The Role of Probiotics in Cancer Prevention


1
Institute of Fermentation Technology and Microbiology, Faculty of Biotechnology and Food Sciences, Lodz University of Technology, Wólczańska 171/173, 90-924 Lodz, Poland
2
Institute of Molecular and Industrial Biotechnology, Faculty of Biotechnology and Food Sciences, Lodz University of Technology, Stefanowskiego 4/10, 90-924 Lodz, Poland
*
Authors to whom correspondence should be addressed.
Cancers 2021, 13(1), 20; https://doi.org/10.3390/cancers13010020
Submission received: 14 November 2020
/
Revised: 17 December 2020
/
Accepted: 21 December 2020
/
Published: 23 December 2020
Abstract
:Simple Summary
Cancer is considered one of the leading causes of human mortality in the world and is the subject of much research. The risk of developing cancer depends on genetic factors, as well as the body’s immune status. The intestinal microbiome plays very important role in maintaining homeostasis in the human body. Probiotics have gained increasing medical significance due to the beneficial effect on the human body associated with the prevention and support of the treatment of many chronic diseases, including cancer in the absence of side effects. The aim of this review was to summarize the knowledge about the effect of probiotic microorganisms in the prevention of cancer. There is a lot of evidence that the use of probiotics can play an important role in cancer prevention and support anti-cancer therapies.
Abstract
The gut microbiome can play important role in maintaining homeostasis in the human body. An imbalance in the gut microbiome can lead to pro-inflammatory immune responses and the initiation of disease processes, including cancer. The research results prove some strains of probiotics by modulating intestinal microbiota and immune response can be used for cancer prevention or/and as adjuvant treatment during anticancer chemotherapy. This review presents the latest advances in research into the effectiveness of probiotics in the prevention and treatment support of cancer. The described issues concern to the anticancer activity of probiotic microorganisms and their metabolites. In addition, we described the potential mechanisms of probiotic chemoprevention and the advisability of using probiotics.
1. Introduction
The human gastrointestinal tract is a reservoir of a complex and dynamic population of microorganisms (the gut microbiota) mainly containing bacteria (in number over 1014), which exerts a significant influence on the host during homeostasis and disease [1]. The presence of such a large count of intestinal bacteria means that the human body has about 10 times more prokaryotic cells than eukaryotic cells [2]. In the human intestines are found bacterial phyla: Firmicutes, Bacteroides, Actinobacteria, Fusobacteria, Proteobacteria, Verrucomicrobia, Sinicobacteria and Spirochaetes [3]. Two bacterial phyla, gram-positive Firmicutes (Bacillus spp., Lactobacillus spp. and Clostridium spp.) and gram-negative Bacteroidetes, predominate in human gut and represent about 90% of the bacterial population [4,5]. The gut microbiota develops and matures during the first 3 years of human life [2]. Enterotype (the type and proportion of microorganisms found in the intestines) may indirectly affect the host’s energy balance. The appropriate balance between bacterial populations ensures homeostasis of the gastrointestinal tract. However, the composition of the intestinal microbiome is susceptible to change. Thus, many factors such as improper diet, stress, gastrointestinal diseases, obesity or taking medications can lead to intestinal homeostasis disorders. As a result of imbalance of the digestive system may be proinflammatory immune responses and initiate disease processes, including cancer. Intestinal dysbiosis may be the reason for the tumorigenesis of both local gastro-intestinal cancers and tumors localized in distant sites of the body [6].
The use of probiotics has a beneficial effect on the human gut microbiome. Their main advantage is the effect on the development of the microbiota inhabiting the organism in the way ensuring proper balance between the bacteria that are necessary for a normal function of the organism and pathogens. Beneficial functions of probiotics lead to the restoration (in case of disturbance) and maintenance of intestinal homeostasis. Probiotics are live microorganisms which, when administered in adequate amounts, confer a health benefit on the host [7,8]. The sources of probiotics in the human diet are mainly silage (e.g., cabbage and cucumbers) and fermented milk products (e.g., yogurt, kefir). Probiotic microorganisms commonly used in human nutrition belong mainly to the genera: Lactobacillus, Bifidobacterium, Lactococcus, Streptococcus and Enterococcus. Moreover, some strains of Bacillus and Saccharomyces are used [9].
2. The Use of Probiotics in the Chemoprevention of Cancer
Goldin and Gorbach [10] were among the first to indicate a relationship between a diet enriched with Lactobacillus and a reduction in the incidence of colorectal cancer (by 37% compared to controls). The results of many in vitro studies indicate beneficial properties of probiotics in modulating the proliferation and apoptosis of cancer cells including, e.g., gastric, colonic, and myeloid leukemia cells (Table 1). Most of the in vitro studies presented were performed on human colonic cancer cells. Many researchers indicate a significant antiproliferative role and/or induction of apoptosis mus musculus colon carcinoma (HGC-27) and human colonic cancer cells (Caco-2, DLD-1, HT-29) [11,12,13,14,15] and also lowering the level of IL–8 [16] by the strain Lactobacillus rhamnosus GG. In addition, scientists’ reports indicate the effectiveness of probiotic microorganisms (e.g., Bacillus: polyfermenticus, subtilis; Bifidobacterium: lactis, adolescentis; Clostridium butyricum; Enterococcus faecium; Lactobacillus: acidophilus, casei, fermentum, delbrueckii, helveticus, paracasei, pentosus, plantarum, salivarius, Lactococcus lactis; Pediococcus pentosaceus, Propionibacterium acidopropionici, Streptococcus thermophilus) in reducing proliferation and/or induction of apoptosis human colonic cancer cells such as Caco-2, HT-29, SW1116, HCT116, SW480, DLD-1, LoVo (Table 1). Moreover, Lactobacillus acidophilus CL1285 and Lactobacillus casei LBC80R (in the presence of 5-FU) induced of apoptosis on human colorectal cells (LS513) [17], while Lactobacillus acidophilus SNUL, Lactobacillus casei YIT9029 and Bifidobacterium longum HY8001 suppressed proliferation of human colorectal (SNUC2A) and gastric carcinoma cells (SNU1) [18]. A drug that is used during chemotherapy, 5–fluorouracil (5–FU), often affects the occurrence of diarrhea. A beneficial effect of lowering cell colony formation in human colonic epithelial cells (NMC460) was found in Bacillus polyfermenticus [19].
Many studies on the anti-tumor effects of probiotics are carried out in animal models. The results of most of this type of research proved to be promising and indicated a potential clinical application. Research related to the effect of probiotic bacteria to animals with tumor-bearing or tumor-induced, is presented in Table 2. These results show that probiotics have anti-cancer properties. The treatment of Bacillus polyfermenticus, Bifidobacterium: infantum, bifidum, and Lactobacillus: acidophilus, casei, lactis, plantarum, rhamnosus, salivarius significantly inhibited the development of colon cancer in rats or mice injected with carcinogenic 1,2-dimethylhydrazine (DMH) (Table 2). In different studies, administration of probiotics such as Pediococcus pentosaceus or Lactobacillus plantarum in mice induced apoptosis and decreased the incidence of azoxymethane (AOM)-induced cancer [23,34]. The reduction the incidence of dextran sulfate sodium (DSS)-induced cancer was observed in the result of the administration in mice probiotics mixture VSL#3 [35] or Lactobacillus plantarum [23]. Probiotics (L. plantarum, L. rhamnosus, mixture VSL#3, B. polyfermenticus, B. lactis KCTC 5727) proved effective in the treatment of cancer induced respectively: CT26 cells injection, MNNG, TNBS, DLD-1 cells injection and in the case no injection (Table 2). Despite the promising results, these results should be interpreted with caution due to the fact that most of the tumors were induced by various chemical agents, which was quite different from the natural process of carcinogenesis.
Due to the antiproliferative role and proapoptotic effects of probiotic strains toward various carcinoma cells (in vitro studies using cell lines) and beneficial effects in animal’s model (in vivo studies), probiotics-based regimens might be used in the cancer prevention and as an adjuvant treatment during anticancer chemotherapy (Table 3).
The results of many clinical studies indicate the effectiveness of probiotics in preventing, treating and reducing the progression of several types of cancer including colorectal, liver, breast, bladder, colon, and cervical in cancer patients (Table 4). The research results indicate, among others that peri-operative administration of probiotics effectively reduces post-operative infectious complications. Infection during abdominal surgery, which is considered a factor in patients’ morbidity, can be reduced by administering probiotics to patients prior to surgery. In addition, it turns out that certain probiotic microorganisms are useful in the control of various intestinal disorders, including fever, postoperative inflammatory diseases, viral diarrhea and antibiotic or chemotherapy/radiotherapy-associated diarrhea.
3. Mechanism of Probiotics Action in Cancer Prevention and Therapy
The anticarcinogenic activity of probiotics is based on: (1) modification of the intestinal microbiota composition, (2) metabolic activity of the intestinal microbiota, (3) production of compounds with anticarcinogenic activity, such as short-chain fatty acids and conjugated linoleic acid, (4) inhibition of cell proliferation and induction apoptosis in cancer cells, (5) influence on other mutagenic and carcinogenic factors, (6) binding and degradation of carcinogenic compounds present in the intestinal lumen, (7) immunomodulation and (8) improvement of the intestinal barrier. Figure 1 shows potential mechanisms of action of probiotics in the prevention of colorectal cancer development.
3.1. Modification of the Intestinal Microbiota Composition
The healthy intestinal microbiota must be properly balanced and diversified to ensure homeostasis (eubiosis). Disturbance of the intestinal microbiota balance may result in a shortage of beneficial bacteria and an excess of pathogens (dysbiosis). Moreover, dysbiosis can cause a chronic inflammation and raise the production of carcinogenic compounds which increases the risk of developing colorectal cancer [67,68].
Sobhani et al. [69] compared samples of feces from healthy people and colorectal cancer patients. Their research shows that the number of Bacteroides and Prevotella genus were significantly higher in the colorectal cancer group. In the intestinal ecosystem, several species of the Lactobacillus type were present in lower amounts than bacteria of the genera Bacteroides, Eubacterium, Fusobacterium, Prevotella and Proteobacteria. It was also found that several species of the genus Salmonella and Clostridium were present in greater numbers in patients with colorectal cancer [70]. Some strains of Bacteroides spp. and Clostridium spp. are classified as bacteria which are involved in the pathogenesis of colorectal cancer. Bacteroides fragilis produces enterotoxigenic toxin (fragilysin), which affects the induction of inflammatory mediators, which leads to the progression of cancer [71]. The pathogenic strain of Escherichia coli can synthesize several toxins, for example, cytotoxic necrotizing agent (CNF), cytolethal distending toxins (CDT), and other various virulence factors. Streptococcus gallolyticus and Enterococcus faecalis can also be connected to colorectal cancer [72,73].
The intestinal microbiota has been linked to development of the gastrointestinal cancers also by production of toxic and genotoxic bacterial metabolites that can lead to mutations by binding specific cell surface receptors and affecting intracellular signal transduction [74].
Competitive exclusion of pathogenic microorganisms by probiotics can be related to competition for nutrients and adhesion to the intestinal mucosa. There are limited nutrients available at the distal part of the colon. Probiotics compete for nutrients and grow at the expense of different intestinal microbiota [75].
Probiotic strains compete with pathogenic microorganisms for adhesion and colonization to biological membranes, forming stable, thin layers called a biofilm [76]. The microbiological biofilm develops as a result of the adhesion of microorganisms to the surface and the production of extracellular polymers that facilitate adhesion and form a structural matrix. Biofilm is characterized by structural heterogeneity, genetic diversity, complexity of interaction and the presence of extracellular substances, i.e., polysaccharides, proteins, nucleic acids, phospholipids. The secretion of these compounds is the result of adaptation to the environment. Polysaccharide polymer substances play a major role in adhesion entirely, bridging the gaps created between microorganisms in biofilm. In the result, microorganisms become focused through polymerization or association. In the first stages of biofilm formation, polysaccharides are secreted the most intensively and help first cells attach to the surface. On the other hand, proteins are first accumulated on the cell surface and then, when released, they associate on the target surface. This is usually a mixture of collagen, elastin and other proteins. They form an extracellular matrix to which microorganisms adhere [77,78].
The results of few clinical trial studies showed the beneficial effect of probiotics on the composition of gut microbiota, thus on the host by improving intestinal barrier integrity, inhibiting growth of pathogens, and reducing metabolism of pro-carcinogenic substances. Ohara et al. investigated the differences between the intestinal flora of colorectal cancer patients and healthy subjects and assessed the possibility of using probiotics to prevent colorectal carcinogenesis [60]. After ingestion of the probiotic (Lactobacillus gasseri OLL2716: LG21), the Lactobacillus detection rate increased, a decrease in the total amount of Clostridium perfringens was found. Moreover, fecal pH indicated acidosis, synthesis of fecal putrefaction products was inhibited, while increase in the short-chain fatty acids and isobutyric acid concentration was observed.
A deterioration of the intestinal environment was observed in the colorectal cancer patients in comparison to the healthy controls, and the intestinal environment improved when probiotics was taken, suggest the possibility of preventing colorectal carcinoma with probiotics. Kotzampassi et al. demonstrated beneficial effects of probiotics (Lactobacillus: acidophilus, plantarum; Bifidobacterium lactis and Saccharomyces boulardii) in patients undergoing colorectal surgery for cancer [63]. The probiotics formulation significantly decreased the risk of postoperative complications, namely mechanical ventilation, infections and anastomotic leakage.
3.2. Metabolic Activity of the Intestinal Microbiota
Some bacteria present in the human intestines are capable of producing carcinogenic compounds from the diet, as well as from the bile salts endogenously produced. This ability is due to the presence and activity of some enzymes, such as azoreductase, β-glucuronidase, β-glucosidase, nitrate reductase, all of which are capable of converting heterocyclic aromatic amines, polycyclic aromatic hydrocarbons, and primary bile acids into active carcinogens and synthetize aglycones, ammonia, cresols, phenols and N-nitroso compounds [79]. These metabolites have genotoxic and cytotoxic activities, which can lead to abnormal cell growth and activation of anti-apoptotic pathways in the colonocytes, thereby contributing to the development of colorectal cancer [67]. Changing the microbial metabolism by modulating the activity of these enzymes may be one of the mechanisms by which the probiotics can reduce the risk of developing colorectal cancer.
