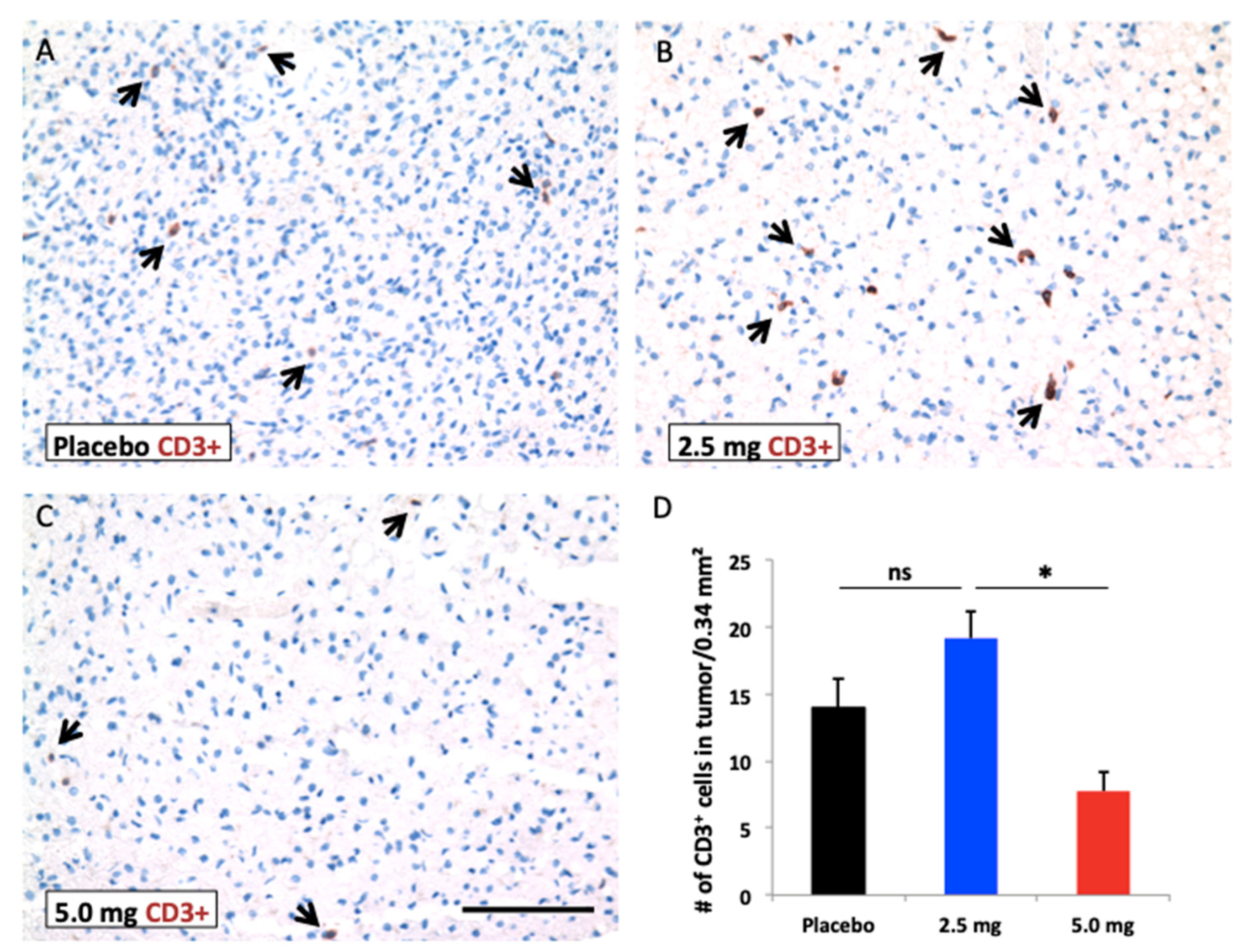
2.6. Efficacy of Sorafenib Against HCC
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Isolation of Peripheral Blood Mononuclear Cells
4.3. Analysis of NFAT1 Nuclear Translocation Activity
4.4. Imaging Flow Cytometry
4.5. Flow Cytometry
4.6. T Cell Responses in Mixed Lymphocyte Reactions (MLRs)
4.7. Dosing Solution Preparation
4.7.1. Long-Term Efficacy Study
4.7.2. Short-Term Efficacy Study
4.8. LC-MS/MS and Noncompartmental Analyses
4.9. Animal Tumor Model and Screening of Tumors
4.10. In Vivo Study Procedures
4.11. In Vivo Animal Dosing with Sorafenib
4.12. Assessment of Treatment Toxicity
4.13. Assessment of Tumor Responses to Treatment
Immunohistochemistry (IHC) and Histological Staining
4.14. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef]
- Surveillance Research Program. National Cancer Institute Fact Stata: An interactive Tool for Access to SEER Cancer Statistics. Available online: http://seer.cancer.gov/faststats (accessed on 20 September 2017).
