Contribution of mTOR and PTEN to Radioresistance in Sporadic and NF2-Associated Vestibular Schwannomas: A Microarray and Pathway Analysis

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Clinical Data
2.2. The mTOR and PTEN Signaling Seem to Be of Potential Interest in Recurrent Irradiated NF2-Associated Vestibular Schwannomas
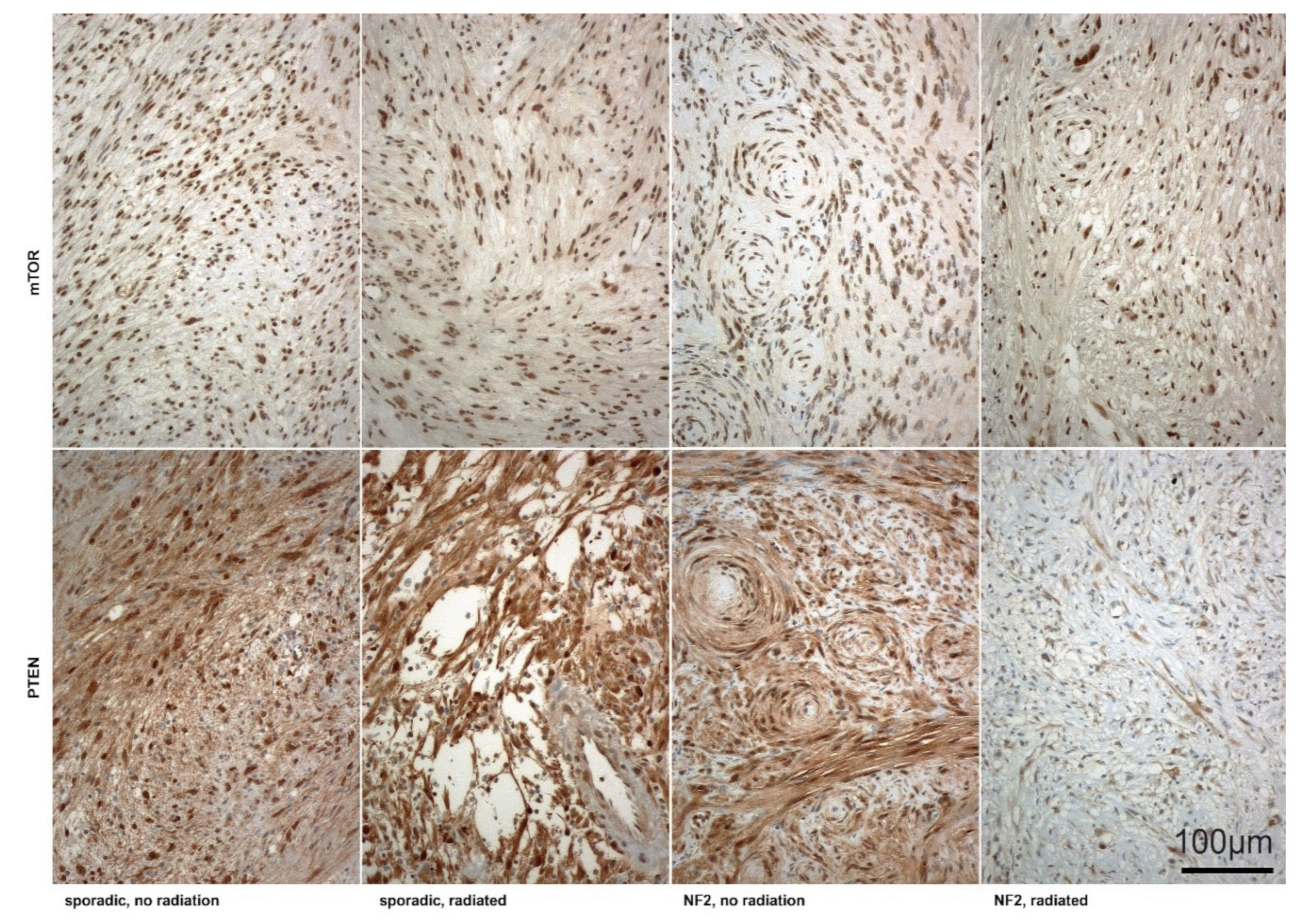
2.3. Irradiated NF2-Associated VS Revealed Significant Lower PTEN Expression Intensity Compared to Non-Irradiated or Sporadic Tumors
3. Discussion
3.1. Low Proliferation Signaling Reveals Radioresistance
3.2. Suppression of Angiogenesis Mediators as an Effect of Irradiation
3.3. Cell Cycle Mediation and Regulation of Apoptosis
3.4. PTEN and mTOR Signaling—Core Pathway in Merlin-Inactive and Progressive NF2-Associated Tumors after Irradiation
3.5. Limitations of the Current Study
4. Material and Methods
4.1. Patients and Tumors
4.2. RNA Extraction and RNA Integrity Number (RIN)
4.3. cDNA Microarray Analysis
4.4. Statistical Array Analysis
4.5. Pathway Analysis
4.6. Immunohistochemistry (IHC)
4.7. Statistical Clinical Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Welling, D.B. Clinical manifestations of mutations in the neurofibromatosis type 2 gene in vestibular schwannomas (acoustic neuromas). Laryngoscope 1998, 108, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Kshettry, V.R.; Hsieh, J.K.; Ostrom, Q.T.; Kruchko, C.; Barnholtz-Sloan, J.S. Incidence of vestibular schwannomas in the United States. J. Neurooncol. 2015, 124, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Neff, B.A.; Welling, D.B.; Akhmametyeva, E.; Chang, L.S. The molecular biology of vestibular schwannomas: Dissecting the pathogenic process at the molecular level. Otol. Neurotol. 2006, 27, 197–208. [Google Scholar] [CrossRef]
- Evans, D.G. Neurofibromatosis type 2 (NF2): A clinical and molecular review. Orphanet J. Rare Dis. 2009, 4, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gugel, I.; Mautner, V.F.; Kluwe, L.; Tatagiba, M.S.; Schuhmann, M.U. Cerebrovascular Insult as Presenting Symptom of Neurofibromatosis Type 2 in Children, Adolescents, and Young Adults. Front. Neurol. 2018, 9, 733. [Google Scholar] [CrossRef] [Green Version]
- Samii, M.; Gerganov, V.; Samii, A. Microsurgery management of vestibular schwannomas in neurofibromatosis type 2: Indications and results. Prog. Neurol. Surg. 2008, 21, 169–175. [Google Scholar] [CrossRef]
- Gugel, I.; Grimm, F.; Teuber, C.; Kluwe, L.; Mautner, V.-F.; Tatagiba, M.; Schuhmann, M.U. Management of NF2-associated vestibular schwannomas in children and young adults: Influence of surgery and clinical factors on tumor volume and growth rate. J. Neurosurg. Pediatr. 2019, 24, 584–592. [Google Scholar] [CrossRef]
- Sakamoto, G.T.; Blevins, N.; Gibbs, I.C. Cyberknife radiotherapy for vestibular schwannoma. Otolaryngol. Clin. N. Am. 2009, 42, 665–675. [Google Scholar] [CrossRef]
- Sweeney, P.; Yajnik, S.; Hartsell, W.; Bovis, G.; Venkatesan, J. Stereotactic radiotherapy for vestibular schwannoma. Otolaryngol. Clin. N. Am. 2009, 42, 655–663. [Google Scholar] [CrossRef]
- Hochart, A.; Gaillard, V.; Baroncini, M.; Andre, N.; Vannier, J.P.; Vinchon, M.; Dubrulle, F.; Lejeune, J.P.; Vincent, C.; Neve, V.; et al. Bevacizumab decreases vestibular schwannomas growth rate in children and teenagers with neurofibromatosis type 2. J. Neurooncol. 2015, 124, 229–236. [Google Scholar] [CrossRef]