Goldin and Gorbach [80] have shown a beneficial effect of Lactobacillus acidophilus (strain N-2 and NCFM) on the activity of azoreductase, nitroreductase and β-glucuronidase in 21 healthy volunteers. These fecal enzymes can catalyze procarcinogens conversion to a proximal carcinogen. Lactobacillus strains caused a significant decline in the specific activity of the three enzymes in all subjects after ten days of feeding.
Some species of pathogenic bacteria, such as Bacteroides, Clostridium, Escherichia coli and Eubacterium exhibit higher activity of enzymes responsible for the synthesis of carcinogenic compounds. In other side, probiotic microorganisms can reduce the population of pathogenic bacteria in the intestinal microbiota and consequently reduce the intestinal production of carcinogenic compounds [81].
Metabolites of lactic acid bacteria (LAB) play an important role in controlling the intestinal microbiota. Among compounds that inhibit the development of pathogenic microorganisms, organic acids are considered to be the most important, in particular lactic and acetic acid as well as hydrogen peroxide and bacteriocins [82].
The antibacterial effect of organic acids can be the result of a sharp decrease in pH outside the optimum value for growth and inhibition of the biochemical activity of microorganisms by undissociated acid molecules. Lactic acid has preservative abilities, in which three factors play an important role: the effect of pH itself, the degree of acid dissociation, the specific activity of the acid molecule. Lactic acid as a weak acid in liquid environments is only partially dissociated. As a lipophilic compound in undissociated form, it can penetrate the lipid cell membrane. Inside the cell, at a higher pH of the cytoplasm, lactic acid dissociates, acidifying the cell content. By releasing potentially toxic hydroxide anions, it interferes with the proton gradient in the membranes, which is the driving force behind active transport. The acidification of the cytoplasm by lactic acid is one of the most important inhibitors of microbial growth [83]. On the other hand, acetic acid can affect cell membranes by neutralizing the electrochemical potential of the cell [84]. The presence of acetic acid can cause denaturation of intracellular proteins and a decrease in the pH value inside the cell. Acetic acid in the presence of lactic acid shows synergism in inhibiting yeast and mold growth, and may also affect the development of putrefactive bacteria, Clostridium and Salmonella. Inhibitors are also other acids, i.e., propionic, formic, benzoic, but they are produced in small quantities, and their action is based on obtaining a synergistic effect [85]. The activity of hydrogen peroxide results from strong oxidizing properties. Hydrogen peroxide can inhibit the development or kill other microorganisms that have low levels of H2O2-degrading enzymes, such as peroxidase, catalase, and superoxide dismutase, protecting against oxidation of disulfide bridges in cellular proteins. In vitro studies confirm the inhibition of various bacteria such as Staphylococcus aureus, Salmonella Typhimurium, Escherichia coli, Clostridium perfringens, Clostridium butyricum and Pseudomonas spp. by hydrogen peroxide [86,87]. On the other hand, it should be emphasized that hydrogen peroxide can also have an effect on all strict anaerobes, including beneficial gut bacteria.
Furthermore, probiotics produce bacteriocins, which are deadly or bacteriostatic on sensitive microorganisms, affecting the cell membranes of bacteria having receptors capable of attaching them. Bacteriocins can cause: a portion of the bacterial cytoplasmic membrane that leads to the dissipation of the transmembrane potential and induces the leakage of K + ions, ATP and amino acids from the cytoplasm of attacked cells; cell lysis and interfering or inhibiting the synthesis of DNA, RNA and proteins (act as DNAase or RNAase) [88,89]. The bacteriocins produced by probiotics e.g., are bifidocin B produced by Bifdobacterium bifidum, nisin from Lactococcus lactis and lactacin B from Lactobacills acidophilus. Bifidocin B, which is produced by Bifidobacterium bifidum NCFB 1454, exerts a strong inhibitory activity against several pathogenic bacteria, including Salmonella Typhimurium SL1344 and Escherichia coli C1845 [90,91]. On the other hand, Drissi et al. explored the role of bacteriocins may have in the GIT. In a genome mining research 641 genomes (307 whole genomes and 334 draft genomes) from microorganisms in the human gut were received. The genomes represented 199 bacterial genera, including Lactobacillus, Streptococcus, Clostridium, and Bacillus. Of the 317 bacteriocins, 175 were from Firmicutes (which includes LAB), 79 from Proteobacteria, 34 from Bacteroidetes, and 25 from Actinobacteria. The authors suggested that bacteriocins in the GIT may have low levels of antimicrobial activity and may thus not have such a drastic effect on microbial populations. However, it was also suggested if bacteriocins play a lesser role in population dynamics, they may have a greater role to play in quorum sensing, or possibly in host immune modulation [92].
3.3. Production of Compounds with Anticarcinogenic Activity, Such as Short-Chain Fatty Acids and Conjugated Linoleic Acid
Short-chain fatty acids (SCFA) are organic acids consisting of 1–6 carbon atoms in the aliphatic chain, including acetic, propionic, butyric, valeric and caproic acid. SCFAs are the main and final products of the metabolism of light-living bacteria large intestine [93]. The daily production of SCFAs in the large intestine of a healthy person should range between 300 and 400 mM, and the total concentration of these acids in the intestinal lumen should be 60–150 mM [94,95]. The molar ratio of acetate, propionate and butyrate produced in the colon is 60:25:15, respectively, but these proportions may be subject to modulation and change [96]. Although their daily production varies around 300 mM, only 10 mmol per day are excreted. This is because SCFAs are absorbed and targeted in the colon at a concentration of 6 to 12 μmol/cm-2/h [97]. SCFAs are a source of energy for colonocytes and promote acidosis and apoptosis of cancer cells, thus promoting an acidic environment that inhibits the formation of high levels of secondary bile acids. Butyric acid helps regulate the balance between proliferation, division, and apoptosis of colonocytes. About 70–90% butyrate is metabolized by colonocytes. The occurrence of that acid in the feces of healthy people is in larger amounts compared to colorectal cancer patients. In addition, it is assumed that a reduction in the 1 µg/L butyrate concentration in stools increases the risk of colon cancer by 84.2%. However, when the concentration of acetic acid is reduced by 1 µg/L, the probability of developing adenoma increases by 71.3% [70].
SCFAs are naturally produced by the bacteria that compose the intestinal microbiota. However, the amount produced may not be sufficient for inhibiting the development of colorectal cancer. Thus, consumption of probiotics may contribute to the increase of the daily production of SCFAs. The presence of SCFAs in the lumen of the large intestine can inhibit the growth of pathogens, e.g., Salmonella Typhimurium in the presence of propionate and butyrate, inhibits the expression of invasive genes encoding Salmonella SPI-1 pathogen islands and prevents attack on cells in tissue culture in vitro [98].
Butyric acid can contribute in the prevention of colorectal cancer because it is capable of improving the intestinal barrier through the increase in mucus production, and in the proliferation of healthy cells (Figure 2).
Butyrate has the ability to decrease the production of inflammatory cytokines by: inhibiting the activation of nuclear transcription factor kappa B, regulating the activity of proteins involved in apoptosis (Bcl-2, Bak, and caspases 3 and 7), increasing the immunogenicity of tumor cells, suppressing cyclooxygenase (COX)-2 activity, increasing the activity of the antioxidant enzyme glutathione S-transferase (GST) and inhibiting the deacetylation of histones. These effects can result in silencing or regulation of genes involved in the control of cell cycle proliferation, differentiation, and apoptosis [101].
SCFAs have immunomodulatory functions that affect the inflammatory response in selected cases by interacting with receptors that are coupled to G protein in the intestine. G protein is a heterotrimeric guanidine nucleotide binding protein. Neurotransmitters or chemokines transmit cell signals with the participation of G protein [102].
Conjugated linoleic acids (CLAs) are isomers of linoleic acid (LA) in which the carbon pairs involved in the formation of double bonds are adjacent to each other. The most commonly found CLA isomers in nature are the cis-9, trans-11 and the trans-10, cis-12. Both isomers are possessing effect against the spread of colon cancer cells [103].
CLAs have influence on the expression of genes involved in the apoptosis process (Bcl-2, caspase 3 and 9) and the cellular response to cell growth factors. In addition, CLAs are able to suppress the production of eicosanoids in colonocytes in two ways. The first consists of the replacement of arachidonic acid in the cell membranes by CLAs, and the second is the result of the interference of CLAs in the activity of the cyclooxygenase and lipoxygenase enzymes, which are responsible for the synthesis of eicosanoids [104].
According to Ewaschuk et al. some species of probiotic bacteria, such as Lactobacillus (acidophilus, casei, delbrueckii, plantarum), Bifidobacterium (breve, infantis, longym), Propionibacterium freudenreichii and Streptococcus salivaris subsp. thermophilus are capable of producing CLAs from linoleic acid [105]. This fatty acid is produced in the distal ileum by bacteria and can be absorbed by or interact with the colonocytes in the intestinal lumen, thus exerting its beneficial effects.
3.4. Inhibition of Cell Proliferation and Induction Apoptosis in Cancer Cells
Proliferation and apoptosis of cancer cells determine the rate of cancer development. During the cancer development process, these cells proliferate more than undergo apoptosis. Probiotics that are able to modulate the cellular proliferation and apoptosis are of great interest because cancer cells would be eliminated less aggressively, and apoptosis brings no damage to the neighbor cells and does not cause inflammation [106]. The apoptosis signaling pathways can be activated by probiotic bacteria (e.g., lactic acid bacteria; LAB) through a mitochondria-dependent (intrinsic) and a death receptor–dependent, mitochondria-independent (extrinsic) pathway (Figure 3) [50].
Baldwin et al. evaluated the antiproliferative activity of Lactobacillus acidophilus and Lactobacillus casei against LS513 gastric cell lines through cellular apoptosis [17]. Hwang et al. demonstrated that probiotic induced apoptosis in gastric cancer cells (KATO3) by inhibiting NF-κB and mTOR-mediated signaling [108]. Cousin et al. demonstrated that probiotic bacteria induce chromatin condensation, apoptotic bodies, DNA fragmentation, caspase activation, inactivation of mitochondrial trans-membrane potential and cell cycle arrest [27]. Probiotic strains such as Lactobacillus reuteri have been reported to influence hematological cancers, which enhanced TNF-induced apoptosis in human chronic myeloid leukemia derived cells [31].
The increased incidence of apoptosis of cancer cells induced by probiotics has been attributed to SCFAs, particularly butyrate, which are able to induce epigenetic changes, paralyze the cell cycle, and stimulate the expression of proapoptotic genes. Relationship exist between the amount of SCFAs in the feces and cell proliferation in the colonic crypts [109].
Probiotic Lactobacillus spp. induced selective genotoxic, cytotoxic, pro-apoptotic effects on leukemia and colon cancer cell lines, and as anti-inflammatory effects on macrophage cells at the molecular level.
In vitro experimental evidence suggests that probiotics used for their anti-cancer activity operate via a process of genotoxicity and cytotoxicity against tumor cells. Liu et al. explored the effects of Lactobacillus casei 01 on 4-nitroquinoline Noxide (4-NQO) induced genotoxicity and colon cancer cell line (HT29) [110]. Nami et al. reported that the metabolites from Lactobacillus acidophilus 36 YL exhibited the most potent cytotoxic effect against human cervical cancer cell lines (HeLa) and colorectal cancer cell lines (HT-29) [111].
3.5. Influence on Other Mutagenic and Carcinogenic Factors
Probiotics may have influence on the other mutagenic and carcinogenic factors, thus contribute to the prevention of cancer. They are able to change the activity of some enzymes involved in the cellular detoxification process, preventing the activity of free radicals and carcinogenic substances.
Glutathione S-transferase (GST) is an antioxidant enzyme with detoxifying activity, which inactivates the carcinogens compounds such as reactive oxygen species (ROS) or xenobionts. GST role is the protection of DNA against oxidative damage, which may lead to mutations, and in consequence, favor carcinogenesis. GST gene polymorphisms may affect the functioning of the encoded enzymes, exerting an effect on the level of DNA damage, and therefore may have an indirect influence on the risk of the development of cancer [113]. Probiotics are able to increase the activity of this enzyme through the action of butyrate, which could change the status of histone acetylation, thus increasing the expression of GST [114].
The bacterial enzyme, β-glucuronidase, with broad substrate specificity can hydrolyze a number of glucuronides, causing the release of carcinogens into the colon, including PAH (e.g., benzo[a]pyrene), an important risk determinant for colorectal cancer [49]. β-glucosidase, breaks down plant glycosides cycasin in the colon, generating aglycones, which are carcinogens [115]. Nitroreductase reduces the N-nitro compounds (e.g., nitrobenzenes) to amines, which are usually mutagenic and carcinogenic, nitrate reductase participates in the generation of highly toxic and carcinogenic nitrite. Nitroreductase activity is significantly higher in colon cancer patients than healthy [116]. Commensal bacteria Bacteroides fragilis, Eschericha coli and Clostridium have exhibited high activities of these pro-carcinogenic enzymes (β-glucosidase, β-glucuronidase, nitroreductase). Studies have also demonstrated Bacteroides fragilis Clostridium and Escherichia coli to be over-presented in colon cancer patients [117].
Probiotics modulate the bacterial enzyme activity in the colon by a combination of mechanisms including: (1) competitive exclusion of pathogenic microflora, (2) ability of synthesizing antibacterial substances that inhibit the growth of other microorganism and (3) ability to generate acids that may lower the pH of the colon [118].