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef]
- Bruix, J.; Han, K.H.; Gores, G.; Llovet, J.M.; Mazzaferro, V. Liver cancer: Approaching a personalized care. J. Hepatol. 2015, 62, S144–S156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, L.C.; Cheng, A.L.; Poon, R.T. Recent advances in the prevention of hepatocellular carcinoma recurrence. Semin. Liver Dis. 2014, 34, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Adnane, L.; Trail, P.A.; Taylor, I.; Wilhelm, S.M. Sorafenib (BAY 43-9006, Nexavar), a dual-action inhibitor that targets RAF/MEK/ERK pathway in tumor cells and tyrosine kinases VEGFR/PDGFR in tumor vasculature. Methods Enzymol. 2006, 407, 597–612. [Google Scholar]
- Wilhelm, S.; Carter, C.; Lynch, M.; Lowinger, T.; Dumas, J.; Smith, R.A.; Schwartz, B.; Simantov, R.; Kelley, S. Discovery and development of sorafenib: A multikinase inhibitor for treating cancer. Nat. Rev. Drug Discov. 2006, 5, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, S.M.; Carter, C.; Tang, L.; Wilkie, D.; McNabola, A.; Rong, H.; Chen, C.; Zhang, X.; Vincent, P.; McHugh, M.; et al. BAY 43-9006 exhibits broad spectrum oral antitumor activity and targets the RAF/MEK/ERK pathway and receptor tyrosine kinases involved in tumor progression and angiogenesis. Cancer Res. 2004, 64, 7099–7109. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.L.; Yan, B.S.; Lu, W.C.; Chen, M.H.; Yu, S.L.; Yang, P.C.; Cheng, A.L. Sorafenib relieves cell-intrinsic and cell-extrinsic inhibitions of effector T cells in tumor microenvironment to augment antitumor immunity. Int. J. Cancer 2014, 134, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, R.; Ararat, M.; Xu, Y.; Brusko, T.; Wasserfall, C.; Atkinson, M.A.; Chang, L.J.; Liu, C.; Nelson, D.R. Immune modulation of effector CD4+ and regulatory T cell function by sorafenib in patients with hepatocellular carcinoma. Cancer Immunol. Immunother. 2013, 62, 737–746. [Google Scholar] [CrossRef]
- Houben, R.; Voigt, H.; Noelke, C.; Hofmeister, V.; Becker, J.C.; Schrama, D. MAPK-independent impairment of T-cell responses by the multikinase inhibitor sorafenib. Mol. Cancer Ther. 2009, 8, 433–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florcken, A.; Takvorian, A.; Van Lessen, A.; Singh, A.; Hopfenmuller, W.; Dorken, B.; Pezzutto, A.; Westermann, J. Sorafenib, but not sunitinib, induces regulatory T cells in the peripheral blood of patients with metastatic renal cell carcinoma. Anticancer Drugs 2012, 23, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.C.; Huang, W.P.; Liu, C.L.; Lee, J.J.; Liu, T.P.; Ko, W.C.; Huang, Y.C.; Hsu, M.L.; Wu, C.H.; Chen, Y.J. Sorafenib induces autophagy in human myeloid dendritic cells and prolongs survival of skin allografts. Transplantation 2013, 95, 791–800. [Google Scholar] [CrossRef] [PubMed]
- Ionescu, A.G.; Cazacu, S.M.; Streba, C.T.; Fortofoiu, M.C.; Ciurea, M.E.; Ionescu, M.; Rogoveanu, O.; Comanescu, V.; Firu, S.G.; Vere, C.C. Interrelations between hepatic stellate cells and immune system cells in patients with hepatocellular carcinoma. Rom. J. Morphol. Embryol. 2015, 56, 481–490. [Google Scholar] [PubMed]