- Plotkin, S.R.; Merker, V.L.; Halpin, C.; Jennings, D.; McKenna, M.J.; Harris, G.J.; Barker, F.G., 2nd. Bevacizumab for progressive vestibular schwannoma in neurofibromatosis type 2: A retrospective review of 31 patients. Otol. Neurotol. 2012, 33, 1046–1052. [Google Scholar] [CrossRef] [PubMed]
- Gugel, I.; Grimm, F.; Liebsch, M.; Zipfel, J.; Teuber, C.; Kluwe, L.; Mautner, V.F.; Tatagiba, M.; Schuhmann, M.U. Impact of Surgery on Long-Term Results of Hearing in Neurofibromatosis Type-2 Associated Vestibular Schwannomas. Cancers 2019, 11, 1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meijer, O.W.; Vandertop, W.P.; Lagerwaard, F.J.; Slotman, B.J. Linear accelerator-based stereotactic radiosurgery for bilateral vestibular schwannomas in patients with neurofibromatosis type 2. Neurosurgery 2008, 62, A37–A42. [Google Scholar] [CrossRef] [PubMed]
- Rowe, J.; Radatz, M.; Kemeny, A. Radiosurgery for type II neurofibromatosis. Prog. Neurol. Surg. 2008, 21, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.G.; Birch, J.M.; Ramsden, R.T.; Sharif, S.; Baser, M.E. Malignant transformation and new primary tumours after therapeutic radiation for benign disease: Substantial risks in certain tumour prone syndromes. J. Med. Genet. 2006, 43, 289–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lustgarten, L. The impact of stereotactic radiosurgery in the management of neurofibromatosis type 2-related vestibular schwannomas. Surg. Neurol. Int. 2013, 4, S151–S155. [Google Scholar] [CrossRef]
- Sharma, M.S.; Singh, R.; Kale, S.S.; Agrawal, D.; Sharma, B.S.; Mahapatra, A.K. Tumor control and hearing preservation after Gamma Knife radiosurgery for vestibular schwannomas in neurofibromatosis type 2. J. Neurooncol. 2010, 98, 265–270. [Google Scholar] [CrossRef]
- Mathieu, D.; Kondziolka, D.; Flickinger, J.C.; Niranjan, A.; Williamson, R.; Martin, J.J.; Lunsford, L.D. Stereotactic radiosurgery for vestibular schwannomas in patients with neurofibromatosis type 2: An analysis of tumor control, complications, and hearing preservation rates. Neurosurgery 2007, 60, 460–468. [Google Scholar] [CrossRef]
- Jacob, A.; Lee, T.X.; Neff, B.A.; Miller, S.; Welling, B.; Chang, L.S. Phosphatidylinositol 3-kinase/AKT pathway activation in human vestibular schwannoma. Otol. Neurotol. 2008, 29, 58–68. [Google Scholar] [CrossRef]
- Asthagiri, A.R.; Parry, D.M.; Butman, J.A.; Kim, H.J.; Tsilou, E.T.; Zhuang, Z.; Lonser, R.R. Neurofibromatosis type 2. Lancet 2009, 373, 1974–1986. [Google Scholar] [CrossRef] [Green Version]
- Irving, R.M.; Moffat, D.A.; Hardy, D.G.; Barton, D.E.; Xuereb, J.H.; Maher, E.R. Somatic NF2 gene mutations in familial and non-familial vestibular schwannoma. Hum. Mol. Genet. 1994, 3, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Kissil, J.L.; Johnson, K.C.; Eckman, M.S.; Jacks, T. Merlin phosphorylation by p21-activated kinase 2 and effects of phosphorylation on merlin localization. J. Biol. Chem. 2002, 277, 10394–10399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irving, R.M.; Moffat, D.A.; Hardy, D.G.; Barton, D.E.; Xuereb, J.H.; Holland, F.J.; Maher, E.R. A molecular, clinical, and immunohistochemical study of vestibular schwannoma. Otolaryngol. Head Neck Surg. 1997, 116, 426–430. [Google Scholar] [CrossRef]
- Ahronowitz, I.; Xin, W.; Kiely, R.; Sims, K.; MacCollin, M.; Nunes, F.P. Mutational spectrum of the NF2 gene: A meta-analysis of 12 years of research and diagnostic laboratory findings. Hum. Mutat. 2007, 28, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sherman, L.S.; Gutmann, D.H. Merlin: Hanging tumor suppression on the Rac. Trends Cell Biol. 2001, 11, 442–444. [Google Scholar] [CrossRef]