Studies have demonstrated that probiotics are able to reduce the activity of above mentioned procarcinogenic enzymes. Lactobacillus rhamnosus GG and Lactobacillus acidophilus treatment led to a significant reduction in β-glucuronidase activity, while the Lactobacillus casei or Lactobacillus plantarum decreased the nitroreductase activity in rats [119]. In different studies Lactobacillus acidophilus KFRI342 inhibited β-glucuronidase and β-glucosidase activity [43]. Bifidobacterium longum was also found to lower these two enzyme activities in healthy rats [120]. Clinical trials showed similar inhibitory effects of probiotic on these carcinogenic enzymes. For instance, studies showed demonstrated that Bifidobacterium species reduced the β-glucuronidase activity in human intestine [121], Lactobacillus rhamnosus GG β-glucuronidase and nitroreductase [122]. Furthermore, Bifidobacterium adolescentis SPM1207 and SPM0212 reduced intestinal β-glucuronidase and β-glucosidase, as well as tryptophanase and urease, producers of putrefactive products linked to higher incidence of colorectal cancer, such as indoles and ammonia [32,120].
3.6. Binding and Degradation of Carcinogenic Compounds Present in the Intestinal Lumen
Carcinogenic compounds may bind to the cell wall of some probiotic bacteria. This to be associated with the occurrence of cationic exchange between the carcinogenic compounds and the peptidoglycan present in the cell walls of some probiotic microorganisms. Thus, carcinogenic compounds would be eliminated together with the bacteria through the feces [123].
Studies have shown, that Bifidobacterium longum, Lactobacillus acidophilus and Streptococcus salivarius strains could bind and cause the release in feces of heterocyclic amines and mutagens such as: 2-amino-3,4-dimethylimidazo [4,5-f] quinoline (MeIQ), 2-amino-3-methyl-3H-imidazo [4,5-f] quinoline (MHIQ), and 5-phenyl-2-amino-l-methylimidazo [4,5-f] pyridine (PhMIP), 3-amino-1-methyl 5 h pyrido[4,3-b] indole acetate (TrpP2) [124,125].
Rowland and Grasso studied in vitro the effect of intestinal microorganisms (of the genera Lactobacillus, Bifidobacterium and Streptococcus) to dimethyl-nitrosamine and showed that bacteria of the genus Lactobacillus most actively degraded these substances [126]. The amine was transformed into its precursor, dimethylamine, as well as to other volatile metabolites. However, Morotomi and Mutai showed a high ability of Lactobacillus casei to detoxify mutagenic heterocyclic amines [127]. They studied effects of live and heat-inactivated bacteria, and only live showed such ability. This may indicate that live bacteria produce metabolites or catalyze reactions that lead to amine detoxification.
The literature also shows that probiotics may have the ability to detoxify mycotoxins that may have carcinogenic properties [128,129]. El-Nezami et al. demonstrated that 5-week supplementation of probiotics reduced the urinary excretion of aflatoxin B(1)-N(7)-guanine, a marker for hepatocyte carcinogenesis, and synbiotic consumption for 12-week significantly reduced colorectal cancer risk [49,128].
3.7. Immunomodulation
Microorganisms of the intestinal microbiota are the main factors stimulating the immune system, which is a condition for the development of lymphoid structures of this system. The immunomodulatory activity of the intestinal microbiota, including probiotic bacteria, is based on three seemingly opposite phenomena: (1) inducing and maintaining a state of immune tolerance to environmental antigens (food and inhalation), (2) induction and control of immune responses against bacterial and viral pathogens and (3) inhibiting autoimmune and allergic reactions.
Probiotic immune stimulation is also manifested in increased production of immunoglobins, increased activity of macrophages, lymphocytes and stimulation of γ-interferon production. The components of the cell wall of lactic acid bacteria stimulate the activity of macrophages, which due to the increased amount of free oxygen radicals and lysosomal enzymes are able to quickly destroy microbes. Probiotic bacteria also have the ability to stimulate cytokines by immunocompetent gastrointestinal cells [130]. Some probiotics can also affect the immune response through the activation of phagocytes and contribute to the maintenance of the state of vigilance, which can eliminate cancer cells in their early stages of development [131].
The adaptive immune response depends on T and B lymphocytes, which are specific for particular antigens whereas innate immune system responds to common structures called pathogen-associated molecular patterns (PAMPs) shared by the vast majority of pathogens. The primary response to pathogens is triggered by pattern recognition receptors (PPRs), which bind PAMPs. The best-studied PPRs are Toll-like receptors (TLRs). TLRs are transmembrane proteins expressed on various immune and nonimmune cells, such as B cells, natural killer cells, macrophages, dendritic cells (DC), epithelial cells, fibroblasts and endothelial cells. Probiotics can suppress intestinal inflammation via the downregulation of TLR expression, secretion of metabolites that may inhibit TNF-α from entering blood mononuclear cells, and inhibition of NF-κB signaling in enterocytes [132].
In the case of colorectal cancer, the proinflammatory cytokines IL-1β, IL-6, IL-8, IL-12, IL-17, and tumor necrosis factor-α (TNF-α) can be associated with the development of cancer [79]. Probiotics are increase the production of anti-inflammatory cytokines and decrease the production of proinflammatory cytokines, and the development of the colon cancer cells can be delayed. In addition, probiotics may decrease the expression of COX-2, an enzyme that catalyzes the production of prostaglandins from arachidonic acid, which has been linked to an increased risk of developing colorectal cancer because it stimulates cell proliferation and the proinflammatory process [67,133].
Probiotic microorganisms also regulate the activity of natural killer (NK) cells. The use of the probiotic Lactobacillus casei subsp. casei in combination with dextran enhances the efficiency of NK cell activity. This property may be linked to intestinal epithelial cell production of IL-15, an important cytokine for NK cells [134]. It was found that Lactobacillus casei Shirota enhanced NK cell activity which was correlated to an IL-12 production, cytokine implicated in NK cells activity [135].
3.8. Improvement of the Intestinal Barrier
The microorganisms of the intestinal microbiota may change the intestinal barrier and permeability. Some of probiotics are able to reduce intestinal permeability because they can modify components of the intestinal barrier, such as intracolonic pH, the production of mucins and the cellular junction proteins [136].
Lower intracolonic pH values (more acidic) inhibit the proliferation of pathogenic and putrefactive bacteria, as well as the activity of bacterial enzymes responsible for the production of carcinogenic compounds [137]. In several in vitro studies has been confirmed that probiotic bacteria inhibit the growth of gram-negative pathogenic microorganisms. This growth-inhibiting activity has generally been attributed to the fact that probiotic strains lower the pH and/or produce lactic acid. For example, strains of L. acidophilus, L. casei subsp. rhamnosus, and Lactobacillus bulgaricus inhibited the growth of clinical isolates of Helicobacter pylori [138,139] and L. casei subsp. rhamnosus strain Lcr35 reduced the growth of enteropathogenic and enterotoxigenic Eschericia coli, and Klebsiella pneumoniae [140]. It was shown that inhibition occurred when the pH of the incubation medium was acid and that no growth inhibition occurred when the pH of the incubation medium was neutral.
Probiotics can regulate intracellular pH, which allows them to survive and maintain metabolic activity at a relatively low pH environment. This is possible due to the rapid secretion of lactic acid from the cells, which causes the cytoplasmic pH in the cells to become more alkaline than the living environment. In addition, biological membranes are relatively impermeable to protons and lactic acid molecules. This creates a pH gradient (ΔpH) between the cytoplasm and their living environment.
The mucin protective layer, which is produced by goblet cells are gel-forming glycoproteins act as lubricants and as a protective barrier between the body and the external environment. At least nine human mucin (MUC) genes have been identified, and MUC1, MUC2, MUC3, MUC4 and MUC5AC are expressed in the human colon. MUC2 is the major gel-forming mucin of the small and large intestines and is the main structural component of the mucus gel. The carcinogenic process decreases the production of mucins and makes their composition less glycosylated. Some probiotics are able to increase the production of mucins by goblet cells through the upregulation of the MUC genes, mainly MUC2. For in vivo studies, Wistar rats were orally administered the probiotic VSL#3 (Lactobacillus: plantarum, acidophilus, casei, delbrueckii subsp. bulgaricus, Bifidobactrium: infantis, breve, longum and Streptococcus salivarius subsp. thermofilus) on a daily basis for seven days. Probiotic significantly stimulated colonic mucin MUC (by 60%) secretion and MUC2 gene expression, (up to 5 times) however, MUC1 and MUC3 gene expression were only slightly elevated [141].
The inflammatory and carcinogenic processes increase intestinal permeability, mainly because they change the structure and expression of the cellular junction proteins, which makes colonocytes adhere to each other. These proteins are found mostly in the apical region between the colonocytes and are formed by a complex of transmembrane proteins that bind to the colonocyte cytoskeleton through the junction transmembrane proteins, forming the tight junctions [76]. Probiotics can reduce intestinal permeability because they can change the distribution of cell junction proteins and improve the distribution of these proteins throughout the colonic epithelium, making it more continuous. n the small intestine of healthy subjects, administration of Lactobacillus plantarum WCFS1 induced changes in the epithelial tight junctions, resulting in increased staining of the scaffold protein zonula occludens-1 and the transmembrane protein occluding. Lactobacillus plantarum induced translocation of zonula occludens-1 to the tight-junction region was also seen in an in vitro model of the human epithelium, and this significantly protected against chemically induced disruption of the tight junction and the associated increase in epithelial permeability. The mechanism was shown to be dependent on Toll-like receptor 2 signaling and highlights the homeostatic role of innate signaling pathways in maintaining human intestinal epithelial barrier functions [142].
4. Conclusions
Probiotics have gained increasing medical significance due to the beneficial effect on the human body associated with the prevention and support of the treatment of many diseases in the absence of side effects. Due to the potential mechanisms of action presented in this paper, probiotic microorganisms can have a beneficial effect both locally and on the body as a whole. The probiotic properties of microorganisms are a strain trait. There is a lot of evidence that the use of probiotics can play an important role in cancer prevention and support anti-cancer therapies. As a result of laboratory research, many promising results were obtained, which indicate the antitumor effect of probiotics. However, the presented research results confirm the effectiveness of probiotics only for potential prevention of cancer or as adjuvant treatment during anticancer chemotherapy. Clinical trials are still not enough to unambiguously confirm the potential of probiotic microorganisms in this regard. Therefore, it is very important and desirable to continue research on the anti-cancerogenic properties of specific probiotic strains and their mechanisms of action (especially during treatment). In addition, a randomized, double-blind, placebo-controlled clinical trial should be conducted to obtain approval from the medical community and validate the potential of probiotics as an alternative cancer therapy.