- Hogan, P.G.; Chen, L.; Nardone, J.; Rao, A. Transcriptional regulation by calcium, calcineurin, and NFAT. Genes Dev. 2003, 17, 2205–2232. [Google Scholar] [CrossRef] [Green Version]
- Macian, F. NFAT proteins: Key regulators of T-cell development and function. Nat. Rev. Immunol. 2005, 5, 472–484. [Google Scholar] [CrossRef] [PubMed]
- Shou, J.; Jing, J.; Xie, J.; You, L.; Jing, Z.; Yao, J.; Han, W.; Pan, H. Nuclear factor of activated T cells in cancer development and treatment. Cancer Lett. 2015, 361, 174–184. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Kang, X.; Cao, S.; Cheng, H.; Wang, D.; Geng, J. Calcineurin/NFATc1 pathway contributes to cell proliferation in hepatocellular carcinoma. Dig. Dis. Sci. 2012, 57, 3184–3188. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Shu, P.; Zou, S.; Shen, X.; Qu, Y.; Zhang, Y.; Sun, K.; Zhang, J. NFATc1 is a tumor suppressor in hepatocellular carcinoma and induces tumor cell apoptosis by activating the FasL-mediated extrinsic signaling pathway. Cancer Med. 2018, 7, 4701–4717. [Google Scholar] [CrossRef]
- Fric, J.; Zelante, T.; Wong, A.Y.; Mertes, A.; Yu, H.B.; Ricciardi-Castagnoli, P. NFAT control of innate immunity. Blood 2012, 120, 1380–1389. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, S.; Avni, O.; Rao, A. Cell-type-restricted binding of the transcription factor NFAT to a distal IL-4 enhancer in vivo. Immunity 2000, 12, 643–652. [Google Scholar] [CrossRef]
- Oum, J.H.; Han, J.; Myung, H.; Hleb, M.; Sharma, S.; Park, J. Molecular mechanism of NFAT family proteins for differential regulation of the IL-2 and TNF-alpha promoters. Mol. Cells 2002, 13, 77–84. [Google Scholar] [PubMed]
- Muller, M.R.; Rao, A. NFAT, immunity and cancer: A transcription factor comes of age. Nat. Rev. Immunol. 2010, 10, 645–656. [Google Scholar] [CrossRef]
- Abe, B.T.; Shin, D.S.; Mocholi, E.; Macian, F. NFAT1 supports tumor-induced anergy of CD4(+) T cells. Cancer Res. 2012, 72, 4642–4651. [Google Scholar] [CrossRef]
- Fehr, T.; Lucas, C.L.; Kurtz, J.; Onoe, T.; Zhao, G.; Hogan, T.; Vallot, C.; Rao, A.; Sykes, M. A CD8 T cell-intrinsic role for the calcineurin-NFAT pathway for tolerance induction in vivo. Blood 2010, 115, 1280–1287. [Google Scholar] [CrossRef]
- Martinez, G.J.; Pereira, R.M.; Aijo, T.; Kim, E.Y.; Marangoni, F.; Pipkin, M.E.; Togher, S.; Heissmeyer, V.; Zhang, Y.C.; Crotty, S.; et al. The transcription factor NFAT promotes exhaustion of activated CD8(+) T cells. Immunity 2015, 42, 265–278. [Google Scholar] [CrossRef]
- Werneck, M.B.; Vieira-de-Abreu, A.; Chammas, R.; Viola, J.P. NFAT1 transcription factor is central in the regulation of tissue microenvironment for tumor metastasis. Cancer Immunol. Immunother. 2011, 60, 537–546. [Google Scholar] [CrossRef]
- Barboza, B.A.; Fonseca, B.P.; Viola, J.P. NFAT1 transcription factor in dendritic cells is required to modulate T helper cell differentiation. Immunobiology 2014, 219, 704–712. [Google Scholar] [CrossRef]
- Tennant, B.C.; Toshkov, I.A.; Peek, S.F.; Jacob, J.R.; Menne, S.; Hornbuckle, W.E.; Schinazi, R.D.; Korba, B.E.; Cote, P.J.; Gerin, J.L. Hepatocellular carcinoma in the woodchuck model of hepatitis B virus infection. Gastroenterology 2004, 127, S283–S293. [Google Scholar] [CrossRef] [PubMed]