- Welling, D.B.; Lasak, J.M.; Akhmametyeva, E.; Ghaheri, B.; Chang, L.S. cDNA microarray analysis of vestibular schwannomas. Otol. Neurotol. 2002, 23, 736–748. [Google Scholar] [CrossRef] [PubMed]
- Blair, K.J.; Kiang, A.; Wang-Rodriguez, J.; Yu, M.A.; Doherty, J.K.; Ongkeko, W.M. EGF and bFGF promote invasion that is modulated by PI3/Akt kinase and Erk in vestibular schwannoma. Otol. Neurotol. 2011, 32, 308–314. [Google Scholar] [CrossRef]
- James, M.F.; Han, S.; Polizzano, C.; Plotkin, S.R.; Manning, B.D.; Stemmer-Rachamimov, A.O.; Gusella, J.F.; Ramesh, V. NF2/merlin is a novel negative regulator of mTOR complex 1, and activation of mTORC1 is associated with meningioma and schwannoma growth. Mol. Cell Biol. 2009, 29, 4250–4261. [Google Scholar] [CrossRef] [Green Version]
- James, M.F.; Stivison, E.; Beauchamp, R.; Han, S.; Li, H.; Wallace, M.R.; Gusella, J.F.; Stemmer-Rachamimov, A.O.; Ramesh, V. Regulation of mTOR complex 2 signaling in neurofibromatosis 2-deficient target cell types. Mol. Cancer Res. 2012, 10, 649–659. [Google Scholar] [CrossRef] [Green Version]
- Rong, R.; Tang, X.; Gutmann, D.H.; Ye, K. Neurofibromatosis 2 (NF2) tumor suppressor merlin inhibits phosphatidylinositol 3-kinase through binding to PIKE-L. Proc. Natl. Acad. Sci. USA 2004, 101, 18200–18205. [Google Scholar] [CrossRef] [Green Version]
- Ammoun, S.; Flaiz, C.; Ristic, N.; Schuldt, J.; Hanemann, C.O. Dissecting and targeting the growth factor-dependent and growth factor-independent extracellular signal-regulated kinase pathway in human schwannoma. Cancer Res. 2008, 68, 5236–5245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sainz, J.; Huynh, D.P.; Figueroa, K.; Ragge, N.K.; Baser, M.E.; Pulst, S.M. Mutations of the neurofibromatosis type 2 gene and lack of the gene product in vestibular schwannomas. Hum. Mol. Genet. 1994, 3, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Aarhus, M.; Bruland, O.; Saetran, H.A.; Mork, S.J.; Lund-Johansen, M.; Knappskog, P.M. Global Gene Expression Profiling and Tissue Microarray Reveal Novel Candidate Genes and Down-Regulation of the Tumor Suppressor Gene CAV1 in Sporadic Vestibular Schwannomas. Neurosurgery 2010, 67, 998–1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caye-Thomasen, P.; Borup, R.; Stangerup, S.E.; Thomsen, J.; Nielsen, F.C. Deregulated Genes in Sporadic Vestibular Schwannomas. Otol. Neurotol. 2009. [Google Scholar] [CrossRef] [Green Version]
- Lasak, J.M.; Welling, D.B.; Akhmametyeva, E.M.; Salloum, M.; Chang, L.S. Retinoblastoma-cyclin-dependent kinase pathway deregulation in vestibular schwannomas. Laryngoscope 2002, 112, 1555–1561. [Google Scholar] [CrossRef] [PubMed]
- Neff, B.A.; Oberstien, E.; Lorenz, M.; Chaudhury, A.R.; Welling, D.B.; Chang, L.S. Cyclin D(1) and D(3) expression in vestibular schwannomas. Laryngoscope 2006, 116, 423–426. [Google Scholar] [CrossRef]
- Lassaletta, L.; Patron, M.; Del Rio, L.; Alfonso, C.; Roda, J.M.; Rey, J.A.; Gavilan, J. Cyclin D1 expression and histopathologic features in vestibular schwannomas. Otol. Neurotol. 2007, 28, 939–941. [Google Scholar] [CrossRef]
- Hanemann, C.O.; Bartelt-Kirbach, B.; Diebold, R.; Kampchen, K.; Langmesser, S.; Utermark, T. Differential gene expression between human schwannoma and control Schwann cells. Neuropathol. Appl. Neurobiol. 2006, 32, 605–614. [Google Scholar] [CrossRef] [Green Version]