Author Contributions
K.Ś. initiated the manuscript concept and wrote the manuscript. P.M.-K. initiated the manuscript concept and wrote the manuscript. W.Ś. helped in preparing the figures. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Liśkiewicz, P.; Pełka-Wysiecka, J.; Wroński, M.; Bąba-Kubiś, A.; Samochowiec, J. Intestinal flora and the pathophysiology of depression and anxiety disorders—Current state of the art and future perspectives. Psychiatry 2018, 15, 70–76. [Google Scholar]
- Gonzalez, A.; Stombaugh, J.; Lozupone, C.; Turnbaugh, P.J.; Gordon, J.I.; Knight, R. The mind-body-microbial continuum. Dialogues. Clin. Neurosci. 2011, 13, 55–62. [Google Scholar]
- Snyder, L.; Peter, J.E.; Henkin, T.M.; Champress, W. Molecular Genetics of Bacteria, 4th ed.; ASM Press: Washington, DC, USA, 1997; pp. 53–66. [Google Scholar]
- Joseph, N.; Vasodavan, K.; Saipudin, N.A.; Yusof, B.N.M.; Kumar, S.; Nordin, S.A. Gut microbiota and short-chain fatty acids (SCFAs) profiles of normal and overweight school children in Selangor after probiotics administration. J. Funct. Foods 2009, 57, 103–111. [Google Scholar] [CrossRef]
- Lach, G.; Schellekens, H.; Dinan, T.G.; Cryan, J.F. Anxiety, depression, and the microbiome: A role for gut peptides. Neurotherapeutics 2018, 15, 36–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodman, B.; Gardner, H. The microbiome and cancer. J. Pathol. 2018, 244, 667–676. [Google Scholar] [CrossRef] [Green Version]
- Food and Agriculture Organization (FAO). Guidelines for the Evaluation of Probiotics in Food; Report of a Joint FAO/WHO Working Group on Drafting Guidelines for the Evaluation of Probiotics in Food; FAO: London, ON, Canada, 2002. [Google Scholar]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S. Expert consensus document: The international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Markowiak, P.; Śliżewska, K. Effects of probiotics, prebiotics, and synbiotics on human health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef]
- Goldin, B.R.; Gorbach, S.L. Effect of Lactobacillus acidophilus dietary supplements on 1,2-dimethylhydrazine dihydrochloride-induced intestinal cancer in rats. J. Natl. Cancer Inst. 1980, 64, 263–265. [Google Scholar] [CrossRef]
- Altonsy, M.O.; Andrews, S.C.; Tuohy, K.M. Differential induction of apoptosis in human colonic carcinoma cells (Caco-2) by Atopobium, and commensal, probiotic and enteropathogenic bacteria: Mediation by the mitochondrial pathway. Int. J. Food Microbiol. 2010, 137, 190–203. [Google Scholar] [CrossRef]
- Borowicki, A.; Michelmann, A.; Stein, K.; Scharlau, D.; Scheu, K. Fermented wheat aleurone enriched with probiotic strains LGG and Bb12 modulates markers of tumor progression in human colon cells. Nutr. Cancer. 2011, 63, 151–160. [Google Scholar] [CrossRef]
- Orlando, A.; Refolo, M.G.; Messa, C.; Amati, L.; Lavermicocca, P.; Guerra, V.; Russo, F. Antiproliferative and proapoptotic effects of viable or heat-killed Lactobacillus paracasei IMPC2.1 and Lactobacillus rhamnosus GG in HGC-27 gastric and DLD-1 colon cell lines. Nutr. Cancer 2012, 64, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- Russo, F.; Orlando, A.; Linsalata, M.; Cavallini, A.; Messa, C. Effects of Lactobacillus rhamnosus GG on the cell growth and polyamine metabolism in HGC-27 human gastric cancer cells. Nutr. Cancer. 2007, 59, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi-Aliabadi, H.; Mohammadi, F.; Fazeli, H.; Mirlohi, M. Effects of Lactobacillus plantarum A7 with probiotic potential on colon cancer and normal cells proliferation in comparison with a commercial strain. Iran. J. Basic. Med. Sci. 2014, 17, 815–819. [Google Scholar]
- Lopez, M.; Li, N.; Kataria, J.; Russell, M.; Neu, J. Live and ultraviolet-inactivated Lactobacillus rhamnosus GG decrease flagellin-induced interleukin-8 production in Caco-2 cells. J. Nutr. 2008, 138, 2264–2268. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, C.; Millette, M.; Oth, D.; Ruiz, M.T.; Luquet, F.M.; Lacroix, M. Probiotic Lactobacillus acidophilus and L. casei mix sensitize colorectal tumoral cells to 5-fluorouracilinduced apoptosis. Nutr. Cancer. 2010, 62, 371–378. [Google Scholar] [CrossRef]
- Lee, J.W.; Shin, J.G.; Kim, E.H.; Kang, H.E.; Yim, I.B.; Kim, J.Y.; Joo, H.G.; Woo, H.J. Immunomodulatory and antitumor effects in vivo by the cytoplasmic fraction of Lactobacillus casei and Bifidobacterium longum. J. Vet. Sci. 2004, 5, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Ma, E.L.; Choi, Y.J.; Choi, J.; Pothoulakis, C.; Rhee, S.H.; Imet, E. The anticancer effect of probiotic Bacillus polyfermenticus on human colon cancer cells is mediated through ErbB2 and ErbB3 inhibition. Int. J. Cancer. 2010, 127, 780–790. [Google Scholar] [PubMed] [Green Version]
- Saxami, G.; Karapetsas, A.; Lamprianidou, E.; Kotsianidis, I.; Chlichlia, A.; Tassou, C.; Zoumpourliset, V.; Galanis, A. Two potential probiotic lactobacillus strains isolated from olive microbiota exhibit adhesion and anti-proliferative effects in cancer cell lines. J. Funct. Foods 2016, 24, 461–471. [Google Scholar] [CrossRef]
- Tiptiri-Kourpeti, A.; Spyridopoulou, K.; Santarmaki, V.; Aindelis, G.; Tompoulidou, E.; Lampranidou, E.; Saxami, G.; Ypsilantis, P.; Lampri, E.; Simopoulos, C.; et al. Lactobacillus casei exerts anti-proliferative effects accompanied by apoptotic cell death and up-regulation of TRAIL in colon carcinoma cells. PLoS ONE 2016, 11, e0147960. [Google Scholar] [CrossRef]
- Han, K.J.; Lee, N.K.; Park, H.; Paik, H.D. Anticancer and anti-inflammatory activity of probiotic Lactococcus lactis nk34. J. Microbiol. Biotechnol. 2015, 25, 1697–1701. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.K.; Son, S.H.; Jeon, E.B.; Jung, G.H.; Lee, J.Y.; Paik, H.D. The prophylactic effect of probiotic Bacillus polyfermenticus KU3 against cancer cells. J. Funct. Foods 2015, 14, 513–518. [Google Scholar] [CrossRef]
- Chen, Z.F.; Ai, L.Y.; Wang, J.L.; Ren, L.L.; Yu, Y.N.; Xu, J.; Chen, H.Y.; Yu, J.; Li, M.; Qin, W.X.; et al. Probiotics Clostridium butyricum and Bacillus subtilis ameliorate intestinal tumorigenesis. Future. Microbiol. 2015, 10, 1433–1445. [Google Scholar] [CrossRef] [PubMed]
- Ghoneum, M.; Gimzewski, J. Apoptotic effect of a novel kefir product, PFT, on multidrug-resistant myeloid leukemia cells via a hole-piercing mechanism. Int. J. Oncol. 2014, 44, 830–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thirabunyanon, M.; Hongwittayakorn, P. Potential probiotic lactic acid bacteria of human origin induce antiproliferation of colon cancer cells via synergic actions in adhesion to cancer cells and short-chain fatty acid bioproduction. Appl. Biochem. Biotechnol. 2013, 169, 511–525. [Google Scholar] [CrossRef]
- Cousin, F.J.; Jouan-Lanhouet, S.; Dimanche-Boitrel, M.T.; Corcos, L.; Jan, G. Milk fermented by Propionibacterium freudenreichii induces apoptosis of HGT-1 human gastric cancer cells. PLoS ONE 2012, 7, 31892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Oh, S.; Yun, H.S.; Oh, S.; Kim, S.H. Cell-bound exopolysaccharide from probiotic bacteria induces autophagic cell death of tumour cells. Lett. Appl. Microbiol. 2010, 51, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.S.; Molina, M.A.; Di Sciullo, P.; Azpiroz, M.B.; Leocata Nieto, F.; Sterín-Speziale, N.B.; Mongini, C.; Manghi, M.A. Beneficial activity of Enterococcus faecalis CECT7121 in the anti-lymphoma protective response. J. Appl. Microbiol. 2010, 109, 1234–1243. [Google Scholar] [CrossRef] [PubMed]
- Thirabunyanon, M.; Boonprasom, P.; Niamsup, P. Probiotic potential of lactic acid bacteria isolated from fermented dairy milks on antiproliferation of colon cancer cells. Biotechnol. Lett. 2009, 31, 571–576. [Google Scholar] [CrossRef]
- Iyer, C.; Kosters, A.; Sethi, G.; Kunnumakkara, A.B.; Aggarwal, B.B.; Versalovic, J. Probiotic Lactobacillus reuteri promotes TNFinduced apoptosis in human myeloid leukemia-derived cells by modulation of NF-kappaB and MAPK signalling. Cell. Microbiol. 2008, 10, 1442–1452. [Google Scholar] [CrossRef]
- Kim, Y.; Lee, D.; Kim, D.; Cho, J.; Yang, J.; Chung, M.; Kim, K.; Ha, N. Inhibition of proliferation in colon cancer cell lines and harmful enzyme activity of colon bacteria by Bifidobacterium adolescentis SPM0212. Arch. Pharm. Res. 2008, 31, 468–473. [Google Scholar] [CrossRef]
- Jan, G.; Belzacq, A.S.; Haouzi, D.; Rouault, A.; Metivier, D.; Kroemer, G.; Brenner, C. Propionibacteria induce apoptosis of colorectal carcinoma cells via short-chain fatty acids acting on mitochondria. Cell Death Differ. 2002, 9, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Dubey, V.; Ghosh, A.R.; Bishayee, K.; Khuda-Bukhsh, A.R. Appraisal of the anti-cancer potential of probiotic Pediococcus pentosaceus GS4 against colon cancer: In vitro and in vivo approaches. J. Funct. Foods 2016, 23, 66–79. [Google Scholar] [CrossRef]
- Talero, E.; Bolivar, S.; Ávila-Román, J.; Alcaide, A.; Fiorucci, S.; Motilva, V. Inhibition of chronic ulcerative colitis-associated adenocarcinoma development in mice by VSL#3. Inflamm. Bowel. Dis. 2015, 21, 1027–1037. [Google Scholar] [PubMed] [Green Version]
- Lenoir, M.; del Carmen, S.; Cortes-Perez, N.G.; Lozano-Ojalvo, D.; Muñoz-Provencio, D.; Chain, F.; Langella, P.; de Moreno de LeBlanc, A.; LeBlanc, J.G.; Bermúdez-Humarán, L.G. Lactobacillus casei BL23 regulates Tregand Th17 T-cell populations and reduces DMH-associated colorectal cancer. J. Gastroenterol. 2016, 51, 862–873. [Google Scholar] [CrossRef] [PubMed]
- Gamallat, Y.; Meyiah, A.; Kuugbee, E.D.; Chiwala, G.; Awadasseid, A.; Bamba, D.; Zhang, X.; Shang, X.; Luo, F.; Xin, Y. Lactobacillus rhamnosus induced epithelial cell apoptosis, ameliorates inflammation and prevents colon cancer development in an animal model. Biomed. Pharmacother. 2016, 83, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Kuugbee, E.D.; Shang, X.; Gamallat, Y.; Bamba, D.; Awadasseid, A.; Suliman, M.A.; Zang, S.; Ma, Y.; Chiwala, G.; Xin, Y.; et al. Structural change in microbiota by a probiotic cocktail enhances the gut barier and reduces cancer via TLR2 signaling in a rat model of colon cancer. Dig. Dis. Sci. 2016, 61, 2908–2920. [Google Scholar] [CrossRef]
- Zhang, M.; Fan, X.; Fang, B.; Zhu, C.; Zhu, J.; Ren, F. Effects of Lactobacillus salivarius Ren on cancer prevention and intestinal microbiota in 1, 2-dimethylhydrazine-induced rat model. J. Microbiol. 2015, 53, 398–405. [Google Scholar] [CrossRef]
- Walia, S.; Kamal, R.; Dhawan, D.K.; Kanwar, S.S. Chemoprevention by probiotics during 1,2-dimethylhydrazine-induced colon carcinogenesis in rats. Dig. Dis. Sci. 2018, 63, 900–909. [Google Scholar] [CrossRef]
- Hu, J.; Wang, C.; Ye, L.; Yang, W.; Huang, H.; Meng, F.; Shi, S.; Ding, Z. Anti-tumour immune effect of oral administration of Lactobacillus plantarum to CT26 tumourbearing mice. J. Biosci. 2015, 40, 269–279. [Google Scholar] [CrossRef]
- Verma, A.; Shukla, G. Synbiotic (Lactobacillus rhamnosus + Lactobacillus acidophilus + inulin) attenuates oxidative stress and colonic damage in 1,2 dimethylhydrazine dihydrochloride-induced colon carcinogenesis in Sprague’ Dawley rats: A long-term study. Eur. J. Cancer Prev. 2014, 23, 550–559. [Google Scholar] [CrossRef]
- Chang, J.H.; Shim, Y.Y.; Cha, S.K.; Reaney, M.J.T.; Chee, K.M. Effect of Lactobacillus acidophilus KFRI342 on the development of chemically induced precancerous growths in the rat colon. J. Med. Microbiol. 2012, 61, 361–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gosai, V.; Ambalam, P.; Raman, M.; Kothari, C.R.; Kothari, R.K.; Vyas, B.R.M.; Sheth, N.R. Protective effect of Lactobacillus rhamnosus 231 against N-Methyl-N’-nitro-N-nitrosoguanidine in animal model. Gut Microbes 2011, 2, 319–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appleyard, C.B.; Cruz, M.L.; Isidro, A.A.; Arthur, J.C.; Jobin, C.; De Simone, C. Pretreatment with the probiotic VSL#3 delays transition from inflammation to dysplasia in a rat model of colitis-associated cancer. Am. J. Physiol. Liver Physiol. 2011, 301, G1004–G1013. [Google Scholar]
- Bertkova, I.; Hijova, E.; Chmelarova, A.; Mojzisova, G.; Petrasova, D.; Strojny, L.; Bomba, A.; Zitnan, R. The effect of probiotic microorganisms and bioactive compounds on chemically induced carcinogenesis in rats. Neoplasma 2010, 57, 422–428. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Singh, N.K.; Sinha, P.R. Inhibition of 1,2-dimethylhydrazine induced colon genotoxicity in rats by the administration of probiotic curd. Mol. Biol. Rep. 2010, 37, 1373–1376. [Google Scholar] [CrossRef]
- Park, E.; Jeon, G.I.; Park, J.S.; Paik, H.D. A probiotic strain of Bacillus polyfermenticus reduces DMH induced precancerous lesions in F344 male. Rat. Biol. Pharm. Bull. 2007, 30, 569–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rafter, J.; Bennett, M.; Caderni, G.; Clune, Y.; Hughes, R.; Karlsson, P.; Klinder, A.; O’Riordan, M.; O’Sullivan, G.; Pool-Zobel, B.; et al. Dietary synbiotics reduce cancer risk factors in polypectomized and colon cancer patients. Am. J. Clin. Nutr. 2007, 85, 488–496. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Zhang, X.; Covasa, M. Emerging roles of lactic acid bacteria in protection against colorectal cancer. World J. Gastroenterol. 2014, 20, 7878–7886. [Google Scholar] [CrossRef] [PubMed]
