Influence of a Major Mountainous Landscape Barrier (Mount Cameroon) on the Spread of Metabolic (GSTe2) and Target-Site (Rdl) Resistance Alleles in the African Malaria Vector Anopheles funestus †

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
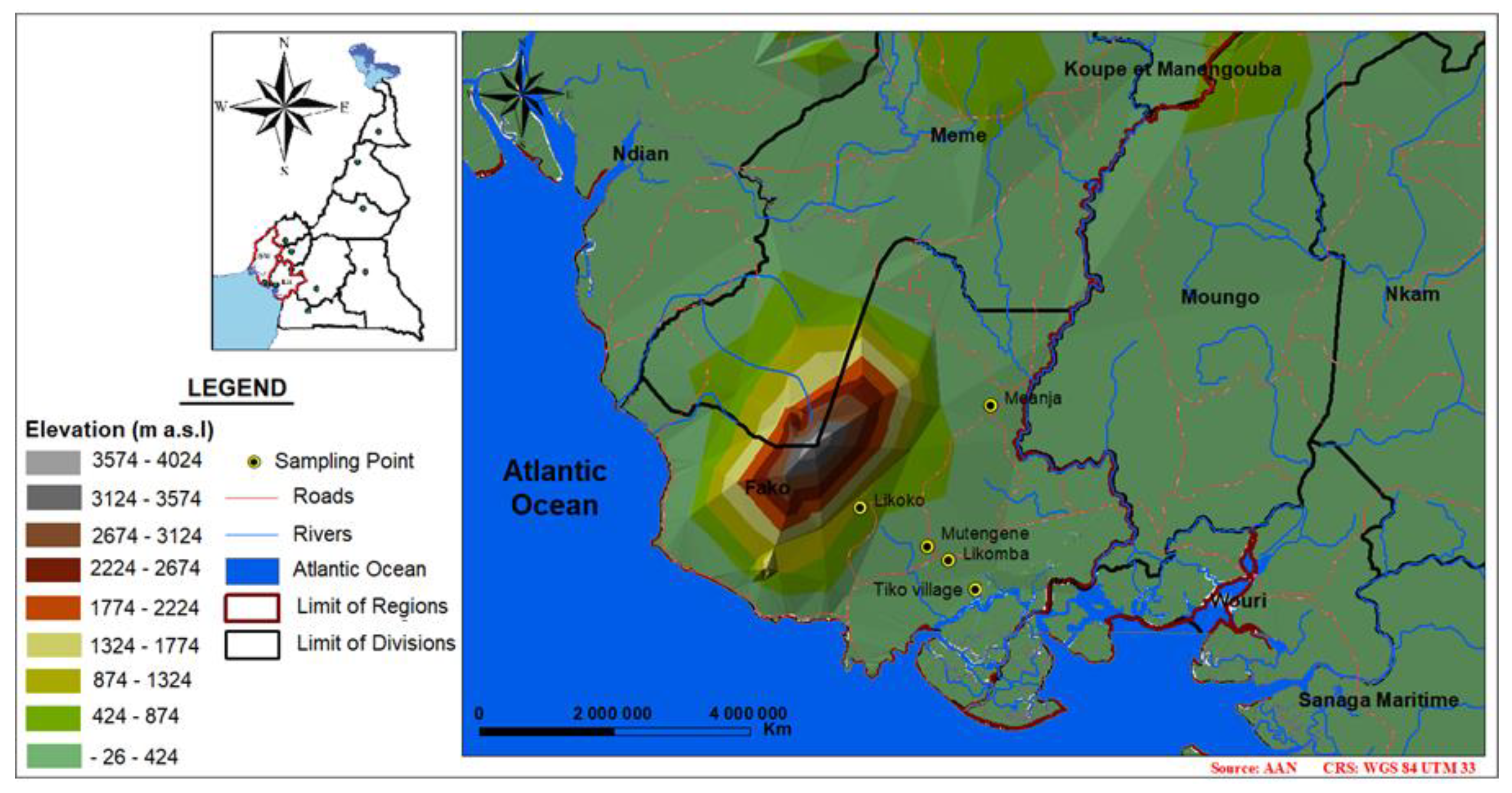
2.1. Study Site and Adult Mosquito Sampling
2.2. Species Identification and Plamodium Infection Rate
2.3. Genotyping of GSTe2 and Rdl Resistance Markers
2.4. Polymorphism Analysis of Resistance Genes
2.4.1. Genetic Variability of An. funestus s.s. across the Mount Cameroon Region Based on GSTe2 and Rdl Full Gene Sequencing
2.4.2. Phylogenetic Trees of Haplotypes
3. Results
3.1. Mosquito Species Composition
3.2. Parity and Plasmodium Infection Rates