- Jain, L.; Woo, S.; Gardner, E.R.; Dahut, W.L.; Kohn, E.C.; Kummar, S.; Mould, D.R.; Giaccone, G.; Yarchoan, R.; Venitz, J.; et al. Population pharmacokinetic analysis of sorafenib in patients with solid tumours. Br. J. Clin. Pharmacol. 2011, 72, 294–305. [Google Scholar] [CrossRef] [Green Version]
- Strumberg, D.; Richly, H.; Hilger, R.A.; Schleucher, N.; Korfee, S.; Tewes, M.; Faghih, M.; Brendel, E.; Voliotis, D.; Haase, C.G.; et al. Phase I clinical and pharmacokinetic study of the Novel Raf kinase and vascular endothelial growth factor receptor inhibitor BAY 43-9006 in patients with advanced refractory solid tumors. J. Clin. Oncol. 2005, 23, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Llovet, J.M.; Ricci, S.; Mazzaferro, V.; Hilgard, P.; Gane, E.; Blanc, J.F.; de Oliveira, A.C.; Santoro, A.; Raoul, J.L.; Forner, A.; et al. Sorafenib in advanced hepatocellular carcinoma. N. Engl. J. Med. 2008, 359, 378–390. [Google Scholar] [CrossRef] [PubMed]
- Bruix, J.; Takayama, T.; Mazzaferro, V.; Chau, G.Y.; Yang, J.; Kudo, M.; Cai, J.; Poon, R.T.; Han, K.H.; Tak, W.Y.; et al. Adjuvant sorafenib for hepatocellular carcinoma after resection or ablation (STORM): A phase 3, randomised, double-blind, placebo-controlled trial. Lancet Oncol. 2015, 16, 1344–1354. [Google Scholar] [CrossRef]
- Lei, C.J.; Liu, J.N.; Wu, R.; Long, Z.X.; Zhang, J.Z.; Tao, D.; Liu, Y.P. Change of the peripheral blood immune pattern and its correlation with prognosis in patients with liver cancer treated by sorafenib. Asian Pac. J. Trop. Med. 2016, 9, 592–596. [Google Scholar] [CrossRef] [Green Version]
- Abe, B.T.; Macian, F. Uncovering the mechanisms that regulate tumor-induced T-cell anergy. Oncoimmunology 2013, 2, e22679. [Google Scholar] [CrossRef] [Green Version]
- Yarchoan, M.; Xing, D.; Luan, L.; Xu, H.; Sharma, R.; Popovic, A.; Pawlik, T.M.; Kim, A.K.; Zhu, Q.; Jaffee, E.M.; et al. Characterization of the Immune Microenvironment in Hepatocellular Carcinoma (HCC). Clin. Cancer Res. 2017, 23, 7333–7339. [Google Scholar] [CrossRef]
- He, G.; Zhang, H.; Zhou, J.; Wang, B.; Chen, Y.; Kong, Y.; Xie, X.; Wang, X.; Fei, R.; Wei, L.; et al. Peritumoural neutrophils negatively regulate adaptive immunity via the PD-L1/PD-1 signalling pathway in hepatocellular carcinoma. J. Exp. Clin. Cancer Res. 2015, 34, 141. [Google Scholar] [CrossRef] [Green Version]
- Duraiswamy, J.; Freeman, G.J.; Coukos, G. Therapeutic PD-1 pathway blockade augments with other modalities of immunotherapy T-cell function to prevent immune decline in ovarian cancer. Cancer Res. 2013, 73, 6900–6912. [Google Scholar] [CrossRef]
- Chen, J.; Ji, T.; Zhao, J.; Li, G.; Zhang, J.; Jin, R.; Liu, J.; Liu, X.; Liang, X.; Huang, D.; et al. Sorafenib-resistant hepatocellular carcinoma stratified by phosphorylated ERK activates PD-1 immune checkpoint. Oncotarget 2016, 7, 41274–41284. [Google Scholar] [CrossRef] [Green Version]