- Ohigashi, S.; Hoshino, Y.; Ohde, S.; Onodera, H. Functional outcome, quality of life, and efficacy of probiotics in postoperative patients with colorectal cancer. Surg. Today 2011, 41, 1200–1206. [Google Scholar] [CrossRef]
- Liu, Z.; Qin, H.; Yang, Z.; Xia, Y.; Liu, W.; Yang, J.; Jiang, Y.; Zhang, H.; Wang, Y.; Zheng, Q. Randomized clinical trial: The effects of perioperative probiotic treatment on barrier function and post-operative infectious complications in colorectal cancer surgery-A double-blind study. Aliment. Pharmacol. Ther. 2011, 33, 50–63. [Google Scholar] [CrossRef]
- Österlund, P.; Ruotsalainen, T.; Korpela, R.; Saxelin, M.; Ollus, A.; Valta, P.; Kouri, M.; Elomaa, I.; Joensuu, H. Lactobacillus supplementation for diarrhoea related to chemotherapy of colorectal cancer: A randomised study. Br. J. Cancer 2007, 97, 1028–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Nezami, H.S.; Polychronaki, N.N.; Ma, J.; Zhu, H.; Ling, W.; Salminen, E.K.; Juvonen, R.O.; Salminen, S.J.; Poussa, T.; Mykkänen, H.M. Probiotic supplementation reduces a biomarker for increased risk of liver cancer in young men from Southern China. Am. J. Clin. Nutr. 2006, 83, 1199–1203. [Google Scholar] [CrossRef] [PubMed]
- Rasouli, B.S.; Ghadimi-Darsajini, A.; Nekouian, R.; Iragian, G.R. In vitro activity of probiotic Lactobacillus reuteri against gastric cancer progression by downregulation of urokinase plasminogen activator/urokinase plasminogen activator receptor gene expression. J. Cancer Res. Ther. 2017, 13, 246–251. [Google Scholar] [PubMed]
- Ghoneum, M.; Felo, N. Selective induction of apoptosis in human gastric cancer cells by Lactobacillus kefiri (PFT), a novel kefir product. Oncol. Rep. 2015, 34, 1659–1666. [Google Scholar] [CrossRef] [Green Version]
- Chitapanarux, I.; Chitapanarux, T.; Traisathit, P.; Kudumpee, S.; Tharavichitkul, E.; Lorvidhaya, V. Randomized controlled trial of live Lactobacillus acidophilus plus Bifidobacterium bifidum in prophylaxis of diarrhea during radiotherapy in cervical cancer patients. Radiat. Oncol. 2010, 5, 31. [Google Scholar] [CrossRef] [Green Version]
- Toi, M.; Hirota, S.; Tomotaki, A. Probiotic beverage with soy isoflavone consumption for breast cancer prevention: A case-control study. Curr. Nutr. Food Sci. 2013, 9, 194–200. [Google Scholar] [CrossRef]
- Pala, V.; Sieri, S.; Berrino, F.; Vineis, P.; Sacerdote, C.; Palli, D.; Masala, G.; Panico, S.; Mattiello, A.; Tumino, R. Yogurt consumption and risk of colorectal cancer in the Italian European prospective investigation into cancer and nutrition cohort. Int. J. Cancer 2011, 129, 2712–2719. [Google Scholar] [CrossRef]
- Ohara, T.; Yoshino, K.; Kitajima, M. Possibility of preventing colorectal carcinogenesis with probiotics. Hepatogastroenterol 2010, 57, 1411–1415. [Google Scholar]
- Hatakka, K.; Holma, R.; El-Nezami, H.; Suomalainen, T.; Kuisma, M.; Saxelin, M.; Poussa, T.; Mykkänen, H.; Korpela, R. The influence of Lactobacillus rhamnosus LC705 together with Propionibacterium freudenreichii ssp. shermanii JS on potentially carcinogenic bacterial activity in human colon. Int. J. Food Microbiol. 2008, 128, 406–410. [Google Scholar] [CrossRef]
- Ohashi, Y.; Nakai, S.; Tsukamoto, T. Habitual intake of lactic acid bacteria and risk reduction of bladder cancer. Urol. Int. 2002, 68, 273–280. [Google Scholar] [CrossRef]
- Kotzampassi, K.; Stavrou, G.; Damoraki, G.; Georgitsi, M.; Basdanis, G.; Tsaousi, G.; Giamarellos-Bourboulis, E.J. A four-probiotics regimen reduces postoperative complications after colorectal surgery: A randomized, double-blind, placebo-controlled study. World J. Surg. 2015, 39, 2776–2783. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.H.; Huang, M.J.; Zhang, X.W.; Wang, L.; Huang, N.Q.; Peng, H.; Lan, P.; Peng, J.S.; Yang, Z.; Xia, Y.; et al. The effects of perioperative probiotic treatment on serum zonulin concentration and subsequent postoperative infectious complications after colorectal cancer surgery: A double-center and doubleblind randomized clinical trial. Am. J. Clin. Nutr. 2013, 97, 117–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.W.; Peng, D.; Yang, B.R.; Gao, J.; Fang, W.J.; Ying, C.M. Preoperative probiotics decrease postoperative infectious complications of colorectal cancer. Am. J. M. Sci. 2012, 343, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Wada, M.; Nagata, S.; Saito, M.; Shimizu, T.; Yamashiro, Y.; Matsuki, T.; Asahara, T.; Nomoto, K. Effects of the enteral administration of Bifidobacterium breve on patients undergoing chemotherapy for pediatric malignancies. Support. Care Cancer 2010, 18, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Dos Reis, S.A.; Da Conceição, L.L.; Siqueira, N.P.; Rosa, D.D.; Da Silva, L.L.; Peluzio, M.C.G. Review of the mechanisms of probiotic actions in the prevention of colorectal cancer. Nutr. Res. 2017, 37, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Koboziev, I.; Webb, C.R.; Furr, K.L.; Grisham, M.B. Role of the enteric microbiota in intestinal homeostasis and inflammation. Free. Radic. Biol. Med. 2013, 68, 122–133. [Google Scholar] [CrossRef]
- Sobhani, I.; Tap, J.; Roudot-Thoraval, F.; Roperch, J.P.; Letulle, S.; Langella, P.; Corthier, G.; Tran Van Nhieu, J.; Furet, J.P. Microbial dysbiosis in colorectal cancer (CRC) patients. PLoS ONE 2011, 6, e16393. [Google Scholar] [CrossRef]
- Kahouli, I.; Tomaro-Duchesneau, C.; Prakash, S. Probiotics in colorectal cancer (CRC) with emphasis on mechanisms of action and current perspectives. J. Med. Microbiol. 2013, 62, 1107–1123. [Google Scholar] [CrossRef] [Green Version]
- Boleij, A.; Hechenbleikner, E.M.; Goodwin, A.C.; Badani, R.; Stein, E.M.; Lazarev, M.G.; Ellis, B.; Carroll, K.C.; Albesiano, E.; Wick, E.C. The bacteroides fragilis toxin gene is prevalent in the colon mucosa of colorectal cancer patients. Clin. Infect. Dis. 2015, 60, 208–215. [Google Scholar] [CrossRef]
- Ambalam, P.; Raman, M.; Purama, R.K.; Doble, M. Probiotics, prebiotics and colorectal cancer prevention. Best. Pract. Res. Clin. Gastroenterol. 2016, 30, 119–131. [Google Scholar] [CrossRef]
- Molska, M.; Reguła, J. Potential mechanisms of probiotics action in the prevention and treatment of colorectal cancer. Nutrients 2019, 11, 2453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javanmard, A.; Ashtari, S.; Sabet, B.; Davoodi, S.H.; Rostami-Nejad, M.; Akbari, M.E.; Niaz, A.; Mortazavian, A.M. Probiotics and their role in gastrointestinal cancers prevention and treatment; an overview. Gastroenterol. Hepatol. 2018, 11, 284–295. [Google Scholar]
- Fooks, L.; Gibson, G. Probiotics as modulators of the gut flora. Br. J. Nutr. 2002, 88, S39–S49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohland, C.L.; MacNaughton, W.K. Probiotic bacteria and intestinal barrier function. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, 807–819. [Google Scholar] [CrossRef] [Green Version]
- Davey, M.E.; O’Toole, G.A. Microbial biofilms: From ecology to molecular genetics. Microbiol. Mol. Biol. Rev. 2000, 64, 847–867. [Google Scholar] [CrossRef] [Green Version]
- Dieltjens, L.; Appermans, K.; Lissens, M.; Lories, B.; Kim, W.; Van der Eycken, E.V.; Foster, K.R.; Steenackers, H.P. Inhibiting bacterial cooperation is an evolutionarily robust anti-biofilm strategy. Nat. Commun. 2020, 107, 1–11. [Google Scholar] [CrossRef]
- Zhu, Q.; Gao, R.; Wu, W.; Qin, H. The role of gut microbiota in the pathogenesis of colorectal cancer. Tumor Biol. 2013, 34, 1285–1300. [Google Scholar] [CrossRef]
- Goldin, B.R.; Gorbach, S.L. The effect of milk and Lactobacillus feeding on human intestinal bacterial enzyme activity. Am. J. Clin. Nutr. 1984, 39, 756–761. [Google Scholar] [CrossRef]
- Tjalsma, H.; Boleij, A.; Marchesi, J.R.; Dutilh, B.E. A bacterial driverpassenger model for colorectal cancer: Beyond the usual suspects. Nat. Rev. Microbiol. 2012, 10, 575–582. [Google Scholar] [CrossRef]
- Rountree, R. Proven therapeutic benefits of high-quality probiotics. Appl. Nutr. Sci. Rep. 2002, 4, 1–6. [Google Scholar]
- Boris, S.; Barbés, C. Role played by lactobacilli in controlling the population of vaginal pathogens. Microbes. Infect. 2000, 2, 543–546. [Google Scholar] [CrossRef]
- Eklund, T. Organic acids and esters. In Mechanism of Actions of Food Preservation Procedures; Gould, G.W., Ed.; Scientific Publishers: London, UK, 1989; pp. 161–200. [Google Scholar]
- Fung, W.Y.; Lye, H.S.; Lim, T.J.; Kuan, C.Y.; Liong, M.T. Role of probiotic on gut health. In Probiotics. Biology, genetics and Health Aspects; Liong, M.T., Ed.; Springer: New York, NY, USA, 2001; pp. 139–166. [Google Scholar]
- Dembele, T.; Obdrzalek, V.; Votava, M. Inhibition of bacterial pathogens by lactobacilli. Zentralbl. Bakteriol. 1998, 288, 395–401. [Google Scholar] [CrossRef]
- Tomás, M.S.; Otero, C.M.; Ocaña, V.; Nader-Macias, E.M. Production of antimicrobial substances by lactic acid bacteria I: Determination of hydrogen peroxide. Meth. Mol. Biol. 2004, 268, 337–346. [Google Scholar]
- Klaenhammer, T.R. Genetics of bacteriocins produced by lactic acid bacteria. FEMS Microbiol. Rev. 1993, 12, 39–86. [Google Scholar] [CrossRef]
- Nazir, Y.; Hussain, S.A.; Hamid, A.A.; Song, Y. Probiotics and their potential preventive and therapeutic role for cancer, high serum cholesterol, and allergic and HIV diseases. BioMed Res. Int. 2018, 2018, 1–17. [Google Scholar] [CrossRef]
- Lievin, V.; Peifer, I.; Hudault, S. Bifdobacterium strains from resident infant human gastrointestinal microfora exert antimicrobial activity. Gut 2000, 47, 646–652. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, D.S.; Cho, G.S.; Hanak, A.; Huch, M.; Franz, C.M.; Arneborg, N. The effect of bacteriocin-producing Lactobacillus plantarum strains on the intracellular pH of sessile and planktonic Listeria monocytogenes single cells. Int. J. Food Microbiol. 2010, 141, S53–S59. [Google Scholar] [CrossRef]
- Drissi, F.; Buffet, S.; Raoult, D.; Merhej, V. Common occurrence of antibacterial agents in human intestinal microbiota. Front. Microbiol. 2015, 6, 441. [Google Scholar] [CrossRef] [Green Version]
- Macfarlane, S.; Macfarlane, G.T. Regulation of short-chain fatty acid production. Proc. Nutr. Soc. 2003, 62, 67–72. [Google Scholar] [CrossRef]
- Roediger, W.E. Role of anaerobic bacteria in the metabolic welfare of the colonic mucosa in man. Gut 1980, 21, 793–798. [Google Scholar] [CrossRef] [Green Version]
- Kuczyńska, B.; Wasilewska, A.; Biczysko, M.; Banasiewicz, T.; Drews, M. Krótkołańcuchowe kwasy tłuszczowe—Mechanizm działania, potencjalne zastosowanie kliniczne oraz zalecenia dietetyczne. Now. Lek. 2011, 80, 299–304. (In Polish) [Google Scholar]
- Topping, D.L.; Clifton, P.M. Short-chain fatty acids and human colonic function: Roles of resistant starch and nonstarch polysaccharides. Physiol. Rev. 2001, 81, 1031–1064. [Google Scholar] [CrossRef] [PubMed]
- Arora, T.; Sharma, R.; Frost, G. Propionate. Anti-obesity and satiety factor? Appetite 2011, 56, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Gantois, I.; Ducatelle, R.; Pasmans, F.; Haesebrouck, F.; Hautefort, I.; Thompson, A.; Hinton, J.C.; Van Immerseel, F. Butyrate specifically down-regulates salmonella pathogenicity island 1 gene expression. Appl. Environ. Microbiol. 2006, 72, 946–949. [Google Scholar] [CrossRef] [Green Version]
- Markowiak-Kopeć, P.; Śliżewska, K. The effect of probiotics on the production of short-chain fatty acids by human intestinal microbiome. Nutrients 2020, 12, 1107. [Google Scholar] [CrossRef]
- Sun, M.; Wu, W.; Liu, Z.; Cong, Y. Microbiota metabolite short chain fatty acids, GPCR, and inflammatory bowel diseases. J. Gastroenterol. 2017, 52, 1–8. [Google Scholar] [CrossRef]
- Kumar, M.; Nagpal, R.; Verma, V.; Kumar, A.; Kaur, N.; Hemalatha, R. Probiotic metabolites as epigenetic targets in the prevention of colon cancer. Nutr. Rev. 2012, 71, 23–34. [Google Scholar] [CrossRef]
- Duc, N.M.; Kim, H.R.; Chung, K.Y. Structural mechanism of G protein activation by G protein-coupled receptor. Eur. J. Pharm. 2015, 763, 214–222. [Google Scholar] [CrossRef]
- Soel, S.M.; Choi, O.S.; Bang, M.H.; Yoon Park, J.H.; Kim, W.K. Influence of conjugated linoleic acid isomers on the metastasis of colon cancer cells in vitro and in vivo. J. Nutr. Biochem. 2007, 18, 650–657. [Google Scholar] [CrossRef]
- Bassaganya-Riera, J.; Viladomiu, M.; Pedragosa, M.; Simone, C.; Hontecillas, R. Immunoregulatory mechanisms underlying prevention of colitis-associated colorectal cancer by probiotic bacteria. PLoS ONE 2012, 7, 1–8. [Google Scholar] [CrossRef]