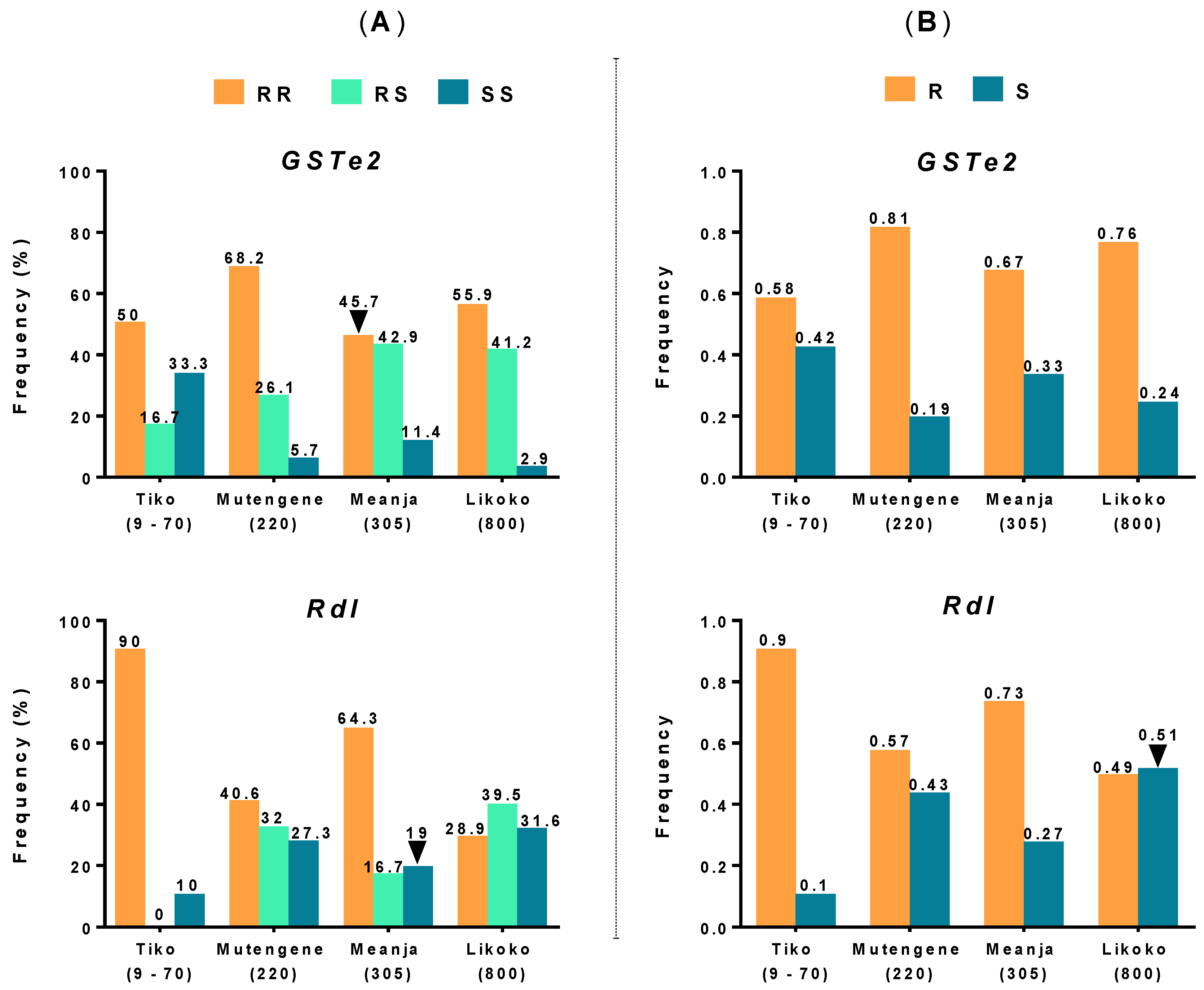
3.3. Detection of Mutations Associated with DDT and Dieldrin Resistance in An. funestus s.s. Mosquitoes
3.4. Analysis of the Polymorphism of GSTe2 and Rdl Genes across Mount Cameroon An. funestus s.s. Populations
3.4.1. Sequence Analysis of Full Length GSTe2 and Rdl Genes
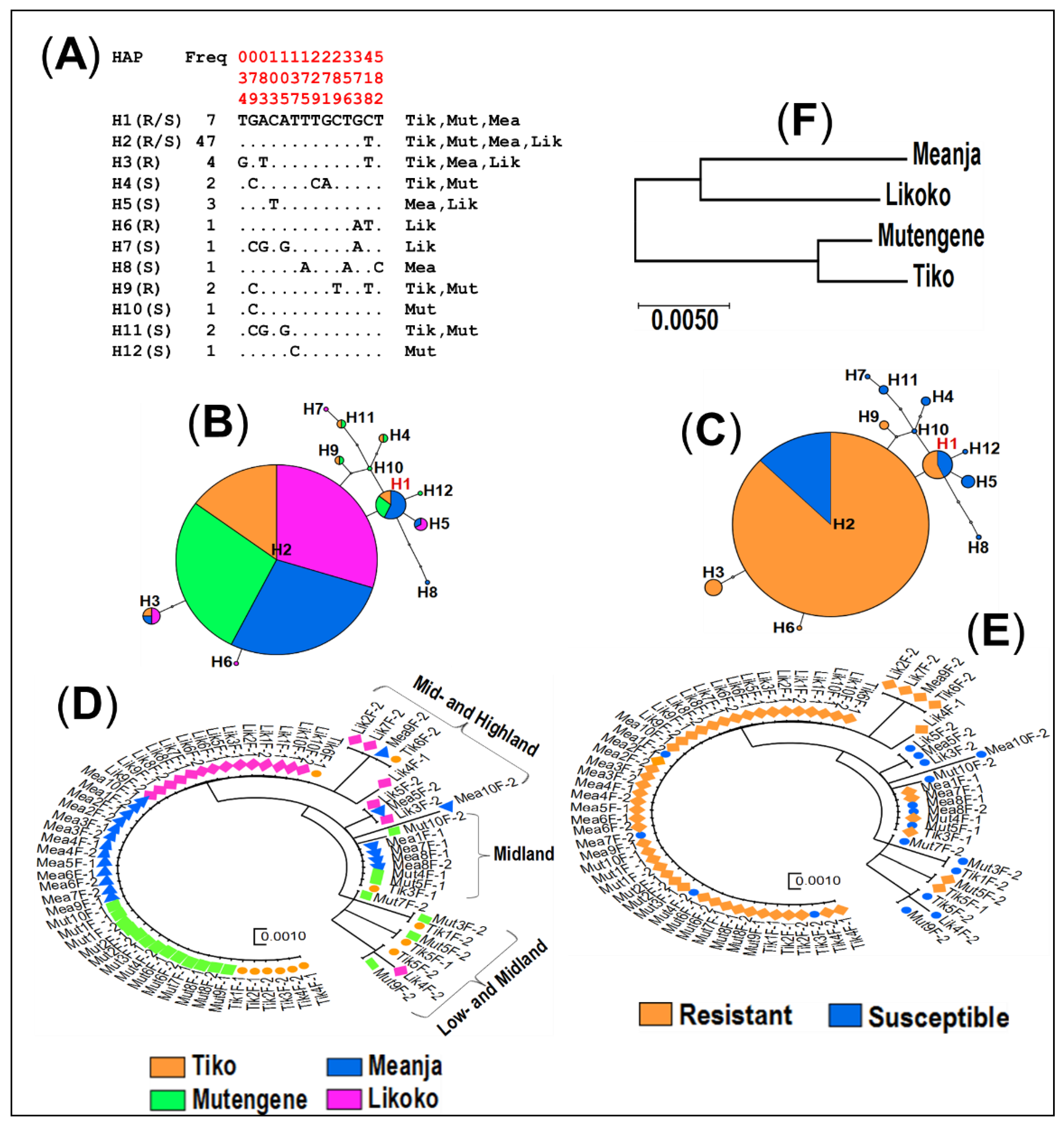
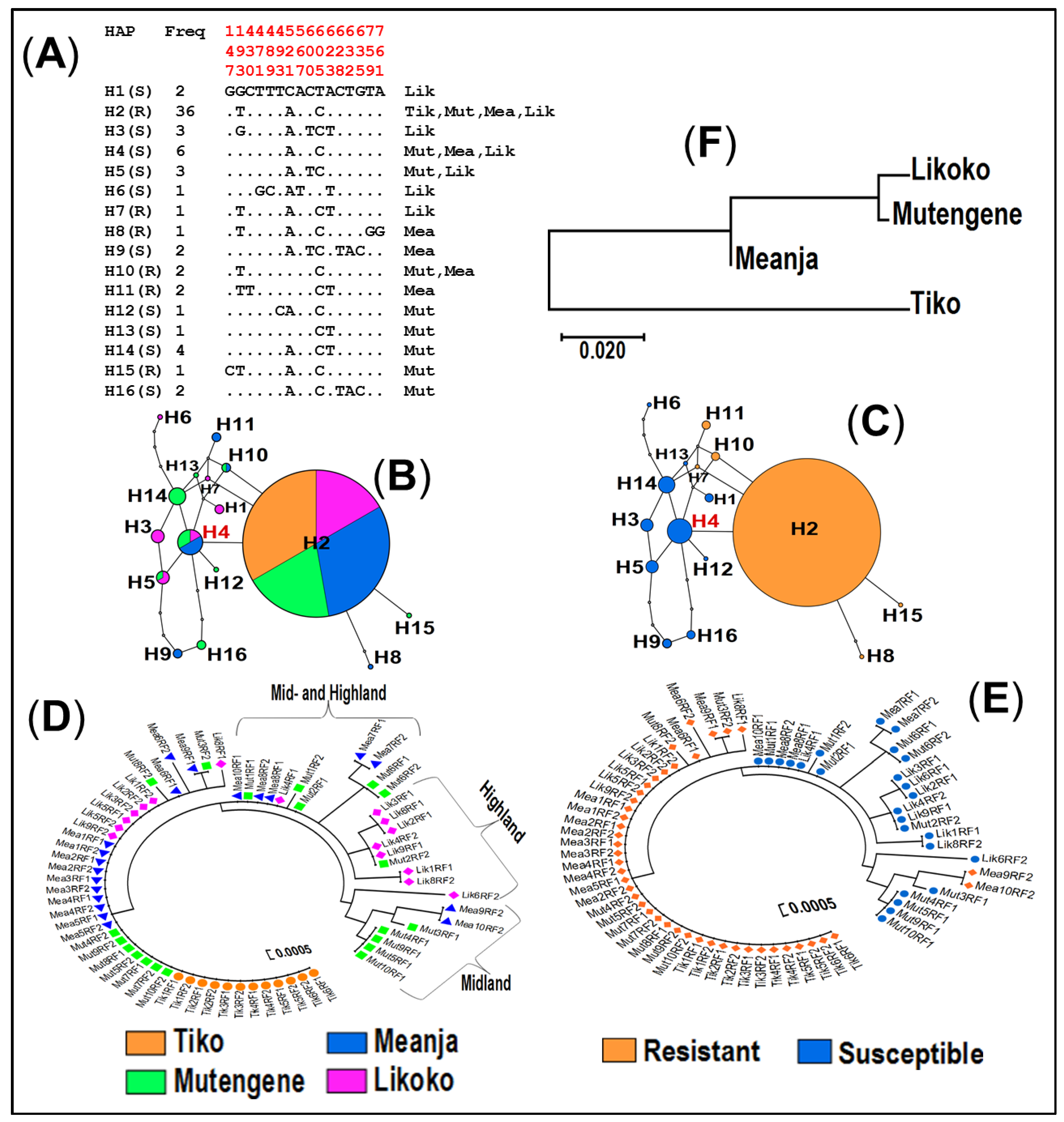
3.4.2. Haplotype Distribution of the GSTe2 Gene
3.4.3. Haplotype Distribution of the GABA-Receptor Gene across Mount Cameroon Populations of An. funestus s.s.
3.5. Population Structure at GSTe2 and Rdl Mutations in An. funestus s.s. across Mount Cameroon
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- WHO. Test Procedure for Insectcides Resistance Monitoring in Malaria Vector Mosquitoes, 2nd ed.; World Health Organisation: Geneva, Switzerland, 2016; pp. 6–8. Available online: http://apps.who.int/iris/bitstream/10665/80139/1/9789241505154_eng.pdf (accessed on 10 July 2020).
- Zaim, M.; Aitio, A.; Nakashima, N. Safety of pyrethroid-treated mosquito nets. Med. Vet. Entomol. 2000, 14, 1–5. [Google Scholar] [CrossRef] [PubMed]
- WHO. Long-Lasting Insecticidal Nets for Malaria Prevention—A Manual for Malaria Programme Managers, Trial ed.; World Health Organisation: Geneva, Switzerland, 2007; pp. 5–9. Available online: http://www.who.int/malaria/publications/atoz/insecticidal_nets_malaria/en/ (accessed on 25 June 2020).