- Kalathil, S.G.; Lugade, A.A.; Miller, A.; Iyer, R.; Thanavala, Y. PD-1+ and Foxp3+ T cell reduction correlates with survival of HCC patients after sorafenib therapy. JCI Insight 2016, 1, 86182. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.; Bannert, C.; Gruber, S.; Klunker, S.; Spittler, A.; Akdis, C.A.; Szepfalusi, Z.; Eiwegger, T. Cord blood derived CD4+ CD25(high) T cells become functional regulatory T cells upon antigen encounter. PLoS ONE 2012, 7, e29355. [Google Scholar] [CrossRef]
- Hogan, P.G. Calcium-NFAT transcriptional signalling in T cell activation and T cell exhaustion. Cell Calcium 2017, 63, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Baine, I.; Abe, B.T.; Macian, F. Regulation of T-cell tolerance by calcium/NFAT signaling. Immunol. Rev. 2009, 231, 225–240. [Google Scholar] [CrossRef]
- Santegoets, S.J.; Dijkgraaf, E.M.; Battaglia, A.; Beckhove, P.; Britten, C.M.; Gallimore, A.; Godkin, A.; Gouttefangeas, C.; de Gruijl, T.D.; Koenen, H.J.; et al. Monitoring regulatory T cells in clinical samples: Consensus on an essential marker set and gating strategy for regulatory T cell analysis by flow cytometry. Cancer Immunol. Immunother. 2015, 64, 1271–1286. [Google Scholar] [CrossRef]
- Gouillat, C.; Manganas, D.; Zoulim, F.; Vitrey, D.; Saguier, G.; Guillaud, M.; Ain, J.F.; Duque-Campos, R.; Jamard, C.; Praves, M.; et al. Woodchuck hepatitis virus-induced carcinoma as a relevant natural model for therapy of human hepatoma. J. Hepatol. 1997, 26, 1324–1330. [Google Scholar] [CrossRef]
- Clark, J.W.; Eder, J.P.; Ryan, D.; Lathia, C.; Lenz, H.J. Safety and pharmacokinetics of the dual action Raf kinase and vascular endothelial growth factor receptor inhibitor, BAY 43-9006, in patients with advanced, refractory solid tumors. Clin. Cancer Res. 2005, 11, 5472–5480. [Google Scholar] [CrossRef]
- Awada, A.; Hendlisz, A.; Gil, T.; Bartholomeus, S.; Mano, M.; de Valeriola, D.; Strumberg, D.; Brendel, E.; Haase, C.G.; Schwartz, B.; et al. Phase I safety and pharmacokinetics of BAY 43-9006 administered for 21 days on/7 days off in patients with advanced, refractory solid tumours. Br. J. Cancer 2005, 92, 1855–1861. [Google Scholar] [CrossRef] [Green Version]
- Therasse, P.; Arbuck, S.G.; Eisenhauer, E.A.; Wanders, J.; Kaplan, R.S.; Rubinstein, L.; Verweij, J.; Van Glabbeke, M.; van Oosterom, A.T.; Christian, M.C.; et al. New guidelines to evaluate the response to treatment in solid tumors. European Organization for Research and Treatment of Cancer, National Cancer Institute of the United States, National Cancer Institute of Canada. J. Natl. Cancer Inst. 2000, 92, 205–216. [Google Scholar] [CrossRef]
- Triolo, M.; Della Corte, C.; Colombo, M. Impact of HBV therapy on the incidence of hepatocellular carcinoma. Liver Int. 2014, 34 (Suppl. 1), 139–145. [Google Scholar] [CrossRef]
- Groebe, K.; Mueller-Klieser, W. On the relation between size of necrosis and diameter of tumor spheroids. Int. J. Radiat. Oncol. Biol. Phys. 1996, 34, 395–401. [Google Scholar] [CrossRef]
- Milross, C.G.; Tucker, S.L.; Mason, K.A.; Hunter, N.R.; Peters, L.J.; Milas, L. The effect of tumor size on necrosis and polarographically measured pO2. Acta Oncol. 1997, 36, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Powers, W.E.; Tolmach, L.J. A multicomponent x-ray survival curve for mouse lymphosarcoma cells irradiated in vivo. Nature 1963, 197, 710–711. [Google Scholar] [CrossRef]