- Ewaschuk, J.B.; Walker, J.W.; Diaz, H.; Madsen, K.L. Bioproduction of conjugated linoleic acid by probiotic bacteria occurs in vitro and in vivo in mice. J. Nutr. 2006, 136, 1483–1487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fearon, E.R. Molecular genetics of colorectal cancer. Annu. Rev. Pathol. Mech. 2011, 6, 479–507. [Google Scholar] [CrossRef] [PubMed]
- De Vries, E.G.; Gietema, J.A.; de Jong, S. Tumor necrosis factor-related apoptosis-inducing ligand pathway and its therapeutic implications. Clin. Cancer Res. 2006, 12, 2390–2393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, J.W.; Baek, Y.M.; Yang, K.E.; Yoo, H.S.; Cho, C.K.; Lee, Y.W. Lactobacillus casei extract induces apoptosis in gastric cancer by inhibiting NF-κB and mTOR-mediated signalling. Integr. Cancer Ther. 2013, 12, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.M.; Yu, Y.N.; Wang, J.L.; Lin, Y.W.; Kong, X.; Yang, C.Q. Decreased dietary fiber intake and structural alteration of gut microbiota in patients with advanced colorectal adenoma. Am. J. Clin. Nutr. 2013, 97, 1044–1052. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.T.; Chu, F.J.; Chou, C.C.; Yu, R.C. Antiproliferative and anticytotoxic effects of cell fractions and exopolysaccharides from Lactobacillus casei 01. Mutat. Res. 2011, 721, 157–162. [Google Scholar] [CrossRef]
- Nami, Y.; Abdullah, N.; Haghshenas, B.; Radiah, D.; Rosli, R.; Khosroushahi, A.Y. Probiotic potential and biotherapeutic effects of newly isolated vaginal Lactobacillus acidophilus 36YL strain on cancer cells. Anaerobe 2014, 28, 29–36. [Google Scholar] [CrossRef]
- Shinnoh, M.; Horinaka, M.; Yasuda, T.; Yoshikawa, S.; Morita, M. Clostridium butyricum MIYAIRI 588 shows antitumor effects by enhancing the release of TRAIL from neutrophils through MMP-8. Int. J. Oncol. 2013, 42, 903–911. [Google Scholar] [CrossRef] [Green Version]
- Klusek, J.; Głuszek, S.; Klusek, J. GST gene polymorphisms and the risk of colorectal cancer development. Contemp. Oncol. (Pozn.) 2014, 18, 219–221. [Google Scholar] [CrossRef] [Green Version]
- Pool-Zobel, B.; Veeriah, S.; Bohmer, F.D. Modulation of xenobiotic metabolising enzymes by anticarcinogens—Focus on glutathione S-transferases and their role as targets of dietary chemoprevention in colorectal carcinogenesis. Mutat. Res. 2005, 591, 74–92. [Google Scholar] [CrossRef]
- McIntosh, F.M.; Maison, N.; Holtrop, G.; Young, P.; Stevens, V.J.; Ince, J.; Johnstone, A.M.; Lobley, G.E.; Flint, H.J.; Louis, P. Phylogenetic distribution of genes encoding b-glucuronidase activity in human colonic bacteria and the impact of diet on faecal glycosidase activities. Environ. Microbiol. 2013, 14, 1876–1887. [Google Scholar] [CrossRef] [PubMed]
- Hamer, H.M.; De Preter, V.; Windey, K.; Verbeke, K. Functional analysis of colonic bacterial metabolism: Relevant to health? Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G1–G9. [Google Scholar] [CrossRef] [PubMed]
- Culpepper, B.S.T.; Mai, V. Evidence for contributions of gut microbiota to colorectal carcinogenesis. Curr. Nutr. Rep. 2013, 2, 10–18. [Google Scholar] [CrossRef] [Green Version]
- Commane, D.; Hughes, R.; Shortt, C.; Rowland, I. The potential mechanisms involved in the anti-carcinogenic action of probiotics. Mutat. Res. 2005, 591, 276–289. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.; Shukla, G. Probiotics Lactobacillus rhamnosus GG, Lactobacillus acidophilus suppresses DMH-induced procarcinogenic fecal enzymes and preneoplastic aberrant crypt foci in early colon carcinogenesis in sprague dawley rats. Nutr. Cancer 2013, 65, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.K.; Jang, S.; Baek, E.H.; Kim, M.J.; Lee, K.S.; Shin, H.S.; Chung, M.J.; Kim, J.E.; Lee, K.O.; Ha, N.J. Lactic acid bacteria affect serum cholesterol levels, harmful fecal enzyme activity, and fecal water content. Lipids Health Dis. 2009, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Bouhnik, Y.; Flourie, B.; Andrieux, C.; Bisetti, N.; Briet, F.; Rambaud, J. Effects of Bifidobacterium sp fermented milk ingested with or without inulin on colonic bifidobacteria and enzymatic activities in healthy humans. Eur. J. Clin. Nutr. 1996, 50, 269–273. [Google Scholar]
- Ling, W.; Korpela, R.; Mykkänen, H.; Salminen, S.; Hänninen, O. Lactobacillus strain GG supplementation decreases colonic hydrolytic and reductive enzyme activities in healthy female adults. J. Nutr. 1994, 124, 18–23. [Google Scholar] [CrossRef]
- Burns, A.J.; Rowland, I.R. Antigenotoxicity of probiotics and prebiotics on faecal water-induced DNA damage in human colon adenocarcinoma cells. Mutat. Res. 2004, 551, 233–243. [Google Scholar] [CrossRef]
- Bolognani, F.; Rumney, C.J.; Rowland, I.R. Influence of carcinogen binding by lactic acid-producing bacteria on tissue distribution and in vivo mutagenicity of dietary carcinogens. Food Chem. Toxicol. 1997, 35, 535–545. [Google Scholar] [CrossRef]
- Orrhage, K.; Sillerstrom, E.; Gustafsson, J.A.; Nord, C.E.; Rafter, J. Binding of mutagenic heterocyclic amines by intestinal and lactic acid bacteria. Mutat. Res. 1994, 311, 239–248. [Google Scholar] [CrossRef]
- Rowland, I.R.; Grasso, P. Degradation of N-nitrosamines by intestinal bacteria. Appl. Envir. Microbiol. 1975, 29, 7–12. [Google Scholar] [CrossRef]
- Morotomi, M.; Mutai, M. In vitro binding of potent mutagenic pyrolysates to intestinal bacteria. J. Nat. Cancer Instit. 1986, 11, 195–201. [Google Scholar]
- El-Nezami, H.; Kankaanp, P.; Salminen, S.; Ahokas, J. Ability of dairy strains of lactic acid bacteria to bind a common food carcinogen, aflatoxin B1. Food Chem. Toxicol. 1998, 36, 321–332. [Google Scholar] [CrossRef]
- Praveena, Y.S.N.; Padmini, P.C. Antibacterial activities of mycotoxins from newly isolated filamentous fungi. Int. J. Plant. Anim. Enviromental Sci. 2011, 1, 8–13. [Google Scholar]
- Gill, H.S.; Cross, M.L. Probiotics and immune function. In Nutrition and Immune Function; Calder, P.C., Field, C.J., Gill, H.S., Eds.; CABI Publishing: Wallingford, UK, 2002; pp. 251–272. [Google Scholar]
- Borchers, A.T.; Selmi, C.; Meyers, F.J.; Keen, C.L.; Gershwin, M.E. Probiotics and immunity. J. Gastroenterol. 2009, 44, 26–46. [Google Scholar] [CrossRef] [Green Version]
- Gόmez-Llorente, C.; Munoz, S.; Gil, A. Role of toll-like receptors in the development of immunotolerance mediated by probiotics. Proc. Nutr. Soc. 2010, 69, 381–389. [Google Scholar] [CrossRef]
- Urbanska, A.M.; Paul, A.; Bhahena, J.; Prakash, S. Suppression of tumorigenesis: Modulation of inflammatory cytokines by oral administration of microencapsulated probiotic yogurt formulation. Int. J. Inflam. 2010, 31, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, T.; Asai, Y.; Tamai, R.; Makimura, Y.; Sakamoto, H.; Hashikawa, S.; Yasuda, K. Natural killer cell activities of synbiotic Lactobacillus casei ssp. casei in conjunction with dextran. Clin. Exp. Immunol. 2006, 143, 103–109. [Google Scholar] [CrossRef]
- Sobhani, I.; Tap, J.; Roudot-Thoraval, F.; Roperch, J.P.; Letulle, S.; Langella, P.; Takeda, K.; Suzuki, T.; Shimada, S.I.; Shida, K.; et al. Interleukin-12 is involved in the enhancement of human natural killer cell activity by Lactobacillus casei shirota. Clin. Exp. Immunol. 2006, 146, 109–115. [Google Scholar]
- Madsen, K.L. Enhancement of epithelial barrier function by probiotics. J. Epithel. Biol. Pharmacol. 2012, 5, 55–59. [Google Scholar] [CrossRef] [Green Version]
- Ohigashi, S.; Sudo, K.; Kobayashi, D.; Takahashi, O.; Takahashi, T.; Asahara, T. Changes of the intestinal microbiota, short chain fatty acids, and fecal pH in patients with colorectal cancer. Dig. Dis. Sci. 2013, 58, 1717–1726. [Google Scholar] [CrossRef]
- Crawford, N.; Brooke, B.N. The pH and buffering power of human bile. Lancet 1955, 268, 1096–1097. [Google Scholar] [CrossRef]
- Shimizu, K.; Ogura, H.; Goto, M.; Asahara, T.; Nomoto, K.; Morotomi, M.; Yoshiya, K.; Matsushima, A.; Sumi, Y.; Kuwagata, Y. Altered gut flora and environment in patients with severe SIRS. J. Trauma 2006, 60, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, H.; Yajima, T. Correlation between water-holding capacity of different types of cellulose in vitro and gastrointestinal retention time in vivo of rats. J. Sci. Food Agric. 1992, 60, 139–146. [Google Scholar] [CrossRef]
- Caballero-Franco, C.; Keller, K.; De Simone, C.; Chadee, K. The VSL#3 probiotic formula induces mucin gene expression and secretion in colonic epithelial cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, 315–322. [Google Scholar]
- Karczewski, J.; Troost, F.J.; Konings, I.; Dekker, J.; Kleerebezem, M.; Brummer, R.J.M. Regulation of human epithelial tight junction proteins by Lactobacillus plantarum in vivo and protective effects on the epithelial barrier. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 289, 851–859. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Potential mechanisms of probiotics action in the prevention of colorectal cancer development. Symbols: ↓ decrease, ↑ increase.
Figure 1.
Potential mechanisms of probiotics action in the prevention of colorectal cancer development. Symbols: ↓ decrease, ↑ increase.

Figure 2.
The role of short-chain fatty acids (SCFAs) in regulation of gut and systemic immunity [99,100]. SCFAs are produced by intestinal microbiome in the fermentation of nondigestible carbohydrates (NDCs), undigested food fiber, or resistant starch (RS). SCFAs regulate intestinal barrier function by inducing intestinal epithelial cell secretion of interleukin-18 (IL-18), mucin (MUC2), antimicrobial peptides, and upregulating the expression of tight junction. Moreover, SCFAs regulate the T cell function through G-protein-coupled receptors (GPR41, GPR43, GPR109A), Olfr78 receptor signaling, and through inhibition of histone deacetylase (HDAC) which affects inhibition of nuclear factors (nuclear factor-κB; NF-κB). SCFAs play an important role in inducing neutrophils migration to inflammatory site and enhancing their phagocytosis. Moreover, SCFAs inhibit intestinal macrophage production of proinflammatory cytokines (e.g., IL-6, IL-8, IL-1β and TNFα), and possibly induce intestinal IgA production of B cells. SCFAs affect the differentiation of regulatory T (Treg) cells and the production of interleukin-10 (IL-10). The direct impact of SCFA as well as SCFA regulation of dendritic cells (DCs) are mediated in the differentiation of T cells. SCFAs regulate the generation of Treg, Th1, and Th17 in different cytokine environment. SCFAs promote apoptosis and suppress proliferation of tumor cells resulting in inhibiting the carcinogenesis. Abbreviations: Aldh1A2—aldehyde dehydrogenase 1A2. Symbols:→ activation; →inhibition.
Figure 2.
The role of short-chain fatty acids (SCFAs) in regulation of gut and systemic immunity [99,100]. SCFAs are produced by intestinal microbiome in the fermentation of nondigestible carbohydrates (NDCs), undigested food fiber, or resistant starch (RS). SCFAs regulate intestinal barrier function by inducing intestinal epithelial cell secretion of interleukin-18 (IL-18), mucin (MUC2), antimicrobial peptides, and upregulating the expression of tight junction. Moreover, SCFAs regulate the T cell function through G-protein-coupled receptors (GPR41, GPR43, GPR109A), Olfr78 receptor signaling, and through inhibition of histone deacetylase (HDAC) which affects inhibition of nuclear factors (nuclear factor-κB; NF-κB). SCFAs play an important role in inducing neutrophils migration to inflammatory site and enhancing their phagocytosis. Moreover, SCFAs inhibit intestinal macrophage production of proinflammatory cytokines (e.g., IL-6, IL-8, IL-1β and TNFα), and possibly induce intestinal IgA production of B cells. SCFAs affect the differentiation of regulatory T (Treg) cells and the production of interleukin-10 (IL-10). The direct impact of SCFA as well as SCFA regulation of dendritic cells (DCs) are mediated in the differentiation of T cells. SCFAs regulate the generation of Treg, Th1, and Th17 in different cytokine environment. SCFAs promote apoptosis and suppress proliferation of tumor cells resulting in inhibiting the carcinogenesis. Abbreviations: Aldh1A2—aldehyde dehydrogenase 1A2. Symbols:→ activation; →inhibition.

Figure 3.
Potential mechanisms of action of lactic acid bacteria in pathways of apoptosis [50,107]. The extrinsic pathway is initiated by FAS ligand-induced activation of death receptors on the cell membrane (e.g., TNF—tumor necrosis factor). The intrinsic pathway is induced radiotherapy and chemotherapy. The mitochondrial localization and activation of Bax and Bak are required in the intrinsic pathway. However, it can be prevented by pharmacologic inhibitors or anti-apoptotic Bcl-2 family proteins. Lactic acid bacteria can enhance the apoptosis induction through 5-fluorouracile (5-FU) that can induce the activation of autophagic cell death promoted by the induction of Beclin-1 and GRP78 or Bcl-2 and Bak. Moreover, LAB may act to prevent cancer via downregulating NF-κB-dependent gene products which regulate cell survival (Bcl-2, Bcl-xl) and proliferation (Cox-2, cyclin D1). Symbols: →activation; →inhibition.