- WHO. WHO Recommended Long-Lasting Insecticidal Mosquito Nets; World Health Organisation: Geneva, Switzerland, 2009. [Google Scholar]
- Pluess, B.; Tanser, F.C.; Lengeler, C.; Sharp, B.L. Indoor residual spraying for preventing malaria. Cochrane Database Syst. Rev. 2010, 2010. [Google Scholar] [CrossRef]
- Bhatt, S.; Weiss, D.; Cameron, E.; Bisanzio, D.; Mappin, B.; Dalrymple, U.; Battle, K.; Moyes, C.; Henry, A.; Eckhoff, P.; et al. The effect of malaria control on Plasmodium falciparum in Africa between 2000 and 2015. Nature 2015, 526, 207–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemingway, J.; Ranson, H. Insecticide resistance in insect vectors of human disease. Annu. Rev. Entomol. 2000, 45, 371–391. [Google Scholar] [CrossRef] [PubMed]
- Hemingway, J.; Field, L.; Vontas, J. An overview of insecticide resistance. Science 2002, 298, 96–97. [Google Scholar] [CrossRef]
- Ranson, H.; N’Guessan, R.; Lines, J.; Moiroux, N.; Nkuni, Z.; Corbel, V. Pyrethroid resistance in African anopheline mosquitoes: What are the implications for malaria control? Trends Parasitol. 2010, 27, 91–98. [Google Scholar] [CrossRef]
- WHO. World Malaria Report 2019; World Health Organisation: Geneva, Switzerland, 2019. [Google Scholar]
- Sinka, M.; Bangs, M.; Manguin, S.; Coetzee, M.; Mbogo, C.; Hemingway, J.; Patil, A.; Temperley, W.; Gething, P.; Kabaria, C.; et al. The dominant Anopheles vectors of human malaria in Africa, Europe and the Middle East: Occurrence data, distribution maps and bionomic précis. Parasit. Vectors 2010, 3, 117. [Google Scholar] [CrossRef] [Green Version]
- Coetzee, M.; Hunt, R.; Wilkerson, R.; Della Torre, A.; Coulibaly, M.; Besansky, N. Anopheles coluzzii and Anopheles amharicus, new members of the Anopheles gambiae complex. Zootaxa 2013, 3619, 246–274. [Google Scholar] [CrossRef] [Green Version]
- Wanji, S.; Tanke, T.; Atanga, S.N.; Ajonina, C.; Tendongfor, N.; Fontenille, D. Anopheles species of the mount Cameroon region: Biting habits, feeding behaviour and entomological inoculation rates. Trop. Med. Int. Health 2003, 8, 643–649. [Google Scholar] [CrossRef] [Green Version]
- Tchuinkam, T.; Simard, F.; Lélé-Defo, E.; Téné-Fossog, B.; Tateng-Ngouateu, A.; Antonio-Nkondjio, C.; Mpoame, M.; Toto, J.; Njiné, T.; Fontenille, D.; et al. Bionomics of Anopheline species and malaria transmission dynamics along an altitudinal transect in Western Cameroon. BMC Infect. Dis. 2010, 10, 119. [Google Scholar] [CrossRef] [Green Version]
- Wondji, C.; Dabire, R.; Tukur, Z.; Irving, H.; Djouaka, R.; Morgan, J. Identification and distribution of a GABA receptor mutation conferring dieldrin the malaria vector Anopheles funestus in Africa. Insect Biochem. Mol. Biol. 2011, 41, 484–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menze, B.; Riveron, J.; Ibrahim, S.; Irving, H.; Antonio-Nkondjio, C.; Awono-Ambene, P.; Wondji, C. Multiple insecticide resistance in the malaria vector Anopheles funestus from northern Cameroon is mediated by metabolic resistance alongside potential target site insensitivity. PLoS ONE 2016, 11, e0163261. [Google Scholar] [CrossRef] [PubMed]
- Irving, H.; Wondji, C. Investigating knockdown resistance (kdr) mechanism against pyrethroids/DDT in the malaria vector Anopheles funestus across Africa. BMC Genet. 2017, 18, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ndo, C.; Kopya, E.; Donbou, M.A.; Njiokou, F.; Awono-Ambene, P.; Wondji, C. Elevated Plasmodium infection rates and high pyrethroid resistance in major malaria vectors in a forested area of Cameroon highlight challenges of malaria control. Parasit. Vectors 2018, 11, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ndo, C.; Kopya, E.; Irving, H.; Wondji, C. Exploring the impact of glutathione S-transferase (GST)-based metabolic resistance to insecticide on vector competence of Anopheles funestus for Plasmodium falciparum version 2; peer review: 2 approved, 1 approved with reservations]. Wellcome Open Res. 2019, 4. [Google Scholar] [CrossRef]
- Riveron, J.; Yunta, C.; Ibrahim, S.; Djouaka, R.; Irving, H.; Menze, B.; Ismail, H.; Hemingway, J.; Ranson, H.; Albert, A.; et al. A single mutation in the GSTe2 gene allows tracking of metabolically based insecticide resistance in a major malaria vector. Genome Biol. 2014, 12, R27. [Google Scholar] [CrossRef] [Green Version]
- Lui, N. Insecticides resistance in mosquitoes: Impact, mechanisms, and research directions. Annu. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef]
- Riveron, J.; Irving, H.; Ndula, M.; Barnes, K.; Ibrahim, S.; Paine, M.; Wondji, C. Directionally selected cytochrome P450 alleles are driving the spread of pyrethroid resistance in the major malaria vector Anopheles funestus. Proc. Natl. Acad. Sci. USA 2013, 110, 252–257. [Google Scholar] [CrossRef] [Green Version]