- Agnihotri, S.; Gugel, I.; Remke, M.; Bornemann, A.; Pantazis, G.; Mack, S.C.; Shih, D.; Singh, S.K.; Sabha, N.; Taylor, M.D.; et al. Gene-expression profiling elucidates molecular signaling networks that can be therapeutically targeted in vestibular schwannoma. J. Neurosurg. 2014, 121, 1434–1445. [Google Scholar] [CrossRef] [Green Version]
- Samii, M.; Matthies, C. Management of 1000 vestibular schwannomas (acoustic neuromas): Hearing function in 1000 tumor resections. Neurosurgery 1997, 40, 248–260. [Google Scholar] [CrossRef]
- Gardner, G.; Robertson, J.H. Hearing preservation in unilateral acoustic neuroma surgery. Ann. Otol. Rhinol. Laryngol. 1988, 97, 55–66. [Google Scholar] [CrossRef] [PubMed]
- House, J.W.; Brackmann, D.E. Facial nerve grading system. Otolaryngol. Head Neck Surg. 1985, 93, 146–147. [Google Scholar] [CrossRef] [PubMed]
- Kelley, K.; Knisely, J.; Symons, M.; Ruggieri, R. Radioresistance of Brain Tumors. Cancers 2016, 8, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeung, A.H.; Sughrue, M.E.; Kane, A.J.; Tihan, T.; Cheung, S.W.; Parsa, A.T. Radiobiology of vestibular schwannomas: Mechanisms of radioresistance and potential targets for therapeutic sensitization. Neurosurg. Focus 2009, 27, E2. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Wang, Z.; Sun, L.; Li, X.; Huang, Q.; Yang, T.; Wu, H. Mutation spectrum and differential gene expression in cystic and solid vestibular schwannoma. Genet. Med. 2014, 16, 264–270. [Google Scholar] [CrossRef] [Green Version]
- Stankovic, K.M.; Mrugala, M.M.; Martuza, R.L.; Silver, M.; Betensky, R.A.; Nadol, J.B., Jr.; Stemmer-Rachamimov, A.O. Genetic determinants of hearing loss associated with vestibular schwannomas. Otol. Neurotol. 2009, 30, 661–667. [Google Scholar] [CrossRef] [Green Version]
- Lassaletta, L.; Martinez-Glez, V.; Torres-Martin, M.; Rey, J.A.; Gavilan, J. cDNA microarray expression profile in vestibular schwannoma: Correlation with clinical and radiological features. Cancer Genet. Cytogenet. 2009, 194, 125–127. [Google Scholar] [CrossRef]
- Torres-Martin, M.; Lassaletta, L.; San-Roman-Montero, J.; De Campos, J.M.; Isla, A.; Gavilan, J.; Melendez, B.; Pinto, G.R.; Burbano, R.R.; Castresana, J.S.; et al. Microarray analysis of gene expression in vestibular schwannomas reveals SPP1/MET signaling pathway and androgen receptor deregulation. Int. J. Oncol. 2013, 42, 848–862. [Google Scholar] [CrossRef] [Green Version]
- Frohnert, P.W.; Stonecypher, M.S.; Carroll, S.L. Constitutive activation of the neuregulin-1/ErbB receptor signaling pathway is essential for the proliferation of a neoplastic Schwann cell line. Glia 2003, 43, 104–118. [Google Scholar] [CrossRef]
- Stonecypher, M.S.; Chaudhury, A.R.; Byer, S.J.; Carroll, S.L. Neuregulin growth factors and their ErbB receptors form a potential signaling network for schwannoma tumorigenesis. J. Neuropathol. Exp. Neurol. 2006, 65, 162–175. [Google Scholar] [CrossRef] [Green Version]