Figure 3.
Potential mechanisms of action of lactic acid bacteria in pathways of apoptosis [50,107]. The extrinsic pathway is initiated by FAS ligand-induced activation of death receptors on the cell membrane (e.g., TNF—tumor necrosis factor). The intrinsic pathway is induced radiotherapy and chemotherapy. The mitochondrial localization and activation of Bax and Bak are required in the intrinsic pathway. However, it can be prevented by pharmacologic inhibitors or anti-apoptotic Bcl-2 family proteins. Lactic acid bacteria can enhance the apoptosis induction through 5-fluorouracile (5-FU) that can induce the activation of autophagic cell death promoted by the induction of Beclin-1 and GRP78 or Bcl-2 and Bak. Moreover, LAB may act to prevent cancer via downregulating NF-κB-dependent gene products which regulate cell survival (Bcl-2, Bcl-xl) and proliferation (Cox-2, cyclin D1). Symbols: →activation; →inhibition.


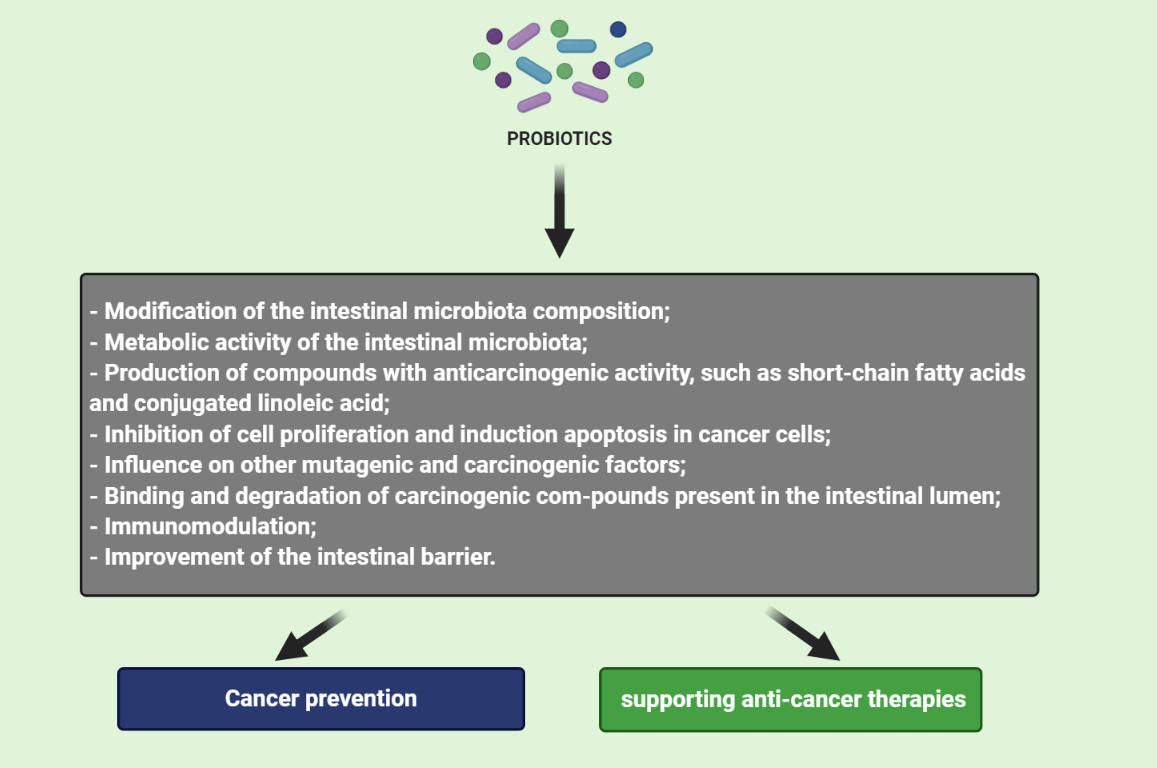{kind=link}
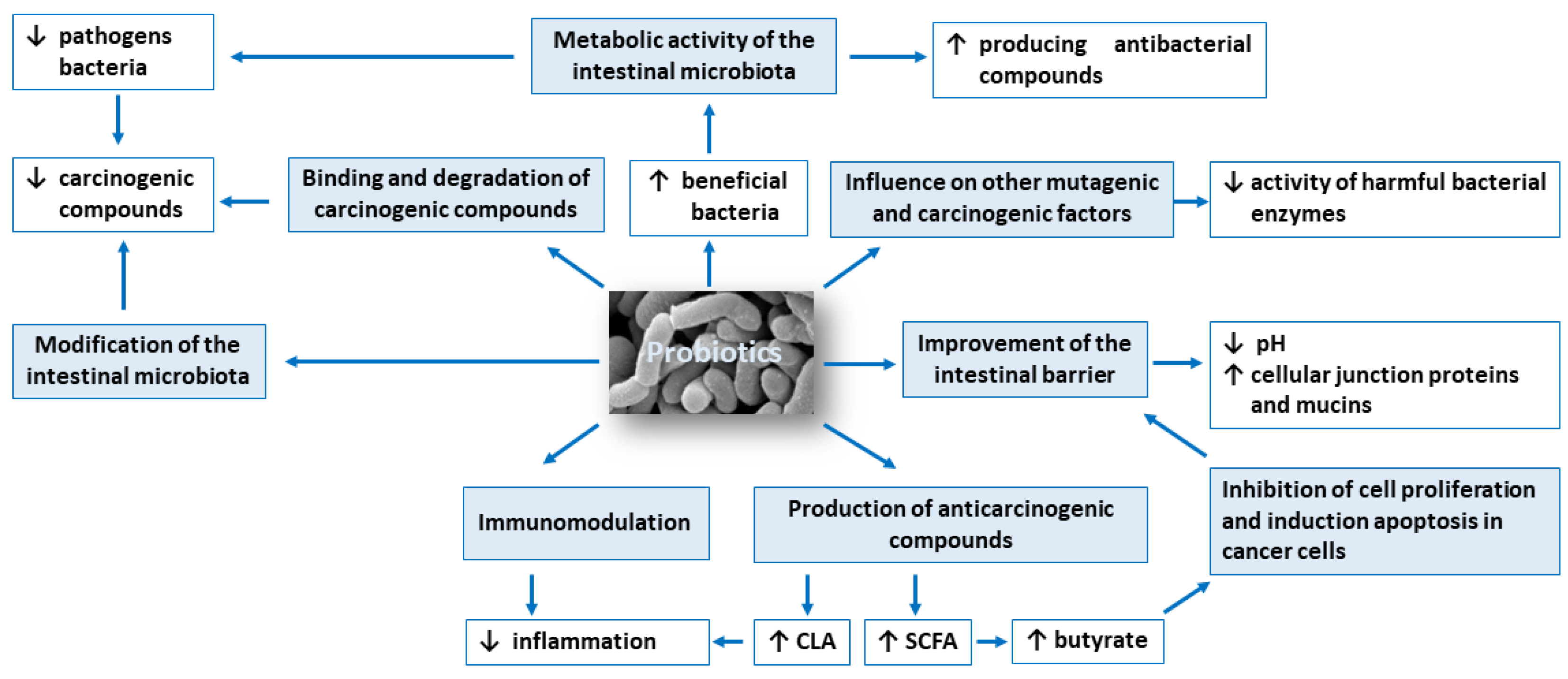{kind=link}
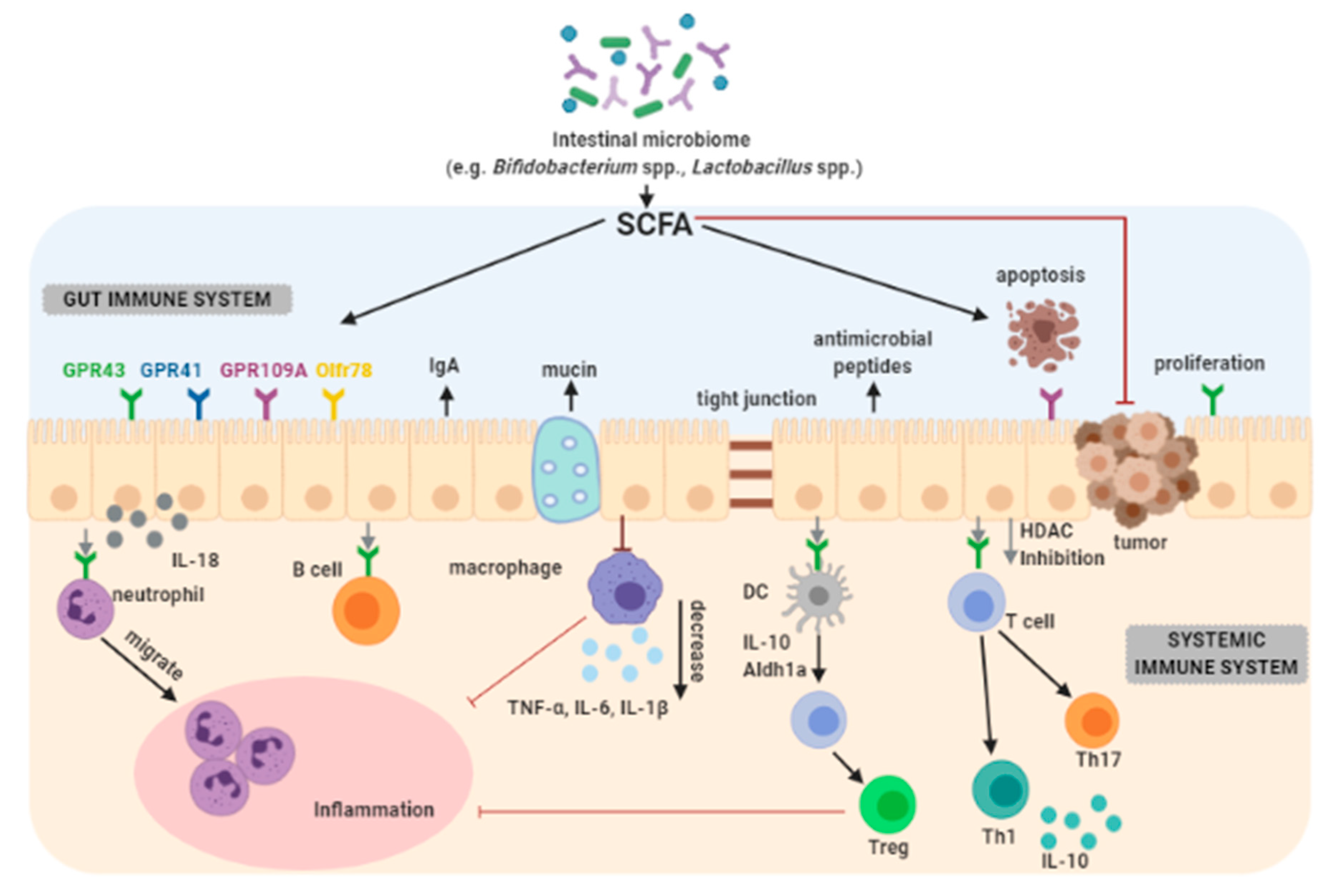{kind=link}
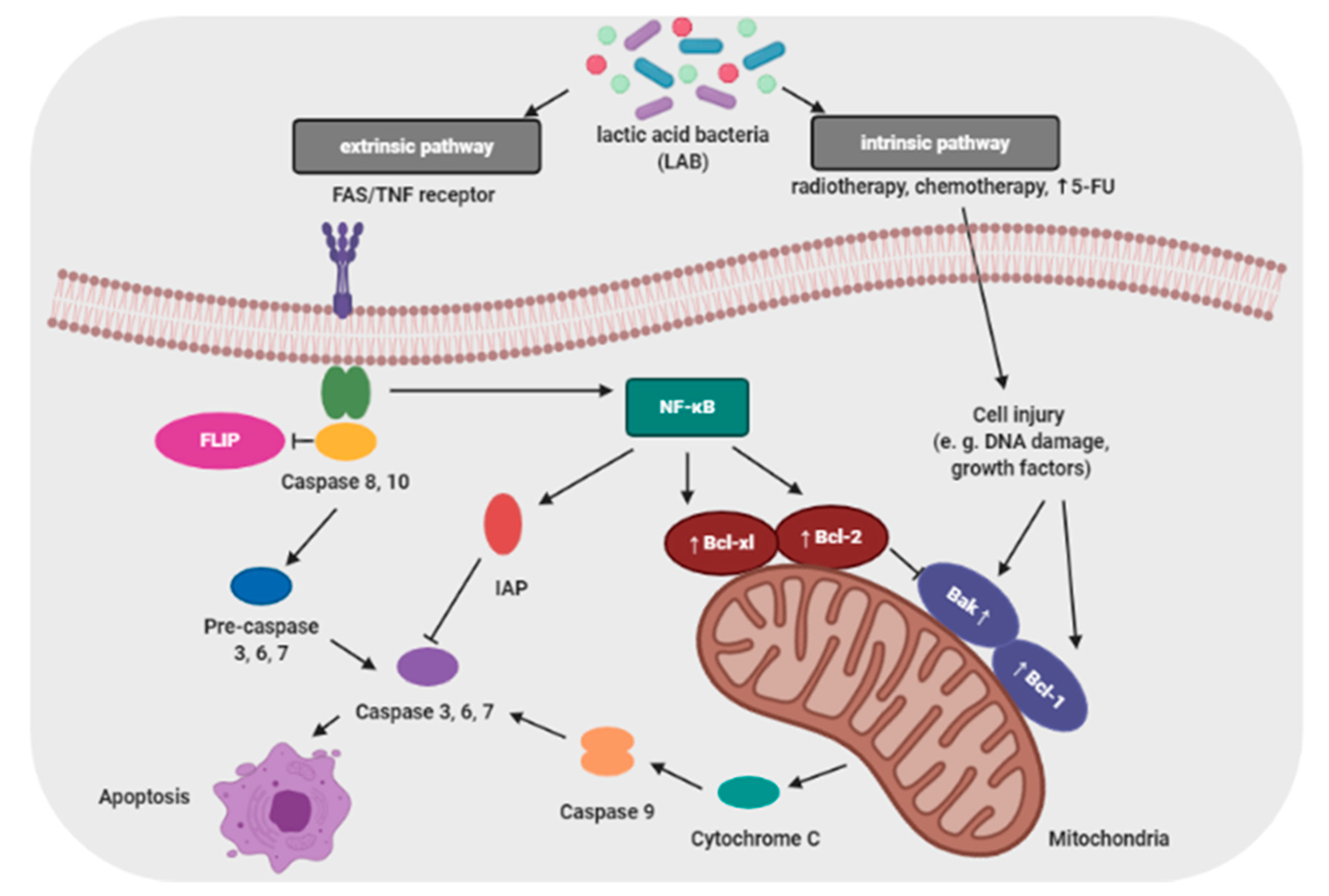{kind=link}
Table 1.