- Riveron, J.; Ibrahim, S.; Chanda, E.; Mzilahowa, T.; Cuamba, N.; Irving, H.; Barnes, K.; Ndula, M.; Wondji, C. The highly polymorphic CYP6M7 cytochrome P450 gene partners with the directionally selected CYP6P9a and CYP6P9b genes to expand the pyrethroid resistance front in the malaria vector Anopheles funestus in Africa. BMC Genom. 2014, 15, 817. [Google Scholar] [CrossRef] [Green Version]
- Caplat, P.; Edelaar, P.; Dudaniec, R.; Green, A.; Okamura, B.; Cote, J.; Ekroos, J.; Jonsson, P.; Löndahl, J.; Tesson, S.; et al. Looking beyond the mountain: Dispersal barriers in a changing world. Front. Ecol. Environ. 2016, 14, 261–268. [Google Scholar] [CrossRef]
- Dhimal, M.; Ahrens, B.; Kuch, U. Climate change and spatiotemporal distributions of vector-borne diseases in Nepal—a systematic synthesis of literature. PLoS ONE 2015, 10, e0129869. [Google Scholar] [CrossRef] [PubMed]
- Zittra, C.; Vitecek, S.; Obwaller, A.; Rossiter, H.; Eigner, B.; Zechmeister, T.; Waringer, J.; Fuehrer, H.-P. Landscape structure affects distribution of potential disease vectors (Diptera: Culicidae). Parasit. Vectors 2017, 10, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haran, J.; Roques, A.; Bernard, A.; Robinet, C.; Roux, G. Altitudinal barrier to the spread of an invasive species: Could the Pyrenean Chain slow the natural spread of the pinewood nematode? PLoS ONE 2015, 10, e0134126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weedall, G.; Mugenzi, L.; Menze, B.; Tchouakui, M.; Ibrahim, S.; Amvongo-Adjia, N.; Irving, H.; Wondji, M.; Tchoupo, M.; Djouaka, R.; et al. A cytochrome P450 allele confers pyrethroid resistance on a major African malaria vector, reducing insecticide-treated bednet efficacy. Sci. Transl. Med. 2019, 11, eaat7386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mugenzi, L.M.J.; Menze, B.D.; Tchouakui, M.; Wondji, M.J.; Irving, H.; Tchoupo, M.; Hearn, J.; Weedall, G.D.; Riveron, J.M.; Wondji, C.S. Cis-regulatory CYP6P9b P450 variants associated with loss of insecticide-treated bed net efficacy against Anopheles funestus. Nat. Commun. 2019, 10, 4652. [Google Scholar] [CrossRef] [Green Version]
- Barnes, K.G.; Irving, H.; Chiumia, M.; Mzilahowa, T.; Coleman, M.; Hemingway, J.; Wondji, C.S. Restriction to gene flow is associated with changes in the molecular basis of pyrethroid resistance in the malaria vector Anopheles funestus. Proc. Natl. Acad. Sci. USA 2017, 114, 286–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weedall, G.D.; Riveron, J.M.; Hearn, J.; Irving, H.; Kamdem, C.; Fouet, C.; White, B.J.; Wondji, C.S. An Africa-wide genomic evolution of insecticide resistance in the malaria vector Anopheles funestus involves selective sweeps, copy number variations, gene conversion and transposons. PLoS Genet. 2020, 16, e1008822. [Google Scholar] [CrossRef] [PubMed]
- Boussougou-Sambe, S.T.; Eyisap, W.E.; Tasse, G.C.T.; Mandeng, S.E.; Mbakop, L.R.; Enyong, P.; Etang, J.; Fokam, E.B.; Awono-Ambene, P.H. Insecticide susceptibility status of Anopheles gambiae (s.l.) in South-West Cameroon four years after long-lasting insecticidal net mass distribution. Parasit. Vectors 2018, 11, 391. [Google Scholar] [CrossRef] [PubMed]
- Awono, A.; Akombi, A.; Owona, H.; Barreau, E. REDD+ around Mount Cameroon, southwest region of Cameroon. In REDD+ on the Ground: A Case Book of Subnational Initiatives across the Globe; Sills, E., Stibniati, S., Atmadja, S., de Sassi, C., Duchelle, A., Kweka, D., Resosudarmo, I., Sunderlin, W., Eds.; CIFOR: Bogor, Indonesia, 2014; pp. 188–202. [Google Scholar]
- Wanji, S.; Enyong, P.; Tendongfor, N.; Ndamukong, J.; Mafo, F.; Kengne-Ouafo, A. Biodiversity of Anopheles Mosquito Species across the Mount Cameroon Region; Research Foundation for Tropical Diseases and Environment: Buea, Cameroon, 2006. [Google Scholar]
- Suchel, J.-B. Les Climats du Cameroun. Ph.D. Thesis, Université de Saint-Etienne, Saint-Etienne, France, 1988. [Google Scholar]
- Wanji, S.; Kengne-Ouafo, A.J.; Eyong, E.E.J.; Kimbi, H.K.; Tendongfor, N.; Ndamukong-Nyanga, J.L.; Nana-Djeunga, H.C.; Bourguinat, C.; Sofeu-Feugaing, D.D.; Charvet, C.L. Genetic diversity of Plasmodium falciparum merozoite surface protein-1 block 2 in sites of contrasting altitudes and malaria endemicities in the Mount Cameroon region. Am. J. Trop. Med. Hyg. 2012, 86, 764–774. [Google Scholar] [CrossRef] [PubMed]
- Anchang-Kimbi, J.K.; Nkweti, V.N.; Ntonifor, H.N.; Apinjoh, T.O.; Tata, R.B.; Chi, H.F.; Achidi, E.A. Plasmodium falciparum parasitaemia and malaria among pregnant women at first clinic visit in the mount Cameroon Area. BMC Infect. Dis. 2015, 15, 439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bigoga, J.; Manga, L.; Titanji, V.; Coetzee, M.; Leke, R. Malaria vectors and transmission dynamics in coastal south-western Cameroon. Malar. J. 2007, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amvongo-Adjia, N.; Wirsiy, E.L.; Riveron, J.M.; Chounna Ndongmo, W.P.; Enyong, P.A.; Njiokou, F.; Wondji, C.S.; Wanji, S. Bionomics and vectorial role of anophelines in wetlands along the volcanic chain of Cameroon. Parasit. Vectors 2018, 11, 471. [Google Scholar] [CrossRef] [PubMed]