- Hansen, M.R.; Roehm, P.C.; Chatterjee, P.; Green, S.H. Constitutive neuregulin-1/ErbB signaling contributes to human vestibular schwannoma proliferation. Glia 2006, 53, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.R.; Linthicum, F.H., Jr. Expression of neuregulin and activation of erbB receptors in vestibular schwannomas: Possible autocrine loop stimulation. Otol. Neurotol. 2004, 25, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Newbern, J.; Birchmeier, C. Nrg1/ErbB signaling networks in Schwann cell development and myelination. Semin. Cell Dev. Biol. 2010, 21, 922–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, M.R.; Clark, J.J.; Gantz, B.J.; Goswami, P.C. Effects of ErbB2 signaling on the response of vestibular schwannoma cells to gamma-irradiation. Laryngoscope 2008, 118, 1023–1030. [Google Scholar] [CrossRef]
- Lee, F.; Linthicum, F., Jr.; Hung, G. Proliferation potential in recurrent acoustic schwannoma following gamma knife radiosurgery versus microsurgery. Laryngoscope 2002, 112, 948–950. [Google Scholar] [CrossRef]
- Plotkin, S.R.; Stemmer-Rachamimov, A.O.; Barker, F.G., 2nd; Halpin, C.; Padera, T.P.; Tyrrell, A.; Sorensen, A.G.; Jain, R.K.; di Tomaso, E. Hearing improvement after bevacizumab in patients with neurofibromatosis type 2. N. Engl. J. Med. 2009, 361, 358–367. [Google Scholar] [CrossRef] [Green Version]
- Dalgorf, D.M.; Rowsell, C.; Bilbao, J.M.; Chen, J.M. Immunohistochemical investigation of hormone receptors and vascular endothelial growth factor concentration in vestibular schwannoma. Skull Base 2008, 18, 377–384. [Google Scholar] [CrossRef] [Green Version]
- Koutsimpelas, D.; Stripf, T.; Heinrich, U.R.; Mann, W.J.; Brieger, J. Expression of vascular endothelial growth factor and basic fibroblast growth factor in sporadic vestibular schwannomas correlates to growth characteristics. Otol. Neurotol. 2007, 28, 1094–1099. [Google Scholar] [CrossRef] [Green Version]
- Mautner, V.F.; Nguyen, R.; Kutta, H.; Fuensterer, C.; Bokemeyer, C.; Hagel, C.; Friedrich, R.E.; Panse, J. Bevacizumab induces regression of vestibular schwannomas in patients with neurofibromatosis type 2. Neuro-Oncol. 2010, 12, 14–18. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Zhao, Y.; Stemmer-Rachamimov, A.O.; Liu, H.; Huang, P.; Chin, S.; Selig, M.K.; Plotkin, S.R.; Jain, R.K.; Xu, L. Anti-VEGF treatment improves neurological function and augments radiation response in NF2 schwannoma model. Proc. Natl. Acad. Sci. USA 2015, 112, 14676–14681. [Google Scholar] [CrossRef] [Green Version]
- D’Avenia, P.; Porrello, A.; Berardo, M.; Angelo, M.D.; Soddu, S.; Arcangeli, G.; Sacchi, A.; D’Orazi, G. Tp53-gene transfer induces hypersensitivity to low doses of X-rays in glioblastoma cells: A strategy to convert a radio-resistant phenotype into a radiosensitive one. Cancer Lett. 2006, 231, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Irving, R.M.; Moffat, D.A.; Hardy, D.G.; Barton, D.E.; Xuereb, J.H.; Maher, E.R. Molecular genetic analysis of the mechanism of tumorigenesis in acoustic neuroma. Arch. Otolaryngol. Head Neck Surg. 1993, 119, 1222–1228. [Google Scholar] [CrossRef] [PubMed]
- Monoh, K.; Ishikawa, K.; Yasui, N.; Mineura, K.; Andoh, H.; Togawa, K. p53 tumor suppressor gene in acoustic neuromas. Acta Otolaryngol. Suppl. 1998, 537, 11–15. [Google Scholar] [PubMed]
- Dent, P.; Yacoub, A.; Fisher, P.B.; Hagan, M.P.; Grant, S. MAPK pathways in radiation responses. Oncogene 2003, 22, 5885–5896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, H.; Sperka, T.; Manent, J.; Giovannini, M.; Ponta, H.; Herrlich, P. Merlin/neurofibromatosis type 2 suppresses growth by inhibiting the activation of Ras and Rac. Cancer Res. 2007, 67, 520–527. [Google Scholar] [CrossRef] [Green Version]
- Tikoo, A.; Varga, M.; Ramesh, V.; Gusella, J.; Maruta, H. An anti-Ras function of neurofibromatosis type 2 gene product (NF2/Merlin). J. Biol. Chem. 1994, 269, 23387–23390. [Google Scholar]