Effects of probiotic microorganisms on cancer cells in in vitro studies.
| Reference | Probiotic Strain | Cell Line | Effect |
|---|---|---|---|
| [20] | Lactobacillus pentosus B281 Lactobacillus plantarum B282 | Caco-2 and HT-29 | ↓ Cell proliferation, Cell cycle arrest (G1) |
| [21] | Lactobacillus casei ATCC 393 | HT29 and CT26 | Induction of apoptosis |
| [22] | Lactococcus lactis NK34 | HT-29, LoVo, AGS | >80% ↓ Cell proliferation |
| [23] | Bacillus polyfermenticus KU3 | LoVo, HT-29, AGS | >90% ↓ Cell proliferation |
| [24] | Clostridium butyricum ATCC Bacillus subtilis ATCC 9398 | HCT116, SW1116, Caco-2 | ↓ Cell proliferation |
| [15] | Lactobacillus plantarum A7 Lactobacillus rhamnosus GG | Caco-2, HT-29 | ↓ Cell proliferation |
| [25] | Lactobacillus kefiri P-IF | MDR | Induction of apoptosis |
| [26] | Pediococcus pentosaceus FP3 Lactobacillus salivarius FP25/FP35 Enterococcus faecium FP51 | Caco-2 | ↓ Cell proliferation, Activation of apoptosis |
| [27] | Propionibacterium freudenreichii ITG P9 | HGT-1 | Induction of apoptosis |
| [13] | Lactobacillus paracasei IMPC2.1 Lactobacillus rhamnosus GG | DLD-1, HGC-27 | ↓ Cell proliferation, Induction of apoptosis |
| [12] | Lactobacillus rhamnosus GG Bifidobacterium lactis Bb12 | HT-29 | Induction of apoptosis |
| [17] | Lactobacillus acidophilus CL1285 Lactobacillus casei LBC80R (in the presence of 5-FU) | LS513 | 40% ↑ apoptosis |
| [28] | Lactobacillus acidophilus 606 | HT-29 | Inhibited proliferation of tumor cells |
| [19] | Bacillus polyfermenticus | NMC460 | ↓ Cell colony formation in cancer cells (N/E on normal colonocytes) |
| [29] | Enterococcus faecalis CECT7121 | LBC | Inhibited proliferation of tumor cells, Induction od apoptosis |
| [11] | Lactobacillus rhamnosus GG Bifidobacterium lactis Bb12 | Caco-2 | ↑ Apoptosis |
| [30] | Enterococcus faecium RM11 Lactobacillus fermentum RM28 | Caco-2 | Cell proliferation: ↓ 21% ↓ 23% |
| [31] | Lactobacillus reuteri ATCC PTA 6475 | KBM-5 | ↑ Apoptosis |
| [16] | Lactobacillus rhamnosus GG | Caco-2 | ↓ level of IL–8 |
| [32] | Bifidobacterium adolescentis SPM0212 | Caco-2, HT-29, SW480 | ↓ Cell proliferation |
| [14] | Lactobacillus rhamnosus GG | HGC-27 | ↓ Cell proliferation Induction of apoptosis |
| [18] | Lactobacillus acidophilus SNUL Lactobacillus casei YIT9029 Bifidobacterium longum HY8001 | SNUC2A, SNU1, NIH/3T3 and Jurkat cell | Suppressed proliferation of tumor cells |
| [33] | Propionibacterium acidopropionici CNRZ80 | HT-29 | ↓ Cell proliferation Induction of apoptosis |
Abbreviations: ↓ Decrease; ↑ increase; N/E no effect; human colonic cancer cells: Caco-2, HT-29, SW1116, HCT116, SW480, DLD-1, LoVo; human colonic epithelial cells: NMC460; human colorectal carcinoma cells: LS513, SNUC2A; human gastric adenocarcinoma cells: AGS; human gastric carcinoma cells: SNU1; human myeloid leukemia-derived cells: MDR, KMB-5; human T lymphocyte cells: Jurkat cel; mus musculus colon carcinoma cells: CT26; HGC-27, HGT-1; mus musculus embryonic fibroblast: NIH/3T3; spontaneous murine T-cell lymphoma cells: LBC, 5FU5-Fluorouracil.
Table 2.
Effects of probiotics on tumor-bearing or tumor-induced in animal models in vivo.
| Reference | Probiotic Strain | Model | Induction | Time of Treatment | Effect |
|---|---|---|---|---|---|
| [36] | Lactobacillus casei BL23 | C57BL/6 mice | DMH | 10 weeks | ↓ TI |
| [34] | Pediococcus pentosaceus GS4 | Swiss albino mice | AOM | 4 weeks | ↓ TP Induction of apoptosis |
| [37] | Lactobacillus rhamnosus GG CGMCC 1.2134 | SD rats | DMH 10 weeks | 25 weeks | ↓ TI ↓ TV ↓ TM Induction of apoptosis |
| [38] | Lactobacillus acidophilus Bifidobacterium bifidum Bifidobacterium infantum | SD rats | antibiotics DMH | 23 weeks | ↓ TI ↓ TV |
| [39] | Lactobacillus salivarius Ren | F344 rats | DMH 10 weeks | 2 weeks a | ↓ TI |
| [40] | Lactobacillus plantarum (AdF10) Lactobacillus rhamnosus GG | SD rats | DMH 4 weeks | One of strains 12 weeks | ↓ TI ↓ TV ↓ TM |
| [15] | VSL#3 (Probiotics mixture) | C57BL/6 mice | DSS | 2 weeks a | ↓ TI ↓ dysplasia |
| [41] | Lactobacillus plantarum | BALB/c mice | CT26 cells injection | 14 weeks | ↓ TV Induction of necrosis |
| [23] | Lactobacillus plantarum | BALB/c mice | AOM, DSS | Nanosized/ Live bacteria 4 weeks | ↓ TI cell cycle arrest Induction of apoptosis |
| [42] | Lactobacillus rhamnosus GG MTCC #1408 Lactobacillus acidophilus NCDC #1 | SD rats | DMH | 19 weeks a | ↓ TI ↓ TM |
| [43] | Lactobacillus acidophilus KFRI342 | F344 rats | DMH | 10 weeks | ↓ ACF ↓ β-glucuronidase and β-glucosidase activity |
| [44] | Lactobacillus rhamnosus 231 (Lr 231) | rats | MNNG | 5 weeks | ↓ fecal activity of azoreductase and nitroreductase, ↓ GST ↑GSH |
| [45] | VSL#3 (Probiotics mixture) | SD rats | TNBS | 10 weeks | None of the animals developed CRC |
| [46] | Lactobacillus plantarum | Wistar albino rats | DMH | 6 weeks | ↓ the activities of bacterial enzymes, the fecal bile acids concentration ↑ serum TNFα level |
| [19] | Bacillus polyfermenticus | CD-1 mice | DLD-1 cells injection | 20 weeks (injection) | ↓ TI ↓ TV |
| [28] | Bifidobacterium lactis KCTC 5727 | SPF C57BL rat | – | 19 weeks | ↓ TI ↓ TV |
| [47] | Lactobacillus acidophilus, Lactobacillus casei Lactobacillus lactis biovar diacetylactis DRC-1 | Rat | DMH | 40 weeks | ↓ TI ↓TV ↓ TM |
| [48] | Bacillus polyfermenticus | F344 rats | DMH | 6 weeks | 50% ↓ ACF, ↑ antioxidant potential |
a Before and until the end of experiment. Abbreviations: ↓ Decrease; ↑ increase; ACF—aberrant crypt foci; AOM—azoxymethane; CRC—colorectal cancer; DMH—1,2-dimethylhydrazine dihydrochloride; DSS—dextran sulfate sodium; GSH—glutathione; GST—glutathione S–transferase; MNNG—N–Methyl–N’–Nitro–Nitrosoguanidine; SD rat—Sprague–Dawley rat; TI—tumor incidence; TM—tumor multiplicity; TNBS—trinitrobenzene sulfonic acid; TNFα level—serum tumor necrosis factor-alpha; TP—tumor progression; TV—tumor volume.
Table 3.
Examples of probiotic bacteria used in the chemoprevention and the anti-cancer therapy.
| Reference | Type of Cancer | Strain/Strains |
|---|---|---|
| [49] | Colorectal carcinoma | Bifidobacterium lactis Bb12, Lactobacillus rhamnosus GG |
| [50] | Bifidobacterium longum, Lactobacillus acidophilus, Enterococcus faecalis | |
| [51] | Bacillus natto, Lactobacillus acidophilus | |
| [52] | Lactobacillus plantarum CGMMCC No 1258, Lactobacillus acidophilus LA-11, Bifidobacterium longum BL-88 | |
| [53] | Lactobacillus rhamnosus 573 | |
| [54] | Liver cancer | Lactobacillus rhamnosus LC705, Propionibacterium freudenreichii subsp. shermanii |
| [55] | Gastric cancer | Lactobacillus reuteri PTCC 1655 |
| [56] | Lactobacillus kefiri P-IF | |
| [57] | Cervical cancer | Lactobacillus acidophilus, Bifidobacterium bifidum |
Table 4.
Results of clinical studies using probiotic strains in the cancer prevention and the treatment.
Table 4.
Results of clinical studies using probiotic strains in the cancer prevention and the treatment.
| Reference | Probiotic Strain | Subject | Time of Treatment | Effect |
|---|---|---|---|---|
| The Prevention | ||||
| [58] | Yakult containing Lactobacillus casei Shirota (LcS) and isoflavones from soy product | 968 breast cancer patients aged 40 to 55. | 2 years | ↓ the incidence of breast cancer in Japanese women correlated with consumption of LcS and isoflavones since adolescence. |
| [59] | Streptococcus thermophilus and Lactobacillus delbruckii subsp. bulgaricus | 45,241 healthy people (14,178 men, 31,063 women) | 12 years | ↓ the risk of colorectal cancer correlated with increased consumption of yogurt (especially in men). |
| [60] | Lactobacillus gasseri OLL2716 (LG21) | 10 people with colorectal cancer and 20 healthy patients | 12 weeks | the number of bacteria from the genus Lactobacillus, synthesis of isobutyric acid, NK cell activity. ↓ the amount of Clostridium perfringens. |
| [19] | Lactobacillus casei Shirota (LcS) | 54 women with an HPV-positive intra epithelial lesion | 6 months | 60 % ↓ in human papilloma virus (HPV) associated infection and cervical cancer precursors |
| [61] | Lactobacillus rhamnosus LC705, Propionibacterium freudenreichii ssp. shermanii JS | 38 men (between 24 and 55 years old) | 4 weeks | ↓ β-glucosidase and urease activity amount of bacteria of the genus Lactobacillus and Propionibacterium |
| [54] | Lactobacillus rhamnosus LC705, Propionibacterium freudenreichii subsp. shermanii | 90 male students with high aflatoxin level in urine | 5 weeks | 61.5% ↓ a liver cancer biomarker which leads to reduced urinary excretion of aflatoxin B1-N7guanine (AFB-N7-guanine) |
| [62] | Lactobacillus acidophilus L1 | 180 people with bladder cancer (mean age:67 years and 445 population-based controls | 10 weeks | ↓ the risk of bladder cancer correlated with habitual intake of lactic acid bacteria |
| The Treatment | ||||
| [63] | Lactobacillus acidophilus, Lactobacillus plantarum, Bifidobacterium lactis, Saccharomyces boulardii | 164 patients with colorectal cancer undergoing colorectal surgery | 30 days | ↓ the risk of postoperative complications. In the probiotic group, a positive correlation was observed between the expression of the SOCS3 gene and the expression of the TNF gene and circulating IL–6. |
| [64] | Lactobacillus plantarum CGMCC, Lactobacillus acidophilus-11, Bifidobacterium longum-88 | 150 patients with colorectal cancer | 6 days preoperatively and 10 days postoperative | ↓ the serum zonulin concentration, ↓ duration of postoperative pyrexia, ↓ duration of antibiotic therapy, ↓ rate of postoperative infectious complications, Inhibited the p38 mitogen-activated protein kinase signaling pathway |
| [65] | Bifidobacterium longum | 60 patients with colorectal cancer undergoing colon resection | 3 days | the count of bacteria of the genus Bifidobacterium ↓ the count of bacteria of the genus Escherichia |
| [66] | Bifidobacterium breve Yakult | 42 patients during chemotherapy | 6 weeks | ↓ the incidence of fever ↓ the need for intravenous antibiotics using compared to the control group. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Śliżewska, K.; Markowiak-Kopeć, P.; Śliżewska, W. The Role of Probiotics in Cancer Prevention. Cancers 2021, 13, 20. https://doi.org/10.3390/cancers13010020
AMA Style
Śliżewska K, Markowiak-Kopeć P, Śliżewska W. The Role of Probiotics in Cancer Prevention. Cancers. 2021; 13(1):20. https://doi.org/10.3390/cancers13010020
Chicago/Turabian StyleŚliżewska, Katarzyna, Paulina Markowiak-Kopeć, and Weronika Śliżewska. 2021. "The Role of Probiotics in Cancer Prevention" Cancers 13, no. 1: 20. https://doi.org/10.3390/cancers13010020
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.