- Gillies, M.; de Meillon, B. The Anophelinae of Africa South of the Sahara (Ethiopian Zoogeographical Region); South African Institute for Medical Research: Johannesburg, South Africa, 1968; Volume 2. [Google Scholar]
- Detinova, T. Age Grouping Methods in Diptera of Medical Importance, with Special Reference to Some Vectors of Malaria; World Health Organization Monographs Series; WHO: Geneva, Switzerland, 1962; Volume 47. [Google Scholar]
- Cornel, A.; Collins, F. PCR of the ribosomal DNA intergenic spacer regions as a method for identifying mosquitoes in the Anopheles gambiae complex. In Species Diagnostics Protocols; Clapp, J., Ed.; Humana Press: Totowa, NJ, USA, 1996; Volume 50, pp. 321–332. [Google Scholar] [CrossRef]
- Scott, J.A.; Brogdon, W.G.; Collins, F.H. Identification of single specimens of the Anopheles gambiae complex by the polymerase chain reaction. Am. J. Trop. Med. Hyg. 1993, 49, 520–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanello, C.; Santolamazza, F.; della Torre, A. Simultaneous identification of species and molecular forms of Anopheles gambiae complex by PCR-RFLP. Med. Vet. Entomol. 2002, 16, 461–464. [Google Scholar] [CrossRef]
- Koekemoer, L.; Kamau, L.; Hunt, R.; Coetzee, M. A cocktail polymerase chain reaction assay to identify members of the Anopheles funestus (Diptera: Culicidae) group. Am. J. Trop. Med. Hyg. 2002, 66, 804–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bass, C.; Nikou, D.; Blagborough, M.; Vontas, J.; Sinden, E.; Williamson, S.; Field, M. PCR-based detection of Plasmodium in Anopheles mosquitoes: A comparison of a new high-throughput assay with existing methods. Malar. J. 2008, 7, 177. [Google Scholar] [CrossRef] [Green Version]
- Padley, D.; Moody, A.H.; Chiodini, P.L.; Saldanha, J. Use of a rapid, single-round, multiplex PCR to detect malarial parasites and identify the species present. Ann. Trop. Med. Parasitol. 2003, 97, 131–137. [Google Scholar] [CrossRef]
- Snounou, G.; Pinheiro, L.; Goncalves, A.; Fonseca, L.; Dias, F.; Brown, K.N.; Dorosario, V.E. The importance of sensitive detection of malaria parasites in the human and insect hosts in epidemiological studies, as shown by the analysis of field samples from Guinea Bissau. Trans. R. Soc. Trop. Med. Hyg. 1993, 87, 646–653. [Google Scholar] [CrossRef]
- Tchouakui, M.; Chiang, M.-C.; Ndo, C.; Kuicheu, C.; Amvongo-Adjia, N.; Wondji, M.; Tchoupo, M.; Kusimo, M.; Riveron, J.; Wondji, C. A marker of glutathione S-transferase-mediated resistance to insecticides is associated with higher Plasmodium infection in the African malaria vector Anopheles funestus. Sci. Rep. 2019, 9, 5772. [Google Scholar] [CrossRef] [Green Version]
- Riveron, J.; Chiumia, M.; Menze, B.; Barnes, K.; Irving, H.; Ibrahim, S.; Weedall, G.; Mzilahowa, T.; Wondji, C. Rise of multiple insecticide resistance in Anopheles funestus in Malawi: A major concern for malaria vector control. Malar. J. 2015, 14, 344. [Google Scholar] [CrossRef] [Green Version]
- Trask, J.; Malhi, R.; Kanthaswamy, S.; Johnson, J.; Garnica, W.; Malladi, V.; Smith, D. The effect of SNP discovery method and sample size on estimation of population genetic data for Chinese and Indian rhesus macaques (Macaca mulatta). Primates 2011, 52, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Thompson, J.; Higgins, D.; Gibson, T. CLUSTALW: Improving the sensitivity of progressive miltiple sequence alignment through sequence weighting, position specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clement, M.; Posada, D.; Crandall, K. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, A.; Cabezas, M.; Tavares, A.; Xavier, R.; Branco, M. tcsBU: A tool to extend TCS network layout and visualization. Bioinformatics 2015, 32, 627–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudson, R.R.; Slatkin, M.; Maddison, W.P. Estimation of levels of gene flow from DNA sequence data. Genetics 1992, 132, 583–589. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Nei, M. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef] [Green Version]
- Antonio-Nkondjio, C.; Ndo, C.; Njiokou, F.; Bigoga, J.; Awono-Ambene, H.; Etang, J.; Ekobo, A.S.; Wondji, C. Review of malaria situation in Cameroon: Technical viewpoint on challenges and prospects for disease elimination. Parasit. Vectors 2019, 12, 501. [Google Scholar] [CrossRef] [Green Version]
- Tabue, R.; Nem, T.; Atangana, J.; Bigoga, J.; Patchoké, S.; Tchouine, F.; Fodjo, B.; Leke, R.; Fondjo, E. Anopheles ziemanni a locally important malaria vector in Ndop health district, north west region of Cameroon. Parasit. Vectors 2014, 7, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanji, S.; Mafo, F.; Tendongfor, N.; Tanga, M.; Tchuente, F.; Bilong Bilong, C.; Njiné, T. Spatial distribution, environmental and physicochemical characterization of Anopheles breeding sites in the Mount Cameroon region. J. Vector Borne Dis. 2009, 46, 75–80. [Google Scholar] [PubMed]