- Song, M.S.; Salmena, L.; Pandolfi, P.P. The functions and regulation of the PTEN tumour suppressor. Nat. Rev. Mol. Cell Biol. 2012, 13, 283–296. [Google Scholar] [CrossRef]
- Blakeley, J. Development of drug treatments for neurofibromatosis type 2-associated vestibular schwannoma. Curr. Opin. Otolaryngol. Head Neck Surg. 2012, 20, 372–379. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Jaquez, H.A.; Keller, J.A.; Foster, K.G.; Ekim, B.; Soliman, G.A.; Feener, E.P.; Ballif, B.A.; Fingar, D.C. Site-specific mTOR phosphorylation promotes mTORC1-mediated signaling and cell growth. Mol. Cell Biol. 2009, 29, 4308–4324. [Google Scholar] [CrossRef] [Green Version]
- Giovannini, M.; Bonne, N.X.; Vitte, J.; Chareyre, F.; Tanaka, K.; Adams, R.; Fisher, L.M.; Valeyrie-Allanore, L.; Wolkenstein, P.; Goutagny, S.; et al. mTORC1 inhibition delays growth of neurofibromatosis type 2 schwannoma. Neuro-Oncol. 2014, 16, 493–504. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Lago, M.A.; Okada, T.; Murillo, M.M.; Socci, N.; Giancotti, F.G. Loss of the tumor suppressor gene NF2, encoding merlin, constitutively activates integrin-dependent mTORC1 signaling. Mol. Cell Biol. 2009, 29, 4235–4249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, H.A.; Wood, J.M.; McSheehy, P.M.; Allegrini, P.R.; Boulay, A.; Brueggen, J.; Littlewood-Evans, A.; Maira, S.M.; Martiny-Baron, G.; Schnell, C.R.; et al. mTOR inhibitor RAD001 (everolimus) has antiangiogenic/vascular properties distinct from a VEGFR tyrosine kinase inhibitor. Clin. Cancer Res. 2009, 15, 1612–1622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karajannis, M.A.; Legault, G.; Hagiwara, M.; Giancotti, F.G.; Filatov, A.; Derman, A.; Hochman, T.; Goldberg, J.D.; Vega, E.; Wisoff, J.H.; et al. Phase II study of everolimus in children and adults with neurofibromatosis type 2 and progressive vestibular schwannomas. Neuro-Oncol. 2014, 16, 292–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantley, L.C.; Neel, B.G. New insights into tumor suppression: PTEN suppresses tumor formation by restraining the phosphoinositide 3-kinase/AKT pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 4240–4245. [Google Scholar] [CrossRef] [Green Version]
- Wick, W.; Furnari, F.B.; Naumann, U.; Cavenee, W.K.; Weller, M. PTEN gene transfer in human malignant glioma: Sensitization to irradiation and CD95L-induced apoptosis. Oncogene 1999, 18, 3936–3943. [Google Scholar] [CrossRef] [Green Version]
- Inaba, N.; Kimura, M.; Fujioka, K.; Ikeda, K.; Somura, H.; Akiyoshi, K.; Inoue, Y.; Nomura, M.; Saito, Y.; Saito, H.; et al. The effect of PTEN on proliferation and drug-, and radiosensitivity in malignant glioma cells. Anticancer Res. 2011, 31, 1653–1658. [Google Scholar]
- Keng, V.W.; Rahrmann, E.P.; Watson, A.L.; Tschida, B.R.; Moertel, C.L.; Jessen, W.J.; Rizvi, T.A.; Collins, M.H.; Ratner, N.; Largaespada, D.A. PTEN and NF1 inactivation in Schwann cells produces a severe phenotype in the peripheral nervous system that promotes the development and malignant progression of peripheral nerve sheath tumors. Cancer Res. 2012, 72, 3405–3413. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, K.; Oda, Y.; Saito, T.; Takahira, T.; Yamamoto, H.; Tamiya, S.; Iwamoto, Y.; Tsuneyoshi, M. Genetic and epigenetic alterations of the PTEN gene in soft tissue sarcomas. Hum. Pathol. 2005, 36, 357–363. [Google Scholar] [CrossRef]