- Moulder, J.E.; Rockwell, S. Tumor hypoxia: Its impact on cancer therapy. Cancer Metastasis Rev. 1987, 5, 313–341. [Google Scholar] [CrossRef] [PubMed]
- Maguire, O.; Tornatore, K.M.; O’Loughlin, K.L.; Venuto, R.C.; Minderman, H. Nuclear translocation of nuclear factor of activated T cells (NFAT) as a quantitative pharmacodynamic parameter for tacrolimus. Cytometry A 2013, 83, 1096–1104. [Google Scholar]
- Maguire, O.; Chen, G.L.; Hahn, T.E.; Brix, L.; McCarthy, P.L.; Wallace, P.K.; Minderman, H. Quantifying MHC dextramer-induced NFAT activation in antigen-specific T cells as a functional response parameter. Methods 2017, 112, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Strumberg, D.; Clark, J.W.; Awada, A.; Moore, M.J.; Richly, H.; Hendlisz, A.; Hirte, H.W.; Eder, J.P.; Lenz, H.J.; Schwartz, B. Safety, pharmacokinetics, and preliminary antitumor activity of sorafenib: A review of four phase I trials in patients with advanced refractory solid tumors. Oncologist 2007, 12, 426–437. [Google Scholar] [CrossRef]
- Yamamoto, Y.; De Velasco, M.A.; Kura, Y.; Nozawa, M.; Hatanaka, Y.; Oki, T.; Ozeki, T.; Shimizu, N.; Minami, T.; Yoshimura, K.; et al. Evaluation of in vivo responses of sorafenib therapy in a preclinical mouse model of PTEN-deficient of prostate cancer. J. Transl. Med. 2015, 13, 150. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PK Parameters | AUCAll (h * ng/mL) | AUC1–7 h (h * ng/mL) | Cmax (ng/mL) | Cmax/D (ng/mL)/(mg/kg) | Tmax (h) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| n, Geo Mean (%CV) | n, Geo Mean (%CV) | n, Geo Mean (%CV) | n, Geo Mean (%CV) | n, Geo Mean (%CV) | ||||||
| Dose Level (mg/kg) | Day 1 | Day 19 | Day 1 | Day 19 | Day 1 | Day 19 | Day 1 | Day 19 | Day 1 | Day 19 |
| 2.5 | 2, 2600 (4.76) | 3, 30,300 (15.8) | 2, 588 (0.648) | 3, 26,800 (15.8) | 2, 136 (3.41) | 3, 4970 (19.8) | 2, 54.4 (3.41) | 3, 1990 (19.8) | 2, 5.92 (23.6) | 3, 4.22 (26.6) |
| 5.0 | 3, 21,000 (70.9) | 3, 43,8000 (67.0) | 3, 5960 (72.6) | 3, 39,100 (65.5) | 3, 1170 (73.7) | 3, 7380 (64.5) | 3, 433 (75.8) | 3, 2000 (54.5) | 3, 3.98 (53.3) | 3, 3.98 (53.3) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iyer, R.V.; Maguire, O.; Kim, M.; Curtin, L.I.; Sexton, S.; Fisher, D.T.; Schihl, S.A.; Fetterly, G.; Menne, S.; Minderman, H. Dose-Dependent Sorafenib-Induced Immunosuppression Is Associated with Aberrant NFAT Activation and Expression of PD-1 in T Cells. Cancers 2019, 11, 681. https://doi.org/10.3390/cancers11050681
Iyer RV, Maguire O, Kim M, Curtin LI, Sexton S, Fisher DT, Schihl SA, Fetterly G, Menne S, Minderman H. Dose-Dependent Sorafenib-Induced Immunosuppression Is Associated with Aberrant NFAT Activation and Expression of PD-1 in T Cells. Cancers. 2019; 11(5):681. https://doi.org/10.3390/cancers11050681
Chicago/Turabian StyleIyer, Renuka V., Orla Maguire, Minhyung Kim, Leslie I. Curtin, Sandra Sexton, Daniel T. Fisher, Sarah A. Schihl, Gerald Fetterly, Stephan Menne, and Hans Minderman. 2019. "Dose-Dependent Sorafenib-Induced Immunosuppression Is Associated with Aberrant NFAT Activation and Expression of PD-1 in T Cells" Cancers 11, no. 5: 681. https://doi.org/10.3390/cancers11050681