- Minakawa, N.; Sonye, G.; Mogi, M.; Githeko, A.; Yan, G. The effects of climatic factors on the distribution and abundance of malaria vectors in Kenya. J. Med. Entomol. 2002, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minakawa, N.; Munga, S.; Atieli, F.; Mushinzimana, E.; Zhou, G.; Githeko, A.; Yan, G. Spatial distribution of anopheline larval habitats in Western Kenyan highlands: Effects of land cover types and topography. Am. J. Trop. Med. Hyg. 2005, 73, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Kweka, E.; Munga, S.; Himeidan, Y.; Githeko, A.; Yan, G. Assessment of mosquito larval productivity among different land use types for targeted malaria vector control in the western Kenya highlands. Parasit. Vectors 2015, 8, 356. [Google Scholar] [CrossRef] [Green Version]
- Prapanthadara, L.; Ketterman, A. Qualitative and quantitative changes in glutathione S-transferases in the mosquito Anopheles gambiae confer DDT-resistance. Biochem. Soc. Trans. 1993, 21, 304S. [Google Scholar] [CrossRef]
- Ranson, H.; Rossiter, L.; Ortelli, F.; Jensen, B.; Wang, X.; Roth, C.; Collins, F.; Hemingway, J. Identification of a novel class of insect glutathione S-transferases involved in resistance to DDT in the malaria vector Anopheles gambiae. Biochem. J. 2001, 359, 295–304. [Google Scholar] [CrossRef]
- Tchakounte, A.; Tchouakui, M.; Mu-Chun, C.; Tchapga, W.; Kopia, E.; Soh, P.; Njiokou, F.; Riveron, J.; Wondji, C. Exposure to the insecticide-treated bednet PermaNet 2.0 reduces the longevity of the wild African malaria vector Anopheles funestus but GSTe2-resistant mosquitoes live longer. PLoS ONE 2019, 14, e0213949. [Google Scholar] [CrossRef] [Green Version]
- Hemingway, J.; Hawkes, N.; Carroll, L.; Ranson, H. The molecular basis of insecticide resistance in mosquitoes. Insect Biochem. Mol. Biol. 2004, 34, 653–665. [Google Scholar] [CrossRef]
- Ffrench-Constant, R. The molecular and population genetics of cyclodiene insecticide resistance. Insect Biochem. Mol. Biol. 1994, 24, 335–345. [Google Scholar] [CrossRef]
- Andreasen, M.; Ffrench-Constant, R. In situ hybridization to the Rdl locus on polytene chromosome 3L of Anopheles stephensi. Med. Vet. Entomol. 2002, 16, 452–455. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; Awolola, T.S.; Howell, P.; Koekemoer, L.L.; Brooke, B.D.; Benedict, M.Q.; Coetzee, M.; Zheng, L. Independent mutations in the Rdl locus confer dieldrin resistance to Anopheles gambiae and An. arabiensis. Insect Mol. Biol. 2005, 14, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Etang, J.; Manga, L.; Chandre, F.; Guillet, P.; Fondjo, E.; Mimpfoundi, R.; Toto, J.-C.; Fontenille, D. Insecticide Susceptibility Status of Anopheles gambiae s.l. (Diptera: Culicidae) in the Republic of Cameroon. J. Med. Entomol. 2003, 40, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Nwane, P.; Etang, J.; Chouaїbou, M.; Toto, J.C.; Koffi, A.; Mimpfoundi, R.; Simard, F. Multiple insecticide resistance mechanisms in Anopheles gambiae s.l. populations from Cameroon, Central Africa. Parasit. Vectors 2013, 6, 41. [Google Scholar] [CrossRef] [Green Version]
- Hancock, P.; Hendriks, C.; Tangena, J.-A.; Gibson, H.; Hemingway, J.; Coleman, M.; Gething, P.; Cameron, E.; Bhatt, S.; Moyes, C. Mapping trends in insecticide resistance phenotypes in African malaria vectors. PLoS Biol. 2020, 18, e3000633. [Google Scholar] [CrossRef]
- Matiya, D.; Philbert, A.; Kidima, W.; Matowo, J. Dynamics and monitoring of insecticide resistance in malaria vectors across mainland Tanzania from 1997 to 2017: A systematic review. Malar. J. 2019, 18, 102. [Google Scholar] [CrossRef]
- Chang, X.; Zhong, D.; Lo, E.; Fang, Q.; Bonizzoni, M.; Wang, X.; Lee, M.-C.; Zhou, G.; Zhu, G.; Qin, Q.; et al. Landscape genetic structure and evolutionary genetics of insecticide resistance gene mutations in Anopheles sinensis. Parasit. Vectors 2016, 9, 228. [Google Scholar] [CrossRef] [Green Version]
- Souris, M.; Marcombe, S.; Laforet, J.; Brey, P.T.; Corbel, V.; Overgaard, H.J. Modeling spatial variation in risk of presence and insecticide resistance for malaria vectors in Laos. PLoS ONE 2017, 12, e0177274. [Google Scholar] [CrossRef] [Green Version]
- Mandeng, S.E.; Awono-Ambene, H.P.; Bigoga, J.D.; Ekoko, W.E.; Binyang, J.; Piameu, M.; Mbakop, L.R.; Fesuh, B.N.; Mvondo, N.; Tabue, R.; et al. Spatial and temporal development of deltamethrin resistance in malaria vectors of the Anopheles gambiae complex from North Cameroon. PLoS ONE 2019, 14, e0212024. [Google Scholar] [CrossRef] [Green Version]
- Wanjala, C.L.; Kweka, E.J. Malaria Vectors Insecticides Resistance in Different Agroecosystems in Western Kenya. Front. Public Health 2018, 6, 55. [Google Scholar] [CrossRef]