- Gregorian, C.; Nakashima, J.; Dry, S.M.; Nghiemphu, P.L.; Smith, K.B.; Ao, Y.; Dang, J.; Lawson, G.; Mellinghoff, I.K.; Mischel, P.S.; et al. PTEN dosage is essential for neurofibroma development and malignant transformation. Proc. Natl. Acad. Sci. USA 2009, 106, 19479–19484. [Google Scholar] [CrossRef] [Green Version]
- Pattje, W.J.; Schuuring, E.; Mastik, M.F.; Slagter-Menkema, L.; Schrijvers, M.L.; Alessi, S.; van der Laan, B.F.; Roodenburg, J.L.; Langendijk, J.A.; van der Wal, J.E. The phosphatase and tensin homologue deleted on chromosome 10 mediates radiosensitivity in head and neck cancer. Br. J. Cancer 2010, 102, 1778–1785. [Google Scholar] [CrossRef]
- Puc, J.; Parsons, R. PTEN loss inhibits CHK1 to cause double stranded-DNA breaks in cells. Cell Cycle 2005, 4, 927–929. [Google Scholar] [CrossRef] [PubMed]
- Kao, G.D.; Jiang, Z.; Fernandes, A.M.; Gupta, A.K.; Maity, A. Inhibition of phosphatidylinositol-3-OH kinase/Akt signaling impairs DNA repair in glioblastoma cells following ionizing radiation. J. Biol. Chem. 2007, 282, 21206–21212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abo-Dalo, B.; Kutsche, K.; Mautner, V.; Kluwe, L. Large intragenic deletions of the NF2 gene: Breakpoints and associated phenotypes. Genes Chromosomes Cancer 2010, 49, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Gentleman, R.C.; Carey, V.J.; Bates, D.M.; Bolstad, B.; Dettling, M.; Dudoit, S.; Ellis, B.; Gautier, L.; Ge, Y.; Gentry, J.; et al. Bioconductor: Open software development for computational biology and bioinformatics. Genome Biol. 2004, 5, R80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautier, L.; Cope, L.; Bolstad, B.M.; Irizarry, R.A. Affy—Analysis of Affymetrix GeneChip data at the probe level. Bioinformatics 2004, 20, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Smyth, G.K. Linear models and empirical bayes methods for assessing differential expression in microarray experiments. Stat. Appl. Genet. Mol. Biol. 2004, 3, Article3. [Google Scholar] [CrossRef]
- Smyth, G.K. Bioinformatics and Computational Biology Solutions using R and Bioconductor. In Bioinformatics and Computational Biology Solutions Using R and Bioconductor; Gentlemen, R., Carey, V., Dudoit, S., Irizarry, R., Huber, W., Eds.; Springer: New York, NY, USA, 2005; pp. 397–420. [Google Scholar]
- Irizarry, R.A.; Bolstad, B.M.; Collin, F.; Cope, L.M.; Hobbs, B.; Speed, T.P. Summaries of Affymetrix GeneChip probe level data. Nucleic Acids Res. 2003, 31, e15. [Google Scholar] [CrossRef]
- Smyth, G.K.; Michaud, J.; Scott, H.S. Use of within-array replicate spots for assessing differential expression in microarray experiments. Bioinformatics 2005, 21, 2067–2075. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Statist. Soc. B 1995, 57, 289–300. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-Irradiated VS (49) | Irradiated VS (8) | |||
|---|---|---|---|---|
| SPO (36) | NF2 (13) | SPO (5) | NF2 (3) | |
| Sex | ||||
| male | 23 | 6 | 1 | 3 |
| female | 13 | 7 | 4 | 0 |
| Operation side | ||||
| left | 24 | 5 | 3 | 0 |
| right | 12 | 8 | 2 | 3 |
| Age at diagnosis in year | 44 ± 12 | 25 ± 8 | 49 ± 10 | 32 ± 12 |
| (mean ± std, range) | 25–76 | 11–40 | 33–62 | 24–49 |
| Age at surgery in year | 45 ± 12 | 25 ± 7 | 51 ± 12 | 33 ± 11 |
| (mean ± std, range) | 25–76 | 11–40 | 33–67 | 25–49 |
| Age at radiation treatment | 49 ± 11 | 29 ± 12 | ||
| (mean ± std, range) | 31–64 | 20–47 | ||
| Tumor size (Hannover classification system [40]) pre-op | ||||
| T1 | 0 | 1 | 0 | 0 |
| T2 | 0 | 1 | 0 | 0 |
| T3 | 11 | 3 | 1 | 1 |
| T4 | 25 | 8 | 4 | 2 |
| G–R Class [41] pre-op | ||||
| 1 | 4 | 1 | 0 | 0 |
| 2 | 11 | 0 | 0 | 0 |
| 3 | 8 | 0 | 2 | 0 |
| 4 | 0 | 0 | 0 | 0 |
| 5 | 4 | 9 | 3 | 2 |
| Not available | 9 | 3 | 0 | 1 |
| H–B Class [42] pre-op | ||||
| I | 28 | 8 | 2 | 0 |
| II | 8 | 1 | 1 | 0 |
| III | 0 | 1 | 1 | 1 |
| IV | 0 | 1 | 0 | 0 |
| V | 0 | 2 | 1 | 2 |
| Clinical symptoms pre-op | ||||
| Tinnitus | 17 | 2 | 1 | 0 |
| Event of sudden hearing loss | 12 | 2 | 0 | 0 2 |
| Vertigo | 19 | 3 | 4 | 0 |
| Gait disturbances | 13 | 8 | 0 | 1 |
| Trigeminal nerve dysfunction (hyp-/dysesthesia, reduced/loss of corneal reflex, neuralgia) | 16 | 6 | 3 | |
| Ingenuity Canonical Pathways | Regulation | ||
|---|---|---|---|
| SPO_RAD-SPO | NF2_RAD-NF2 | SPO-NF2 | |
| Notch signaling | 1 | nr | nr |
| TGF-β signaling | 1 | nr | nr |
| Wnt/β-catenin signaling | 0 | −1 | nr |
| PTEN signaling | 0 | −1 | nr |
| IGF-1 signaling | −1 | −1 | nr |
| VEGF signaling | −1 | −1 | nr |
| cMet/HGF signaling | −1 | −1 | nr |
| ErbB2–ErbB3 signaling | −1 | −1 | nr |
| Neuregulin signaling | −1 | −1 | nr |
| EGF/EGFR signaling | 1 | −1 | nr |
| PDGF signaling | 1 | −1 | nr |
| p53 signaling | 1 | −1 | nr |
| STAT3 Pathway | 1 | −1 | 1 |
| NF-κB signaling | 1 | −1 | −1 |
| FGF signaling | 1 | −1 | −1 |
| mTOR signaling | 0 | 1 | −1 |
| Methods | No of Tumor Samples | No and Type of Control Samples | Author, Year |
|---|---|---|---|
| cDNA Microarray, RT-PCR | 31 CYS-VS 6 SPO-VS | No | [45] |
| cDNA Microarray, Cell cultures, Western Blotting | 36 SPO-VS 13 NF2-VS | 7 vestibular nerves | [39] |
| cDNA Microarray, RT-PCR, MLPA analysis of NF2 LOH analysis | 28 SPO-VS 3 NF2-VS | 2 auricular nerves 2 cervical nerves 1 facial nerve 1 vestibular nerve 1 nerve from the VIII cranial pair 1 commercial normal human adult Schwann cell (HSC) RNA | [48] |
| cDNA Microarray, RT-PCR, IHC | 15 SPO-VS 7 CYS-VS 3 RAD-VS | 3 tibial nerves | [33] |
| cDNA Microarray, RT-PCR, IHC | 13 VS | No | [46] |
| cDNA Microarray | 16 SPO-VS | 3 vestibular nerves | [34] |
| cDNA Microarray, RT-PCR | 11 SPO-VS | 11 blood samples | [47] |
| cDNA Microarray, RT-PCR, IHC | 3 SPO-VS 1 CYS-VS 3 RAD-VS | 1 vestibular nerve | [26] |
| cDNA Microarray, IHC | 36 SPO-VS 13 NF2-VS 9 CYS-VS 9 RAD-VS | 7 vestibular nerves | Current study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gugel, I.; Ebner, F.H.; Grimm, F.; Czemmel, S.; Paulsen, F.; Hagel, C.; Tatagiba, M.; Nahnsen, S.; Tabatabai, G. Contribution of mTOR and PTEN to Radioresistance in Sporadic and NF2-Associated Vestibular Schwannomas: A Microarray and Pathway Analysis. Cancers 2020, 12, 177. https://doi.org/10.3390/cancers12010177
Gugel I, Ebner FH, Grimm F, Czemmel S, Paulsen F, Hagel C, Tatagiba M, Nahnsen S, Tabatabai G. Contribution of mTOR and PTEN to Radioresistance in Sporadic and NF2-Associated Vestibular Schwannomas: A Microarray and Pathway Analysis. Cancers. 2020; 12(1):177. https://doi.org/10.3390/cancers12010177
Chicago/Turabian StyleGugel, Isabel, Florian H. Ebner, Florian Grimm, Stefan Czemmel, Frank Paulsen, Christian Hagel, Marcos Tatagiba, Sven Nahnsen, and Ghazaleh Tabatabai. 2020. "Contribution of mTOR and PTEN to Radioresistance in Sporadic and NF2-Associated Vestibular Schwannomas: A Microarray and Pathway Analysis" Cancers 12, no. 1: 177. https://doi.org/10.3390/cancers12010177