- Touré, Y.T.; Petrarca, V.; Traoré, S.F.; Coulibaly, A.; Maïga, H.M.; Sankaré, O.; Sow, M.; Di Deco, M.A.; Coluzzi, M. Ecological genetic studies in the chromosomal form Mopti of Anopheles gambiae s.str. in Mali, West Africa. Genetica 1994, 94, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Petrarca, V.; Nugud, A.D.; Ahmed, M.A.E.; Haridi, A.M.; Di Deco, M.A.; Coluzzi, M. Cytogenetics of the Anopheles gambiae complex in Sudan, with special reference to An. arabiensis: Relationships with East and West African populations. Med. Vet. Entomol. 2000, 14, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Ayala, D.; Ullastres, A.; González, J. Adaptation through chromosomal inversions in Anopheles. Front. Genet. 2014, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, K.; Weedall, G.; Ndula, M.; Irving, H.; Mzihalowa, T.; Hemingway, J.; Wondji, C. Genomic footprints of selective sweeps from metabolic mesistance to pyrethroids in African malaria vectors are driven by scale up of insecticide-based vector control. PLoS Genet. 2017, 13, e1006539. [Google Scholar] [CrossRef] [PubMed]
- Simard, F.; Lehmann, T.; Lemasson, J.-J.; Diatta, M.; Fontenille, D. Persistence of Anopheles arabiensis during the severe dry season conditions in Senegal: An indirect approach using microsatellite loci. Insect Mol. Biol. 2000, 9, 467–479. [Google Scholar] [CrossRef]
- Yang, M.; Ma, Y.; Wu, J. Mitochondrial genetic differentiation across populations of the malaria vector Anopheles lesteri from China (Diptera: Culicidae). Malar. J. 2011, 10, 216. [Google Scholar] [CrossRef] [Green Version]
- Riveron, J.; Huijben, S.; Tchapga, W.; Tchouakui, M.; Wondji, M.; Tchoupo, M.; Irving, H.; Cuamba, N.; Maquina, M.; Paaijmans, K.; et al. Escalation of pyrethroid resistance in the malaria vector Anopheles funestus induces a loss of efficacy of piperonyl butoxide-based insecticide-treated nets in Mozambique. J. Infect. Dis. 2019, 220, 467–475. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
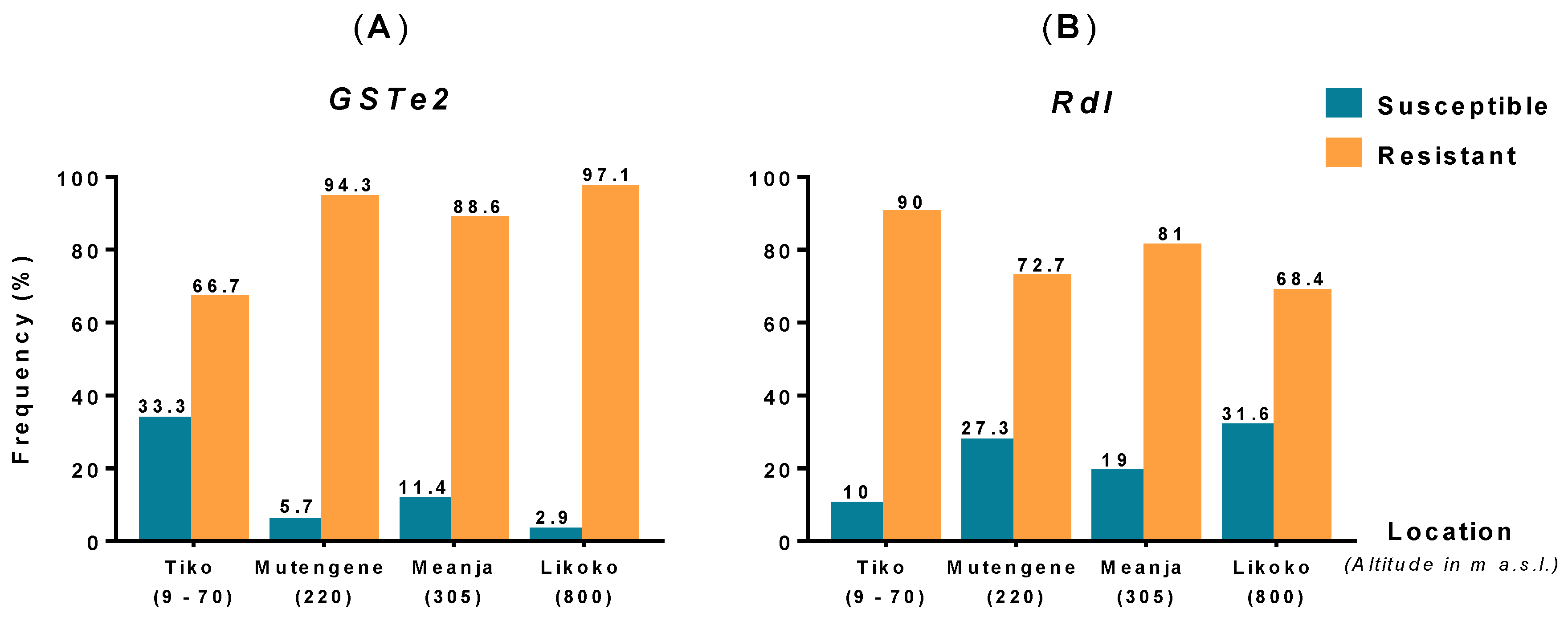{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | N | S * | h (Hd) | Syn | NSyn | π (k) | D | F * |
|---|---|---|---|---|---|---|---|---|
| Per allele | ||||||||
| S | 18 | 11 | 9 (0.91) | 4 | 2 (D61E, L119F) | 0.003 (2.69) | −0.84 ns | −0.70 ns |
| R | 54 | 6 | 5 (0.36) | 2 | 1 (L119F) | 0.001 (0.60) | −1.36 ns | −0.27 ns |
| Per locality (altitude in m a.s.l.) | ||||||||
| Tiko (9–70) | 12 | 8 | 6 (0.68) | 4 | 1 (L119F) | 0.003 (1.97) | −1.38 ns | −1.77 ns |
| Mutengene (220) | 20 | 8 | 7 (0.58) | 4 | 1 (L119F) | 0.002 (1.38) | −1.31 ns | −2.13 ns |
| Meanja (305) | 20 | 7 | 5 (0.56) | 2 | 2 (D61E, L119F) | 0.001 (1.04) | −1.55 ns | −2.54 ns |
| Likoko (800) | 20 | 7 | 5 (0.51) | 1 | 1 (L119F) | 0.002 (1.32) | −1.40 ns | −0.76 ns |
| ALL | 72 | 14 | 12 (0.56) | 6 | 2 | 0.002 (1.36) | −1.62 ns | −1.02 ns |
| Samples | N | S * | h (Hd) | Syn | NSyn | π (k) | D | F * |
|---|---|---|---|---|---|---|---|---|
| Per Allele | ||||||||
| S | 25 | 11 | 10 (0.90) | 0 | 0 | 0.003 (2.53) | −0.44 ns | −0.53 ns |
| R | 43 | 6 | 6 (0.30) | 1 | 0 | 0.0005 (0.54) | −1.61 ns | −1.65 ns |
| Per locality (altitude in m a.s.l.) | ||||||||
| Tiko (9–70) | 12 | 0 | 1 (0.00) | 0 | 0 | 0 (0.00) | n.a. | n.a. |
| Mutengene (220) | 20 | 9 | 9 (0.85) | 1 | 1 (A296S) | 0.002 (1.97) | −0.76 ns | −0.39 ns |
| Meanja (305) | 20 | 10 | 6 (0.68) | 0 | 1 (A296S) | 0.002 (2.00) | −1.02 ns | 0.04 ns |
| Likoko (800) | 16 | 8 | 7 (0.83) | 0 | 1 (A296S) | 0.002 (2.38) | −0.05 ns | −0.21 ns |
| ALL | 68 | 16 | 16 (0.71) | 1 | 1 | 0.002 (1.85) | −1.32 ns | −1.87 ns |
| Tiko | Mutengene | Meanja | |
|---|---|---|---|
| Mutengene | −0.027 ns (0.00) | ||
| Meanja | 0.002 ns (57.05) | 0.003 ns (43.40) | |
| Likoko | −0.007 ns (0.00) | 0.010 ns (12.19) | −0.0006 ns (0.00) |
| Tiko | Mutengene | Meanja | |
|---|---|---|---|
| Mutengene | 0.125 ** (1.24) | ||
| Meanja | 0.037 * (2.08) | 0.0098 ns (15.82) | |
| Likoko | 0.158 ** (1.16) | 0.007 ns (25.82) | 0.035 * (14.54) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amvongo-Adjia, N.; Riveron, J.M.; Njiokou, F.; Wanji, S.; Wondji, C.S. Influence of a Major Mountainous Landscape Barrier (Mount Cameroon) on the Spread of Metabolic (GSTe2) and Target-Site (Rdl) Resistance Alleles in the African Malaria Vector Anopheles funestus. Genes 2020, 11, 1492. https://doi.org/10.3390/genes11121492
Amvongo-Adjia N, Riveron JM, Njiokou F, Wanji S, Wondji CS. Influence of a Major Mountainous Landscape Barrier (Mount Cameroon) on the Spread of Metabolic (GSTe2) and Target-Site (Rdl) Resistance Alleles in the African Malaria Vector Anopheles funestus. Genes. 2020; 11(12):1492. https://doi.org/10.3390/genes11121492
Chicago/Turabian StyleAmvongo-Adjia, Nathalie, Jacob M. Riveron, Flobert Njiokou, Samuel Wanji, and Charles S. Wondji. 2020. "Influence of a Major Mountainous Landscape Barrier (Mount Cameroon) on the Spread of Metabolic (GSTe2) and Target-Site (Rdl) Resistance Alleles in the African Malaria Vector Anopheles funestus" Genes 11, no. 12: 1492. https://doi.org/10.3390/genes11121492